

Лесоведение, 2021, № 5, стр. 494-508
Изменчивость признаков вегетативных органов лиственницы Гмелина в Забайкальском крае
В. П. Макаров a, *, О. Ф. Малых a, Т. В. Желибо a
a Институт природных ресурсов, экологии и криологии СО РАН
672014 Чита, ул. Недорезова, 16а, Россия
* E-mail: vm2853@mail.ru
Поступила в редакцию 22.10.2020
После доработки 27.12.2020
Принята к публикации 03.06.2021
Аннотация
В статье впервые обобщены результаты исследования изменчивости морфологических органов 38 популяций лиственницы Гмелина (Larix gmelinii (Rupr.) Rupr.) в разнообразных природных условиях Забайкальского края. Представлены особенности признаков ствола, кроны, хвои, интенсивности семеношения. Показана корреляционная связь признаков с условиями окружающей среды и между собой. Выделены насаждения, отличающиеся важными селекционными хозяйственно-полезными признаками и свойствами. Установлено, что изменчивость прямоствольности ствола в ряде популяций лиственницы достигает очень высокого уровня. Величина признака статистически достоверно связана с крутизной и экспозицией склона, типом гидротопа, а также высотой над уровнем моря и географическими координатами местообитания. Изменчивость протяженности кроны по отношению к высоте ствола находится в широких пределах, от очень низкого до высокого уровня. Обнаружена корреляционная связь признака с географической долготой и с типом гидротопа. Ширина кроны также варьирует в значительных пределах. Признак достоверно корреляционно связан с географическими координатами и высотой над уровнем моря, а также с экологическими условиями местообитания – крутизной и экспозицией склона, типом гидротопа. Средние величины угла отхождения ветвей от ствола в популяциях находятся в пределах 60°–90°. Редко встречаются деревья с тупым углом отхождения ветвей. Признак достоверно коррелирует с географическими и экологическими параметрами местообитания. Установлено, что число хвоинок в пучке коррелирует с географическими координатами и высотой над уровнем моря, а также экологическими условиями местообитания. Длина хвои корреляционно связана с географической широтой. Не установлена связь признака с экологическими условиями местообитания. Интенсивность семеношения образует тесную корреляционную связь с географической широтой, высотой над уровнем моря и типом гидротопа. Индивидуальная изменчивость признака достигает повышенного уровня. В статье также приведены результаты корреляционной связи морфологических признаков лиственницы Гмелина между собой.
Площадь Забайкальского края – около 6% территории Восточной Сибири. Покрытые лесной растительностью земли составляют 86.6% от общей площади региона. Доминирующей лесной породой в крае является лиственница Гмелина, занимающая около 57% от лесопокрытой площади.
Потепление климата на планете в значительной мере повлияло на состояние забайкальских лесов. Пожары привели к уничтожению и деградации значительной площади лесов, сокращению ценного генофонда древесных растений. Такая тенденция, вероятно, сохранится. В то же время, изученность разнообразия лесных сообществ и древесных пород на территории края остается низкой. Поэтому продолжение исследований в этой области необходимо для выявления наиболее ценных насаждений и экотипов, селекционной работы, организации охраны, успешного лесовосстановления и создания насаждений различного назначения.
Результаты исследований фенотипической и генетической изменчивости лиственницы в Забайкальском крае отражены в работах (Дылис, 1947; Круклис, Милютин, 1977; Дылис, 1981; Макаров и др., 2002; Милютин, 2003; Ирошников, 2004; Макаров, 2005; Барченков, Милютин, 2007; Абаимов, Коропачинский, 2008; Орешкова, 2009; Абаимов и др., 2010) и др. Однако этих исследований недостаточно, чтобы дать относительно полную характеристику множеству популяций лиственницы Гмелина. Значительная, столь обширного ареала, площадь Забайкальского края, горный характер рельефа, нахождение на границе тайги и сухих степей, между ареалами лиственниц сибирской и Каяндера, предполагает формирование в природе различных экотипов лиственницы Гмелина.
В соседнем регионе, Китайской народной республике, также проводятся исследования географической изменчивости лиственницы Гмелина и близких к ней видов (Yang Chuanping et al., 1991; Yu Bingjun, 1994; Yang Chuanping et al., 1997; Hu X.-S., Ennos R.A. 2001; Zhang Xin-bo, et al., 2001; Jia Qingbin et al., 2016; Dao Guang Zhu et al., 2020).
Новизна представленных исследований состоит в том, что получены более полные сведения по фенотипическому разнообразию вегетативных органов лиственницы Гмелина в Забайкалье. Они вносят дополнительный вклад в познание морфологического разнообразия лиственницы Гмелина. Цель исследований – изучить изменчивость фенотипических признаков вегетативных органов лиственницы Гмелина в Забайкальском крае, а также их связь с условиями окружающей среды.
ОБЪЕКТЫ И МЕТОДИКА ИССЛЕДОВАНИЯ
Исследования проведены в Забайкальском крае, в бассейнах рр. Аргунь, Ингода, Онон, Хилок и Шилка, в течение 2007–2012 гг. Пробные площади размещались в естественно разреженных насаждениях лиственницы Гмелина с низкой сомкнутостью крон, для снижения влияния конкурентных отношений между деревьями на развитие генеративных и вегетативных органов. Всего исследовано 38 насаждений лиственницы Гмелина (рис. 1).
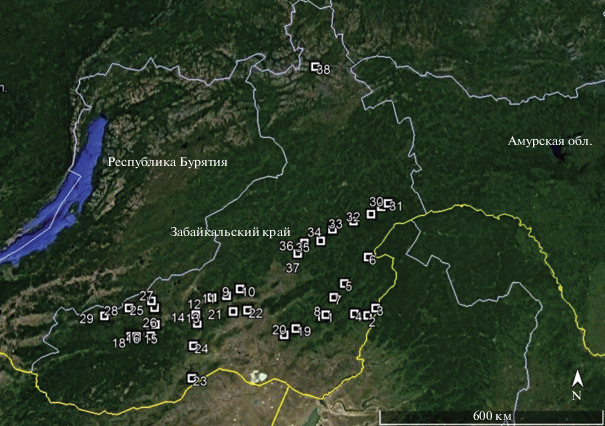
На пробных площадях фиксировались географические координаты, высота над уровнем моря, крутизна и направление склонов, тип гидротопа по П.С. Погребняку (1955), тип растительного сообщества (Панарин, 1965; Рысин, 2010), сомкнутость крон, средний диаметр и высота деревьев (табл. 1).
Таблица 1.
