

Лесоведение, 2021, № 6, стр. 627-644
Поливариантность развития подроста ели сибирской в темнохвойных лесах северного Предуралья
Т. Ю. Браславская a, *, А. С. Ефименко a
a Центр по проблемам экологии и продуктивности лесов РАН
117997 Москва, Профсоюзная ул., д. 84/32, стр. 14, Россия
* E-mail: t-braslavskaya@yandex.ru
Поступила в редакцию 26.03.2021
После доработки 25.05.2021
Принята к публикации 21.07.2021
Аннотация
На основе анализа морфометрических показателей и хода линейного роста модельных растений (высотой до 3.5 м) сформулированы диагностические признаки онтогенетических состояний im1, im2, v1 и уровней жизненности в них. В качестве критерия перспективности подроста использована сделанная на основе анализа хода роста модельных растений оценка способности достичь высоты генеративных деревьев, сформировавшихся в тех же условиях. Выявлено, что достичь высоты генеративных деревьев в сопоставимом с ними возрасте способен подрост в онтогенетических состояниях im2 и v1, имеющих нормальную жизненность; эти растения отнесены к категории перспективного подроста. Признаки внешнего строения у перспективного подроста – относительная протяженность живой кроны не менее 4/5 от общей высоты растения и число живых ветвей более 3-х во всех годичных мутовках на стволе. К категории средней перспективности отнесен подрост в онтогенетических состояниях im2 и v1 с пониженной жизненностью, к категории сомнительной перспективности – подрост в онтогенетических состояниях im2 и v1 с низкой жизненностью. У подроста в онтогенетическом состоянии im1 перспективы еще недостаточно определенные.
Для понимания и прогнозирования лесовозобновительных процессов требуется знать закономерности развития растений лесообразующих видов, начиная с ранних этапов. В лесоведении и лесоводстве к подросту относят растения лесообразующих видов, имеющие календарный возраст от 2–5 (в условиях средней и северной тайги – от 10) лет; завершением этого этапа считается переход растений в категорию молодняка (молодого древостоя), для которой в качестве диагностического критерия указано начало двух процессов – интенсивного роста растений и смыкания их крон (ОСТ 56-108-98). Определения подроста, произрастающего в лесных сообществах, нередко дополняют критериями размеров: диаметра ствола на высоте 1.3 м или высоты – например, указанием, что его высота не превышает 25% от высоты господствующего яруса древостоя (ГОСТ 18486-87; Методы …, 2002). Для оценки качества подроста, то есть его жизнеспособности и перспектив развития, многие исследователи проводили анализ его морфометрических показателей и скорости роста (Касимов, 1960; Дерябин, Букштынов, 1970; Злобин, 1970, 1975; Титов, 1970; Максимов, 1971; Алексеев, 1978; Диагнозы …, 1989; Antos et al., 2000; Романовский, 2001; Claveau et al., 2002; Prévost et al., 2016). Показано, в том числе и для темнохвойных видов, что темпы развития подроста сильно зависят от фитоценотических условий, в которых он произрастает, поэтому наблюдаемое у него разнообразие габитуса (внешнего облика), размеров и морфологии часто не проявляет тесной связи с его календарным возрастом (Диагнозы …, 1989; Романовский, 2001; Евстигнеев, 2014; Ставрова и др., 2017). Большая вариабельность темпов индивидуального развития (онтогенеза) – это вообще характерное свойство растений, что обобщено в концепции их биологического возраста (Gatzuk et al., 1980; Диагнозы …, 1989; Злобин, 1989; Evstigneev, Korotkov, 2016; Omelko et al., 2018). В рамках этой концепции в онтогенезе выделяют по морфологическим критериям категории биологического возраста – онтогенетические состояния, а в каждом из них описывают морфологическую дифференциацию растений по уровням жизненного состояния (жизненности, виталитета), характеризующего жизнеспособность (Gatzuk et al., 1980; Злобин, 1989); такая дифференциация получила название поливариантности развития (Gatzuk et al., 1980; Диагнозы …, 1989). У лесообразующих видов к категории подроста относятся растения нескольких прегенеративных онтогенетических состояний (Диагнозы …, 1989; Evstigneev, Korotkov, 2016): младшего имматурного (im1), старшего имматурного (im2), младшего виргинильного (v1) и старшего виргинильного (v2), в которое у многих видов включают также и растения с диаметром ствола 6 см и более (Диагнозы …, 1989; Романовский, 2001), относящиеся к древостою (Методы …, 2002).
В связи с большим разнообразием лесорастительных условий в России требуется изучать в каждом регионе особенности развития подроста основных лесообразователей. У видов ели развитие исследовано с применением концепции биологического возраста в лесах северной тайги (Ставрова и др., 2017; Мишко, 2019), подзоны хвойно-широколиственных лесов (Романовский, 2001; Давыдычев, Кулагин, 2010). В исследованиях подроста в среднетаежных темнохвойных лесах Северного Предуралья (Бобкова, Бессонов, 2009; Бобкова и др., 2010) эта концепция не применялась. Цель нашей работы – охарактеризовать для ели сибирской диагностические признаки начальных онтогенетических состояний и уровней жизненности, удобные для применения во время массовых полевых учетов подроста и информативные для оценки перспектив его дальнейшего развития.
Большинство подходов к оценке качества подроста основано на анализе таких его показателей как средняя длина годичного линейного прироста ствола за несколько последних лет или ее отношение к средней (за те же годы) длине годичных линейных приростов на ветвях 2-го порядка, общая форма кроны (результат соотношения между скоростью удлинения ствола и боковых ветвей), относительная протяженность живой кроны (Касимов, 1960; Науменко, 1962; Дерябин, Букштынов, 1970; Злобин, 1970, 1975; Титов, 1970; Максимов, 1971; Алексеев, 1978; Диагнозы …, 1989; Antos et al., 2000; Романовский, 2001; Claveau et al., 2002; Энциклопедия …, 2006; Prévost et al., 2016; Ставрова и др., 2017). Также учитываются и другие параметры габитуса и структуры кроны: максимальный порядок ветвления (Диагнозы …, 1989; Романовский, 2001), общее число живых ветвей 2-го порядка (Мишко, 2019) или их среднее число в ежегодно формируемых хвойными видами мутовках на стволе (Дерябин, Букштынов, 1970; Злобин, 1975; Prévost et al., 2016), или же менее четко определенный показатель “густота кроны” (Алексеев, 1989; Методы …, 2002). В описаниях онтогенетических состояний и уровней жизненности учитывается формирование корки на стволе и высота, до которой она развита (Диагнозы …, 1989; Романовский, 2001; Мишко, 2019). Нередко обращается внимание на наличие повреждений кроны и фотосинтезирующих органов различными патогенами: долю участия сухих ветвей в верхней половине кроны (Алексеев, 1989; Методы …, 2002), густоту облиствения и наличие хлорозов и некрозов на листьях/хвое (Касимов, 1960; Дерябин, Букштынов, 1970; Титов, 1970; Алексеев, 1989; Методы …, 2002; Мишко, 2019). Так как ухудшение состояния древесных растений под влиянием патогенов может быть непродолжительным и обратимым, то оценки должны основываться на различных показателях, учитывать реакцию и на эти факторы, и на фоновые фитоценотические условия. Действующие правила лесовосстановления определяют следующие признаки жизнеспособного подроста хвойных видов: густая хвоя, зеленая или темно-зеленая окраска хвои, заметно выраженная мутовчатость, островершинная или конусообразная симметричная густая или средней густоты крона протяженностью до 1/3 высоты ствола в группах и до 1/2 высоты ствола – при одиночном размещении, прирост по высоте за последние 3–5 лет не утрачен, прирост вершинного побега равен (или более) приросту боковых ветвей верхней половины кроны, стволики прямые неповрежденные, гладкая или мелкочешуйчатая кора без лишайников (Правила …, 2020).
В ходе проведенных исследований были выявлены корреляции между различными морфометрическими показателями подроста и сделано заключение, что для оценки его жизненного состояния достаточно рассматривать только некоторые показатели – наиболее информативные (тесно связанные с большим числом остальных) и удобные в практическом применении (Титов, 1970; Злобин, 1975, 1989; Алексеев, 1978).
