

Лесоведение, 2021, № 5, стр. 472-493
Начальный радиальный прирост разных поколений сосны обыкновенной в средневозрастных лесах Кольского полуострова
П. Н. Катютин a, *, Н. И. Ставрова a, В. В. Горшков a, b, И. Ю. Баккал a, С. А. Михайлов a
a Ботанический институт им. В.Л. Комарова РАН
196376 Санкт-Петербург, ул. Проф. Попова, 2, Россия
b Санкт-Петербургский государственный лесотехнический университет им. С.М. Кирова
194021 Санкт-Петербург, Институтский пер., 5, литер У, Россия
* E-mail: PauRussia@binran.ru
Поступила в редакцию 17.01.2020
После доработки 16.10.2020
Принята к публикации 06.04.2021
Аннотация
Исследование выполнено в западной части Кольского полуострова в районе среднего течения р. Лива (67°30′–68°10′ с.ш., 33°57′–34°21′ в.д.) в 4 типах сообществ: лишайниковых сосновых редколесьях, сосняках лишайниковых, сосняках лишайниково-зеленомошных и сосняках зеленомошных. На 12 постоянных пробных площадях по кернам, спилам или срезам, отобранным у основания ствола, определен начальный радиальный прирост (за первые 5 и 20 лет жизни) у особей сосны обыкновенной, относящихся к разным возрастным поколениям, и проанализировано его изменение в зависимости от времени появления поколения после пожара. Установлен ступенчатый характер снижения начального прироста 10-летних поколений в послепожарном временнóм ряду, имеющий значимые типологические различия. Наиболее важными предикторами различий выступают средние, максимальные и минимальные значения прироста на 1- и 2-ом этапах динамики и соотношение значений среднего прироста на 1- и 2-ом этапах. Показано, что наличие допожарных деревьев и повышенная доля березы в составе формирующихся послепожарных сообществ оказывают значимое влияние на характер динамики начального радиального прироста послепожарных поколений сосны.
Исследованию закономерностей динамики радиального прироста сосны обыкновенной под влиянием различных природных факторов (гелиогеофизических, климатических, почвенно-грунтовых) посвящена обширная литература (Битвинскас, 1974; Schweingruber, 1996; Kalela-Brundin, 1999; Lindholm et al., 2000; Шиятов и др., 2000; Pärn, 2002; Алексеев, Сорока, 2003; Румянцев, 2004; Wilson et al., 2005; Кузьмин и др., 2011; Koprowski et al., 2012; Тюкавина, Лежнева, 2014; Махныкина и др., 2017; Железнова, Тобратов, 2019; Матвеев, Тимощук, 2019). Значительное внимание уделяется и вопросам влияния на радиальный прирост сосны обыкновенной антропогенных воздействий, таких как рубки, мелиорация, промышленное загрязнение среды (Таранков, Матвеев, 1994; Кучеров, Мулдашев, 2003; Pärn, 2006; Fedorkov, 2007; Щекалев, Тарханов, 2007).
В то же время для корректной интерпретации природных и антропогенных воздействий необходим учет внутриценотических факторов, оказывающих влияние на радиальный прирост, прежде всего, конкурентных взаимоотношений особей (Nilsson, Gemmel, 1993; Monserud, Sterba, 1996; Bigler et al., 2003; Das, 2012), которые проявляются на фоне определенных условий местообитания. Выявление различий в динамике роста особей разных возрастных групп и разного ценотического статуса позволяет более объективно подойти к интерпретации их реакции на действие экологических факторов, в том числе современных глобальных климатических перемен.
Сосна обыкновенная является важнейшим лесообразователем светлохвойных лесов на территории России, этот вид господствует на площади более 100 млн га. На Кольском полуострове сосновые леса встречаются в широком спектре местообитаний и занимают 43% покрытой лесом площади, или 2.2 млн га. Изучение сосновых лесов Кольского полуострова, их распространения, особенностей экологии, видового состава и структуры проводилось, начиная с 20–30-х годов прошлого века (Regel, 1923; Цинзерлинг, 1934; Некрасова, 1935; Боброва, Качурин, 1936; Цветков, Чертовской, 1978). Примерно с 60-х годов внимание исследователей стали все чаще стали привлекать проблемы разрушения и восстановления экосистем сосновых лесов Кольского севера после различных внешних воздействий: рубок (Леса …, 1961; Цветков, 1986, 2002), пожаров (Пушкина, 1960; Горшков, 1993, 2001; Ставрова, Горшков, 2004), промышленного загрязнения (Цветков, 1985; Алексеев, Ярмишко, 1990; Влияние …, 1990; Ярмишко, 1997; Ганичева и др., 2004; Лукина и др., 2005), которые остаются актуальными и в последнее десятилетие (Цветков В., Цветков И., 2012; Ставрова и др., 2016; Иванова, Лукина, 2017; Ярмишко, Игнатьева, 2019). Одним из важных аспектов при изучении динамики сосновых лесов региона является анализ роста и развития особей основного лесообразующего вида. Можно отметить целый ряд публикаций этого направления, посвященных лесам Кольского полуострова, в которых рассматривались показатели радиального и линейного роста деревьев сосны обыкновенной, различающихся по возрасту (Ярмишко, 1997; Полоскова и др., 2013), жизненному состоянию (Katjutin et. al., 2020), условиям произрастания (Alekseev, Soroka, 2002; Fedorkov, 2007; Ярмишко, 2012; Ярмишко и др., 2017).
Целью данной работы является анализ радиального прироста разных поколений сосны обыкновенной в начальный период жизни (первые 5 и 20 лет) с целью оценки характера и степени дифференциации особей по скорости роста в разных типах средневозрастных сосновых лесов.
ОБЪЕКТЫ И МЕТОДИКА
Исследование выполнено в западной части Кольского полуострова в районе среднего течения р. Ливы (67°30′–68°10′ с.ш., 33°57′–34°21′ в.д.) в 4 типах сообществ: лишайниковых сосновых редколесьях (Subpinetum cladinosum), сосняках лишайниковых (Pinetum cladinosum), сосняках лишайниково-зеленомошных (Pinetum hylocomioso-cladinosum) и сосняках зеленомошных (Pinetum empetroso-myrtillosum).
На Кольском полуострове лишайниковые сосновые леса и редколесья распространены на вершинах и в верхних частях склонов холмов, на речных террасах, лишайниково-зеленомошные и зеленомошные сосновые леса – преимущественно в средних и нижних частях склонов и на равнинах. Под сосновыми лесами и редколесьями на песчаных, ледниковых и водно-ледниковых отложениях формируются подзолы иллювиально-железистые ненасыщенные, мелкоподзолистые, иллювиально-малогумусовые, песчаные, среднескелетные (Классификация …, 2004), относящиеся к отделу Al-Fe-гумусовых почв. Они отличаются коротким почвенным профилем (до 60 см) и накоплением оксалаторастворимых соединений Al и Fe в иллювиальном горизонте при низком содержании в нем фульватного гумуса (Никонов, Переверзев, 1989; Переверзев, 2004). Почвы изученных лишайниковых сообществ отличаются малой толщиной органогенного горизонта (менее 2 см) и низким (менее 2%) содержанием гумуса в иллювиальном горизонте. Для зеленомошных лесов характерны почвы более высокой влажности, имеющие мощность органогенного горизонта до 7–10 см и бόльшее количество вмытого гумуса (до 3–8%) по сравнению с лишайниковыми сообществами.
Климатические условия района исследований определяются особенностями его географического положения в центральной части Кольского полуострова. Климат территории переходный от морского к континентальному, с прохладным летом, сравнительно теплой осенью, относительно мягкой зимой и затяжной поздней весной (Яковлев, Козлова, 1971). Среднегодовая температура воздуха по данным метеостанции г. Мончегорска составляет –0.5°С (Научно-прикладной …, 1988). Среднесуточная температура воздуха самых холодных месяцев – января и февраля составляет –12–13°С, самого теплого месяца – июля – +13–14°С. Общее годовое количество осадков – 550–600 мм, примерно половина выпадает в период с июня по октябрь. Вегетационный период длится 100–130 дней. Для роста растений наиболее важен период со среднесуточной температурой не менее 10°С, который продолжается в районе исследований с июня по август.
Согласно имеющимся данным (NASA …, 2020) по близкой к району исследований метеостанции Sodankyla (67°25' с.ш., 26°35' в.д.) в рассматриваемый временной интервал (1926–2015 гг.) средняя температура июня, июля и августа направленно не изменялась (F = 0.33–2.14; p = 0.15–0.57) и составляла соответственно 11 ± 2°С (6.9–15.8°C), 14 ± 2°С (10.7–17.9°С) и 12 ± 1°С (8.9–15.0°С). Среднее суммарное количество осадков в июне и июле направленно не изменялось (F = 0.52–0.73; p = 0.12–0.41) и составило соответственно 58 ± 28 (6–121) и 70 ± 32 мм (8–139 мм). Среднее количество осадков в августе линейно снизилось (F = = 4.60; p = 0.03; r = –0.22) с 81 ± 32 в период с 1926 по 1946 г. до 56 ± 27 мм в период с 1995 по 2015 г. Практическое отсутствие однонаправленных климатических трендов на территории района исследований позволяет сопоставлять параметры радиального прироста разных поколений сосны, заселившихся в период с 1926 по 1995 г.
Изученные сообщества сформировались после обширных пожаров, которые наблюдались в 1926 и 1937 г. Давность пожара устанавливали по кернам и спилам, которые отбирались у живых деревьев, имеющих пожарные повреждения.
В исследованных сообществах древесный ярус сформирован Pinus sylvestris L. с участием Betula pubescens Ehrh. (табл. 1). В травяно-кустарничковом ярусе преобладают Vaccinium vitis-idea L., Vaccinium myrtillus L., Empetrum hermaphroditum Hagerup. В формировании мохово-лишайникового покрова участвуют зеленые мхи (в основном Pleurozium schreberi (Brid) Mitt. и Dicranum polysetum Sw.) и лишайники (Cladonia stellaris (Opiz.) Brodo, C. rangiferina (L.) Nyl., C. mitis (Sandst.) Hustich).
Таблица 1.