Характеристика пробных площадей лиственницы Гмелина
| Пробная площадь | Географические координаты, ° (система WGS-84) |
Абсолютная высота, м | Экспозиция, крутизна склона | Тип гидротопа | Тип лиственничного леса | Средний диаметр, см | Средняя высота, м | Сомкнутость крон, % | |
|---|---|---|---|---|---|---|---|---|---|
| с.ш. | в.д. | ||||||||
| 1 | 51.423978 | 118.135639 | 960 | Западный-35° | Сухой | Разнотравный | 35 | 20 | 50 |
| 2 | 51.338967 | 119.635657 | 646 | Северный-5° | Свежий | Ерниковый | 24 | 18 | 40 |
| 3 | 51.486078 | 119.939194 | 580 | Восточный-30° | Свежий | Разнотравный | 21 | 13 | 20 |
| 4 | 51.398283 | 119.163172 | 730 | Южный-5° | Свежий | Разнотравный | 22 | 12 | 30 |
| 5 | 52.086994 | 118.861792 | 690 | Северный-5° | Свежий | Разнотравный | 20 | 14 | 10 |
| 6 | 52.627753 | 119.80325 | 480 | Южный-5° | Сухой | Разнотравный | 26 | 17 | 30 |
| 7 | 51.797940 | 118.45280 | 651 | Южный-5° | Свежий | Разнотравный | 24 | 16 | 30 |
| 8 | 51.389720 | 118.00650 | 884 | Западный-25° | Свежий | Разнотравный | 27 | 16 | 10 |
| 9 | 52.064472 | 115.097389 | 792 | Западный-15° | Сухой | Разнотравный | 30 | 17 | 50 |
| 10 | 52.063944 | 115.095278 | 760 | Пологий-0° | Сырой | Приручейниковый | 47 | 21 | 50 |
| 11 | 51.919444 | 114.610778 | 715 | Западный-15° | Сухой | Разнотравный | 41 | 21 | 20 |
| 12 | 51.879139 | 114.091167 | 825 | Юго-вост.-10° | Свежий | Разнотравный | 32 | 16 | 10 |
| 13 | 51.306000 | 113.570139 | 833 | Пологий-0° | Сырой | Приручейниковый | 33 | 17 | 20 |
| 14 | 51.497222 | 113.516361 | 670 | Пологий-0 | Влажный | Приручейниковый | 35 | 16 | 10 |
| 15 | 51.262861 | 112.092139 | 854 | Пологий-0° | Сырой | Приручейниковый | 38 | 18 | 10 |
| 16 | 51.006500 | 111.936556 | 803 | Западный-35° | Сухой | Разнотравный | 36 | 22 | 50 |
| 17 | 50.977130 | 111.228044 | 1014 | Западный-5° | Свежий | Разнотравный | 32 | 20 | 10 |
| 18 | 50.979361 | 111.434972 | 905 | Юго-вост.-5° | Влажный | Разнотравный | 37 | 15 | 10 |
| 19 | 51.153110 | 117.047960 | 847 | Восточный-5° | Свежий | Разнотравный | 31 | 14 | 10 |
| 20 | 51.024890 | 116.632050 | 927 | Северо-зап.-15° | Свежий | Разнотравный | 25 | 13 | 20 |
| 21 | 51.568400 | 114.840000 | 852 | Южный-5° | Сырой | Приручейниковый | 46 | 19 | 30 |
| 22 | 51.591150 | 115.355350 | 850 | Северо-зап.-5° | Свежий | Разнотравный | 25 | 10 | 50 |
| 23 | 50.079300 | 113.398340 | 891 | Северо-зап.-5° | Сухой | Разнотравный | 41 | 15 | 10 |
| 24 | 50.80692 | 113.44053 | 880 | Пологий-0° | Свежий | Разнотравный | 35 | 16 | 20 |
| 25 | 51.633590 | 112.045610 | 938 | Пологий-0° | Влажный | приручейниковый | 60 | 20 | 5 |
| 26 | 51.622100 | 112.050670 | 953 | Северо-зап.-10° | Свежий | Приручейниковый | 32 | 22 | 50 |
| 27 | 51.781200 | 111.928370 | 910 | Южный-5° | Сухой | Разнотравный | 30 | 13 | 10 |
| 28 | 51.603250 | 111.132600 | 894 | Пологий-0° | Влажный | Приручейниковый | 30 | 15 | 30 |
| 29 | 51.398500 | 110.261990 | 807 | Пологий-0° | Сырой | Приручейниковый | 35 | 15 | 10 |
| 30 | 53.713117 | 120.444550 | 555 | Северный-5° | Сырой | Ерниковый | 28 | 18 | 30 |
| 31 | 53.774750 | 120.714317 | 742 | Северный-10° | Свежий | Лишайниковый скальный | 22 | 15 | 20 |
| 32 | 53.569317 | 120.032433 | 503 | Южный-40° | Сухой | Разнотравный | 38 | 19 | 10 |
| 33 | 53.446717 | 119.354317 | 604 | Западный-5° | Свежий | Разнотравный | 31 | 13 | 10 |
| 34 | 53.298350 | 118.541833 | 480 | Пологий-5° | Свежий | Разнотравный | 29 | 17 | 20 |
| 35 | 53.058667 | 118.084300 | 651 | Южный-5° | Свежий | Разнотравный | 44 | 19 | 10 |
| 36 | 53.030483 | 117.479850 | 884 | Западный-25° | Свежий | Лишайниковый скальный | 24 | 15 | 30 |
| 37 | 52.809683 | 117.217133 | 792 | Южный-15° | Сухой | Разнотравный | 36 | 14 | 5 |
| 38 | 56.910682 | 118.281972 | 705 | Пологий-0° | Влажный | Ерниковый | 28 | 18 | 30 |
На пробных площадях исследовали 30 деревьев. Измеряли диаметр ствола на высоте груди мерной вилкой, высоту ствола высотомером, угол отхождения ветвей от ствола в средней части кроны с помощью транспортира, определяли прямоствольность ствола по 5-бальной шкале: 5 – прямой одноствольный; 4 – прямой двуствольный; 3 – слабо искривленный одноствольный; 2 – слабо искривленный двуствольный; 1 – сильно и многократно искривленный). Интенсивность семеношения определяли по 5-бальной шкале: 1 – нет шишек; 2 – единично разбросанные шишки (не более 7 на каждый сук); 3 – до 20 шишек; 4 – до 50 шишек; 5 – вся крона обильно усеяна шишками. Также измеряли протяженность и ширину кроны. Форму кроны определяли как отношение протяженности кроны к ее ширине, относительную протяженность кроны – как процент отношения протяженности кроны к высоте ствола. Хвою отбирали с 10 деревьев на побегах 2 и 3-го годов жизни, с юго-восточной стороны средней части кроны. Определяли число хвоинок в 10 пучках. В лабораторных условиях измеряли длину 10 случайно отобранных хвоинок.
Для математического анализа полученных данных использовали программу Microsoft Office Excel, 2007. Изменчивость признаков вегетативных органов (эндогенную, индивидуальную и популяционную) оценивали по шкале С.А. Мамаева (C.V., % до 7 – уровень изменчивости очень низкий; 7–15 – низкий; 16–25 – средний; 26–35 – повышенный; 36–50 – высокий и более 50 – очень высокий) (Мамаев, 1972).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Прямоствольность
Прямоствольность - важный признак дерева, определяющий качество древесного сырья. Лучшие насаждения по этому признаку (средний балл более 4.5) находятся на пр. пп. 2, 4, 8, 9, 16, 17, 34, 38. Худшее качество стволов (средний балл около 3.0) отмечено в насаждениях на пр. пп. 7 и 22 (рис. 2).
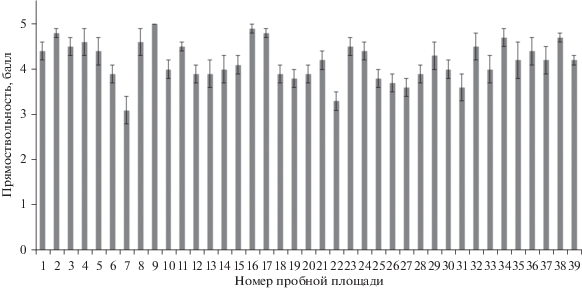
Средняя географическая изменчивость этого признака в регионе находится на низком уровне (C.V., % = 10.6). В то же время индивидуальная изменчивость признака в отдельных насаждениях достигает высокого (C.V., % = 31.3–39.4) и очень высокого значения (C.V., % = 41.2) (табл. 2).
Таблица 2.