Одним из закономерных явлений в развитии деревьев лесообразующих видов и проявлением их благополучия является ускорение линейного (в высоту) и радиального (по диаметру) роста на достаточно раннем этапе – в период завершения имматурного состояния и перехода в виргинильное (Серебряков, 1962; Диагнозы …, 1989; Evstigneev, Korotkov, 2016). Это учитывалось в подходах к оценке качества подроста: например, было предложено учитывать не только длину годичных линейных приростов ствола, но и проявление в их динамике тенденции неубывания (Дерябин, Букштынов, 1970; Методы …, 2002). Но немногие современные исследователи (Antos et al., 2020) проводят анализ хода линейного роста ствола – из-за его трудоемкости даже при изучении подроста.
Некоторые исследователи обосновывали оценки жизненного состояния и/или перспектив развития древесных растений результатами сравнения их размеров и возраста со значениями, выбранными в качестве эталонов (Декатов, Кузнецов, 1968; Абатуров и др., 2012; Antos et al., 2020).
Задачей нашего исследования было охарактеризовать начальные онтогенетические состояния ели сибирской и уровней жизненности в них теми морфометрическими показателями, которые отражают многолетние тенденции роста, согласованы с остальными показателями и удобны для оценки во время полевых исследований популяций.
ОБЪЕКТЫ И МЕТОДИКА
Исследования проведены в 2012 г. в лесном массиве Верхне-Печорского лесничества Печоро-Илычского государственного природного биосферного заповедника (Республика Коми, Троицко-Печорский район), в бассейне р. Большой Порожней (левого притока р. Печоры в верхнем течении). Эта территория относится к Западно-Уральской предгорной увалисто-равнинной провинции Уральской горно-увалисто-равнинной области (Смолоногов, 1995), в системе природной зональности – к средней тайге (Атлас …, 2001), в пределах Печоро-Илычского заповедника – к восточной части Предгорного ландшафтного района (Бобрецов, Теплова, 2000) с абсолютными высотными отметками 250–300 м. Климат Предгорного района – умеренно-континентальный с умеренно суровой зимой. Среднегодовая температура воздуха равна минус 0.4°С; средняя температура января – минус 18.5°С, июля – 16.6°С. Период с отрицательными температурами воздуха длится 175–185 дней, безморозный период – 80–110, вегетационный период – 140–150, период активной вегетации – 90–100 дней (в это время сумма температур воздуха выше 5°С составляет 1600–1800°С). Среднемноголетняя годовая сумма осадков – 627 мм. Снеговой покров устанавливается в середине октября – начале ноября и держится около 180–190 дней, средняя высота снегового покрова – 105 см (Атлас …, 2001; Бобрецов, Теплова, 2000). Почвы в бассейне р. Большой Порожней – средне- и тяжелосуглинистые горно-лесные. В растительном покрове бассейна преобладают перестойные темнохвойные леса с древостоями, сформированными елью сибирской (Picea obovata Ledeb.) и пихтой сибирской (Abies sibirica Ledeb.) с участием березы (Betula pubescens Ehrh.) и примесью кедра (Pinus sibirica Du Tour). Пихто-ельники чернично-зеленомошные и бореально-мелкотравные произрастают на иллювиально-гумусово-железистых (нередко – глееватых) почвах, пихто-ельники высокотравные – на бурых лесных грубогумусированных (Смирнов, Браславская, 2010; Семиколенных и др., 2013).
Подрост ели сибирской исследован в бореально-мелкотравном пихто-ельнике с кедром и березой, который расположен (62°02′17.0135″ с.ш., 58°57′51.7483″ в.д.) на абсолютной высоте приблизительно 280 м, на склоне восточной экспозиции с уклоном 5°. Размещение деревьев в сообществе – групповое, в пологе крон выражены “окна” площадью до 70–80 м2, общая сомкнутость полога – приблизительно 50%. Сообщество сформировалось после пирогенных нарушений (Смирнова и др., 2007), давность которых не менее 180–200 лет (датировка по возрасту самых старых деревьев ели в послепожарном древостое – см. ниже). Для характеристики древостоя и подроста была заложена постоянная пробная площадь (пр. пл.) размером 50 × 50 м, на которой проведен сплошной перечет всех стволов древесных растений с диаметром от 2 см на высоте 1.3 м и закартированы их основания и горизонтальные проекции крон с помощью электронно-измерительного комплекса Field-map® (IFER, Field-Map Technology, 2009, http://www.field-map.cz). При перечете стволы с диаметром от 6 см отнесены к древостою, менее 6 см – к подросту (Методы …, 2002); у всех стволов регистрировали вид, диаметр на высоте 1.3 м, высоту – общую и прикрепления нижних ветвей живой кроны, состояние (живые/сухие) и повреждения; у хвойных деревьев проверяли наличие шишек. Для определения календарного возраста деревьев и наиболее крупных экземпляров подроста были выборочно взяты керны в основании стволов (на минимально возможной высоте) с помощью возрастного бурава. Характеристика древостоя в сообществе (на основе результатов перечета) представлена в таблице 1. Численность подроста с диаметром ствола 2–6 см – 196 шт. · га–1, состав – 5Е5Пх (где Е – ель, Пх – пихта), средняя высота – 2.87 м, средний диаметр – 4 см; при этом подрост высотой более 3.5 м был единичен.
Таблица 1.
Некоторые таксационные характеристики пр. пл. 9 в бореально-мелкотравном пихто-ельнике
| Формула древостоя* | Порода | Число стволов на 1 га | Возраст, лет | Средние параметры деревьев |
Сумма площадей поперечного сечения стволов, м2 · га–1 | Сумма площадей проекций крон, м2 · га–1 | Запас древесины, м3 · га–1 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| живые | сухие | всего | мин. | макс. | среднее | диаметр (на высоте 1.3 м), см | высота, м | площадь проекции кроны, м2 | живая | сухая | всего | ||||
| 7Е 1Пх 1К 1Б | Ель | 340 | 100 | 440 | 99 | 203 | 143 | 28.5 | 22.1 | 7.8 | 24.5 | 2642.9 | 261.5 | 27.9 | 289.4 |
| Пихта | 224 | 52 | 276 | 78 | 146 | 116 | 16.8 | 14.4 | 5.5 | 5.6 | 1207.7 | 39.6 | 6.7 | 46.3 | |
| Кедр | 8 | 0 | 8 | 150 | 150 | 150 | 63.2 | 28.5 | 38.8 | 2.6 | 310.6 | 35.3 | 0.0 | 35.3 | |
| Береза | 56 | 20 | 76 | Нет данных | Нет данных | Нет данных | 30.1 | 19.9 | 16.6 | 3.8 | 929.5 | 36.6 | 2.3 | 38.9 | |
| Всего | 628 | 172 | 800 | – | – | – | – | – | – | 36.5 | 5090.4 | 373.0 | 36.9 | 409.9 | |
В этом сообществе отобрано 45 модельных растений подроста ели высотой от 0.17 м (минимальная высота уже начавших ветвиться, т.е. имматурных, растений) до 3.5 м. В сообществе не было подроста с признаками ослабления, учитываемыми в шкале В.А. Алексеева (1989), т.е. с усыханием ветвей в верхней части кроны и/или хлорозами и некрозами хвои; выбранные модельные растения относились в этой шкале к категории здоровых. В ходе полевых исследований у каждой модели были определены доступные для быстрого измерения морфометрические показатели, используемые в диагностике онтогенетических состояний и жизненности (Злобин, 1975; Алексеев, 1978; Диагнозы …, 1989; Романовский, 2001; Ставрова и др., 2017; Мишко, 2019): общая высота и высота нижней границы живой кроны (уровень прикрепления к стволу самых нижних живых ветвей 2-го порядка); 4 радиуса кроны (максимальный, противоположный ему и 2 перпендикулярных) на уровне ее нижней границы; длины годичных линейных приростов ствола за последние 5 лет (границы приростов определяли по расположению мутовок ветвей или рубцам от почечных чешуй); диаметр ствола у основания. Также определяли длину покрытой коркой нижней части ствола, длину его верхней части, покрытой первичной корой (не опробковевшей покровной тканью с незасохшими листовыми подушками под хвоинками). Кроме того, подсчитывали общее число живых ветвей 2-го порядка, а на самых нижних из них – максимальный порядок ветвления.
Для определения календарного возраста и анализа хода линейного роста у моделей высотой более 0.5 м отобрали серии поперечных спилов ствола с шагом 10 см от корневой шейки до последнего годичного прироста (оставшуюся часть верхушки ствола сохраняли вместе со спилами). Во время камерального этапа работы каждый спил был высушен до воздушно-сухого состояния, затем отшлифован шлифовальной машиной МШУ-1,3П-180 с использованием абразивного материала крупной (Р100) и мелкой (Р320, Р600, P1500) зернистости. С помощью стереоскопического микроскопа МБС-10 при увеличении 80× на каждом спиле подсчитано число годичных колец; из-за неясностей, возникших в ходе этого подсчета, 3 модели были исключены из дальнейшего анализа. У моделей высотой до 0.5 м для определения календарного возраста и анализа хода линейного роста подсчитывали годичные линейные приросты стволика (все они были визуально различимы, границы определяли по расположению мутовок ветвей и рубцам от почечных чешуй) и измеряли их длины.