Таксационные характеристики древостоев средневозрастных сосновых лесов
| Постоянная пробная площадь | Вид | Возраст, лет | D1.3м, см | Н, м | N, экз. га–1 | S, м2 га–1 |
|---|---|---|---|---|---|---|
| Сосновые лишайниковые редколесья | ||||||
| 1 (56) | С* | 220 | 38.2 | 12.6 | 11 | 1.2 |
| С | 71 | 9.6 | 8.0 | 1067 | 7.8 | |
| Б | – | 8.8 | 6.0 | 8 | 0.5 | |
| 2 (92) | С* | 188 | 34.2 | 12.0 | 13 | 1.2 |
| С | 72 | 11.8 | 9.2 | 453 | 5.0 | |
| Б | – | 5.3 | 4.1 | 40 | 0.1 | |
| 3 (96) | С | 73 | 20.0 | 9.3 | 293 | 9.3 |
| Б | – | 6.6 | 5.3 | 270 | 1.0 | |
| Сосняки лишайниковые | ||||||
| 4 (81) | С* | 315 | 48.1 | 14.0 | 10 | 1.8 |
| С | 72 | 13.0 | 10.1 | 1080 | 14.3 | |
| Б | – | 10.1 | 8.8 | 30 | 0.2 | |
| 5 (82) | С* | 313 | 35.6 | 17.7 | 40 | 4.0 |
| С | 76 | 13.5 | 12.6 | 541 | 9.5 | |
| Б | – | 5.5 | 6.1 | 27 | 0.1 | |
| 6 (97) | С | 76 | 10.8 | 9.8 | 1670 | 15.4 |
| Б | – | 6.5 | 6.9 | 6 | 0.2 | |
| Сосняки лишайниково-зеленомошные | ||||||
| 7 (75) | С | 70 | 11.9 | 11.5 | 1156 | 12.9 |
| Б | – | 7.9 | 7.7 | 478 | 2.3 | |
| 8 (90) | С* | 103 | 30.0 | 16.7 | 73 | 5.3 |
| С | 81 | 17.6 | 15.0 | 420 | 10.2 | |
| Б | – | 4.5 | 3.6 | 32 | 0.1 | |
| 9 (91) | С | 69 | 15.9 | 12.0 | 655 | 13.1 |
| Б | – | 7.7 | 8.2 | 273 | 1.3 | |
| Сосняки зеленомошные | ||||||
| 10 (93) | С | 69 | 10.9 | 10.5 | 3110 | 29.0 |
| Б | – | 5.3 | 6.1 | 120 | 0.3 | |
| 11 (94) | С | 70 | 14.5 | 14.6 | 1500 | 24.6 |
| Б | – | 6.1 | 8.4 | 140 | 0.4 | |
| 12 (99) | С | 75 | 15.0 | 14.2 | 1420 | 25.1 |
| Б | – | 7.3 | 7.9 | 320 | 1.3 | |
Исследование проводилось на постоянных пробных площадях размером 0.10–0.15 га. Возраст и величина радиального прироста особей, имеющих диаметр ствола на высоте груди более 4 см, устанавливались по кернам, отобранным у основания ствола. Отбор кернов в большинстве случаев проводили на высоте 10–15 см при наклонном положении бурава с фиксацией угла наклона для последующего пересчета и определения истинной величины радиального прироста. При наличии понижений микрорельефа вблизи ствола керны отбирались стандартным способом. Радиальный прирост особей меньшего размера определялся по спилам модельных экземпляров, которые отбирались за пределами пробной площади. На пробных площадях в составе ценопопуляций сосны анализировался прирост особей каждого из 10-летних поколений, которые появились с момента пожара (1926 или 1937 г.).
Измерение величины радиального прироста проводилось с использованием микроскопа МБС–10 и прибора LINTAB-6 с точностью 0.01 мм. На основании измеренных величин рассчитывался средний годичный радиальный прирост в течение первых 5 лет жизни особей – Z5:
где $z_{i}^{1},$ $z_{i}^{2},$ …, $z_{i}^{5}$ – измеренные годичные приросты i-го дерева за 1-, 2-, …, 5-й годы роста (от сердцевины), мм; N – число анализируемых особей.
Аналогичным образом рассчитывался средний годичный радиальный прирост за первые 20 лет жизни особей (Z20).
Статистическая обработка данных выполнена с использованием корреляционного, дисперсионного и дискриминантного анализа. Сравнение средних значений выборок ввиду их ограниченного объема проводилось на основе непараметрического критерия Краскелла–Уоллиса, с учетом результатов многоуровневого критерия Фишера (LSD) при сравнении между собой более двух выборок.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Численность особей в составе ценопопуляций сосны в исследованных средневозрастных сосновых лесах и редколесьях составляет от ~2.5 до 20 тыс. экз. га–1. Древостои на 80–100% по числу и запасу сформированы особями, появившимися на протяжении первых 3-х послепожарных десятилетий (табл. 2). Густота изученных древостоев варьирует от 300–500 до 1500–3000 экз. га–1. В большинстве сообществ, имеющих густоту древостоя от ~300 до 1000 экз. га–1, резко выраженный максимум численности приходится на поколения, появившиеся в последние два десятилетия (8–9-е), доля которых составляет от 76 до 92% (табл. 2). В сообществах, имеющих густоту древостоя 1500–1800 экз. га–1, наблюдается содоминирование по числу особей 1–3 и 8–9-го поколений. При наиболее высокой густоте древостоя (более 3000 экз. га–1) ценопопуляция сосны состоит практически полностью из особей, заселившихся в течение 1–3-го десятилетий после пожара.
Таблица 2.
Характеристики 10-летних поколений в составе ценопопуляций и древостоев сосны обыкновенной в средневозрастных сосновых лесах
| Пробная площадь |
Параметры поколений |
10-летние поколения | В целом | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |||
| Сосновые лишайниковые редколесья | |||||||||||
| 92 | Npop. | $\frac{{47}}{{0.9}}$ | $\frac{{226}}{{4.3}}$ | $\frac{{213}}{4}$ | $\frac{{20}}{{0.4}}$ | $\frac{{20}}{{0.4}}$ | $\frac{{100}}{{1.9}}$ | $\frac{{220}}{{4.1}}$ | $\frac{{1960}}{{36.9}}$ | $\frac{{2507}}{{47.1}}$ | $\frac{{5313}}{{100}}$ |
| Nstand | $\frac{{47}}{{10.3}}$ | $\frac{{226}}{{50}}$ | $\frac{{180}}{{39.7}}$ | – | – | – | – | – | – | $\frac{{453}}{{100}}$ | |
| М | $\frac{{7.7}}{{30.7}}$ | $\frac{{13.4}}{{53}}$ | $\frac{{4.1}}{{16.3}}$ | – | – | – | – | – | – | $\frac{{25.2}}{{100}}$ | |
| 96 | Npop. | $\frac{{53}}{{0.5}}$ | $\frac{{133}}{{1.4}}$ | $\frac{{47}}{{0.5}}$ | $\frac{{40}}{{0.4}}$ | $\frac{{67}}{{0.7}}$ | $\frac{{47}}{{0.5}}$ | $\frac{{313}}{{3.2}}$ | $\frac{{3807}}{{38.5}}$ | $\frac{{5366}}{{54.3}}$ | $\frac{{9873}}{{100}}$ |
| Nstand | $\frac{{53}}{{18.2}}$ | $\frac{{133}}{{45.5}}$ | $\frac{{47}}{{15.9}}$ | $\frac{{20}}{{6.8}}$ | $\frac{{40}}{{13.6}}$ | – | – | – | – | $\frac{{293}}{{100}}$ | |
| М | $\frac{{15.6}}{{32.2}}$ | $\frac{{25.8}}{{53.2}}$ | $\frac{{6.2}}{{12.8}}$ | $\frac{{0.5}}{1}$ | $\frac{{0.4}}{{0.8}}$ | – | – | – | – | $\frac{{48.5}}{{100}}$ | |
| Сосняки лишайниковые | |||||||||||
| 81 | Npop. | $\frac{{120}}{{0.5}}$ | $\frac{{400}}{{1.8}}$ | $\frac{{580}}{{2.6}}$ | $\frac{{70}}{{0.3}}$ | $\frac{{70}}{{0.3}}$ | $\frac{{300}}{{1.3}}$ | $\frac{{3810}}{{}}$ | $\frac{{3330}}{{14.7}}$ | $\frac{{14000}}{{61.7}}$ | $\frac{{22{\kern 1pt} 680}}{{100}}$ |
| Nstand | $\frac{{120}}{{11.1}}$ | $\frac{{400}}{{37}}$ | $\frac{{530}}{{49.1}}$ | $\frac{{30}}{{2.8}}$ | – | – | – | – | – | $\frac{{1080}}{{100}}$ | |
| М | $\frac{{32.4}}{{41.5}}$ | $\frac{{25.3}}{{32.4}}$ | $\frac{{19.6}}{{25.2}}$ | $\frac{{0.7}}{{0.9}}$ | 1 | – | – | – | – | $\frac{{78.0}}{{100}}$ | |
| 97 | Npop. | $\frac{{340}}{8}$ | $\frac{{660}}{{15.5}}$ | $\frac{{680}}{{15.9}}$ | $\frac{{440}}{{10.3}}$ | $\frac{{70}}{{1.6}}$ | $\frac{{30}}{{0.7}}$ | $\frac{{390}}{{9.1}}$ | $\frac{{910}}{{21.3}}$ | $\frac{{750}}{{17.6}}$ | $\frac{{4270}}{{100}}$ |
| Nstand | $\frac{{340}}{{20.3}}$ | $\frac{{657}}{{39.2}}$ | $\frac{{645}}{{38.5}}$ | $\frac{{33}}{2}$ | – | – | – | – | – | $\frac{{1675}}{{100}}$ | |
| М | $\frac{{43.5}}{{54.1}}$ | $\frac{{26.0}}{{32.3}}$ | $\frac{{10.8}}{{13.4}}$ | $\frac{{0.2}}{{0.2}}$ | – | – | – | – | – | $\frac{{80.5}}{{100}}$ | |
| Сосняки лишайниково-зеленомошные | |||||||||||
| 75 | Npop. | $\frac{{111}}{{4.6}}$ | $\frac{{700}}{{29}}$ | $\frac{{511}}{{21.2}}$ | $\frac{{233}}{{9.7}}$ | $\frac{{89}}{{3.7}}$ | $\frac{{100}}{{4.1}}$ | $\frac{{302}}{{12.5}}$ | $\frac{{368}}{{15.2}}$ | – | $\frac{{2414}}{{100}}$ |
| Nstand | $\frac{{111}}{{9.5}}$ | $\frac{{694}}{{60}}$ | $\frac{{331}}{{28.6}}$ | $\frac{{21}}{{1.9}}$ | – | – | – | – | – | $\frac{{1156}}{{100}}$ | |
| М | $\frac{{41.9}}{{54.7}}$ | $\frac{{30.1}}{{39.4}}$ | $\frac{{4.4}}{{5.8}}$ | $\frac{{0.1}}{{0.1}}$ | – | – | – | – | – | $\frac{{76.5}}{{100}}$ | |
| 91 | Npop. | $\frac{{36}}{{0.7}}$ | $\frac{{346}}{7}$ | $\frac{{285}}{6}$ | $\frac{{18}}{{0.4}}$ | $\frac{{18}}{{0.4}}$ | $\frac{{24}}{{0.5}}$ | $\frac{{103}}{{2.1}}$ | $\frac{{42}}{{0.9}}$ | $\frac{{4000}}{{82}}$ | $\frac{{4873}}{{100}}$ |
| Nstand | $\frac{{36}}{{5.6}}$ | $\frac{{346}}{{52.8}}$ | $\frac{{261}}{{39.8}}$ | $\frac{{12}}{{1.9}}$ | – | – | – | – | – | $\frac{{655}}{{100}}$ | |
| М | $\frac{{8.7}}{{11.1}}$ | $\frac{{56.0}}{{71.1}}$ | $\frac{{13.9}}{{17.6}}$ | $\frac{{0.2}}{{0.2}}$ | – | – | – | – | – | $\frac{{78.8}}{{100}}$ | |
| Сосняки зеленомошные | |||||||||||
| 93 | Npop. | $\frac{{2530}}{{80.6}}$ | $\frac{{390}}{{12.4}}$ | $\frac{{190}}{{6.1}}$ | $\frac{{10}}{{0.3}}$ | $\frac{{20}}{{0.6}}$ | – | – | – | – | $\frac{{3140}}{{100}}$ |
| Nstand | $\frac{{2530}}{{81.4}}$ | $\frac{{390}}{{12.5}}$ | $\frac{{190}}{{6.1}}$ | – | – | – | – | – | – | $\frac{{3110}}{{100}}$ | |
| М | $\frac{{152.3}}{{94.8}}$ | $\frac{{7.1}}{{4.4}}$ | $\frac{{1.3}}{{0.8}}$ | – | – | – | – | – | – | $\frac{{160.7}}{{100}}$ | |
| 94 | Npop. | $\frac{{1180}}{{38.4}}$ | $\frac{{240}}{{7.8}}$ | $\frac{{80}}{{2.6}}$ | – | – | $\frac{{10}}{{0.3}}$ | $\frac{{60}}{2}$ | $\frac{{1500}}{{48.9}}$ | – | $\frac{{3070}}{{100}}$ |
| Nstand | $\frac{{1180}}{{78.7}}$ | $\frac{{240}}{{16}}$ | $\frac{{80}}{{5.3}}$ | – | – | – | – | – | – | $\frac{{1500}}{{100}}$ | |
| М | $\frac{{160.0}}{{92.8}}$ | $\frac{{8.9}}{{5.2}}$ | $\frac{{3.1}}{2}$ | – | – | – | – | – | – | $\frac{{172.0}}{{100}}$ | |
Примечание: Npop. – численность (экз. га–1) разных поколений в составе ценопопуляции; Nstand – численность (экз. га–1) разных поколений в составе древостоя; М – величина запаса (м3 га–1) разных поколений в составе древостоя. В числителе: абсолютное значение показателя, в знаменателе: относительная доля (%). “–” – поколение отсутствует.