Индивидуальная и географическая изменчивость признаков вегетативных органов лиственницы Гмелина
| Пробная площадь | Параметры | Прямо ствольность, балл |
Протя-женность кроны, м | Ширина кроны, м | Форма кроны | Относительная протяженность кроны, % | Угол отхождения ветвей от ствола дерева, ° | Число хвоинок в пучке | Длина хвои, мм | Интенсивность семеношения, балл |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | X ± mx | 4.4 ± 0.2 | 16.0± 0.3 | 11.7± 0.9 | 1.6± 0.2 | 95.6± 0.4 | 79.5± 2.1 | 32.5± 1.1 | 17.1± 0.6 | 4.9± 0.1 |
| min–max | 3.0–5.0 | 13.2–17.8 | 4.0–20.0 | 0.9–4.4 | 90.6–98.9 | 60.0–100.0 | 25.2–38.5 | 13.8–18.8 | 4.0–5.0 | |
| C.V., % | 21.5 | 9.1 | 33.4 | 48.6 | 2.0 | 11.9 | 10.7 | 10.6 | 7.6 | |
| 2 | X ± mx | 4.8± 0.1 | 16.4± 0.5 | 8.7± 0.4 | 2.0± 0.1 | 90.2± 0.4 | 81.9± 2.5 | 30.0± 1.4 | 19.9± 0.8 | 4.6± 0.1 |
| min–max | 3.0–5.0 | 10.7–21.7 | 5.0–14.0 | 1.3–3.0 | 84.4–94.3 | 45.0–90.0 | 24.2–37.6 | 15.7–23.5 | 3.0–5.0 | |
| C.V., % | 11.2 | 15.3 | 25.8 | 22.7 | 2.5 | 15.4 | 13.9 | 12.9 | 14.1 | |
| 3 | X ± mx | 4.5± 0.2 | 11.2± 1.0 | 7.1± 0.5 | 1.6± 0.1 | 83.5± 1.7 | 79.3± 2.8 | 28.6± 1.0 | 25.5± 1.2 | 3.8± 0.2 |
| min–max | 2.0–5.0 | 5.5–17.5 | 4.0–12.0 | 0.8–2.9 | 68.8–95.3 | 45.0–90.0 | 23.5–34.5 | 21.1–33.9 | 3.0–5.0 | |
| C.V., % | 21.9 | 40.5 | 29.1 | 41.1 | 9.1 | 16.0 | 11.0 | 15.6 | 22.1 | |
| 4 | X ± mx | 4.6± 0.3 | 11.1± 0.4 | 4.6± 0.2 | 2.5± 0.1 | 94.5± 1.1 | 90.0± 0.0 | 27.1± 1.5 | 24.3± 1.6 | 3.9± 0.2 |
| min–max | 3.0–5.0 | 9.5–13.3 | 4.0–5.5 | 2.1–3.0 | 87.7–99.2 | 90.0–90.0 | 21.6–39.1 | 17.4–32.9 | 3.0–5.0 | |
| C.V., % | 18.3 | 10.8 | 12.1 | 8.8 | 3.7 | 0.0 | 17.9 | 20.8 | 18.9 | |
| 5 | X ± mx | 4.4± 0.3 | 11.5± 0.6 | 6.2± 0.5 | 1.9± 0.1 | 84.1± 1.9 | 82.0± 1.3 | 31.0± 1.0 | 18.5± 1.0 | 4.4± 0.2 |
| min–max | 3.0–5.0 | 7.2–13.7 | 4.0–8.0 | 1.4–2.5 | 72.0–91.3 | 75.0–90.0 | 25.7–35.4 | 14.6–24.4 | 4.0–5.0 | |
| C.V., % | 22.0 | 17.6 | 23.6 | 19.3 | 7.1 | 5.1 | 10.2 | 16.4 | 11.7 | |
| 6 | X ± mx | 3.9± 0.2 | 15.0± 0.2 | 8.9± 0.4 | 1.8± 0.1 | 89.7± 0.5 | 84.1± 2.0 | 27.0± 1.3 | 25.2± 0.9 | 3.4± 0.2 |
| min–max | 3.0–5.0 | 12.5–16.7 | 6.0–12.0 | 1.1–2.7 | 83.3–92.8 | 60.0–95.0 | 21.3–34.9 | 22.1–32.0 | 2.0–5.0 | |
| C.V., % | 24.9 | 7.3 | 18.9 | 24.9 | 2.5 | 11.1 | 14.7 | 11.4 | 28.3 | |
| 7 | X ± mx | 3.1± 0.3 | 14.6± 0.3 | 7.6± 0.5 | 2.0± 0.1 | 89.5± 0.3 | 88.5± 0.8 | 25.6± 1.4 | 19.9± 0.9 | 3.7± 0.2 |
| min–max | 1.0–5.0 | 13.2–15.5 | 4.5–9.5 | 1.4–3.0 | 88.0–91.2 | 85.0–90.0 | 19.0–31.9 | 16.1–25.2 | 3.0–4.0 | |
| C.V., % | 32.1 | 6.5 | 20.5 | 22.1 | 1.0 | 2.7 | 16.9 | 14.1 | 13.1 | |
| 8 | X ± mx | 4.6± 0.3 | 13.2± 0.5 | 8.6± 0.6 | 1.6± 0.2 | 81.2± 2.6 | 89.0± 0.7 | 29.4± 1.1 | 19.0± 1.0 | 4.2± 0.2 |
| min–max | 3.0–5.0 | 10.2–15.0 | 4.5–11.5 | 0.9–3.0 | 61.1–88.7 | 85.0–90.0 | 24.5–35.1 | 15.0–24.3 | 3.0–5.0 | |
| C.V., % | 18.3 | 12.9 | 22.1 | 34.7 | 10.1 | 2.4 | 12.3 | 16.5 | 18.8 | |
| 9 | X ± mx | 5.0± 0.0 | 14.8± 0.6 | 7.5± 0.1 | 1.9± 0.1 | 87.9± 1.6 | 73.0± 2.6 | 32.8± 1.1 | 25.2± 1.0 | 4.2± 0.1 |
| min–max | 5.0–5.0 | 10.0–20.7 | 5.9–8.7 | 1.3–2.5 | 66.7–97.8 | 50.0–90.0 | 29.3–38.4 | 22.1–32.5 | 3.0–5.0 | |
| C.V., % | 0.0 | 23.6 | 11.0 | 22.3 | 9.7 | 19.8 | 10.2 | 12.9 | 14.5 | |
| 10 | X ± mx | 4.0± 0.2 | 15.4± 0.6 | 10.2± 0.3 | 1.5± 0.1 | 73.9± 1.6 | 81.2± 1.7 | 30.1± 0.9 | 27.8± 0.9 | 4.8± 0.1 |
| min–max | 2.0–5.0 | 10.0–20.0 | 7.1–13.6 | 1.0–2.3 | 52.4–90.0 | 60.0–90.0 | 25.0–33.3 | 24.2–33.8 | 4.0–5.0 | |
| C.V., % | 26.4 | 22.2 | 16.2 | 22.2 | 12.1 | 11.6 | 9.4 | 10.1 | 7.8 | |
| 11 | X ± mx | 4.5± 0.1 | 15.6± 0.5 | 8.8± 0.4 | 1.8± 0.1 | 72.5± 1.8 | 83.8± 1.4 | 38.0± 1.3 | 24.6± 1.3 | 4.3± 0.1 |
| min–max | 3.0–5.0 | 10.0–23.0 | 4.9–14.0 | 1.1–2.8 | 50.0–89.5 | 55.0–90.0 | 32.0–45.3 | 18.7–31.6 | 3.0–5.0 | |
| C.V., % | 18.1 | 18.9 | 23.6 | 24.4 | 13.7 | 9.4 | 11.0 | 16.8 | 13.9 | |
| 12 | X ± mx | 3.9± 0.2 | 14.3± 0.4 | 8.2± 0.3 | 1.8± 0.1 | 87.1± 1.2 | 75.5± 1.8 | 30.5± 1.2 | 25.0± 0.8 | 4.4± 0.1 |
| min–max | 3.0–5.0 | 11.0–18.0 | 6.9–11.7 | 1.3–2.5 | 73.3–96.4 | 60.0–90.0 | 25.3–36.6 | 21.4–29.5 | 4.0–5.0 | |
| C.V., % | 24.6 | 14.0 | 17.1 | 16.6 | 7.5 | 12.9 | 12.2 | 9.6 | 11.3 | |
| 13 | X ± mx | 3.9± 0.3 | 14.4± 0.4 | 8.2± 0.3 | 1.8± 0.1 | 87.5± 1.1 | 65.0± 1.7 | 33.3± 1.1 | 21.1± 0.9 | 4.4± 0.1 |
| min–max | 1.0–5.0 | 10.5–18.0 | 5.8–10.7 | 1.2–2.4 | 80.0–95.8 | 60.0–90.0 | 28.3–38.5 | 17.5–24.4 | 4.0–5.0 | |