Во время камерального этапа работы вычисления, построение графиков и диаграмм проводили в программных пакетах Excel 2010, Statistica 6.0 и PAST (Hammer et al., 2001).
Для всех моделей путем интерполяции рассчитаны значения высот ствола при заданных значениях возраста с шагом 5 лет (5, 10, 15, 20 лет и т.д.) и построены графики хода роста в высоту. Выраженность тенденции к ускорению или замедлению роста верифицировали путем расчета корреляции (коэффициент Спирмена) между рангами секций ствола (выделенных, начиная от корневой шейки, и ранжированных в порядке снизу вверх) и значениями скорости роста, соответствующими этим секциям. У моделей высотой до 0.5 м секции ствола выделили по границам годичных линейных приростов. У моделей высотой более 0.5 м выделили секции с шагом 20 см (поскольку годичные линейные приросты ствола иногда превышали 10 см, то есть шаг в сериях спилов) и рассчитали для каждой секции среднюю скорость линейного роста (см·год–1).
Чтобы оценить теоретические перспективы развития исследованных модельных растений ели, рассчитали ход их роста в высоту до 200 лет (максимальный возраст, выявленный у генеративных деревьев ели в данном сообществе); графики, построенные по результатам расчета, сравнили с высотами и возрастами генеративных деревьев ели в данном сообществе – нанесли на диаграмму соответствующие им точки. Расчеты провели на основе функции А. Митчерлиха, известной в отечественном лесоводстве также под названием формулы В.Н. Дракина – Д.И. Вуевского (Кузьмичев, 1977) и описывающей зависимость высоты дерева от его возраста: H = Hmax × (1 – e–kA)m, где H – высота, A – календарный возраст, Hmax – максимальная возможная в данных условиях высота, k и m – коэффициенты. Согласно свойствам этой функции, при малых значениях возраста рост в высоту происходит медленно, затем постепенно ускоряется, а потом снова постепенно замедляется. Для каждого модельного растения индивидуально рассчитаны параметры этой функции Hmax, k и m на основе выявленного при анализе хода роста ряда высот и соответствующих им возрастов; расчет проводили по алгоритму О.А. Трулля (1966), для подбора функции, наиболее точно описывающей ряд фактических данных, использовали метод наименьших квадратов. У подобранной функции определяли среднеквадратичную ошибку и отклонение расчетного значения общей высоты модельного растения от фактического.
Для моделей также рассчитали дополнительные морфометрические показатели: длину живой кроны (Lcr = H – Hcrb, где Lcr – длина живой кроны, H – общая высота модельного растения, Hcrb – высота нижней границы живой кроны); относительную протяженность живой кроны (Lcr : H); средний диаметр кроны внизу (Dcrb – как полусумму 4-х радиусов); отношение длины кроны к ее среднему диаметру внизу (Lcr : Dcrb); относительную протяженность зоны корки на стволе (Lcrust : H, где Lcrust – длина покрытой коркой нижней части); относительную протяженность зоны первичной коры на стволе (Lprim : H, где Lprim – длина верхней части ствола, покрытой первичной корой); среднее число ветвей 2-го порядка в одной годичной мутовке на стволе (как отношение NLivBr : Acr, где NLivBr – общее число живых ветвей 2-го порядка, Acr – возраст ствола на уровне ответвления самых нижних живых ветвей 2-го порядка, то есть продолжительность нарастания живой части кроны).
Анализ дифференциации модельных растений по сходству/различию комплекса морфометрических показателей провели при помощи многомерной ординации – неметрического многомерного шкалирования (NMDS). Мерой сходства/различия в ординации служило Евклидово расстояние. Его рассчитали с использованием показателей H, Lcr : H, Lcr : Dcrb, ZH5 (среднее значение линейного прироста ствола за последние 5 лет), Lcrust : H, Lprim : H, NLivBr : Acr; выбор определялся тем, что их значения нетрудно оценивать глазомерно во время полевых учетов подроста. Выбранные морфометрические показатели слабо коррелируют друг с другом (табл. 2), т.е. в их совместном анализе информация не избыточна. Для выполнения расчетов Евклидова расстояния значения этих показателей были нормированы (из каждого значения вычиталось среднее в соответствующей выборке, полученная разность делилась на стандартное отклонение выборки). По итогам ординации были сформированы группы наиболее сходных между собой модельных растений. Эти группы интерпретировали как уровни жизненности в разных онтогенетических состояниях; при интерпретации были учтены результаты анализа хода роста у растений в составе каждой группы. Для выделенных групп проверена статистическая значимость различий (U-тест Манна–Уитни) по всем морфометрическим показателям и календарному возрасту.
Таблица 2.
Матрица корреляций между морфометрическими показателями и календарным возрастом модельных растений ели
| H* | D0 | Dcrb | Lcr : H | Lprim : H | Lcrust : H | Lcr : Dcrb | ZH5 | NLivBr : Acr | BrOrd | |
|---|---|---|---|---|---|---|---|---|---|---|
| H | ||||||||||
| D0 | 0.973** | |||||||||
| Dcrb | 0.898 | 0.894 | ||||||||
| Lcr : H | х*** | х | х | |||||||
| Lprim : H | –0.586 | –0.639 | –0.500 | 0.625 | ||||||
| Lcrust : H | 0.572 | 0.603 | 0.529 | –0.562 | –0.725 | |||||
| Lcr : Dcrb | –0.329 | –0.373 | –0.519 | 0.538 | 0.596 | –0.566 | ||||
| ZH5 | 0.632 | 0.567 | 0.725 | 0.559 | х | х | х | |||
| NLivBr : Acr | х | х | 0.398 | 0.567 | 0.366 | –0.323 | х | 0.757 | ||
| BrOrd | 0.882 | 0.921 | 0.826 | х | –0.743 | 0.553 | –0.408 | 0.445 | х | |
| A | 0.789 | 0.825 | 0.682 | –0.541 | –0.919 | 0.798 | –0.562 | х | х | 0.851 |
Примечание. A – календарный возраст, BrOrd – максимальный порядок ветвления, Dcrb – средний диаметр живой кроны внизу, D0 – диаметр ствола в основании, H – общая высота, Lcr : Dcrb – отношение протяженности живой кроны к ее среднему диаметру внизу, Lcr : H – относительная протяженность живой кроны, Lcrust : H – относительная протяженность зоны корки на стволе, Lprim : H – относительная протяженность зоны первичной коры на стволе, NLivBr : Acr – среднее число живых ветвей в годичной мутовке на стволе, ZH5 – средний линейный прирост ствола за последние 5 лет. Жирный шрифт – p < 0.01; простой шрифт – p < 0.05, “х” — корреляции не значимы.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Графики хода линейного роста показывают (рис. 1), что устойчивая тенденция его ускорения была выражена почти у половины (11 из 27) модельных растений, имевших высоту более 0.5 м (рис. 1а). У большинства таких растений эта тенденция проявилась при достижении высоты 0.4–0.5 м (в возрасте 15–20 лет), у отдельных (№№ 99, 110) – при высоте 0.25–0.3 м в более молодом возрасте (10 лет) или при высоте более 1 м уже в возрасте 45–60 лет (№ 109). У одной из моделей (№ 84: возраст – около 100 лет, высота – чуть более 2 м), наоборот, итоговой тенденцией было замедление роста; но при этом в начальные 5 лет своей жизни растение росло гораздо быстрее, чем все остальные в таком же возрасте, а затем рост постепенно замедлялся. У других моделей высотой более 0.5 м ход роста не имел выраженных тенденций: в течение их жизни многократно чередовались не очень продолжительные периоды относительно быстрого и медленного роста. Выявленное разнообразие многолетних тенденций роста обусловлено неравномерной освещенностью в сообществе, где размещение деревьев – групповое, а общая сомкнутость полога древостоя – не очень большая.
Рис. 1.
Графики хода линейного роста модельных растений высотой более 0.5 м (а) и до 0.5 м в сравнении с некоторыми растениями более 0.5 м (б). 1 – рост не имеет значимых тенденций, 2 – тенденция ускорения (p < 0.05), 3 – тенденция ускорения (p < 0.01), 4 – тенденция замедления (p < 0.01). На рис. 1 и 2 на графиках подписаны номера характерных моделей, обсуждаемых в тексте.