Анализ динамики радиального прироста сосны обыкновенной за первые 20 лет жизни особей (рис. 1) свидетельствует, что она имеет существенные различия в зависимости от типологической принадлежности сообществ и возраста поколений и, в некоторых случаях, индивидуальных особенностей отдельных особей в составе поколений. В качестве примера на рис. 1 представлена динамика роста особей сосны обыкновенной, относящихся к отдельным возрастным поколениям в двух исследованных сообществах – сосняке лишайниковом и сосняке зеленомошном.
Рис. 1.
Динамика радиального прироста особей сосны обыкновенной за первые 20 лет жизни в составе разных послепожарных поколений в сосняке лишайниковом (а) и сосняке зеленомошном (б) на территории Кольского полуострова. 1–5 особи первого 10-летнего поколения, 6–8 – особи третьего 10-летнего поколения; 9–11 особи шестого 10-летнего поколения.
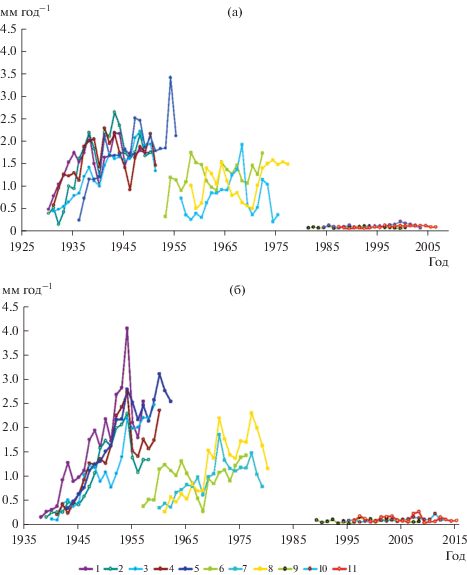
В лишайниковом сообществе (рис. 1а) радиальный прирост за первые 20 лет у особей 1-го послепожарного поколения варьирует в разные годы от 0.1 до 2.5–2.7 мм год–1, в отдельных случаях он может достигать 3.5 мм год–1. Для всех особей поколения в течение исследованного периода характерно линейное увеличение годичного прироста (r = 0.52–0.73, p < 0.001–0.05). У особей сосны 3-го послепожарного поколения начальный прирост варьирует от 0.2 до 2.0 мм год–1. Направленного изменения прироста в течение первых 20 лет жизни при этом не наблюдается. Особи 6-го поколения отличаются низкими значениями и крайне слабым варьированием, т.е. фактической стабильностью начального радиального прироста.
В сосняке зеленомошном (рис. 1б) радиальный прирост за первые 20 лет у особей 1-го послепожарного поколения варьирует от 0.1 до 2.5–3.0 мм год–1, в единичных случаях достигая 4.0 мм год–1. У всех особей величина годичного прироста в течение исследованного периода линейно возрастает (r = = 0.78–0.95, p < 0.001). Начальный прирост особей сосны, относящихся к 3-му послепожарному поколению, изменяется в отдельные годы от 0.2–0.3 до 2.2–2.4 мм год–1. У большинства особей он линейно возрастает (r = 0.65–0.77, p < 0.01), однако часть особей характеризуется отсутствием направленного изменения прироста при его существенных (до 5–7 крат) колебаниях. У особей 6-го послепожарного поколения годичный прирост составляет от 0.02 до 0.3 мм год–1 и в течение первого 20-летия жизни направленно не изменяется.
Изменение среднегодового радиального прироста поколений сосны обыкновенной за первые 5 и 20 лет жизни в зависимости от времени их заселения после пожара в большинстве случаев достаточно точно может быть аппроксимировано линейным (r = –0.76–0.98) или экспоненциальным (r = –0.84–0.99) уравнениями. Величина прироста за первые 5 лет снижается от 0.55–1.25 мм год–1 у поколений, появившихся в первое десятилетие после пожара, до 0.09–0.16 мм год–1 у особей, заселившихся через 50 и более лет после пожара; прирост за первые 20 лет снижается соответственно от 1.0–2.0 мм год–1 до 0.1–0.3 мм год–1 (рис. 2).
Рис. 2.
Изменение начального радиального прироста (первые 5 (а, в, д, ж) и 20 (б, г, е, з) лет) генераций сосны обыкновенной в разных типах северотаежных сосновых лесов. а, б – сосновые лишайниковые редколесья; в, г – сосняки лишайниковые; д, е – сосняки лишайниково-зеленомошные; ж, з – сосняки зеленомошные.
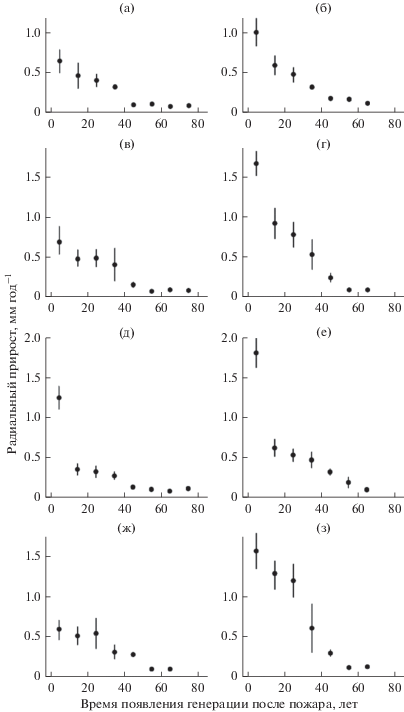
Наиболее высоким начальным приростом отличаются особи сосны, появившиеся в течение 1-го, реже первых 2–3-х десятилетних поколений (табл. 3, 4). Сравнение начального радиального прироста разных поколений сосны в исследованных сосновых лесах показало, что средний прирост особей 1-го поколения за первые 5 и 20 лет является примерно на 30% более высоким в лишайниково-зеленомошных и зеленомошных лесах по сравнению с лишайниковыми лесами и редколесьями (табл. 5). Средний начальный радиальный прирост особей сосны 2- и 3-го десятилетних послепожарных поколений соответственно на 30–40 и 45–50% выше в зеленомошных сосняках по сравнению с остальными типами сообществ. Начальный прирост всех последующих поколений сосны не имеет достоверных различий в разных типах сосновых лесов. Стабилизация начального прироста на нижнем пределе наблюдается через ~50 лет после пожара, его средняя величина за первые 5 и 20 лет составляет соответственно 0.10 и 0.14 мм год–1 (табл. 5). Взаимная связь прироста разных поколений за первые 5 лет проявляется в единичных случаях и является слабой. Более четко выражена связь прироста разных поколений за первые 20 лет. Прирост 2-, 3- и 4-го поколений достоверно положительно скоррелирован с приростом 1-го поколения (r = 0.61–0.64; p < 0.05). Более поздние поколения не имеют связи с приростом 1-го поколения. Довольно тесная положительная связь (r = 0.80–0.89; p < 0.01) выявлена между приростом за первые 20 лет у 2- и 3-, 4- и 5-, 5- и 6-го поколений.
Таблица 3.
Радиальный прирост особей сосны обыкновенной за первые 5 и 20 лет жизни в лишайниковых сосновых лесах и редколесьях
| Пробная площадь |
Показатели прироста поколений |
10-летние поколения | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | ||
| Сосновые лишайниковые редколесья | |||||||||
| 56 | C5 | 0.35 ± 0.08a* | 0.24 ± 0.13b | 0.26 ± 0.09b | 0.20 ± 0.10b | 0.13 ± 0.03c | 0.16 ± 0.08c | 0.11 ± 0.03c | 0.10 ± 0.04c |
| N | 7 | 14 | 36 | 13 | 4 | 3 | 14 | 15 | |
| C5int | 0.35 ± 0.08 | 0.24 ± 0.10 (K–W = 3.84; p = 0.06) | 0.11 ± 0.04 (K–W = 2.86; p = 0.24) | ||||||
| 56 | C20 | 0.58 ± 0.2a | 0.36 ± 0.21b | 0.36 ± 0.17b | 0.30 ± 0.17b | 0.15 ± 0.06c | 0.18 ± 0.08 c | 0.12 ± 0.03d | – |
| N | 6 | 14 | 36 | 13 | 4 | 3 | 14 | ||
| C20int | 0.58 ± 0.2 | 0.34 ± 0.18 (K–W = 2.47; p = 0.29) | 0.17 ± 0.03 (K–W = 4.06; p = 0.11) | 0.12 ± 0.03d | |||||
| 92 | C5 | 0.64 ± 0.26a | 0.42 ± 0.17b | 0.37 ± 0.15b | – | 0.11 ± 0.03c | 0.12 ± 0.05 c | 0.09 ± 0.04c | 0.11 ± 0.03c |
| N | 6 | 12 | 12 | 4 | 7 | 17 | 9 | ||
| C5int | 0.64 ± 0.26 | 0.39 ± 0.16 (K–W = 0.27; p = 0.60) | 0.10 ± 0.04 (K–W = 5.78; p = 0.22) | ||||||
| 92 | C20 | 0.93 ± 0.38a | 0.55 ± 0.17b | 0.48 ± 0.21b | – | 0.17 ± 0.01c | 0.18 ± 0.05 c | 0.13 ± 0.03d | – |
| N | 6 | 12 | 12 | 4 | 7 | 15 | |||
| C20int | 0.93 ± 0.38 | 0.51 ± 0.19 (K–W = 1.61; p = 0.2) | 0.18 ± 0.04 (K–W = 0.14; p = 0.71) | 0.13 ± 0.03d | |||||