| C.V., % | 41.2 | 15.9 | 20.2 | 22.1 | 6.7 | 14.4 | 10.5 | 13.5 | 11.3 | |
| 14 | X ± mx | 4.0± 0.3 | 13.6± 0.5 | 7.2± 0.3 | 1.9± 0.1 | 78.2± 1.2 | 79.5± 2.9 | 32.2± 1.8 | 24.5± 0.7 | 4.7± 0.1 |
| min–max | 1.0–5.0 | 9.5–19.0 | 5.2–9.6 | 1.4–2.8 | 67.5–87.1 | 40.0–90.0 | 23.4–42.2 | 20.6–27.2 | 4.0–5.0 | |
| C.V., % | 39.4 | 19.3 | 22.0 | 20.8 | 8.7 | 19.9 | 17.8 | 9.5 | 9.9 | |
| 15 | X ± mx | 4.1± 0.2 | 13.7± 0.4 | 7.9± 0.4 | 1.8± 0.1 | 77.8± 1.5 | 74.5± 2.7 | 33.1± 0.9 | 22.0± 1.2 | 4.5± 0.1 |
| min–max | 3.0–5.0 | 9.5–19.0 | 3.2–11.7 | 1.0–3.4 | 55.9–90.6 | 30.0–90.0 | 30.3–38.0 | 17.2–28.9 | 3.0–5.0 | |
| C.V., % | 24.4 | 17.8 | 26.9 | 27.8 | 10.8 | 20.0 | 8.8 | 17.1 | 13.9 | |
| 16 | X ± mx | 4.9± 0.1 | 18.8± 0.6 | 8.1± 0.2 | 2.3± 0.1 | 85.8± 1.4 | 84.5± 1.3 | 32.6± 1.4 | 20.4± 1.0 | 3.3± 0.1 |
| min–max | 3.0–5.0 | 13.0–26.0 | 5.1–11.0 | 1.7–3.3 | 59.1–97.1 | 50.0–90.0 | 28.9–41.4 | 15.3–24.9 | 3.0–4.0 | |
| C.V., % | 10.4 | 17.1 | 15.3 | 17.2 | 9.2 | 8.7 | 13.4 | 15.6 | 14.4 | |
| 17 | X ± mx | 4.8± 0.1 | 15.2± 0.5 | 7.6± 0.2 | 2.0± 0.1 | 77.1± 0.9 | 87.0± 0.6 | 30.3± 0.8 | 20.1± 0.9 | 4.1± 0.1 |
| min–max | 3.0–5.0 | 11.5–21.5 | 5.8–9.4 | 1.6–2.9 | 66.7–82.7 | 80.0–90.0 | 27.7–33.6 | 16.9–25.8 | 4.0–5.0 | |
| C.V., % | 12.7 | 18.5 | 14.2 | 17.8 | 6.3 | 3.9 | 8.4 | 13.9 | 7.4 | |
| 18 | X ± mx | 3.9± 0.2 | 12.9± 0.3 | 8.5± 0.3 | 1.5± 0.0 | 85.0± 0.6 | 80.0± 1.9 | 31.0± 1.4 | 19.1± 1.1 | 4.9± 0.0 |
| min–max | 2.0–5.0 | 10.0–15.8 | 6.4–11.2 | 1.1–2.1 | 76.9–90.0 | 60.0–90.0 | 21.5–36.7 | 11.8–24.7 | 4.0–5.0 | |
| C.V., % | 29.6 | 11.6 | 16.6 | 17.2 | 3.8 | 12.8 | 14.1 | 17.6 | 5.1 | |
| 19 | X ± mx | 3.8± 0.2 | 11.0± 0.3 | 6.9± 0.4 | 1.7± 0.1 | 80.0± 1.3 | 65.3± 3.1 | 29.8± 1.0 | 22.0± 0.8 | 4.4± 0.1 |
| min–max | 2.0–5.0 | 7.0–14.5 | 4.2–11.0 | 1.1–2.2 | 58.3–89.3 | 40.0–90.0 | 25.9–34.5 | 17.4–28.0 | 4.0–5.0 | |
| C.V., % | 27.9 | 17.2 | 29.7 | 17.5 | 9.4 | 26.7 | 11.1 | 11.9 | 11.4 | |
| 20 | X ± mx | 3.9± 0.2 | 10.9± 0.3 | 6.9± 0.3 | 1.7± 0.1 | 81.3± 1.5 | 76.0± 2.7 | 30.3± 1.2 | 20.1± 0.8 | 4.2± 0.1 |
| min–max | 1.0–5.0 | 6.5–12.5 | 3.0–9.5 | 1.2–3.8 | 65.0–95.5 | 45.0–90.0 | 24.5–36.5 | 16.7–25.2 | 3.0–5.0 | |
| C.V., % | 32.0 | 14.2 | 27.3 | 39.3 | 10.2 | 19.7 | 12.9 | 13.2 | 13.1 | |
| 21 | X ± mx | 4.2± 0.2 | 15.4± 0.3 | 10.0± 0.3 | 1.6± 0.0 | 82.7± 0.6 | 77.0± 3.1 | 24.0± 1.3 | 24.9± 0.9 | 3.0± 0.0 |
| min–max | 3.0–5.0 | 14.0–17.5 | 7.0–11.0 | 1.3–2.0 | 77.8–87.5 | 45.0–90.0 | 20.5–28.7 | 22.3–27.6 | 3.0–3.0 | |
| C.V., % | 23.7 | 9.2 | 15.8 | 15.4 | 3.8 | 21.9 | 12.3 | 8.4 | 0.0 | |
| 22 | X ± mx | 3.3± 0.2 | 6.9± 0.1 | 6.6± 0.2 | 1.1± 0.0 | 70.3± 1.8 | 80.8± 0.8 | 23.9± 1.9 | 30.4± 1.8 | 3.5± 0.1 |
| min–max | 2.0–5.0 | 5.5–7.8 | 5.0–8.5 | 0.8–1.6 | 55.0–86.7 | 75.0–90.0 | 19.1–29.7 | 23.7–36.0 | 3.0–4.0 | |
| C.V., % | 38.1 | 11.0 | 16.9 | 24.1 | 13.9 | 5.6 | 19.1 | 14.3 | 14.5 | |
| 23 | X ± mx | 4.5± 0.2 | 12.9± 0.2 | 10.2± 0.4 | 1.3± 0.1 | 85.5± 0.9 | 76.5± 2.1 | 33.5± 1.1 | 16.4± 0.7 | 4.9± 0.1 |
| min–max | 2.0–5.0 | 11.0–15.0 | 6.0–14.0 | 0.8–2.3 | 76.7–93.8 | 45.0–90.0 | 27.8–37.8 | 13.0–19.3 | 4.0–5.0 | |
| C.V., % | 23.2 | 10.0 | 23.9 | 29.1 | 6.0 | 15.2 | 10.2 | 12.9 | 6.2 | |
| 24 | X ± mx | 4.4± 0.2 | 14.1± 0.3 | 9.9± 0.3 | 1.5± 0.1 | 87.9± 0.9 | 75.0± 2.5 | 28.9± 1.9 | 20.3± 0.8 | 5.0± 0.0 |
| min–max | 3.0–5.0 | 11.1–16.3 | 7.0–13.0 | 1.1–2.0 | 76.3–92.9 | 50.0–90.0 | 17.6–38.0 | 16.6–24.2 | 5.0–5.0 | |
| C.V., % | 21.2 | 13.2 | 16.9 | 18.8 | 5.3 | 18.2 | 20.4 | 13.1 | 0.0 | |
| 25 | X ± mx | 3.8± 0.2 | 15.0± 0.4 | 7.5± 0.4 | 2.5± 0.2 | 83.8± 1.1 | 81.0± 2.3 | 32.5± 0.8 | 22.2± 1.0 | 5.0± 0.0 |
| min–max | 1.0–5.0 | 9.2–20.0 | 4.5–14.5 | 1.3–4.5 | 58.8–90.9 | 40.0–90.0 | 28.8–36.9 | 15.0–25.5 | 5.0–5.0 | |
| C.V., % | 31.3 | 18.0 | 34.2 | 36.6 | 8.7 | 18.1 | 8.1 | 13.8 | 0.0 | |
| 26 | X ± mx | 3.7± 0.2 | 12.5± 0.6 | 5.4± 0.3 | 2.5± 0.2 | 74.8± 2.5 | 91.0± 0.6 | 35.6± 1.4 | 18.9± 0.5 | 4.5± 0.1 |
| min–max | 2.0–5.0 | 6.0–17.8 | 3.0–10.0 | 1.3–4.5 | 50.0–91.4 | 90.0–100.0 | 28.4–42.9 | 15.9–21.3 | 4.0–5.0 | |
| C.V., % | 30.2 | 26.1 | 34.4 | 36.6 | 18.3 | 3.4 | 12.7 | 8.8 | 11.3 | |
| 27 | X ± mx | 3.6± 0.2 | 11.5± 0.3 | 6.5± 0.2 | 1.8± 0.1 | 85.0± 0.7 | 77.5± 3.5 | 35.6± 1.4 | 21.5± 0.8 | 4.2± 0.1 |
| min–max | 1.0–5.0 | 8.0–14.0 | 4.5–7.2 | 1.1–2.4 | 78.6–89.3 | 40.0–90.0 | 28.7–42.4 | 16.6–24.2 | 4.0–5.0 | |