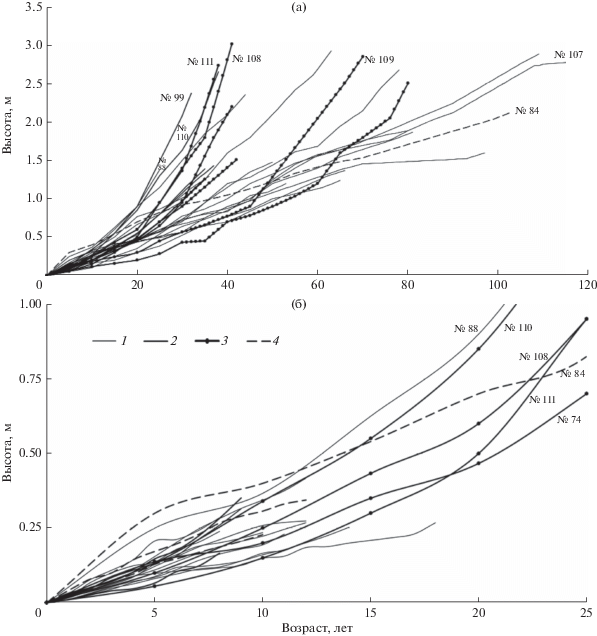
Среди 15 моделей высотой до 0.5 м (рис. 1б) у двух была выражена тенденция к ускорению роста, еще у двух – к замедлению, а у остальных значимых тенденций не было из-за резких межгодовых колебаний длин годичных приростов. При этом у всех моделей высотой до 0.5 м скорости роста были в пределах того же диапазона значений, в каком они были и у моделей высотой более 0.5 м в начальные 15–20 лет их жизни. Это свидетельствует, что подрост обеих этих генераций поселялся и вначале развивался в сходных условиях. Но так как у многих моделей высотой более 0.5 м после завершения начального периода развития существенно менялись темпы роста (рис. 1а), можно и для моделей высотой до 0.5 м предполагать, что их рост в момент исследования еще не приобрел определенного характера, даже если выявлены значимые тенденции. В связи с этим для моделей высотой до 0.5 м не выполнялся расчетный прогноз хода роста.
Для прогноза развития моделей высотой более 0.5 м оснований больше, особенно для растений с большим календарным возрастом. Графики их роста в высоту до 200 лет, построенные на основе функции Митчерлиха с индивидуально подобранными параметрами, представлены на рис. 2. У семи из этих моделей рассчитанное значение высоты в том возрасте, в котором растение находилось в момент исследования (23–125 лет), отклоняется от фактического значения не более чем на 10%. Из них только у одной (№ 88: 23 г., высота – 1.38 м, ZH5 – 12 см · год–1) расчет прогнозирует достижение к 200-летнему возрасту хотя бы минимальной высоты генеративных деревьев, произрастающих в том же сообществе (причем подобранная для нее функция занижает высоту в момент исследования на 9% по сравнению с фактической, то есть если бы график был точнее, то проходил бы выше и достигал минимальной высоты генеративных деревьев при меньшем значении возраста). Для других моделей, у которых удовлетворительна точность подобранных функций роста, рассчитанный прогноз демонстрирует неспособность достичь высоты генеративных деревьев в сопоставимом с ними возрасте. Две из них (№№ 84 и 107) относятся к категории нежизнеспособных по критериям действующих правил лесовосстановления (относительная протяженность живой кроны составляет менее 1/2 от общей высоты, в расположении ветвей на стволе не выражена мутовчатость). Но несколько моделей с таким неблагоприятным прогнозом соответствуют критериям (предложенным разными исследователями и современным официальным) жизнеспособного подроста: эти модели имеют относительную протяженность живой кроны более 1/2 от общей высоты, конусообразную крону, выраженное мутовчатое ветвление ствола. Их возраст – 33–78 лет, а негативные тенденции их роста, ставшие причиной неблагоприятного прогноза – небольшие значения ZH5 (5–8 см · год–1) и отсутствие выраженного ускорения роста.
Рис. 2.
Графики прогноза линейного роста модельных растений (высотой более 0.5 м). 1–3 – для возраста в момент исследования точность высоты, рассчитанной на основе функции Митчерлиха, находится в пределах 10%: 1 – тенденция ускорения роста (p < 0.01), 2 – рост не имеет значимых тенденций, 3 – тенденция замедления (p < 0.01); 4–5 – для возраста в момент исследования рассчитанная высота занижена более чем на 10%: 4 – тенденция ускорения роста (p < 0.01), 5 –тенденция ускорения роста (p < 0.05); 6–8 – для возраста в момент исследования рассчитанная высота завышена более чем на 10%: 6 – тенденция ускорения роста (p < 0.01), 7 – тенденция ускорения роста (p < 0.05), 8 – рост не имеет значимых тенденций; 9 – возрасты и высоты генеративных деревьев в том же сообществе; 10 – минимальная высота, выявленная у генеративных деревьев.
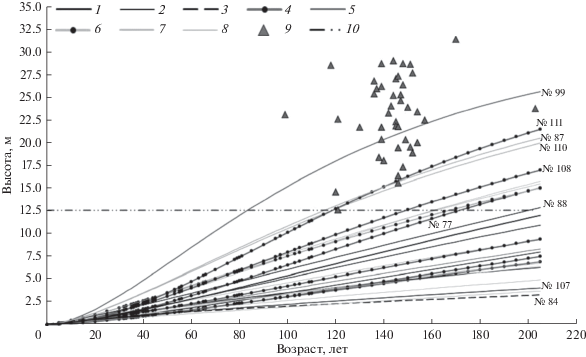
У остальных моделей рассчитанные по функциям роста значения высот в возрасте на момент исследования отклонялись от фактических более чем на 10%, поэтому при обсуждении полученных прогнозов их роста необходимо учитывать, что точность не является высокой. Однако в некоторых характерных случаях это не становится препятствием для заключения о перспективах развития. Так, у четырех растений, для которых теоретически прогнозируется достижение высоты генеративных деревьев (хотя бы минимальной), оказалось занижено значение высоты, рассчитанное для возраста в момент исследования: № 99 – на 46%, № 108 – на 23%, № 111 – на 21%, № 77 – на 20%. Если бы построенные для этих моделей графики проходили через точки с их фактическими значениями высот в этом возрасте, то располагались бы выше, т.е. еще более гарантированно давали бы прогноз достижения высоты генеративных деревьев; поэтому можно обоснованно считать перспективы развития этих моделей вполне благополучными.
У пяти других моделей, для которых тоже прогнозируется способность достичь высоты генеративных деревьев, завышена высота, рассчитанная для возраста в момент исследования: например, у № 87 – на 119%, у № 110 – на 20%, у остальных трех – на 25–178%; в связи с этим нет достаточных оснований считать, что перспективы этих моделей благополучны. Также нельзя сделать уверенное заключение о перспективах тех моделей, у которых не прогнозируется способность достичь высоты генеративных деревьев, но при этом оказалась низкой точность расчетов высот на основе подобранных функций.
Рост в высоту имеет значение для деревьев ели, возможно, не только как способ преодолеть затенение. Есть данные, что у темнохвойных видов в верхней части кроны преобладают женские шишки и что их морфогенез регулируется водным потенциалом в клетках апикальных меристем, поэтому переходу дерева в генеративное состояние способствует достижение некоторой пороговой высоты: после этого апикальные меристемы на верхних ветвях периодически испытывают недостаток воды и под его влиянием формируют женские шишки (Минина, 1960; Некрасова, Рябинков, 1978). Исходя из этого, возможность для подроста ели достичь высоты, наблюдаемой у генеративных деревьев в тех же условиях, – это критерий перспективности, обоснованный не только хозяйственными соображениями, но и популяционно-биологическими свойствами вида. Скорость роста в высоту нередко рассматривалась в предшествующих исследованиях подроста ели как один из критериев оценки его перспективности/благонадежности (Касимов, 1960; Дерябин, Букштынов, 1970; Максимов, 1971; Алексеев, 1978). На основе наших результатов можно охарактеризовать наиболее перспективный в условиях пихто-ельника бореально-мелкотравного подрост ели (то есть способный к моменту достижения возраста генеративных деревьев вырасти до сопоставимой с ними высоты) как достигший высоты 2.50–3.00 м и развивший ZH5 в диапазоне 15–21 см · год–1. Проведенный анализ показал, что при таких значениях ZH5 у растений выражено значимое ускорение роста.