| 96 | C5 | 0.83 ± 0.50a | 0.78 ± 0.32a | 0.28 ± 0.20b | – | 0.35 ± 0.13b | 0.11 ± 0.04 c | 0.14 ± 0.08c | 0.11 ± 0.03c |
| N | 8 | 14 | 4 | 6 | 6 | 10 | 33 | ||
| C5int | 0.79 ± 0.38 (K–W = 0.02; p = 0.89) | 0.32 ± 0.16 (K–W = 0.18; p = 0.67) | 0.12 ± 0.05 (K–W = 3.44; p = 0.33) | ||||||
| 96 | C20 | 1.32 ± 0.43a | 1.36 ± 0.40 a | 0.65 ± 0.24b | – | 0.46 ± 0.14b | 0.21 ± 0.10 c | 0.20 ± 0.09c | – |
| N | 10 | 12 | 4 | 6 | 6 | 7 | |||
| C20int | 1.34 ± 0.40 (K–W = 2.25; p = 0.13) | 0.53 ± 0.20 (K–W = 1.14; p = 0.29) | 0.20 ± 0.09 (K–W = 0.05; p = 0.83) | ||||||
| Сосняки лишайниковые | |||||||||
| 81 | C5 | 0.69 ± 0.35a | 0.48 ± 0.21b | 0.51 ± 0.24b | 0.22 ± 0.19c | 0.16 ± 0.07c | 0.08 ± 0.02d | 0.10 ± 0.04d | 0.09 ± 0.03d |
| N | 5 | 21 | 25 | 3 | 9 | 14 | 52 | 11 | |
| C5int | 0.69 ± 0.35 | 0.50 ± 0.22 (K–W = 0.34; p = 0.56) | 0.18 ± 0.10 (K–W = 0.08; p = 0.78) | 0.09 ± 0.03 (K–W = 2.64; p = 0.27) | |||||
| 81 | C20 | 1.68 ± 0.31a | 0.93 ± 0.39b | 0.80 ± 0.32b | 0.36 ± 0.07c | 0.25 ± 0.12c | 0.10 ± 0.04d | 0.10 ± 0.04d | – |
| N | 5 | 21 | 26 | 3 | 9 | 14 | 48 | ||
| C20int | 1.68 ± 0.31 | 0.86 ± 0.36 (K–W = 0.66; p = 0.42) | 0.28 ± 0.11 (K–W = 1.93; p = 0.16) | 0.10 ± 0.04 (K–W = 0.16; p = 0.69) | |||||
| 82 | C5 | 0.57 ± 0.23a | 0.39 ± 0.17b | 0.14c | 0.14 ± 0.02 c | 0.12 ± 0.05c | 0.09 ± 0.02 c | 0.11 ± 0.04c | 0.11 ± 0.03c |
| N | 32 | 10 | 1 | 6 | 6 | 4 | 14 | 21 | |
| C5int | 0.57 ± 0.23 | 0.39 ± 0.17 | 0.11 ± 0.04 (K–W = 9.23; p = 0.10) | ||||||
| 82 | C20 | 0.96 ± 0.36a | 0.68 ± 0.29b | 0.21c | 0.23 ± 0.06c | 0.19 ± 0.03c | 0.13 ± 0.0d | 0.15 ± 0.04d | – |
| N | 32 | 10 | 1 | 6 | 6 | 4 | 6 | ||
| C20int | 0.96 ± 0.36 | 0.68 ± 0.29 | 0.21 ± 0.05 (K–W =1.78; p = 0.41) | 0.14 ± 0.03 (K–W = 0.41; p = 0.52) | |||||
| 97 | C5 | 0.54 ± 0.24a | 0.41 ± 0.20b | 0.41 ± 0.21b | 0.39 ± 0.24b | 0.07c | 0.07c | 0.09 ± 0.03c | 0.11 ± 0.04c |
| N | 20 | 22 | 33 | 4 | 1 | 1 | 9 | 18 | |
| C5int | 0.54 ± 0.24 | 0.41 ± 0.21 (K–W = 0.22; p = 0.89) | 0.10 ± 0.04 (K–W = 4.21; p = 0.24) | ||||||
| 97 | C20 | 1.00 ± 0.35a | 0.67 ± 0.32b | 0.59 ± 0.25b | 0.32 ± 0.18c | 0.16d | 0.10d | 0.09 ± 0.02d | – |
| N | 20 | 22 | 33 | 4 | 1 | 1 | 7 | ||
| C20int | 1.00 ± 0.35 | 0.62 ± 0.28 (K–W = 0.78; p = 0.38) | 0.32 ± 0.18 | 0.10 ± 0.03 (K–W = 2.74; p = 0.25) | |||||
Примечание: * – буквенными индексами (a, b, c, d) отмечены значения, достоверно различающиеся при уровне значимости p < 0.05 (критерий Краскелла–Уоллиса); C5, C20 – среднее значение (мм год–1) и стандартное отклонение радиального прироста за первые 5 и 20 лет; C5int, C20int – среднее значение (мм год–1) и стандартное отклонение радиального прироста за первые 5 и 20 лет по выделенным временным интервалам; “–" – поколение отсутствует.
Таблица 4.
Радиальный прирост особей сосны обыкновенной за первые 5 и 20 лет жизни в лишайниково-зеленомошных и зеленомошных сосновых лесах
| Пробная площадь |
Показатели прироста поколений |
10-летние поколения | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | ||
| Сосняки лишайниково-зеленомошные | |||||||||
| 75 | C5 | 1.25 ± 0.29a | 0.36 ± 0.15b | 0.33 ± 0.15b | 0.34 ± 0.04b | 0.16 ± 0.07c | 0.08 ± 0.01d | 0.09 ± 0.02d | 0.12 ± 0.04d |
| N | 4 | 47 | 27 | 4 | 9 | 2 | 6 | 2 | |
| C5int | 1.25 ± 0.29 | 0.35 ± 0.15 (K–W = 0.73; p = 0.69) | 0.16 ± 0.07 | 0.09 ± 0.03 (K–W = 3.73; p = 0.15) | |||||
| 75 | C20 | 1.81 ± 0.37a | 0.63 ± 0.22b | 0.54 ± 0.16b | 0.59 ± 0.09b | 0.31 ± 0.09c | 0.12 ± 0.03d | 0.11 ± 0.01d | |
| N | 4 | 47 | 27 | 4 | 9 | 2 | 5 | ||
| C20int | 1.81 ± 0.37 | 0.60 ± 0.20 (K–W = 3.43; p = 0.18) | 0.31 ± 0.09 | 0.11 ± 0.02 (K–W = 0.6; p = 0.44) | |||||
| 90 | C5 | 0.70 ± 0.31a | 0.28 ± 0.15b | 0.25 ± 0.14b | 0.17 ± 0.04b | 0.09 ± 0.02c | 0.08 ± 0.02c | 0.11 ± 0.03c | 0.13 ± 0.03c |
| N | 40 | 11 | 20 | 8 | 2 | 16 | 43 | 15 | |
| C5int | 0.70 ± 0.31 | 0.24 ± 0.13 (K–W = 1.75; p = 0.42) | 0.10 ± 0.03 (K–W = 14.2; p = 0.01) | ||||||
| 90 | C20 | 1.24 ± 0.44a | 0.44 ± 0.20b | 0.36 ± 0.19b | 0.30 ± 0.21b | 0.13 ± 0.01c | 0.10 ± 0.03c | 0.12 ± 0.03c | |
| N | 40 | 11 | 20 | 8 | 2 | 16 | 35 | ||
| C20int | 1.24 ± 0.44 | 0.37 ± 0.20 (K–W =3.72; p = 0.16) | 0.12 ± 0.03 (K–W =3.8; p = 0.15) | ||||||
| 91 | C5 | 1.10 ± 0.47a | 0.49 ± 0.18b | 0.43 ± 0.16b | 0.16c | 0.19 ± 0.03c | 0.12 ± 0.04d | 0.11 ± 0.04d | 0.10 ± 0.02d |
| N | 2 | 10 | 5 | 1 | 3 | 13 | 15 | 3 | |
| C5int | 1.10 ± 0.47 | 0.47 ± 0.17 (KW = 0.54; p = 0.46) | 0.18 ± 0.03 (K–W = 1.8; p = 0.18) | 0.12 ± 0.04 (K–W = 0.33; p = 0.85) | |||||
| 91 | C20 | 2.08 ± 0.37a | 1.22 ± 0.31b | 0.93 ± 0.35b | 0.52c | 0.31 ± 0.02c | 0.21 ± 0.08d | 0.14 ± 0.04e | – |
| N | 2 | 10 | 5 | 1 | 3 | 13 | 15 | ||
| C20int | 2.08 ± 0.37 | 1.14 ± 0.34 (K–W = 2.0; p = 0.16) | 0.36 ± 0.11 (K–W = 1.8; p = 0.18) | 0.21 ± 0.08 | 0.14 ± 0.04 | ||||
| Сосняки зеленомошные | |||||||||
| 93 | C5 | 0.77 ± 0.27a | 0.72 ± 0.20a | 0.45 ± 0.06b | 0.34 ± 0.13b | – | – | – | – |
| N | 64 | 7 | 3 | 4 | |||||
| C5int | 0.76 ± 0.26 (K–W = 0.93; p = 0.33) | 0.39 ± 0.12 (K–W = 2.0; p = 0.16) | |||||||
| 93 | C20 | 1.25 ± 0.30a | 0.88 ± 0.26b | 0.83 ± 0.12b | 0.40 ± 0.09c | – | – | – | – |
| N | 64 | 7 | 3 | 4 | |||||
| C20int | 0.87 ± 0.22 (K–W = 0.01; p = 0.91) | ||||||||
| 94 | C5 | 0.83 ± 0.28a | 0.67a | 0.77 ± 0.18a | 0.27 ± 0.18b | 0.12c | 0.10 ± 0.01c | 0.13 ± 0.02c | – |
| N | 26 | 1 | 4 | 2 | 1 | 2 | 3 | – | |
| C5int | 0.82 ± 0.26 (K–W = 0.48; p = 0.79) | 0.12 ± 0.02 (K–W = 3.52; p = 0.17) | |||||||
| 94 | C20 | 1.73 ± 0.38a | 1.37b | 1.31 ± 0.21b | 0.25 ± 0.07с | 0.12c | 0.11 ± 0.01c | – | – |
| N | 26 | 1 | 4 | 2 | 1 | 2 | |||
| C20int | 1.32 ± 0.19 (K–W = 0.00; p =1.0) | 0.11 ± 0.01 (K–W =1.5; p =0.22) | – | ||||||
| 99 | C5 | 0.59 ± 0.25a | 0.51 ± 0.23a | 0.55 ± 0.33a | 0.22 ± 0.02b | 0.28 ± 0.06b | 0.10 ± 0.01c | 0.10 ± 0.02c | |
| N | 58 | 12 | 5 | 3 | 5 | 5 | 6 | ||
| C5int | 0.58 ± 0.25 (K–W =1.27; p = 0.53) | 0.26 ± 0.05 (K–W = 2.69; p = 0.10) | 0.10 ± 0.02 (K–W = 0.21; p = 0.64) | ||||||
| 99 | C20 | 1.57 ± 0.45a | 1.30 ± 0.36b | 1.26 ± 0.39b | 0.31 ± 0.08c | 0.29 ± 0.09c | 0.12 ± 0.02d | 0.13d | |
| N | 58 | 12 | 5 | 3 | 5 | 5 | 1 | ||
| C20int | 1.32 ± 0.19 (K–W = 0.01; p = 0.92) | 0.30 ± 0.08 (K–W = 0.02; p = 0.88) | 0.12 ± 0.02 (K–W = 0.09; p = 0.77) | ||||||
Примечание. Обозначения см. табл. 3.
Таблица 5.