| C.V., % | 36.2 | 14.1 | 12.7 | 17.6 | 4.4 | 24.4 | 12.8 | 11.5 | 9.7 | |
| 28 | X ± mx | 3.9± 0.2 | 12.4± 0.4 | 5.4± 0.1 | 2.4± 0.1 | 80.0± 1.1 | 86.0± 0.9 | 37.3± 1.0 | 19.0± 1.1 | 3.0± 0.0 |
| min–max | 3.0–5.0 | 8.0–15.0 | 4.0–6.0 | 1.5–3.8 | 70.6–88.2 | 80.0–90.0 | 32.6–43.2 | 13.2–24.0 | 3.0–3.0 | |
| C.V., % | 24.6 | 18.2 | 12.5 | 26.8 | 7.7 | 5.8 | 8.9 | 17.8 | 0.0 | |
| 29 | X ± mx | 4.3± 0.3 | 12.7± 0.3 | 7.0± 0.1 | 1.8± 0.1 | 83.9± 0.9 | 72.5± 3.4 | 33.5± 1.5 | 22.8± 1.0 | 3.8± 0.1 |
| min–max | 1.0–5.0 | 10.5–16.5 | 5.5–8.0 | 1.3–2.4 | 73.3–91.7 | 45.0–90.0 | 25.2–39.6 | 19.3–28.5 | 3.0–4.0 | |
| C.V., % | 33.5 | 14.7 | 10.3 | 18.6 | 5.7 | 25.9 | 14.0 | 13.6 | 10.7 | |
| 30 | X ± mx | 4.0± 0.2 | 12.8± 0.4 | 6.6± 0.3 | 2.0± 0.1 | 72.2± 2.7 | 89.8± 0.3 | 31.4± 1.6 | 19.0± 0.7 | 4.0± 0.0 |
| min–max | 3.0–5.0 | 10.0–15.0 | 4.5–8.5 | 1.1–3.3 | 55.0–88.2 | 85.0–90.0 | 26.3–41.0 | 15.3–22.9 | 4.0–4.0 | |
| C.V., % | 25.6 | 13.3 | 17.2 | 28.7 | 16.5 | 1.2 | 16.3 | 12.3 | 0.0 | |
| 31 | X ± mx | 3.6± 0.3 | 11.5± 0.8 | 6.1± 0.5 | 2.2± 0.2 | 75.1± 3.0 | 91.5± 1.1 | 29.6± 1.8 | 20.9± 0.8 | 4.3± 0.3 |
| min–max | 3.0–5.0 | 7.0–15.5 | 3.5–7.5 | 1.3–3.7 | 58.3–86.1 | 90.0–100.0 | 22.4–40.3 | 15.9–23.8 | 3.0–5.0 | |
| C.V., % | 26.8 | 23.4 | 26.5 | 34.3 | 12.6 | 3.7 | 19.5 | 11.5 | 19.1 | |
| 32 | X ± mx | 4.5± 0.3 | 16.8± 1.5 | 8.2± 0.7 | 2.1± 0.2 | 87.3± 2.4 | 88.5± 0.8 | 28.6± 1.4 | 19.5± 0.3 | 4.5± 0.2 |
| min–max | 3.0–5.0 | 8.4–23.2 | 5.5–12.5 | 1.2–3.6 | 76.4–96.7 | 85.0–90.0 | 24.4–35.1 | 18.4–21.8 | 4.0–5.0 | |
| C.V., % | 18.9 | 28.1 | 27.7 | 32.0 | 8.6 | 2.7 | 15.6 | 5.5 | 11.7 | |
| 33 | X ± mx | 4.0± 0.3 | 11.2± 0.4 | 8.6± 0.2 | 1.3± 0.1 | 87.4± 1.5 | 69.5± 5.9 | 27.2± 1.0 | 20.7± 0.6 | 3.2± 0.2 |
| min–max | 3.0–5.0 | 8.8–13.0 | 7.5–9.8 | 1.1–1.7 | 80.0–96.2 | 45.0–90.0 | 21.6–31.0 | 16.8–23.2 | 2.0–4.0 | |
| C.V., % | 23.6 | 12.4 | 9.0 | 15.5 | 5.5 | 27.0 | 11.8 | 8.5 | 19.8 | |
| 34 | X ± mx | 4.7± 0.2 | 14.3± 0.4 | 6.8± 0.5 | 2.2± 0.2 | 84.8± 1.8 | 61.5± 5.6 | 27.7± 0.8 | 17.5± 0.6 | 4.8± 0.1 |
| min–max | 3.0–5.0 | 12.0–16.5 | 4.5–10.5 | 1.3–3.2 | 75.0–91.7 | 45.0–90.0 | 23.4–32.4 | 15.1–19.8 | 4.0–5.0 | |
| C.V., % | 14.4 | 8.9 | 25.5 | 24.4 | 6.7 | 28.9 | 9.5 | 10.2 | 8.8 | |
| 35 | X ± mx | 4.2± 0.4 | 16.4± 1.2 | 10.2± 0.7 | 1.6± 0.1 | 86.2± 2.0 | 80.0± 4.1 | 32.5± 1.3 | 19.9± 0.6 | 4.3± 0.2 |
| min–max | 2.0–5.0 | 9.5–23.0 | 7.5–15.7 | 1.1–2.0 | 75.0–92.0 | 45.0–90.0 | 26.9–39.4 | 16.1–23.1 | 3.0–5.0 | |
| C.V., % | 31.3 | 23.7 | 22.3 | 21.9 | 7.3 | 16.4 | 12.3 | 9.0 | 15.7 | |
| 36 | X ± mx | 4.4± 0.3 | 12.1± 0.4 | 6.1± 0.3 | 2.0± 0.1 | 85.5± 1.2 | 86.5± 1.5 | 36.6± 1.3 | 20.8± 1.6 | 4.5± 0.2 |
| min–max | 3.0–5.0 | 9.5–13.0 | 4.8–7.7 | 1.4–2.6 | 79.2–89.3 | 80.0–90.0 | 31.2–44.8 | 14.7–29.1 | 3.0–5.0 | |
| C.V., % | 22.0 | 9.3 | 17.4 | 19.2 | 4.5 | 5.5 | 10.9 | 24.4 | 15.7 | |
| 37 | X ± mx | 4.2± 0.3 | 9.4± 1.1 | 8.9± 0.4 | 1.1± 0.1 | 64.7± 6.7 | 81.0± 5.3 | 32.5± 1.5 | 18.2± 0.4 | 5.0± 0.0 |
| min–max | 3.0–5.0 | 3.0–16.0 | 7.0–10.3 | 0.4–1.6 | 30.0–94.1 | 50.0–95.0 | 23.4–40.0 | 15.2–20.5 | 5.0–5.0 | |
| C.V., % | 24.6 | 38.1 | 14.4 | 31.8 | 32.6 | 20.5 | 14.8 | 7.8 | 0.0 | |
| 38 | X ± mx | 4.7± 0.1 | 16.0± 1.1 | 7.7± 0.6 | 2.1± 0.2 | 77.2± 2.5 | 89.5± 0.8 | 28.8± 1.3 | 17.2± 0.7 | 4.1± 0.2 |
| min–max | 4.0–5.0 | 10.0–23.0 | 6.0–12.0 | 1.4–3.3 | 58.8–88.5 | 85.0–95.0 | 24.6–35.3 | 14.4–21.1 | 3.0–5.0 | |
| C.V., % | 9.9 | 22.9 | 25.3 | 27.2 | 10.9 | 3.0 | 12.8 | 13.0 | 11.6 | |
| Географическая изменчивость | X ± mx | 4.2± 0.1 | 13.5± 0.4 | 7.8± 0.2 | 1.8± 0.1 | 82.5± 1.1 | 80.4± 1.2 | 30.9± 0.5 | 21.4± 0.5 | 4.2± 0.1 |
| min–max | 3.1–5.0 | 6.9–18.8 | 4.6–11.7 | 1.1–2.5 | 64.7–95.6 | 61.5–91.5 | 23.9–38.0 | 16.4–30.4 | 3.0–5.0 | |
| C.V., % | 10.6 | 17.2 | 19.7 | 19.3 | 8.1 | 9.3 | 10.8 | 14.7 | 13.2 |
Коэффициент корреляции Пирсона свидетельствует о положительной достоверной связи прямоствольности ствола с крутизной склона (R = 0.36). Коэффициент корреляции Спирмена также указывает на положительную достоверную связь прямоствольности с крутизной склона (P = = 0.58), но также с экспозицией склона (P = 0.59), типом гидротопа (P = 0.46); на отрицательную достоверную связь с координатами северной широты (P = –0.69), восточной долготы (P = –0.35), высотой над уровнем моря (P = –0.61).