В литературе описано ускорение роста в высоту у подроста разных видов ели при улучшении освещенности (Науменко, 1962; Тимофеев, 1968; Максимов, 1971; Алексеев, 1978; Дерюгин и др., 2000; Prévost et al., 2016). Но высказывается точка зрения, что с некоторого критического возраста развитие подроста вступает в период необратимого старения, после чего ускорение его роста уже невозможно; такой возраст оценивается: для подзон хвойно-широколиственных лесов и южной тайги – 60–80 годами (Сеннов, 1999; Абатуров, Меланхолин, 2004), для подзоны средней тайги – 80 годами (Мелехов, 1966; Дыренков, 1966 – цит. по: Алексеев, 1978). Последняя оценка применима и к району нашего исследования; основываясь на ней, можно предполагать, что модели в возрасте до 80 лет, возможно, смогли бы ускорить линейный рост и нормально развиваться при улучшении условий в сообществе, но для моделей старше 80 лет это маловероятно.
Многомерная ординация моделей по комплексу морфометрических признаков (рис. 3) выявила их дифференциацию, прежде всего, по размерам: на ординационной диаграмме это отражается в виде расхождения групп вдоль вектора H, т.е. градиента общей высоты (с которой коррелируют диаметр ствола и диаметр кроны – см. табл. 2). Размерная дифференциация интерпретируется как различие онтогенетических состояний (рис. 3, табл. 3), поэтому закономерно, что с ней заметно согласована дифференциация по градиенту протяженности молодой (покрытой первичной корой) части ствола (вектор Lprim : H). Так, самые крупные растения с небольшой относительной протяженностью молодой части ствола отнесены к состоянию v1, самые мелкие (уже ветвящиеся) и при этом отличающиеся наибольшей относительной протяженностью молодой части ствола – к im1, растения промежуточного размера – к im2.
Рис. 3.
Дифференциация онтогенетических состояний и уровней жизненности у модельных растений ели на основе NMDS-ординации по сходству/различию морфометрических показателей. Для каждой ординационной оси в скобках указана соответствующая ей доля объясненной дисперсии в данных. Векторы, проведенные из начала координат, – градиенты морфометрических показателей (направлены в сторону возрастания значений; обозначения показателей – см. табл. 2). Символы – модельные растения (для характерных случаев, обсуждаемых в тексте, подписаны номера): 1 – с ускорением линейного роста; 2 – без значимых тенденций линейного роста, 3 – с замедлением линейного роста, 4 – с ускорением линейного роста и способные достичь размеров генеративных деревьев, 5 – без значимых тенденций линейного роста, но способные достичь размеров генеративных деревьев. Группы онтогенетических состояний (im1, im2, v1) разделены по уровням жизненности контурами: I – нормальная жизненность, II – пониженная, III – низкая.
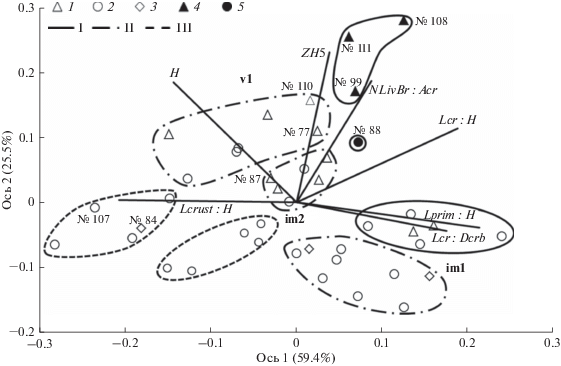
Таблица 3.
Морфометрические показатели онтогенетических состояний у подроста ели
| Онто-генетич. состояние | Уровень жизненности | n | H, м | D0, см | Dcrb, м | BrOrd | Lcr : Dcrb | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Me | min | max | Me | min | max | Me | min | max | Me | min | max | Me | min | max | |||
| im1 | Нормальная | 6 | 0.3 (α) | 0.2 | 0.4 | 0.4 (α) | 0.3 | 0.6 | 0.1 (α) | 0.1 | 0.2 | 3 (α) | 3 | 4 | 1.7 (α) | 1.3 | 2.3 |
| Пониженная | 9 | 0.3 (α) | 0.2 | 0.3 | 0.4 (α) | 0.1 | 0.5 | 0.1 (α) | 0.1 | 0.2 | 4 (α) | 3 | 4 | 2.0 (α) | 1.0 | 2.8 | |
| im2 | Нормальная | 1 | 1.4 | – | – | 2.3 | – | – | 0.8 | – | – | 5 | – | – | 1.3 | – | – |
| Пониженная | 6 | 1.4 (β) | 1.3 | 1.5 | 2.5 (β) | 2.1 | 3.0 | 1.0 (β) | 0.8 | 1.2 | 5 (β,γ) | 5 | 6 | 0.9 (β) | 0.7 | 1.2 | |
| Низкая | 5 | 1.4 (β) | 1.2 | 1.9 | 2.8 (β,γ) | 1.5 | 3.5 | 0.6 (γ) | 0.6 | 0.8 | 5 (β,δ) | 5 | 6 | 0.8 (β) | 0.7 | 1.3 | |
| v1 | Нормальная | 3 | 2.7 (γ) | 2.4 | 3.0 | 3.7 (γ) | 3.7 | 4.3 | 1.3 (β,δ) | 1.1 | 1.4 | 6 (β,γ,δ) | 5 | 6 | 1.8 (α,γ) | 1.4 | 2.5 |
| Пониженная | 7 | 2.7 (γ) | 2.2 | 2.9 | 4.7 (γ) | 3.6 | 6.3 | 1.7 (δ) | 1.3 | 1.9 | 7 (γ,δ) | 5 | 8 | 1.2 (β,γ) | 1.0 | 1.3 | |
| Низкая | 5 | 2.1 (γ,β) | 1.6 | 2.9 | 3.7 (β,γ) | 2.9 | 6.2 | 1.0 (β) | 0.8 | 1.1 | 6 (β,γ,δ) | 5 | 7 | 1.2 (β,γ) | 0.6 | 1.6 | |
Примечание. n – объем выборки. BrOrd – максимальный порядок ветвления, Dcrb – средний диаметр живой кроны внизу, D0 – диаметр ствола в основании, H – общая высота, Lcr : Dcrb – отношение протяженности живой кроны к ее среднему диаметру внизу. Me – медиана выборки (в скобках греческими буквами отмечены варианты значений: разными буквами обозначены варианты, имеющие значимые различия (тест Манна–Уитни, p < 0.05), одинаковыми буквами – варианты, не имеющие значимых различий). При n = 1 значимость различий неизвестна.
В группах онтогенетических состояний, особенно в более взрослых (im2 и v1), выражена согласованная дифференциация модельных растений по показателям ZH5, Lcr : H и NLivBr : Acr. Это можно интерпретировать как проявление разных уровней жизненности: в каждом состоянии к нормальному уровню жизненности относятся растения с наибольшим средним за 5 лет приростом в высоту, наибольшей относительной протяженностью живой кроны, наибольшим средним числом живых ветвей в годичной мутовке на стволе, но с наименьшей относительной протяженностью зоны корки на стволе. В группе v1 такое сочетание признаков характерно для растений, не только проявивших тенденцию ускорения роста в высоту, но и уже способных вырасти до размеров генеративных деревьев. В группе im2 только одно растение (№ 88) сильно отличается по значениям ZH5, Lcr : H и NLivBr : Acr. При этом оно было самым молодым из im2, но уже приобрело способность вырасти до размеров генеративных деревьев (рис. 2), хотя значимая тенденция ускорения роста у него еще не проявилась (резкое увеличение прироста произошло совсем незадолго до исследования).
Меньшие значения ZH5, Lcr : H, NLivBr : Acr и, наряду с этим, большее значение Lcrust : H отражают ухудшение жизненности. При этом в группе im1 мы выделили нормальный и пониженный уровни жизненности (не выделив низкий), поскольку ординация дифференцировала ее только на две подгруппы (рис. 3), а перспективы развития растений еще нельзя считать достаточно определенными в этом онтогенетическом состоянии. В группах im2 и v1 растения с уровнем жизни хуже нормального можно по морфометрическим показателям дифференцировать на пониженный и низкий уровни жизненности (табл. 4). В группе im2 морфометрическая дифференциация этих уровней выражена отчетливо (рис. 3). В группе v1 она менее отчетливая, но дополнительным обоснованием проведенной дифференциации служит то, что внешние признаки самого плохого жизненного состояния характерны для растений с календарным возрастом более 80 лет (табл. 4), для которых теоретически уже не предполагается способность развиваться сопоставимо с генеративными деревьями, даже при улучшении условий произрастания.