Начальный прирост (мм год–1) послепожарных поколений сосны обыкновенной в сообществах с разными ценотическими характеристиками
| Поко- ления |
L | Прирост | K–W | p | Spf | Прирост | K–W | p | SBp | Прирост | K–W | p |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| За первые 5 лет | ||||||||||||
| 1 | >94 | 0.60 ± 0.16 | 4.0 | <0.05 | <1–5 | 0.73 ± 0.24 | – | – | <1–3 | 0.73 ± 0.24 | – | – |
| 10–65 | 0.87 ± 0.25 | |||||||||||
| 2 | >55 | 0.43 ± 0.16 | 4.7 | <0.05 | <1 | 0.56 ± 0.16 | 4.1 | <0.05 | <1–3 | 0.49 ± 0.16 | – | – |
| <20 | 0.62 ± 0.09 | 1–5 | 0.36 ± 0.10 | |||||||||
| 3 | >55 | 0.33 ± 0.11 | 5.3 | <0.05 | <1 | 0.46 ± 0.16 | 4.0 | <0.05 | <1–3 | 0.38 ± 0.16 | – | – |
| <20 | 0.59 ± 0.16 | 1–5 | 0.30 ± 0.13 | |||||||||
| 4 | <20–99 | 0.24 ± 0.09 | – | – | <1 | 0.29 ± 0.08 | 5.2 | <0.05 | <1–3 | 0.24 ± 0.09 | – | – |
| 1–5 | 0.18 ± 0.03 | |||||||||||
| 5 | <20–99 | 0.16 ± 0.08 | – | – | <1–5 | 0.16 ± 0.08 | – | – | < 1 | 0.11 ± 0.03 | 6.6 | <0.01 |
| 1–3 | 0.25 ± 0.09 | |||||||||||
| 6–7 | <20–99 | 0.10 ± 0.02 | – | – | <1–5 | 0.10 ± 0.02 | – | – | <1–3 | 0.10 ± 0.02 | – | – |
| За первые 20 лет | ||||||||||||
| 1 | <65 | 1.62 ± 0.33 | 4.3 | <0.05 | <1 | 1.54 ± 0.37 | 4.1 | <0.05 | < 1 | 1.17 ± 0.39 | 4.2 | <0.05 |
| >94 | 1.08 ± 0.38 | 1–5 | 1.08 ± 0.41 | 1–3 | 1.70 ± 0.32 | |||||||
| 2 | <20 | 1.25 ± 0.24 | 4.0 | <0.05 | <1 | 1.09 ± 0.33 | 4.8 | <0.05 | <1–3 | 0.89 ± 0.38 | – | – |
| >55 | 0.77 ± 0.34 | 1–5 | 0.60 ± 0.22 | |||||||||
| 3 | <20 | 1.12 ± 0.23 | 6.3 | <0.05 | <1 | 0.88 ± 0.29 | 6.4 | <0.05 | <1–3 | 0.70 ± 0.34 | – | – |
| >55 | 0.56 ± 0.23 | 1–5 | 0.44 ± 0.22 | |||||||||
| 4 | <20–99 | 0.39 ± 0.15 | – | – | <1–5 | 0.39 ± 0.15 | – | – | < 1 | 0.32 ± 0.10 | – | – |
| 1–3 | 0.56 ± 0.06 | |||||||||||
| 5 | <20–99 | 0.22 ± 0.11 | – | – | <1–5 | 0.22 ± 0.11 | – | – | < 1 | 0.16 ± 0.05 | 7.4 | <0.01 |
| 1–3 | 0.35 ± 0.07 | |||||||||||
| 6–7 | <20–99 | 0.14 ± 0.05 | – | – | <1–5 | 0.14 ± 0.05 | – | – | < 1 | 0.13 ± 0.04 | 5.1 | <0.05 |
| 1–3 | 0.19 ± 0.04 | |||||||||||
Численность особей в составе 1-го десятилетнего поколения сосны является существенно более высокой в зеленомошных лесах (K – W = 6.2; p < 0.05), чем в остальных типах сообществ. Различие средних величин достигает 8 крат (1623 ± 453 и 201 ± 60 экз. га–1). Число особей сосны 2- и 3-го поколений не имеет значимых различий в разных типах сообществ и составляет в среднем, соответственно 313 ± 59 и 332 ± 87 экз. га–1. Согласно результатам множественного дисперсионного анализа густота древостоя (сформированного особями 1–3-го послепожарных 10-летних поколений) и тип леса в совокупности значимо влияют (F = 49.38 и 90.57; p < 0.001) на суммарную численность особей 4–7-го поколений, формирующих основу полога подроста (особи в возрасте более 20 лет). В сосняках зеленомошных при густоте древостоя более 1500 экз. га–1 и доминировании зеленых мхов в составе напочвенного покрова (>80% по покрытию) суммарная численность подроста 4–7-го послепожарных поколений является наиболее низкой (67 ± 19 экз. га–1). Наиболее высокая численность подроста (4206 ± 15 экз. га–1) регистрируется в сосняках лишайниковых, если густота древостоя не превышает 1000 экз. га–1. В лишайниковых редколесьях, сосняках лишайниковых с густотой древостоя более 1500 экз. га–1 и сосняках лишайниково-зеленомошных плотность подроста составляет в среднем 594 ± 122 экз. га–1.
Численность особей в составе отдельных поколений не влияет на величину их начального прироста. В частности, при 20-кратных различиях численности особей 1-го послепожарного поколения (соответственно, 1623 ± 453 и 73 ± 17 экз. га–1) величина их среднего прироста за первые 20 лет может быть одинаковой (соответственно, 1.52 ± 0.25 и 1.56 ± 0.45 мм год–1). В то же время при отсутствии достоверных различий численности особей 2-го послепожарного поколения в разных типах сосновых лесов наблюдаются существенное (примерно 2-кратное) превышение начального прироста этого поколения в сосняках зеленомошных по сравнению с остальными типами сообществ. Густота древостоя, оказывая значимое влияние на плотность подроста, не влияет (K–W = 0.37–3.2; p > 0.05) на величину его начального прироста.
Помимо типологической принадлежности сообществ, существенным фактором, оказывающим влияние на начальный прирост поколений сосны, является наличие на гари остатков допожарного соснового древостоя. Согласно результатам дисперсионного анализа, величина среднего годичного радиального прироста за первые 5 лет у 2-, 3- и 4-го поколений сосны в сообществах с наличием допожарного компонента (1–5 м2 га–1) является на 35–40% более низкой, чем при его отсутствии в составе древостоя (табл. 5). При анализе прироста за первые 20 лет выявлена в целом аналогичная картина: достоверное снижение прироста (на 30–50%) регистрируется у 1-, 2- и 3-го послепожарных поколений сосны.
Положительное влияние на начальный радиальный прирост поколений сосны оказывает участие в составе сообществ березы пушистой (1–2.5 м2 га–1). Оно характерно в основном для лишайниково-зеленомошных и зеленомошных сосновых лесов, но может отмечаться и в лишайниковых редколесьях. Выполненный анализ показал, что этот фактор слабо сказывается на приросте за первые 5 лет, но его влияние четко проявляется у ряда поколений на величине прироста за первые 20 лет (табл. 5). В последнем случае при участии березы в составе древесного яруса регистрируется увеличение прироста 1-го послепожарного поколения (на 45%), а также 4, 5 и 6-го поколений (от 45% до 2 крат).
Изменение начального прироста поколений в послепожарном временнóм ряду имеет неравномерный характер. Достоверно выделяются 3–4 основных уровня значений (этапа динамики) начального радиального прироста, последовательно сменяющихся от первых послепожарных поколений к последним (табл. 6).
Таблица 6.
Параметры начального радиального прироста (мм год–1) сосны обыкновенной на разных этапах послепожарной динамики в разных типах сосновых лесов
| Этап | Тип леса1 | Средний | K–W (p) | Тип леса | Ср. максимум | K–W (p) | Тип леса | Ср. минимум | K–W (p) |
|---|---|---|---|---|---|---|---|---|---|
| За первые 5 лет | |||||||||
| I | 1–4 | 0.74 ± 0.25 | – | 3 | 1.51 ± 0.10 | 4.8 (<0.05) |
3 | 0.64 ± 0.35 | 4.5 (<0.05) |
| 1, 2, 4 | 1.11 ± 0.29 | 1, 2, 4 | 0.20 ± 0.07 | ||||||
| II | 1–4 | 0.34 ± 0.09 | – | 2, 3 | 0.81 ± 0.18 | 7.4 (<0.01) |
1–3 | 0.11 ± 0.05 | 5.4 (<0.05) |
| 1, 4 | 0.48 ± 0.12 | 4 | 0.18 ± 0.03 | ||||||
| III | 1–4 | 0.12 ± 0.03 | – | 1–3 | 0.28 ± 0.08 | 4.1 (<0.05) |
1, 2 | 0.05 ± 0.01 | 5.0 (<0.05) |
| 4 | 0.16 ± 0.06 | 3, 4 | 0.08 ± 0.04 | ||||||
| За первые 20 лет | |||||||||
| I | 3, 4 | 1.61 ± 0.33 | 4.3 (<0.05) |
3, 4 | 2.32 ± 0.28 | 5.9 (<0.05) |
1–4 | 0.79 ± 0.48 | – |
| 1, 2 | 1.08 ± 0.38 | 1, 2 | 1.67 ± 0.51 | ||||||
| II | 4 | 1.16 ± 0.25 | 5.3 (<0.05) |
1–4 | 1.32 ± 0.34 | – | 4 | 0.77 ± 0.21 | 5.8 (<0.05) |
| 1–3 | 0.63 ± 0.25 | 1–3 | 0.27 ± 0.16 | ||||||
| III | 1–4 | 0.26 ± 0.09 | – | 1–4 | 0.38 ± 0.12 | – | 1–4 | 0.15 ± 0.08 | – |
| IV | 1–4 | 0.13 ± 0.03 | – | 1–4 | 0.18 ± 0.07 | – | 1–4 | 0.09 ± 0.02 | – |
Примечание. 1 Тип леса: 1 – сосновые лишайниковые редколесья, 2 – сосняки лишайниковые, 3 – сосняки лишайниково-зеленомошные, 4 – сосняки зеленомошные. K–W – критерий Краскелла–Уоллиса; p – уровень значимости.
Годичный прирост за первые 5 лет на 1-, 2- и 3-ем этапах динамики в среднем одинаков во всех типах сосновых лесов и составляет, соответственно, 0.74 ± 0.25, 0.34 ± 0.09, 0.12 ± 0.03 мм год–1. Однако средние максимумы и минимумы прироста, а также время и степень скачкообразных снижений прироста (переходов с одного этапа на другой) имеют определенные различия в разных типах сосновых лесов (табл. 6). В зеленомошных лесах 1-й этап динамики имеет бóльшую продолжительность – 27 ± 6 лет по сравнению с остальными типами сообществ (в среднем 11 ± 3 года). Продолжительность 2-го этапа в зеленомошных сосняках (15 ± 6 лет) является, напротив, наиболее низкой среди других изученных типов сообществ, в которых она составляет в среднем 24 ± 7 лет. Время перехода на последний этап динамики в разных типах сообществ достоверно не различается и происходит в среднем через 41 ± 9 лет после пожара.
При переходе с 1 на 2-ой этап динамики снижение прироста за первые 5 лет является более резким в лишайниково-зеленомошных и зеленомошных лесах (в среднем на 61 ± 8%), чем в лишайниковых лесах и редколесьях (в среднем на 36 ± 13%). Второе скачкообразное снижение прироста существенно не различается в разных типах сообществ (в среднем на 62 ± 9%).
Наиболее высокие значения максимума и минимума радиального прироста за первые 5 лет (соответственно 1.51 ± 0.10 и 0.64 ± 0.35 мм год–1) на 1-ом этапе динамики характерны для лишайниково-зеленомошных сосновых лесов. В остальных типах сообществ эти величины не различаются и в среднем составляют соответственно примерно 75 и 30% от вышеуказанных значений. В течение 2-го этапа более высокие максимумы начального радиального прироста регистрируются в лишайниковых и лишайниково-зеленомошных лесах (в среднем 0.81 ± 0.18 мм год–1), в лишайниковых редколесьях и сосняках зеленомошных средний максимум на 40% ниже. Средние минимумы прироста в зеленомошных сосняках (0.18 ± ± 0.03 мм год–1) на 60% выше, чем в остальных типах сообществ. На 3-ем этапе средний максимум начального прироста в сосняках зеленомошных (0.16 ± 0.06 мм год–1) составляет не более 60% от соответствующей величины в других типах сообществ; а минимальные значения являются на 40% более низкими в лишайниковых сообществах по сравнению с лишайниково-зеленомошными и зеленомошными.