Протяженность кроны
Протяженность кроны зависит как от условий произрастания, так и от наследственных свойств лиственницы. Средняя протяженность кроны в исследованных насаждениях равна 13.5 м. Географическая изменчивость признака характеризуется средним уровнем (C.V., % = 17.2). Индивидуальная изменчивость различна, в насаждениях от очень низкого (C.V., % = 6.5) до высокого (C.V., % = 40.5).
Лиственницы с наиболее протяженной кроной (20 м и более) отмечены в насаждения на пр.пп. 2, 9, 10, 11, 16, 25, 32, 35, 38. Низкая протяженность кроны лиственницы (3–5 м), встречается на пр. пп. 22, 26, 37. Коэффициент корреляции Спирмена указывает на отрицательную достоверную связь признака с географической долготой (P = –0.45).
Ширина кроны
Ширина кроны, также как и ее протяженность, зависит от условий произрастания растения и наследственных свойств. Ширина кроны напрямую влияет на полноту, а также плотность создаваемых насаждений. По мнению Л.И. Милютина (Круклис, Милютин, 1977) ширококронные деревья лиственницы являются более быстрорастущими. Существует прямая зависимость между урожаем шишек на деревьях лиственницы и объемами их крон. Поэтому ширококронные деревья лиственницы являются более ценными для лесного хозяйства.
Средняя ширина кроны в исследованных насаждения равна 7.8 м. Географическая изменчивость признака характеризуется средним уровнем (C.V., % = 19.7). Индивидуальная изменчивость признака в насаждениях находится в пределах от низкого (C.V., % = 9.0–12.5) до высокого (C.V., % = = 33.4–34.4). Наиболее широкая крона (20 м) обнаружена в насаждении на пр. пп. 1, в районе с. Кличка, а самая узкая крона (около 3 м) в насаждениях на пр. пп. 15, 20, 26.
Обнаружена отрицательная достоверная связь ширины кроны с географической широтой (P = –0.55) и восточной долготы (P = –0.49), а также высотой над уровнем моря (P = –0.43). Положительная достоверная связь признака отмечается с крутизной (P = 0.35) и экспозицией склона (P = 0.50), а также типом гидротопа (P = 0.39).
Форма кроны
Форма кроны лиственницы отличается большим разнообразием, зависит от возраста и условий произрастания, а также наследственных свойств. В литературе встречаются термины: пирамидальная, конусовидная, колонновидная, яйцевидно-шатровидная и др. (Дылис, 1947). По данным Л.И. Милютина (Круклис, Милютин, 1977) при селекционной инвентаризации насаждений лиственницы Чекановского в Забайкальском крае выявлено, что, несмотря на некоторую зависимость встречаемости различных форм кроны от условий произрастания, доля участия той или иной формы кроны во всех насаждениях относительно стабильна. Практически во всех популяциях преобладает цилиндрическая форма кроны, встречаются также флагообразная, шаровидная, зонтикообразная и коническая формы. Согласно исследованиям на севере Забайкалья морфологическое разнообразие лиственницы по форме кроны обусловлено преимущественно высотной поясностью. С увеличением абсолютной высоты произрастания форма кроны меняется от яйцевидной до столбовидной (Барченков, 2008). В наших исследованиях для упрощения оценки форма кроны определена как отношение протяженности кроны к ее ширине. Средняя величина этого отношения в исследованных насаждениях равна 1.8. Географическая изменчивость признака характеризуется средним уровнем (C.V., % = = 19.3). Индивидуальная изменчивость находится в пределах от очень низкой (C.V., % = 8.8) до очень высокой (C.V., % = 48.6).
В ряде насаждений встречаются деревья, у которых протяженность кроны превосходит ширину в четыре и более раза, например на пр. пп. 1, 20, 25, 28, 31, 32, а также деревья с приблизительно равной протяженностью и шириной кроны: это, например, деревья на пр. пп. 1, 3, 6 и др. (табл. 2).
Выявлена отрицательная достоверная связь признака с широтой (P = –0.53) и долготой местопроизрастания (P = –0.24), а также положительная достоверная связь с крутизной (P = 0.38) и экспозицией склона (P = 0.62), типом гидротопа (P = 0.51).
Относительная протяженность кроны
Относительная протяженность кроны характеризует в процентах долю протяженности кроны к высоте ствола дерева. Средняя величина относительной протяженности кроны в регионе 82.5 ± ± 1.1%. Средние пределы признака в насаждениях 64.7–95.6%. Минимальная относительная протяженность кроны отмечена в насаждениях на пр. пп. 11, 26, 37 (30.0–50.0%), а максимальное значение на пр. пп. 1, 4 (98.9–99.2%).
Признак характеризуется низким уровнем географической изменчивости (C.V., % = 8.1). Индивидуальная изменчивость признака находится в пределах от очень низкой до средней. Исключение составляет насаждение на пр. пп. 37, в районе пос. Жирекен. Здесь индивидуальная изменчивость признака находится на высоком уровне (C.V., % = 32.6). Что, возможно, связано с изреживанием древостоя после пожаров. Отмечена положительная достоверная связь признака с географической долготой (P = 0.39) и отрицательная связь – с типом гидротопа (P = –0.47).
Угол отхождения ветвей от ствола
Имеются сведения о связи этого признака с темпом роста растения, семенной продуктивностью (Дебринюк, 2011). Угол отхождения сучьев от ствола является генетически обусловленным признаком, показателем быстрорастущих особей сосны обыкновенной, наиболее устойчивых к рекреационным воздействиям (Тырченкова, 2017).
Географическая изменчивость признака в регионе низкая (C.V., % = 9.3). Средний угол отхождения ветвей от ствола дерева равен 80.4 ± 1.2°. Средние величины признака в исследованных насаждениях находятся в пределах 61.5°–91.5°. Индивидуальная изменчивость угла отхождения ветвей от ствола в насаждениях находится в пределах от очень низкой до повышенной.
Минимальные значения признака 30°–45° отмечены в насаждениях на пр .пп. 2. 3, 14, 15, 27, 29, 33, 34, а максимальные (100°) в ценопопуляциях на пр. пп. 1, 26, 31. Прямой угол отхождения ветвей характерен для насаждений на пр. пп. 4, 8, 26, 31, 30, 38 (рис. 3).
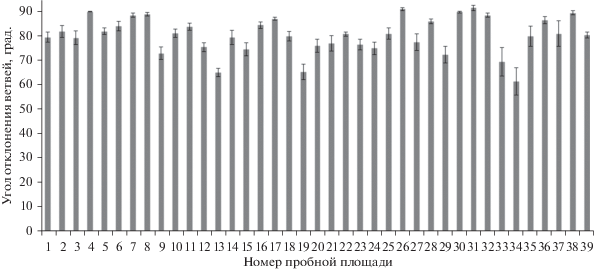
Выявлена положительная корреляция признака с географической широтой (P = 0.56) и географической долготой (P = 0.50), с высотой над уровнем моря (P = 0.43), а также отрицательная корреляционная связь с крутизной склона (P = –0.35) и типом гидротопа (P = –0.47).
Число хвоинок в пучке
Хвоя – самый чувствительный орган, быстро реагирующий на условия окружающей среды и определяющий рост и развитие других органов растения, поэтому размеры хвои очень изменчивы даже в пределах кроны одного дерева (Барченков, Милютин, 2007). Ряд авторов указывает на видоспецифичность этого признака и отмечает большее число хвоинок у лиственницы Гмелина по сравнению с лиственницами сибирской и Каяндера (Круклис, Милютин, 1977; Абаимов, Коропачинский, 1984).
Среднее число хвоинок в пучке на исследованных пробных площадях равно 30.9 ± 0.5. Средняя величина признака в насаждениях варьирует в пределах 23.9–38.0 шт. Географическая изменчивость числа хвоинок в пучке в регионе характеризуется низким уровнем (C.V., % = 10.8). Индивидуальная изменчивость признака находится в пределах от низкой до средней. На отдельных деревьях среднее число хвои в пучке достигает 45 шт. (пр. пп. 11, 36). Минимальное же число хвоинок в пучке на деревьях составляет 18–19 шт.
Средняя эндогенная изменчивость признака в регионе характеризуется средним уровнем (C.V., % = 16.4). В насаждениях она варьирует в пределах от низкой (C.V., % = 9.7) до повышенной (C.V., % = 22.3).