Таблица 4.
Морфометрические показатели жизненности и календарный возраст подроста ели
| Онтогенетич. состояние | Уровень жизненности | n | ZH5, см год–1 | NLivBr : Acr | Lcr : H | Lcrust : H | Lprim : H | A, годы | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Me | min | max | Me | min | max | Me | min | max | Me | min | max | Me | min | max | Me | min | max | |||
| im1 | Нормальная | 6 | 3.5 (α) | 2.5 | 5.0 | 1.8 (α) | 1.5 | 2.6 | 0.9 (α) | 0.8 | 0.9 | 0.0 (α) | 0.0 | 0.0 | 0.6 (α) | 0.4 | 0.9 | 9 (α) | 8 | 12 |
| Пониженная | 9 | 2.1 (β) | 1.6 | 2.4 | 1.2 (β) | 0.6 | 1.8 | 0.7 (β) | 0.5 | 0.8 | 0.0 (α) | 0.0 | 0.0 | 0.3 (β) | 0.2 | 0.7 | 12 (β) | 10 | 18 | |
| im2 | Нормальная | 1 | 12.1 | – | – | 3.5 | – | – | 0.8 | – | – | 0.0 | – | – | 0.36 | – | – | 23 | – | – |
| Пониженная | 6 | 5.3 (γ) | 3.9 | 5.7 | 3.3 (γ) | 2.4 | 4.2 | 0.7 (β,γ) | 0.6 | 0.8 | 0.0 (α,β) | 0.00 | 0.1 | 0.2 (γ,ε) | 0.1 | 0.3 | 36 (γ) | 33 | 42 | |
| Низкая | 5 | 2.8 (α,ζ) | 1.9 | 3.1 | 1.0 (β) | 0.7 | 1.3 | 0.5 (δ) | 0.3 | 0.6 | 0.1 (β) | 0.0 | 0.2 | 0.0 (δ) | 0.0 | 0.1 | 65 (δ) | 53 | 80 | |
| v1 | Нормальная | 3 | 18.2 (δ) | 15.0 | 20.6 | 4.2 (γ) | 3.1 | 5.0 | 0.9 (α) | 0.8 | 0.9 | 0.0 (β) | 0.0 | 0.0 | 0.3 (β,ε) | 0.2 | 0.4 | 38 (γ,ε) | 32 | 41 |
| Пониженная | 7 | 8.6 (ε) | 5.8 | 15.6 | 2.5 (γ) | 1.0 | 4.2 | 0.7 (γ) | 0.7 | 0.8 | 0.3 (β) | 0.00 | 0.7 | 0.2 (γ) | 0.0 | 0.2 | 63 (δ,ε) | 38 | 79 | |
| Низкая | 5 | 2.3 (β,ζ) | 1.9 | 3.1 | 0.8 (β) | 0.4 | 1.1 | 0.5 (δ) | 0.4 | 0.6 | 0.7 (γ) | 0.6 | 1.0 | 0.0 (δ) | 0.0 | 0.1 | 103 (ζ) | 81 | 125 | |
A – календарный возраст, Lcr : H – относительная протяженность живой кроны, Lcrust : H – относительная протяженность зоны корки на стволе, Lprim : H – относительная протяженность зоны первичной коры на стволе, NLivBr : Acr – среднее число живых ветвей в годичной мутовке на стволе, ZH5 – средний линейный прирост ствола за последние 5 лет. Остальные обозначения – см. табл. 3.
Описанные морфометрические различия выделенных групп связаны с их дифференциацией по темпам развития: в каждом онтогенетическом состоянии растения пониженной и низкой жизненности старше по календарному возрасту, чем растения нормальной жизненности (табл. 4). Среди старших имматурных (im2) единственное растение, жизненность которого была оценена как нормальная, было самым молодым, и у него еще не успела проявиться значимая тенденция ускорения роста, но оно уже развило такую скорость роста в высоту, что было способно вырасти так же, как генеративные деревья в том же сообществе. Растения im2 пониженной жизненности имели такой же календарный возраст, как v1 нормальной жизненности, но значимо отличались от последних по скорости роста и размерам, еще не приобрели способность вырасти до размеров генеративных деревьев к моменту достижения сопоставимого с ними возраста. Поэтому и было сделано заключение, что ход развития таких растений не вполне нормален.
Ход развития подроста хвойных видов и его жизненность часто характеризуют такими количественными признаками как длина годичного линейного прироста верхушки ствола (обычно – среднее значение за несколько последних лет) и ее соотношение с длиной годичных линейных приростов у ветвей 2-го порядка (за последний год или за несколько последних лет), или же заменяют второй из этих признаков характеристикой общей формы живой кроны, под которой понимается отношение ее длины к максимальному диаметру (в нижней части) (Касимов, 1960; Титов, 1970; Алексеев, 1978; Романовский, 2001; Claveau et al., 2002; Ставрова и др., 2017). Проведенный нами анализ показал, что признак ZH5 четко дифференцирует у подроста разные уровни жизненности (табл. 4), но использоваться в диагностике онтогенетических состояний может только в случаях нормальной и пониженной жизненности. К тому же абсолютные значения этого признака зависят от почвенно-гидрологических условий (Чиндяев, 1986), поэтому для его применения в диагностике требуется систематизированное исследование его вариабельности в разных типах леса. Соотношение между длиной линейного прироста на верхушке ствола и ветвей 2-го порядка, если его рассматривать не за последний год, а как среднее за несколько лет (что более корректно), – не очень удобный признак для применения в ходе массовых учетов подроста. Что касается общей формы кроны, которая, по мнению ряда исследователей (Claveau et al., 2002; Ставрова и др., 2017), отражает это соотношение именно как среднемноголетнее, проведенный нами анализ не выявил достаточно четких различий между онтогенетическими состояниями и уровнями жизненности по признаку Lcr : Dcrb (табл. 3), характеризующему форму кроны.
Относительную протяженность кроны и мутовчатость ветвления ствола часто учитывают в диагностике жизненности (Касимов, 1960; Дерябин, Букштынов, 1970; Злобин, 1970; Максимов, 1971; Claveau et al., 2002; Правила …, 2020). Комбинация этих показателей наиболее четко дифференцировала исследованные нами модели по уровням жизненности во всех онтогенетических состояниях (табл. 4, рис. 3).
По итогам проведенного анализа предлагается следующий ключ для диагностики у подроста ели сибирской онтогенетических состояний im1, im2 и v1, а также уровней жизненности в этих состояниях.
1. Ветвящееся растение высотой не более 0.5 м, порядок ветвления – не более 4-го………………………… …………………………………………….. im1; см. ступень 2
– Высота растения более 0.5 м (до 3–4 м) ……… ……….................…………… im2 или v1; см. ступень 3
2. Относительная протяженность живой кроны составляет 3/4 и более от общей высоты растения; в кроне на всех годичных приростах стволика развиты ветви 2-го порядка и обычно их не менее 2-х на приросте …...................................... ......……… im1 нормальной жизненности (рис. 4а)
Рис. 4.
Схематические изображения внешнего облика ели сибирской в онтогенетических состояниях im1 (а, б), im2 (в–д) и v1 (е–з). Уровни жизненности: а, в, е – нормальная, б, г, ж – пониженная, д, з – низкая.