Анализ изменения радиального прироста поколений сосны за первые 20 лет показал, что в процессе послепожарной сукцессии в абсолютном большинстве случаев в сообществах достоверно выделяется четыре уровня его значений, то есть четыре основных этапа динамики. Временные границы скачкообразного снижения прироста в изученных типах сосновых лесов достоверно не различаются при переходе c 1- на 2-ой (через 11 ± 3 лет) и со 2- на 3-ий (через 33 ± 8 лет) этапы. Переход на последний этап в лишайниковых редколесьях наблюдается на 20 лет позже (через 67 ± 6 лет), чем в других типах сообществ (через 47 ± 7 лет).
На 1-ом этапе величина среднего прироста поколений сосны за первые 20 лет в лишайниковых лесах и редколесьях является примерно на 35% более низкой, чем в лишайниково-зеленомошных и зеленомошных лесах, в которых она составляет в среднем 1.61 ± 0.33 мм год–1 (табл. 6). На 2-ом этапе наиболее высоким средним приростом (1.16 ± 0.25 мм год–1) отличаются зеленомошные леса; в остальных типах сообществ соответствующая величина является на 45% более низкой. На 3- и 4-ом этапах достоверных различий прироста особей сосны за первые 20 лет в разных типах сообществ не наблюдается, его средняя величина составляет соответственно 0.26 ± 0.09 и 0.13 ± 0.03 мм год–1.
Снижение среднего прироста на границе 1- и 2-го этапов является существенно более слабым в зеленомошных лесах (на 24 ± 6%), чем в остальных типах сообществ (на 49 ± 14%). Переход со 2 на 3-й этап связан во всех сообществах с одинаковым и довольно существенным (на 64 ± 11%) снижением прироста. Последнее снижение, отмечающее переход с 3-го на последний, 4-ый этап, наиболее слабо выражено в лишайниковых редколесьях (29 ± 1%) по сравнению с другими типами сообществ (на 58 ± 15%).
Средний максимум прироста за первые 20 лет на 1-ом этапе динамики является более высоким в лишайниково-зеленомошных и зеленомошных лесах (2.32 ± 0.28 мм год–1) по сравнению с лишайниковыми сообществами, в которых его величина примерно на 30% ниже (табл. 6). Средний минимум в разных типах сообществ достоверно не различается и составляет 0.79 ± 0.48 мм год–1. На 2-ом этапе величина среднего максимума прироста за первые 20 лет в разных типах сосновых лесов достоверно не различается 1.32 ± 0.34 мм год–1, тогда как средний минимум имеет существенные различия: в зеленомошных лесах его величина (0.77 ± 0.21 мм год–1) почти в 3 раза выше, чем в остальных типах сообществ. На двух последних этапах величины среднего максимума и минимума в разных типах сосновых лесов не имеют существенных различий, на 3-ем этапе они составляют, соответственно, 0.38 ± 0.12 и 0.15 ± 0.08 мм год–1, на 4-ом этапе – 0.18 ± 0.07 и 0.09 ± 0.02 мм год–1.
Таким образом, на основе выполненного анализа было выявлено различие ряда параметров динамики начального радиального прироста послепожарных поколений сосны в разных группах сообществ. Есть параметры, которые достоверно различаются в группах лишайниковых лесов и редколесий (1) и лишайниково-зеленомошных и зеленомошных лесов (2). Другие параметры имеют отличие в зеленомошных лесах или лишайниковых редколесьях (1) по сравнению с остальными типами сообществ (2). Для того, чтобы учесть все эти данные и на основе наиболее информативных параметров выделить достоверно различающиеся по динамике начального прироста поколений группы типов леса, проведен дискриминантный анализ, в котором использованы данные по приросту за первые 20 лет. По его результатам в исследованной выборке средневозрастных сосновых лесов достоверно выделяются три группы сообществ: первая включает лишайниковые сосновые леса и редколесья, вторая – сосняки лишайниково-зеленомошные, третья – сосняки зеленомошные. Предикторами, наиболее точно отражающими различия этих групп сообществ, являются: средний, максимальный и минимальный начальный прирост особей сосны в течение 1- и 2-го этапов динамики, процентное соотношение среднего прироста на 1- и 2-ом этапах динамики. Уровень значимости уравнений 1- и 2‑ой дискриминантных функций: p < 0.001 и p < < 0.05. В уравнении 1-ой функции наиболее высокими значениями коэффициентов отличаются параметры прироста на 1-ом этапе динамики, в уравнении 2-ой функции – параметры прироста на 2-ом этапе динамики.
Во всех исследованных сообществах, независимо от их таксономической принадлежности, густоты, суммы площадей сечений и запаса древостоев, а также общей численности ценопопуляций сосны выявлен в целом однотипный характер изменения начального радиального прироста в ряду послепожарных поколений. Начальный прирост особей сосны обыкновенной, которые заселились на протяжении первых 3-х послепожарных десятилетий, в 3–10 раз превышает соответствующие величины у особей, появившихся в течение 3-х последних десятилетий. Полученные оценки среднего начального радиального прироста деревьев старших поколений сосны, формирующих основу средневозрастных древостоев, согласуются с соответствующими данными других авторов для того же региона (Alekseev, Soroka, 2002; Yarmishko, 2015).
Исследователями неоднократно подчеркивалась связь скорости роста деревьев сосны обыкновенной с типологической принадлежностью сообществ и густотой древостоев (Бузыкин и др., 2002; Усольцев, 2002; Цветков, 2002; Демаков и др., 2019). Установлено, что среднее значение прироста в молодых древостоях сосны, формирующихся в более сухих и бедных элементами питания местообитаниях (сосняки лишайниковые), на 20–30% ниже, чем на более богатых и влажных почвах в сообществах сосняков черничных. Проведенное нами исследование показало, что типологические различия среднего прироста за первые 5 и 20 лет характерны только для деревьев, относящихся к первым трем 10-летним послепожарным поколениям, причем величина прироста в сосняках лишайниково-зеленомошных и зеленомошных на 30–50% выше, чем в лишайниковых лесах и редколесьях. Начиная с 4-го десятилетия после пожара начальный прирост поколений сосны в разных типах средневозрастных сосновых лесов достоверных различий не имеет.
В отношении типологических различий численности особей в составе отдельных послепожарных поколений можно отметить следующую закономерность: наиболее благоприятные условиях для выживания всходов и сеянцев сосны складываются на свежих гарях зеленомошных сосновых лесов, о чем свидетельствует на порядок более высокая, чем в остальных типах сообществ, густота особей, заселившихся в первое десятилетие после пожара. Скорее всего, это обусловлено неполным выгоранием слоя подстилки, имеющего в зеленомошниках значительную толщину, поскольку наличие даже небольшого слоя органики существенно повышает влагоемкость субстрата на гари. В последующие два послепожарных десятилетия различия микроклиматических условий и влажности верхних горизонтов на гарях разных типов сосновых лесов, по-видимому, относительно выравниваются благодаря формированию защитного покрова из трав, кустарничков и подроста березы. Это выражается в одинаковой в среднем густоте 2- и 3-го поколений сосны в лишайниковых, лишайниково-зеленомошных и зеленомошных сообществах. В дальнейшем, на протяжении 4-го и последующих десятилетий, по мере увеличения полноты и сомкнутости крон формирующегося древостоя, восстановления плотного зеленомошного покрова и толщины лесной подстилки, условия для прорастания семян и выживания всходов в сосняках зеленомошных становятся значительно менее благоприятными, чем в лишайниковых и лишайниково-зеленомошных сообществах. Это приводит к существенно более низкой численности новых поколений сосны, однако на их начальном приросте указанные факторы не сказываются: он остается в среднем одинаковым в разных типах сообществ и при разной густоте древостоев.
При анализе изменения начального прироста от поколения к поколению было установлено, что этот процесс, хотя и может быть удовлетворительно описан линейным уравнением, не является равномерным. Величина прироста изменяется скачкообразно. Первое скачкообразное снижение прироста происходит относительно рано – в абсолютном большинстве случаев через ~10 лет после пожара, второе – через 30–40 лет. Стабилизация начального прироста, обусловленная достижением минимальных значений, обеспечивающих выживание особей сосны в конкретных условиях местообитания (типа леса), наблюдается через 40–50 лет после пожара. Снижение прироста при переходе с одной ступени на другую является довольно значительным – до 50–65%, особенно в середине и второй половине послепожарного периода. Главной его причиной является внутрипопуляционное конкурентное взаимодействие поколений. Восстановительная динамика параметров напочвенного покрова и лесной подстилки не может являться причиной скачкообразного изменения начального прироста послепожарных поколений, поскольку представляет собой последовательный непрерывный процесс (Горшков, 1995; Горшков и др., 2005).
Более низкая скорость роста молодых генераций древесных растений по отношению к старшим – хорошо известное явление, причем в сосновых лесах, особенно в разреженных северотаежных, оно обусловлено в первую очередь корневой конкуренцией (Листов, 1986; Kuuluvainen et al., 1993). Это еще раз подтвердили результаты специальных исследований (Санникова, 2003; Санников и др., 2012). Однако этот фактор не может объяснить первое скачкообразное снижение начального радиального прироста особей сосны, которое в абсолютном большинстве случаев наблюдается уже через ~10 лет после пожара. Густота особей первого послепожарного 10‑летнего поколения, как было показано на примере исследованных сообществ, не влияет на прирост 2-го поколения. Наиболее вероятной причиной того, что начальный прирост у особей, появившихся в течение 2-го послепожарного десятилетия, существенно более низкий, чем у особей, появившихся ранее, является 1.5–2-кратное увеличение содержания подвижных форм элементов минерального питания растений (кальция, калия, магния, фосфора, ряда микроэлементов) в верхних горизонтах почв (Neff et al., 2005; Сосорова и др., 2013; Дымов и др., 2014). Через 10 лет после пожара это положительное для развития подроста явление практически полностью сходит на нет, и начальный прирост вновь появляющихся поколений падает. Исключением является прирост особей сосны за первые 5 лет в зеленомошных сообществах, скачкообразное снижение которого происходит не через 10, а через 20–30 лет после пожара. Это можно объяснить более благоприятными для начальных стадий развития сеянцев условиями на гарях сосняков зеленомошных: в частности, наличием недогоревшего слоя подстилки, существенно повышающего обеспеченность субстрата элементами питания и влагой, что не характерно для лишайниковых и лишайниково-зеленомошных лесов.
Второе и последующие скачкообразные снижения начального прироста сосны, очевидно, связаны с повышением напряженности корневой конкуренции и уменьшением количества доступных для новых генераций сосны ресурсов питания и влаги. Известно, что к 30–40-летнему возрасту у деревьев сосны первых послепожарных поколений происходит переход от экстенсивного роста корневых систем к интенсивному. На фоне многократного снижения скорости нарастания стержневого и главных боковых корней резко возрастает степень насыщенности сосущими корнями корнеобитаемого пространства за счет прогрессирующего увеличения числа корней высоких порядков (Шиманюк, 1955; Калинин, 1978; Санникова 1979, 2003; Санников и др., 2012). Согласно имеющимся данным (Санникова, 1992; Санников и др., 2012), годичный прирост подроста сосны имеет значимую отрицательную связь с индексом корневой конкуренции древостоя (r = –0.54…–0.67; η = = 0.60–0.89; p < 0.01), который в свою очередь положительно связан с насыщенностью ризосферы тонкими корнями деревьев (Санникова, Локосова, 2001). Приведенные факты позволяют объяснить установленное в нашем исследовании второе, наиболее существенное скачкообразное снижение прироста новых поколений сосны, наблюдающееся примерно через 30–40 лет после пожара. Корненасыщенность верхних горизонтов почв является более высокой в сообществах с более высокой густотой и продуктивностью древостоя. Было показано, что в редкостойных сообществах с лишайниковым покровом она примерно в 2 раза ниже, чем в лесах с участием в покрове лишайников и зеленых мхов. Интенсивность микоризации поглощающих корней в обоих случаях примерно одинакова, однако уровень жизненности и, соответственно, функциональной активности эктомикориз существенно выше при высокой корненасыщенности почв в более благоприятных условиях местообитания (Веселкин, 2001).