Отмечена отрицательная достоверная корреляция признака с координатами восточной долготы (R = –0.48) и положительная связь с высотой над уровнем моря (R = 0.37). Ранговый коэффициент корреляции указывает на положительную достоверную связь с географической широтой (P = 0.54) и высотой над уровнем моря (P = 0.69) и отрицательную связь с крутизной (P = –0.37) и экспозицией склона (P = –0.73), с типом гидротопа (P = –0.65).
Длина хвои
Этот признак ряд исследователей также считает видоспецифичным (Круклис, Милютин, 1977). В среднем, у лиственницы Гмелина хвоя короче, чем у лиственниц сибирской, Чекановского и Каяндера. Однако отмечается значительное влияние на длину хвои степени увлажнения почвы и климатических условий местообитания. Поэтому использование признака для определения видов лиственницы затруднительно (Абаимов, Коропачиснкий, 1984). На севере Забайкалья в пойменной популяции длина хвои значительно превышает показатели, зарегистрированные в среднегорье. Это свидетельствует о том, что изменчивость этого признака в значительной степени определяется влажностью климата (Барченков, 2008).
Географическая изменчивость признака характеризуется средним уровнем (C.V., % = 14.7). Средняя длина хвои в исследованных насаждениях региона равна 21.4 ± 0.5 мм. Средняя длина хвои на пробной площади колеблется в пределах 16.4–30.4 мм. Индивидуальная изменчивость длины хвои в ценопопуляциях находится в пределах от очень низкой до повышенной. Минимальная средняя длина хвои на дереве (11.8 мм) обнаружена в насаждении на пр. пп. 18, а максимальная 36.0 мм на пр. пп. 22.
Средняя эндогенная изменчивость признака в регионе характеризуется средним уровнем (C.V., % = 18.4), в насаждениях она варьирует в пределах от низкой (C.V., % = 12.0) до повышенной (C.V., % = 25.0).
В наших исследованиях признак положительно достоверно связан с географической широтой (P = 0.44). Возможно, это объясняется большим количеством осадков в северных районах региона. Влияния степени увлажнения почвы на длину хвои лиственницы не обнаружено.
Интенсивность семеношения
Важный признак, связанный с урожайностью семян лиственницы; показывает примерное число шишек в кроне растущего дерева в баллах. Хотя он и не относится к вегетативным органам, интересно выяснить его связь с рассматриваемыми в статье признаками.
Средний балл интенсивности семеношения в регионе 4.2 ± 0.1. Средняя величина в насаждениях на пробных площадях изменяется от 3.0 до 5.0 баллов (рис. 4).
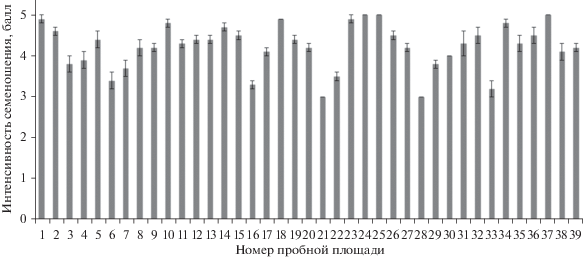
Географическая изменчивость признака характеризуется средним уровнем (C.V., % = 13.2). Индивидуальная изменчивость в популяциях различна и находится в пределах от очень низкой до повышенной (C.V., % = 0.0 до 28.3). Можно выделить насаждения с высокой интенсивностью семеношения (более 4.5 балла). Это популяции лиственницы на пр. пп. 1, 10, 14, 18, 24, 25, 34, 37.
Признак достоверно корреляционно связан с географической широтой (P = 0.34), высотой над уровнем моря (P = 0.69) и типом гидротопа (P = = 0.46).
Связь признаков вегетативных органов между собой
Для проведения отбора лиственницы с желательными признаками и свойствами важно знать взаимную связь между ними. Для установления корреляционной связи использовали два коэффициента корреляции – Пирсона и Спирмена (табл. 3).
Таблица 3.
Корреляция признаков вегетативных органов лиственницы
| Признак | Коэффициент корреляции Пирсона (R) | Критерий T | Коэффициент корреляции Спирмена (P) | Критерий T |
|---|---|---|---|---|
| Прямоствольность | ||||
| Протяженность кроны | 0.45 | 3.01 | 0.30 | 1.86 |
| Ширина кроны | 0.20 | 1.20 | 0.20 | 1.25 |
| Форма кроны | 0.14 | 0.85 | 0.36 | 2.30 |
| Относительная протяженность кроны | 0.23 | 1.40 | 0.14 | 0.84 |
| Угол отхождения ветвей от ствола дерева | –0.02 | –0.12 | 0.31 | 1.94 |
| Число хвоинок в пучке | 0.12 | 0.72 | 0.47 | 3.22 |
| Длина хвои | –0.23 | –1.41 | 0.16 | 0.94 |
| Высота ствола | 0.13 | 0.80 | 0.38 | 2.44 |
| Диаметр ствола | 0.17 | 1.03 | 0.41 | 2.67 |
| Интенсивность семеношения | 0.24 | 1.44 | 0.42 | 2.79 |
| Протяженность кроны | ||||
| Ширина кроны | 0.50 | 3.38 | 0.85 | 9.53 |
| Форма кроны | 0.36 | 2.27 | 0.39 | 2.55 |
| Относительная протяженность кроны | 0.37 | 2.38 | 0.36 | 2.30 |
| Угол отхождения ветвей от ствола дерева | 0.09 | 0.53 | 0.31 | 1.99 |
| Число хвоинок в пучке | 0.10 | 0.61 | 0.62 | 4.79 |
| Длина хвои | –0.18 | –1.10 | 0.22 | 1.37 |
| Высота ствола | 0.31 | 1.90 | 0.71 | 6.10 |
| Диаметр ствола | 0.40 | 2.58 | 0.67 | 5.44 |
| Интенсивность семеношения | 0.23 | 1.43 | 0.57 | 4.11 |
| Ширина кроны | ||||
| Форма кроны | –0.57 | –4.10 | 0.04 | 0.27 |
| Относительная протяженность кроны | 0.18 | 1.09 | 0.20 | 1.19 |
| Угол отхождения ветвей от ствола дерева | –0.22 | –1.35 | 0.19 | 1.18 |
| Число хвоинок в пучке | –0.11 | –0.67 | 0.53 | 3.72 |
| Длина хвои | –0.08 | –0.45 | 0.23 | 1.45 |
| Высота ствола | 0.19 | 1.17 | 0.66 | 5.31 |
| Диаметр ствола | 0.37 | 2.39 | 0.64 | 5.02 |
| Интенсивность семеношения | 0.18 | 1.09 | 0.48 | 3.30 |
| Форма кроны | ||||
| Относительная протяженность кроны | 0.21 | 1.26 | 0.46 | 3.14 |
| Угол отхождения ветвей от ствола дерева | 0.36 | 2.27 | 0.68 | 5.60 |
| Число хвоинок в пучке | 0.24 | 1.47 | 0.51 | 3.55 |
| Длина хвои | –0.18 | –1.10 | 0.38 | 2.49 |
| Высота ствола | 0.03 | 0.20 | 0.54 | 3.82 |
| Диаметр ствола | –0.07 | –0.39 | 0.47 | 3.20 |
| Интенсивность семеношения | 0.10 | 0.57 | 0.44 | 2.91 |
| Относительная протяженность кроны | ||||
| Угол отхождения ветвей от ствола дерева | –0.20 | –1.23 | 0.17 | 1.06 |
| Число хвоинок в пучке | –0.15 | –0.89 | 0.16 | 0.94 |
| Длина хвои | –0.14 | –0.81 | –0.02 | –0.13 |
| Высота ствола | –0.25 | –1.53 | 0.21 | 1.32 |
| Диаметр ствола | –0.16 | –0.93 | 0.18 | 1.12 |
| Интенсивность семеношения | 0.09 | 0.53 | 0.34 | 2.14 |
| Угол отхождения ветвей от ствола дерева | ||||
| Число хвоинок в пучке | 0.04 | 0.24 | 0.30 | 1.88 |
| Длина хвои | –0.10 | –0.57 | 0.23 | 1.41 |
| Высота ствола | 0.22 | 1.34 | 0.56 | 4.02 |
| Диаметр ствола | –0.17 | –1.01 | 0.35 | 2.23 |
| Интенсивность семеношения | –0.17 | –1.05 | 0.23 | 1.43 |
| Число хвоинок в пучке | ||||
| Длина хвои | –0.27 | –1.66 | 0.35 | 2.21 |
| Высота ствола | –0.08 | –0.49 | 0.67 | 5.35 |
| Диаметр ствола | 0.19 | 1.12 | 0.84 | 9.22 |
| Интенсивность семеношения | 0.07 | 0.40 | 0.69 | 5.78 |
| Длина хвои | ||||
| Высота ствола | –0.20 | –1.22 | 0.25 | 1.57 |
| Диаметр ствола | –0.06 | –0.38 | 0.43 | 2.82 |
| Интенсивность семеношения | –0.36 | –2.28 | 0.10 | 0.61 |
| Высота ствола | ||||
| Диаметр ствола | 0.62 | 4.72 | 0.82 | 8.45 |
| Интенсивность семеношения | 0.10 | 0.61 | 0.61 | 4.57 |
| Диаметр ствола | ||||
| Интенсивность семеношения | 0.20 | 1.21 | 0.71 | 6.00 |
Согласно коэффициенту Пирсона, статистически достоверные связи образованы между прямоствольностью и протяженностью кроны (R = 0.45), шириной кроны и диаметром ствола (R = 0.37), а также между длиной хвои и интенсивностью семеношения (R = –0.36).