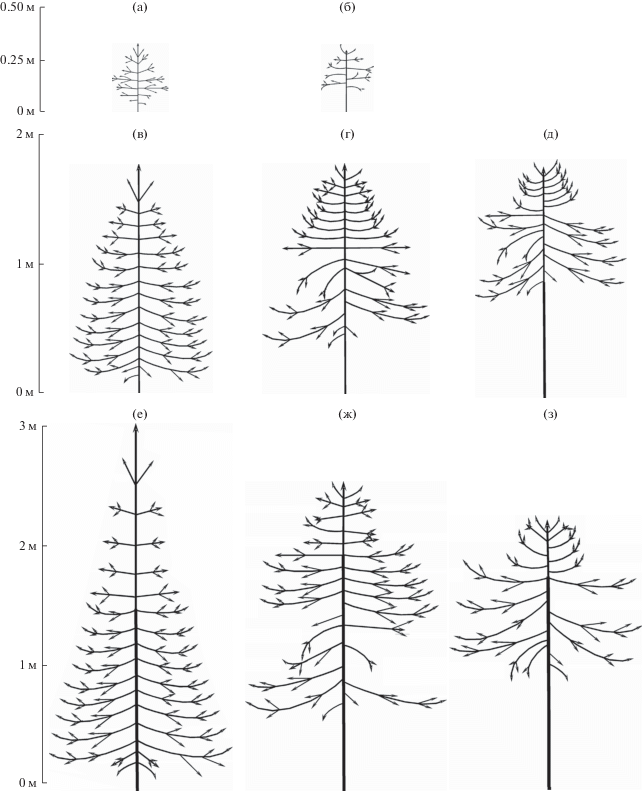
– Относительная протяженность живой кроны не превышает 3/4 от общей высоты растения; на годичных приростах стволика часто бывает развито по 1 ветви 2-го порядка, а у некоторых растений ветви 2-го порядка могут быть развиты не на всех годичных приростах ……………………………………… …………… im1 пониженной жизненности (рис. 4б)
3. Протяженность живой кроны составляет не менее 4/5 от общей высоты растения; в кроне на всех годичных приростах ствола развиты мутовки ветвей 2-го порядка и каждая из них включает более 2 ветвей; корка на стволе отсутствует ……… …………………...............…………………… im2 или v1 нормальной жизненности; см. ступень 4
– Протяженность живой кроны – менее 4/5 от общей высоты; в кроне на некоторых годичных приростах ствола может быть развито по 1–2 ветви 2-го порядка или могут совсем отсутствовать боковые ветви; в нижней части ствола нередко развита корка...........… пониженная или низкая жизненность; см. ступень 5
4. Максимальный порядок ветвления (на самых старых живых ветвях в нижней части кроны) – не более 5-го………………………………………… im2 нормальной жизненности (рис. 4в)
– В нижней части кроны максимальный порядок ветвления – обычно 6-й (у некоторых растений – 5-й, но в таких случаях протяженность зоны первичной коры – не более 1/5 от общей высоты) …………………v1 нормальной жизненности (рис. 4е)
5. В кроне на каждом годичном приросте есть хотя бы 1 ветвь 2-го порядка; корка в нижней части ствола обычно отсутствует (а если есть, то ее протяженность не превышает 1/10 от общей высоты); протяженность живой кроны составляет не менее 1/2 от общей высоты (часто – не менее 2/3) ………………………………… пониженная жизненность; см. ступень 6
– В кроне на некоторых годичных приростах ствола могут отсутствовать ветви 2-го порядка; в нижней части ствола обычно развита корка; протяженность зоны первичной коры составляет не более 1/10 от общей высоты; протяженность живой кроны часто составляет менее 1/2 от общей высоты ……………..…..……………… низкая жизненность; см. ступень 7
6. Максимальный порядок ветвления – не больше 6-го; протяженность зоны первичной коры составляет до 1/3 от общей высоты, а протяженность зоны корки – не более 1/10 ………………… …………………………………… im2 пониженной жизненности (рис. 4г)
– Максимальный порядок ветвления – 6-й или более высокий; протяженность зоны первичной коры составляет не более 1/5 от общей высоты, а зоны корки – до 2/3 (иногда – не более 1/10, но в этом случае общая высота растения превышает 2 м); ……………………………..……………… v1 пониженной жизненности (рис. 4ж)
7. Протяженность зоны корки в нижней части ствола составляет не более 1/10 от общей высоты; максимальный порядок ветвления – не больше 6-го …………………………………………………… im2 низкой жизненности (рис. 4д)
– Протяженность зоны корки – не менее 1/2 от общей высоты, иногда ствол покрыт коркой до самого верха; максимальный порядок ветвления – 6-й или более высокий ……..............………………………… ……………………… v1 низкой жизненности (рис. 4з)
Заключение. Проведенный анализ показал, что в темнохвойных таежных лесах растения ели сибирской в начальных онтогенетических состояниях, относящиеся к категории “здоровые” (Алексеев, 1989) или “жизнеспособные” (Правила …, 2020), сильно дифференцированы по темпам своего развития; это проявляется в их значимых различиях по календарному возрасту и морфометрическим показателям габитуса. Такая поливариантность развития отражается на способности подроста ели перейти в генеративное состояние и на общем ходе возобновительных процессов в древостое, поэтому ее необходимо учитывать при проведении исследований динамики популяций ели и темнохвойных лесов, при планировании лесохозяйственной деятельности.
Проявление поливариантности развития у подроста ели сибирской, то есть его дифференциацию по уровням жизненности в онтогенетических состояниях im1, im2, v1, наиболее четко отражают значения таких показателей как среднее число ветвей в годичных мутовках на стволе и относительная протяженность живой кроны. Относительная протяженность зон ствола, покрытых первичной корой и коркой, – это показатели как биологического возраста подроста, так и его жизненности; их применение в диагностике онтогенетических состояний возможно, если одновременно учитываются признаки морфологии кроны, позволяющие корректно диагностировать жизненность. Дифференциация онтогенетических состояний также проявляется в показателях общего размера (высоте, диаметре ствола, диаметре живой кроны внизу) и структурной сложности (максимальный порядок ветвления). Длины годичных линейных приростов ствола тоже очень тесно связаны с уровнем жизненности. Поэтому их среднее значение за последние 5 лет (ZH5) можно использовать для оценки перспектив растений в сообществе (особенно у im2 и v1), но в диагностике онтогенетических состояний – только в случае нормальной жизненности (проявившейся в параметрах кроны – см. выше) и только как дополнительный признак. Для уточнения диагностических критериев размера и линейного прироста ствола у онтогенетических состояний необходим сравнительный количественный анализ варьирования этих признаков подроста в условиях разных типов леса.
По критерию темпов роста в высоту можно выделить у ели сибирской несколько категорий перспективности (благонадежности) подроста. Высокую перспективность имеет подрост в онтогенетических состояниях im2 и v1 с нормальной жизненностью. Внешнее строение таких растений характеризуется относительной протяженностью живой кроны не менее 4/5 от общей высоты и числом живых ветвей более трех во всех годичных мутовках на стволе; благополучие перспектив обеспечивается тем, что у них выражена тенденция увеличения годичных линейных приростов ствола, в частности – ZH5 уже составляет более 10 см год–1. Среднюю перспективность имеет подрост в онтогенетических состояниях im2 и v1 с пониженной жизненностью, если у него выражена тенденция увеличения годичных линейных приростов; внешним признаком такого состояния растений может служить относительная протяженность живой кроны от 2/3 до 4/5 от общей высоты. Перспективность следует считать сомнительной у подроста в онтогенетических состояниях im2 и v1 при низкой жизненности (ZH5 менее 4 см), а также если в этих состояниях при пониженной жизненности не выражена тенденция ускорения линейного роста; у таких растений относительная протяженность живой кроны составляет менее 2/3 общей высоты, на многих годичных линейных приростах ствола развито только по одной живой ветви или ветви совсем отсутствуют. У подроста в онтогенетическом состоянии im1 перспективы еще не определенные.
Список литературы
Абатуров А.В., Меланхолин П.Н. Естественная динамика леса на постоянных пробных площадях в Подмосковье. Тула: Гриф и К°, 2004. 333 с.
Абатуров А.В., Кулешов А.П., Браславская Т.Ю. Оценка успешности возобновления ели под пологом лиственных лесов в национальном парке “Лосиный остров” // Известия Самарского НЦ РАН. 2012. Т. 14. № 1(8). С. 1941–1945.
Алексеев В.А. Диагностика жизненного состояния деревьев и древостоев // Лесоведение. 1989. № 4. С. 51–57.
Алексеев В.И. Возобновление ели на вырубках. М.: Наука, 1978. 130 с.
Атлас Республики Коми / Под ред. Коми НЦ Уральского отд-ния РАН. М.: Дизайн. Информация. Картография, 2001. 551 с.
Бобкова К.С., Бессонов И.М. Естественное возобновление в среднетаежных ельниках Европейского северо-востока // Лесоведение. 2009. № 5. С. 10–16.
Бобкова К.С., Робакидзе Е.А., Галенко Э.П. Жизненное состояние древостоев и подроста коренных ельников предгорий Урала бассейна реки Печоры // Сибирский экологический журн. 2010. № 2. С. 271–280.
Бобрецов А.В., Теплова В.П. Природные условия Печоро-Илычского заповедника // Закономерности полувековой динамики биоты девственной тайги Северного Предуралья. Сыктывкар: Госкомстат Республики Коми, 2000. С. 6–21.
ГОСТ 18486-87. Государственный стандарт Союза ССР. Лесоводство. Термины и определения. Утвержден и введен в действие Постановлением Государственного комитета СССР по стандартам от 10.12.1987 № 4445. URL: http://docs.cntd.ru/document/1200022985 (дата обращения – 02.05.2021).