Анализируя результаты выполненного нами исследования, с учетом приведенных выше сведений, можно предположить следующее. По мере послепожарного формирования разных по густоте и сумме площадей сечений древостоев в разных типах сосновых лесов и связанной с этим разной корненасыщенности верхних почвенных горизонтов, а также компенсирующих эти различия механизмов, с течением времени происходит постепенное стирание различий в параметрах роста последовательно появляющихся поколений сосны. Типологические различия величин, хронологии и интенсивности изменения начального радиального прироста в ряду поколений проявляются почти исключительно в течение первых 30–40 лет после пожара, т.е. в течение выделенных 1- и 2-го этапов динамики. Указанная закономерность в концентрирированном виде отражается в перечне предикторов, дискримирующих типы сосновых лесов по параметрам начального радиального прироста послепожарных поколений сосны. Это позволяет заключить, что при давности пожара более 40 лет конкурентное воздействие сформировавшегося разреженного низкополнотного древостоя на бедных и сухих почвах аналогично воздействию высокополнотного древостоя на более богатых и более увлажненных почвах.
Общеизвестно, что сохранившиеся допожарные деревья сосны существенно подавляют рост молодых генераций. Это явление в условиях северной тайги было исследовано, в частности, А.А. Листовым (1986). Согласно данным настоящего исследования наличие на гари деревьев допожарного происхождения оказывает значимое влияние на динамику начального радиального прироста первых трех послепожарных генераций сосны. Но в наибольшей степени его отрицательное влияние сказывается на приросте 2- и 3-го поколений, величина которого снижается примерно в 2 раза. При этом важно отметить, что этот вывод получен на основе анализа выборки сообществ (лишайниковые редколесья, сосняки лишайниковые, сосняки лишайниково-зеленомошные), в пределах которой средний прирост 2- и 3-его десятилетних поколений не имеет достоверных типологических различий. Зеленомошные сосновые леса, в которых прирост является в среднем достоверно более высоким, чем в остальных сообществах, не входят в эту выборку из-за отсутствия в древостоях изученных зеленомошных лесов допожарного компонента. Более слабая реакция начального прироста 1-го поколения на конкурентное воздействие со стороны допожарных деревьев (на приросте за первые 5 лет она вообще не проявляется) по сравнению со 2- и 3-им поколениями, скорее всего, объясняется улучшением условий минерального питания в первые годы после пожара, о чем говорилось выше. Это явление может оказывать положительное влияние на прирост сеянцев сосны, появляющихся на гари в первые годы после пожара и в определенной степени уменьшать отрицательное влияние конкуренции со стороны допожарных деревьев сосны.
Значимым фактором, положительно влияющим на величину начального прироста поколений сосны, может выступать участие в составе сообществ березы пушистой. Этот вид является характерным компонентом древесного яруса северотаежных лесов и после пожара быстро восстанавливается за счет вегетативного возобновления. В районе исследований участие березы является наиболее значимым в лишайниково-зеленомошных сообществах, где она находит наиболее оптимальные условия. Сосняки лишайниково-зеленомошные более благоприятны для березы по уровню увлажнения, чем сосняки лишайниковые, и более благоприятны по уровню освещенности, чем сосняки зеленомошные. Однако заметное участие березы можно наблюдать и в некоторых зеленомошных сосновых лесах, и даже в лишайниковых редколесьях. В первом случае это, как правило, связано с относительной неравномерностью пространственной структуры соснового древостоя, наличием участков с редким стоянием деревьев и лучшей освещенностью. В лишайниковых редколесьях участие березы, как показывают наши наблюдения, поддерживается при наличии выраженного микрорельефа (небольших понижений диаметром от 1.5 до 3 м), в которых, очевидно, складываются более благоприятные локальные условия увлажнения, о чем свидетельствует примесь зеленых мхов в составе напочвенного покрова. Следует признать, что установленное благоприятное влияние березы на начальную скорость роста 1-го послепожарного поколения может быть косвенным результатом выявленных различий его прироста по типам леса, хотя определенную роль может играть и улучшение характеристик среды на открытой гари за счет затенения и снижения перепадов температур. Что касается увеличения прироста 4, 5 и 6-го поколений, прирост которых одинаков в среднем во всех изученных типах сообществ, есть все основания предполагать, что в этом случае положительный эффект связан с обогащением верхнего горизонта почв питательными веществами в результате поступления быстро разлагающегося лиственного опада. Это особенно значимо для особей, появившихся через 30–60 лет после пожара и испытывающих существенное конкурентное воздействие со стороны предшествующих поколений сосны. В ряде работ можно найти указание на более высокие значения прироста сосны в высоту и по диаметру при наличии небольшой примеси березы в молодых сосновых лесах (Сеннов, 1963; Неволин, 1968; Луганский, Макаренко, 1976). Отдельными авторами отмечалось также улучшение условий почвенного питания и лучшее развитие корневой системы сосны в результате благотворного влияния микрофлоры ризосферы березы (Рахтеенко и др., 1976; Лебедев, 2010).
ЗАКЛЮЧЕНИЕ
1) Влияние типа местообитания достоверно сказывается на величине начального (за первые 5 и 20 лет жизни) радиального прироста только первых 3-х послепожарных 10-летних поколений сосны, формирующих основу древостоя; прирост последующих поколений, находящихся под их конкурентным воздействием, одинаков во всех исследованных типах сообществ. Это означает, что конкурентное воздействие низкополнотного древостоя на бедных и сухих почвах аналогично воздействию высокополнотного древостоя на более богатых и более увлажненных почвах.
2) Снижение начального радиального прироста в ряду послепожарных поколений сосны обыкновенной отличается неравномерным, скачкообразным характером. В большинстве сообществ за послепожарный период продолжительностью 80 лет сменяется 3–4 уровня (ступени) значений начального радиального прироста.
3) Скорость изменения начального прироста, его средние, максимальные и минимальные значения на разных ступенях динамики зависят от типологической принадлежности сообществ. По совокупности признаков, характеризующих изменение начального прироста поколений сосны, в исследованной выборке сообществ достоверно выделяются три типологические группы: 1) лишайниковые сосновые леса и редколесья, 2) сосняки лишайниково-зеленомошные, 3) сосняки зеленомошные.
4) Наличие на гари остатков допожарного соснового древостоя приводит к снижению начального радиального прироста трех первых послепожарных поколений сосны. Повышенная доля участия березы пушистой в составе сообществ положительно влияет на прирост наиболее угнетенных поколений, появившихся через 30 и более лет после пожара.
Список литературы
Алексеев А.С., Сорока А.Р. Анализ долговременных тенденций роста Pinus sylvestris на северо-западе Кольского полуострова // Ботанический журн. 2003. Т. 88. № 6. С. 59–75.
Алексеев В.А., Ярмишко В.Т. Влияние атмосферного загрязнения двуокисью серы с примесью тяжелых металлов на строение и продуктивность северо-таежных древостоев // Лесные экосистемы и атмосферное загрязнение. Л.: Наука, 1990. С. 105–115.
Битвинскас Т.Т. Дендроклиматические исследования. Л.: Гидрометеоиздат, 1974. 172 с.
Боброва Л.И., Качурин М.Х. Очерк растительности Монче-тундры // Труды Совета по изучению производительных сил АН СССР. Сер. Кольская. 1936. Вып. 2. С. 95–121.
Бузыкин А.И., Пшеничникова Л.С., Суховольский В.Г. Густота и продуктивность древесных ценозов. Новосибирск: Наука, 2002. 150 с.
Влияние промышленного атмосферного загрязнения на сосновые леса Кольского полуострова. Л.: Ботанический институт им. В.Л. Комарова РАН, 1990. 195 с.
Веселкин Д.В. Структура эктомикориз сосны обыкновенной в связи с корневой конкуренцией древостоя // Генетические и экологические исследования в лесных экосистемах. Екатеринбург: УрО РАН, 2001. С. 113–126.
Ганичева С.Н., Лукина Н.В., Костина В.А., Никонов В.В. Техногенная дигрессия и восстановительная сукцессия в хвойных лесах Кольского полуострова // Лесоведение. 2004. № 3. С. 57–67.
Горшков В.В. Характеристики восстановления лесных экосистем после пожаров // Доклады АН России. 1993. Т. 333. № 6. С. 111–114.
Горшков В.В. Послепожарное восстановление сосновых лесов Европейского Севера. Автореф. дис. … д-ра. биол. наук. (03.00.05 – Ботаника) CПб.: Ботанический институт им. В.Л. Комарова РАН, 2001. 35 с.
Горшков В.В. Послепожарное восстановление мохово-лишайникового яруса в сосновых лесах Кольского полуострова // Экология. 1995. № 3. С. 179–183.
Горшков В.В., Ставрова Н.И., Баккал И.Ю. Динамика восстановления лесной подстилки в бореальных сосновых лесах после пожаров // Лесоведение. 2005. № 3. С. 37–45.
Демаков Ю.П., Демитрова И.П., Нуреева Т.В., Симатова Т.Ю. Влияние начальной густоты и интенсивности изреживания древостоя в культурах сосны на прирост и плотность древесины // Вестник Поволжского государственного технологического университета. Серия: Лес. Экология. Природопользование. 2019. № 4(42). С. 26–40.
Дымов А.А., Дубровский Ю.А., Габов Д.Н. Пирогенные изменения подзолов иллювиально-железистых (средняя тайга, Республика Коми) // Почвоведение. 2014. № 2. С. 144–154.
Железнова О.С., Тобратов С.А. Влияние климата на радиальный прирост сосны обыкновенной (Pinus sylvestris L.) в различных местообитаниях Мещерской низменности // Известия РАН. Серия географическая. 2019. Т. 5. С. 67–77.
Иванова Е.А., Лукина Н.В. Варьирование массы и фракционного состава древесного опада в сосняках кустарничково-лишайниковых при аэротехногенном загрязнении // Лесоведение. 2017. № 5. С. 47–58.
Классификация и диагностика почв России. Смоленск: Ойкумена, 2004. 342 с.
Калинин М.И. Моделирование лесных насаждений. Львов: Вища школа. 1978. 207 с.
Кузьмин А.В., Полоскова Е.Ю., Гончарова О.А. Определение временных интервалов влияния температурных факторов на формирование радиального прироста в сосновых древостоях Кольского региона // Вестник Мурманского государственного технического университета. 2011. Т. 14. № 3. С. 565–576.
Кучеров С.Е., Мулдашев А.А. Радиальный прирост сосны обыкновенной в районе Карабашского медеплавильного комбината // Лесоведение. 2003. № 2. С. 43–49
Лебедев Е.В. Фотосинтез, минеральное питание и биологическая продуктивность растений березы повислой и сосны обыкновенной при совместном выращивании // Лесной вестник. 2010. № 5. С. 15–19.
Леса Кольского полуострова и их возобновление. М.: Изд-во. АН СССР, 1961. 188 с.
Листов А.А. Боры-беломошники. М.: Агропромиздат, 1986. 181 с.
Луганский Н.А., Макаренко Г.П. Влияние густоты и состава березово-сосновых молодняков на рост деревьев сосны // Леса Урала и хозяйство в них. 1976. Вып. 9. С. 84–89.