Коэффициент Спирмена обнаруживает большее число достоверных связей между признаками. Интересны связи числа хвоинок в пучке с прямоствольностью (P = 0.47), протяженностью кроны (P = 0.62), формой кроны (P = 0.51), высотой (P = 0.67), и диаметром ствола (P = 0.84), а также с интенсивностью семеношения (P = 0.69); длины хвои – с формой кроны (P = 0.38), числом хвоинок в пучке (P = 0.35) и диаметром ствола (P = 0.43).
Важный признак лиственницы, влияющий на урожайность семян – интенсивность семеношения. Он достоверно образует корреляционную связь с высотой (P = 0.61) и диаметром ствола (P = 0.71), протяженностью (P = 0.57) и шириной кроны (P = 0.48), относительной протяженностью (P = 0.34) и формой кроны (P = 0.44), прямоствольностью (P = 0.42), а также числом хвоинок в пучке (P = 0.69).
Угол отхождения ветвей от ствола коррелятивно связан с высотой (P = 0.56) и диаметром ствола (P = 0.35), а также формой кроны (P = 0.68).
Выводы. 1. Географическая изменчивость признаков вегетативных органов лиственницы Гмелина в регионе характеризуется низким и средним уровнями. Большей вариабельностью отличаются протяженность, ширина и форма кроны, а также длина хвои.
2. Индивидуальная изменчивость формы кроны достигает очень высокого уровня; прямоствольности, протяженности и ширины кроны – высокого уровня; угла отхождения ветвей, длины хвои и интенсивности семеношения – повышенного уровня; относительной протяженности кроны – среднего уровня изменчивости.
3. Эндогенная изменчивость длины хвои и числа хвоинок в пучке достигает в популяциях повышенного уровня.
4. Высокой теснотой корреляционной связи (P = 0.7–0.8) характеризуются протяженность кроны с шириной и высотой ствола; диаметр ствола с числом хвоинок в пучке и интенсивностью семеношения.
5. Исследованные признаки вегетативных органов лиственницы Гмелина корреляционно достоверно связаны с географическими координатами, высотой над уровнем моря и экологическими условиями места произрастания.
6. В регионе встречаются насаждения лиственницы Гмелина, значительно отличающиеся параметрами признаков вегетативных органов, что позволяет судить о перспективности обора ценопопуляций и деревьев для селекции и хозяйственного использования.
Список литературы
Абаимов А.П., Коропачинский И.Ю. Лиственницы Гмелина и Каяндера. Новосибирск. Наука, 1984. 120 с.
Абаимов А.П., Адрианова И.Ю., Артюкова Е.В., Барченков А.П., Васюткина Е.А., Гуков Г.В., Журавлев Ю.Н., Зырянова О.А., Исаев А.П., Катышев А.И., Карпюк Т.В., Квитко О.В., Князева С.Г., Козыренко М.М., Константинов Ю.М., Ларионова А.Я., Милютин Л.И., Милютина И.Л., Михалева Л.Г., Муратова Е.Н., Орешкова Н.В., Пименов А.В., Реунова Г.Д., Седельникова Т.С., Сизых О.А., Судачкова Н.Е., Чикидов И.И. Биоразнообразие лиственниц Азиатской России. Новосибирск: ГЕО, 2010. 160 с.
Барченков А.П. Изменчивость лиственницы в Северном Забайкалье (Становое нагорье) // Вестник Томского государственного университета. Биология. 2008. № 3(4). С. 7–15.
Барченков А.П., Милютин Л.И. Морфологическая изменчивость лиственницы в Средней Сибири // Хвойные бореальной зоны. 2007. Т. 24. № 4–5. С. 367–372.
Дебринюк Ю.М. Формовое разнообразие лиственницы европейской (Larix decidua Mill.) как интродуцента в Прикарпатье // Плодоводство, семеноводство, интродукция древесных растений. 2011. № 14. С. 38–41.
Дылис Н.В. Лиственница. М.: Лесн. пром-ость., 1981. 96 с.
Дылис Н.В. Сибирская лиственница. Материалы к систематике, географии и истории. М.: изд-во МОИП, 1947. 137 с.
Ирошников А.И. Лиственницы России. Биоразнообразие и селекция. М.: ВНИИЛМ, 2004. 182 с.
Круклис М.В., Милютин Л.И. Лиственница Чекановского. М.: Наука, 1977. 212 с.
Макаров В.П. Изменчивость морфологических видов и климатипов лиственницы в географических культурах (Восточное Забайкалье) // Лесоведение. 2005. № 4. С. 67–75.
Макаров В.П., Бобринев В.П., Милютин Л.И. Географические культуры лиственницы в Восточном Забайкалье. Улан-Удэ: Изд-во Бурятского НЦ СО РАН, 2002. 187 с.
Мамаев С.А. Формы внутривидовой изменчивости древесных растений. М.: Наука, 1972. 284 с.
Милютин Л.И. Биоразнообразие лиственниц России // Хвойные бореальной зоны. 2003. Т. 21. № 1. С. 6–9.
Орешкова Н.В. Популяционно-генетические параметры лиственницы Гмелина в Восточном Забайкалье (Читинская область) // Вестник Томского государственного университета. 2009. № 28. С. 193–198.
Панарин И.И. Типы лиственничных лесов Читинской области. М.: Наука, 1965. 103 с.
Погребняк П.С. Основы лесной типологии. Киев: Изд-во АН УССР, 1955. 456 с.
Рысин Л.П. Лиственничные леса России. М.: Товарищество научных изданий КМК, 2010. 343 с.
Тырченкова И.В. Фенотипические признаки устойчивости сосны обыкновенной к рекреационному воздействию // Лесотехнический журнал. 2017. № 2. С. 115–121.
Zhu Dao Guang, Xu Xue Hong, Yang Li Bin, Fan Yang, Li Jin Bo, Cui Fu Xing, Ye Wan Hu, Ni Hong Wei. Characteristics of leaves, stems and roots of dominant trees in Larix gmelinii forest in the cold temperate zone // Scienta sinica Vitae. 2020. V. 50. № 4. P. 398–405.
Zhang Xin-bo, Feng Jing-hua, Ren Jian-ru. Morphological variance and natural types' division of plus trees of Larix principis-rupprechtii Mayr // J. Northeast Forestry University. 2001. V. 12. P. 123–127.
Yu Bingjun. A study on the geographic variation and selection of the best provenance of Larix olgensis // J. Northeast Forestry University. 1994. V. 5. P.1–9.
Yang Chuanping, Zhang Wei, Yu Bingjun, Qin Sihua. Study on geographic variation of Dahurian larch in China // J. Northeast Forestry University. 1991. V. 2. P. 34–41.
Yang Chuanping, Liu Guifeng, Xu Jiachun, Cai Baoming, Xia Dean, Yang Shuwen, Zhang Peigao. Geographic variation pattern of Larix olgensis // J. Forestry Research. 1997. V. 8. P. 3–9.
Jia QingBin, Zhang Hanguo, Zhang Lei, Zhang Haixiao. Variation analysis of hybrid Larch families and superior famliies selection // J. Northeast Forestry University. 2016. V. 4. P. 1–7.
Hu X.-S., Ennos R.A. Population structure and genetic relationships of taxa in the Larix Gmelinii complex in China // Forest Geneyics. 2001. V. 8. № 3. P. 225–232.
Дополнительные материалы отсутствуют.