Давыдычев А.Н., Кулагин А.Ю. Характеристика предгенеративного периода онтогенеза ели сибирской (Picea obovata Ledeb.) в подзоне широколиственно-хвойных лесов Уфимского плато // Бюллетень Моск. общества испытателей природы. Отд. биол. 2010. Т. 115. Вып. 2. С. 59–66.
Декатов Н.Н., Кузнецов А.Н. Рост древостоев, возникших из подроста // Лесное хоз-во. 1968. № 8. С. 46–48.
Дерюгин А.А., Рубцов М.В., Серяков А.Д. Рост ели после рубки березняков с сохранением подроста в южной тайге // Там же. 2000. № 5. С. 30–31.
Дерябин Д.И., Букштынов А.Д. Лесоводственное значение хвойного подроста. М.: Лесная пром-сть. 1970. 96 с.
Диагнозы и ключи возрастных состояний лесных растений. Деревья и кустарники. М.: Прометей,1989. 102 с.
Евстигнеев О.И. Поливариантность сосны обыкновенной в Брянском полесье // Лесоведение. 2014. № 2. С. 69–77.
Злобин Ю.А. Оценка качества подроста древесных растений // Там же. 1970. № 3. С. 96–102.
Злобин Ю.А. Факторный анализ параметров морфогенеза подроста древесных пород // Известия вузов. Лесной журн. 1975. № 1. С. 19–21.
Злобин Ю.А. Принципы и методы изучения ценотических популяций растений. Учебно-методическое пособие. Казань: Изд-во Казанского гос. университета, 1989. 146 с.
Касимов В.Д. Классификация елового подроста // Лесное хоз-во. 1960. №7. С. 9–11.
Кузьмичев В.В. Закономерности роста древостоев. Новосибирск: Наука, 1977. 160 с.
Максимов В.Е. Физиологические особенности и рост елового подроста на лесосеках постепенных и сплошных рубок // Лесоведение. 1971. № 1. С. 84–88.
Методы изучения лесных сообществ. СПб.: НИИХимии СПбГУ, 2002. 240 с.
Минина Е.Г. Определение пола у лесных древесных растений // Труды Института леса АН СССР. 1960. Т. 47. С. 76–163.
Мишко А.Е. Онтоморфогенез ели сибирской (Picea obovata Ledeb.) в северотаежных лесах (на примере Кольского полуострова): Дис. … канд. биол. наук (специальность 03.02.08). СПб.: Ботанический ин-т им. В.Л. Комарова РАН, 2019. 146 с.
Науменко З.М. Жизнеспособность елового подроста на сплошных вырубках Ленинградской области // Сборник работ по лесному хозяйству. М.: Гослесбумиздат, 1962. Вып. 5. С. 158–182.
Некрасова Т.П., Рябинков А.П. Плодоношение пихты сибирской. Новосибирск: Наука, 1978. 151 с.
ОСТ 56-108-98. Стандарт отрасли. Лесоводство. Термины и определения. Утвержден и введен в действие Приказом Рослесхоза от 03.12.1998 № 203. URL: https://klh.mosreg.ru/dokumenty/spravochnaya-literatura/ gosty-i-osty/ost-56-108-98-lesovodstvo-terminy-i-opredeleniya (дата обращения – 02.05.2021)
Правила лесовосстановления. Утверждены приказом Министерства природных ресурсов Российской Федерации № 1014 от 04.12.2020. Зарегистрированы в Министерстве юстиции Российской Федерации: № 61556 от 18.12.2020. URL: http://www.consultant.ru/document/ cons_doc_LAW_370561/42d0d93b6e5c84eb17e29477246f258ffa5566b7/ (дата обращения – 02.05.2021).
Романовский А.М. Поливариантность онтогенеза Picea abies (Pinaceae) в Брянском полесье // Ботанический журн. 2001. Т. 86. № 8. С. 72–85.
Семиколенных А.А., Бовкунов А.Д., Алейников А.А. Почвы и почвенный покров таежного пояса Северного Урала (верховья реки Печора) // Почвоведение. 2013. № 8. С. 911–923.
Сеннов С.Н. Итоги 60-летних наблюдений за динамикой леса. СПб.: СПб. НИИ лесного хоз-ва, 1999. 98 с.
Серебряков И.Г. Экологическая морфология растений. М.: Высшая школа, 1962. 378 с.
Смирнов Н.С., Браславская Т.Ю. Растительность темнохвойных лесов нижней части бассейна реки Большая Порожняя (приток р. Печора) // Тр. Печоро-Илычского гос. заповедника. Сыктывкар: Изд-во Коми НЦ УрО РАН, 2010. Вып. 16. С. 149–156.
Смирнова О.В., Бобровский М.В., Ханина Л.Г., Смирнов В.Э. Биоразнообразие и сукцессионный статус темнохвойных лесов Шежимопечорского и Большепорожного ботанико-географических районов Печоро-Илычского заповедника // Труды Печоро-Илычского гос. заповедника. Сыктывкар: Институт геологии Коми НЦ Уральского отд-ния РАН, 2007. Т. 15. С. 28-47.
Смолоногов Е.П. Комплексное районирование Урала // Леса Урала и хозяйство в них. Екатеринбург: Уральская гос. лесотехническая академия, 1995. Вып. 18. С. 24–41.
Ставрова Н.И., Горшков В.В., Мишко А.Е. Онтогенез ели сибирской Picea obovata Ledeb. (Pinaceae) в малонарушенных северотаежных кустарничково-зеленомошных сосново-еловых лесах // Ботанический журн. 2017. Т. 102. № 2. С. 163–185.
Тимофеев В.П. Роль елового подроста в восстановлении вырубок // Лесное хоз-во. 1968. № 12. С. 9–15.
Титов Н.А. О закономерных связях морфологических признаков елового подроста // Известия вузов. Лесной журн. 1970. № 6. С. 30–33.
Трулль О.А. Математическая статистика в лесном хозяйстве. Минск: Высшая школа, 1966. 233 с.
Чиндяев А.С. Влияние осушения на прирост по высоте у подроста ели предварительной генерации в условиях Среднего Урала // Экология и физиология основных лесообразующих видов Урала. Свердловск: Уральский НЦ АН СССР, 1986. С. 77–86.
Энциклопедия лесного хозяйства: в 2-х томах. Т. 2. М.: Всеросс. НИИ лесного хоз-ва и механизации, 2006. С. 145.
Antos J.A., McPherson A., Guest H. Growth rates and crown morphology of Abies amabilis in the seedling bank of an ancient, subalpine conifer forest // Canadian J. Forest Research. 2020. V. 50. № 11. P. 1124–1130.
Antos J.A., Parish R., Conley K. Age structure and growth of the tree-seedling bank in subalpine spruce-fir forests of south-central British Columbia // American Middle Naturalist. 2000. V. 143. № 2. P. 342–354.
Claveau Y., Messier C., Comeau P.G., Coates K.D. Growth and crown morphological responses of boreal conifer seedlings and saplings with contrasting shade tolerance to a gradient of light and height // Canadian J. Forest Research. 2002. V. 32. № 3. P. 458–468.
Evstigneev O.I., Korotkov V.N. Ontogenetic stages of trees: an overview // Russian J. Ecosystem Ecology. 2016. V. 2. № 2. P. 1–31.
Gatzuk L.E., Smirnova O.V., Vorontzova L.I., Zaugolnova L.B., Zhukova L.A. Age states of plants of various growth forms: a review // J. Ecology. 1980. V. 68. № 3. P. 675–696.
Hammer Ø., Harper D.A.T., Ryan P.D. Past: Paleontological Statistics software package for education and data analysis // Palaeontologia Electronica. 2001. V. 4. № 1. P. 1–9. URL: http://palaeo-electronica.org/2001_1/past/issue1_01.htm
Omelko A., Ukhvatkina O., Zhmerenetsky A., Sibirina L., Petrenko T., Bobrovsky M. From young to adult trees: How spatial patterns of plants with different life strategies change during age development in an old-growth Korean pine-broadleaved forest // Forest Ecology & Management. 2018. V. 411. № 1. P. 46–66. https://doi.org/10.1016/j.foreco.2018.01.023
Prévost M., Dumais D., De Blois J. Morphological response of conifer advance growth to canopy opening in mixedwood stands, in Quebec, Canada // Trees. 2016. V. 30. № 5. P. 1735–1747.
Дополнительные материалы отсутствуют.