Лукина Н.В., Сухарева Т.А., Исаева Л.Г. Техногенные дигрессии и восстановительные сукцессии в северотаежных лесах. М.: Наука, 2005. 244 с.
Матвеев С.М., Тимащук Д.А. Дендроклиматический анализ 200-летнего древостоя сосны обыкновенной в Воронежском биосферном заповеднике // Лесоведение. 2019. № 2. С. 93–104.
Махныкина А.В., Верховец С.В., Кошурникова Н.Н. Воздействие нарушающих факторов различного происхождения на радиальный прирост в сосновых насаждениях Центральной Сибири // Известия РАН. Серия биологическая. 2017. № 4. С. 476–482.
Неволин О.А. Взаимодействие сосны и березы и рубки ухода в сосново-березовых насаждениях Архангельской области // Вопросы лесоустройства и таксации лесов Европейского Севера. 1968. Вып. 1. С. 95–103.
Некрасова Т.П. Очерк растительности Лапландского заповедника // Труды Ленинградского общества естествоиспытателей. 1935. Т. 64. Вып. 2. С. 239–272.
Никонов В.В., Переверзев В.Н. Почвообразование в Кольской субарктике. Л.: Наука, 1989. 168 с.
Переверзев В.Н. Лесные почвы Кольского полуострова. М.: Наука, 2004. 232 с.
Полоскова Е.Ю., Гончарова О.А., Кузьмин А.В. Возрастные особенности и динамика радиального прироста сосны обыкновенной на Кольском полуострове // Вестник Мурманского гос. техн. университета. 2013. Т. 16. № 3. С. 519–525.
Пушкина Н.М. Естественное возобновление растительности на лесных гарях // Труды Лапландского гос. заповедника. 1960. Вып. 4. С. 5–125.
Рахтеенко H.H., Мартинович B.C., Крот Л.А., Кабашникова Г.И. О взаимодействии сосны и березы в чистых и смешанных насаждениях // Эколого-физиологические основы взаимодействия растений в фитоценозах. Минск: Наука и техника, 1976. С. 67–94.
Румянцев Д.Е. Влияние климатических факторов на рост сосны в Южной Карелии // Лесоведение. 2004. № 5. С. 73–75.
Санникова Н.С. Микроэкосистемный анализ структуры и функций лесных биогеоценозов. Экология. 2003. № 2. С. 90–95.
Санников С.Н., Санникова Н.С., Петрова И.В. Очерки по теории лесной популяционной биологии. Екатеринбург: РИО УрО РАН, 2012. 281 с.
Санникова Н.С. К количественной оценке корневой конкуренции одиночного дерева сосны //Экологические исследования в лесных и луговых биогеоценозах равнинного Зауралья. Свердловск: УНЦ АН СССР. 1979. С. 21–26.
Санникова Н.С., Локосова Е.И. Микроэкосистемный анализ структурно-функциональных связей в лесных биогеоценозах // Генетические и экологические исследования в лесных экосистемах. Екатеринбург: УрО РАН. 2001. С. 73–94.
Сеннов С.Н. Формирование молодняков на вырубках в сосновых борах // Сб. научно-исследовательских работ по лесному хозяйству. Л.: ЛенНИИ лесного хозяйства, 1963. Вып. 7. С. 136–169.
Сосорова С.Б., Меркушева М.Г., Убугунов Л.Л. Пирогенное изменение содержания микроэлементов в почвах и растениях сосновых лесов Западного Забайкалья // Сибирский экологический журн. 2013. № 5. С. 661–674.
Ставрова Н.И., Горшков В.В. Влияние параметров лесных сообществ на семенную продуктивность и возобновление Pinus sylvestris L. на разных этапах послепожарных сукцессий на Европейском севере // Растительные ресурсы. 2004. Т. 40(3). С. 1–15.
Ставрова Н.И., Горшков В.В., Катютин П.Н. Формирование структуры ценопопуляций лесообразующих видов в процессе послепожарного восстановления северотаежных лесов // Тр. Карельского НЦ РАН. Серия “Биогеография”. 2016. Т. 3. С. 10–28.
Таранков В.И., Матвеев С.М. Радиальный прирост древостоев сосны обыкновенной в зоне действия промышленного загрязнения // Известия высших учебных заведений. Лесной журн. 1994. № 4. С. 47–51.
Тюкавина О.Н., Лежнева С.В. Черты единства в приросте сосны при разных условиях произрастания в Архангельском лесничестве // Вестник Северного (Арктического) федерального университета. Серия: Естественные науки. 2014. № 4. С. 98–111.
Усольцев В.А. Фитомасса лесов северной Евразии: нормативы и элементы географии. Екатеринбург: Изд-во УрО РАН, 2002. 762 с.
Цветков В.Ф. Состояние лесов и лесовозобновление в районе деятельности комбината “Североникель” в Мурманской области // Проблемы биосферы. 1985. Инф. бюлл. № 11. С. 39–60.
Цветков В.Ф. Типы формирования насаждений на вырубках сосновых лесов Мурманской области // Лесоведение. 1986. № 3. С. 3–18.
Цветков В.Ф. Сосняки Кольской лесорастительной области и ведение хозяйства в них. Архангельск: Архангельский гос. техн. университет, 2002. 380 с.
Цветков, В.Ф., Чертовской В.Г. Классификационные типологические схемы и лесорастительное районирование Мурманской области. Архангельск: Архангельский институт леса и лесохимии, 1978. 35 с.
Цветков В.Ф., Цветков И.В. Промышленное загрязнение окружающей среды и лес. Архангельск: Изд-во Северного (Арктического) федерального университета, 2012. 312 с.
Цинзерлинг Ю.Д. География растительного покрова Северо-Запада европейской части СССР // Труды Геоморфологического института АН СССР. Сер. физико-географическая. 1934. Вып. 4. С. 1–377.
Шиманюк А.П. Естественное возобновление на концентрированных вырубках. М.: Изд-во АН СССР. 1955. 355 с.
Шиятов С.Г., Ваганов Е.А., Кирдянов А.В., Круглов В.Б., Мазепа В.С., Наурзбаев М.М., Хантемиров Р.М. Методы дендрохронологии. Сбор и получение древесно-кольцевой информации. Красноярск: ИЦ Красноярского гос. университета, 2000. 80 с.
Щекалев Р.В., Тарханов С.Н. Радиальный прирост сосны обыкновенной при аэротехногенном загрязнении в бассейне Северной Двины // Лесоведение 2007. № 2. С. 45–50.
Яковлев Б.А., Козлова Л.Г. Климатические карты: осадки, температура воздуха // Атлас Мурманской области. М.: Изд-во Главного управления геодезии и картографии при СМ СССР, 1971. С. 9–11.
Ярмишко В.Т. Сосна обыкновенная и атмосферное загрязнение на Европейском Севера. СПб.: НИИ химии СПбГУ, 1997. 210 с.
Ярмишко В.Т. Ход роста Pinus sylvestris L. на северном пределе распространения в условиях атмосферного загрязнения // Известия Самарского НЦ РАН 2012. Т. 14. № 1(6). С. 1576–1580.
Ярмишко В.Т., Игнатьева О.В. Многолетний импактный мониторинг состояния сосновых лесов в центральной части Кольского полуострова // Известия РАН. Сер. биологическая. 2019. № 6. С. 658–668.
Ярмишко В.Т., Лянгузова И.В., Лянгузов А.Ю. Изменение годичного прироста стволов Pinus sylvestris (Pinaceae) при снижении аэротехногенного загрязнения // Растительные ресурсы. 2017. Т. 53. № 4. С. 527–542.
Alekseev A.S., Soroka A.R. Scots Pine Growth Trends in Northwestern Kola Peninsula as an Indicator of Positive Changes in the Carbon Cycle // Climatic change. 2002. V. 55(1–2). P. 183–196.
Bigler Ch., Gričar J., Bugmanna H., Čufar K. Growth patterns as indicators of impending tree death in silver fir // Forest Ecology & Management. 2003. V. 199, Is. 2–3(11). P. 183–190.
Das A. The effect of size and competition on tree growth rate in old-growth coniferous forests // Canadian Journal of Forest Research. 2012. V. 42(11). P. 1983–1995.
Fedorkov A. Effect of heavy metal pollution of forest soil on radial growth of Scots pine // Forest Pathology. 2007. V. 37. № 2. P. 136–142.
Kalela-Brundin M. Climatic information from tree-rings of Pinus sylvestris L. and reconstruction of summer temperatures back to AD 1500 in Femund-marka, Eastern Norway, using partial least squares regression (PSL) analysis // Holocene. 1999. № 9. P. 59–69.
Katjutin P.N., Stavrova N.I., Gorshkov V.V., Lyanguzov A.Yu., Bakkal I.Ju., Mikhailov S.A. Radial growth of trees differing in their vitality in the middle-aged Scots pine forests in the Kola peninsula // Silva Fennica V. 54. № 3. Article ID 10 263. 10 p.
Koprowski M., Przybylak R., Zielski A., Pospieszynska A. Tree rings of Scots pine (Pinus sylvestris L.) as a source of information about past climate in northern Poland // International J. Biometeorology. 2012. V. 56. P. 1–10.
Kuuluvainen T., Hokkanen T., Pukkala T. Factors related to seedlings growth in a boreal Scots pine stand: a spatial analisys of a vegetation-soil system // Can. J. For. Res. 1993. V. 23. P. 2101–2109.
Lindholm M., Lehtonen H., Koström T., Meriläinen J., Eronen M., Timonen M. Climatic signals extracted from ring-width chronologies of Scots pines from the northern, middle and southern parts of the boreal forest belt in Finland // Silva Fennica. 2000. V. 34/4. P. 317–330.
Monserud R.A., Sterba H. A basal area increment model for individual trees growing in even- and uneven-aged forest stands in Austria // Forest Ecology & Management. 1996. V. 80. P. 57–80.
NASA National Aeronautics and Space Administration. Goddard Institute for Space Studies. GISS Surface Temperature Analysis, 2020. https://data.giss.nasa.gov/gistemp/station_data_v4_globe
Neff J., Harden J., Gleixner G. Fire effect on soil organic matter content, composition and nutrients in boreal interior Alaska // Canadian J. Forest Research. 2005. V. 35. P. 2178–2187.
Nilsson U., Gemmel P. Changes in growth and allocation of growth in young Pinus sylvestris and Picea abies due to competition // Scandinavian J. Forest Research. 1993. V. 8. P. 213–222.
Pärn H. Relationships between radial growth of Scots pine and climate in the northeastern industrial region of Estonia // Forestry Studies. 2002. V. 36. P. 47–61.
Pärn H. Radial growth of conifers in regions of different cement dust loads // Proceedings of the Estonian Academy of Science. Biology, Ecology. 2006. V. 55(2). P. 108–122.
Regel K. Die Pflanzendecke der Halbinsel Kola // Memories de la faculte des sciences de l’universite de Lithuanie. Tail 2. Lapponia Ponoensis. 1923. P. 164–293.
Schweingruber F.H. Tree-Rings and environment. Dendroecology. Berne; Stuttgart; Vienna: Swiss Federal Institute for Forest, Snowand Landscape Research, 1996. 609 p.
Wilson R.J.S., Luckman B.H., Esper J. A 500-year dendroclimatic reconstruction of spring/summer precipitation from the lower Bavarian Forest Region, Germany // International Journal of Climatology. 2005. V. 25(5). P. 611–630.
Yarmishko V.T. Radial growth of Scots pine (Pinus sylvestis L.) in varied environment influenced be air pollution in the European North of Russia // Forestry Ideas (Bulgaria). 2015. V. 21(2 (50)). P. 96–105.
Дополнительные материалы отсутствуют.