

Лесоведение, 2021, № 6, стр. 566-580
Функциональная классификация лесов: актуальность и подходы к разработке
Н. В. Лукина a, *, А. П. Гераськина a, А. И. Кузнецова a, В. Э. Смирнов a, А. В. Горнов a, Н. Е. Шевченко a, Е. В. Тихонова a, Д. Н. Тебенькова a, Е. В. Басова a
a Центр по проблемам экологии и продуктивности лесов РАН
117997 Москва, ул. Профсоюзная, 84/32, стр. 14, Россия
* E-mail: nvl07@yandex.ru
Поступила в редакцию 30.06.2021
После доработки 13.07.2021
Принята к публикации 21.07.2021
Аннотация
В статье обосновывается актуальность разработки функциональной классификации лесов на основе эффективности выполнения ими экосистемных функций. На примере девяти типов хвойно-широколиственных лесов, доминирующих в европейской части России и функционирующих в автономных позициях ландшафта, демонстрируется возможный подход к оценке выполнения ими функции регулирования цикла углерода на основе взаимосвязей между информативными индикаторами состава растительности, почвенной макрофауны и экосистемных процессов, связанных с секвестрацией почвенного углерода. Самым низким уровнем аккумуляции почвенного углерода характеризуются хвойно-широколиственные леса на песчаных почвах, отличающиеся небольшим функциональным разнообразием и низкой биомассой дождевых червей. Высокий запас почвенного углерода демонстрируют леса с более равномерным распределением в составе растительности функциональных групп растений с быстро- и медленноразлагаемым опадом. Это объясняется созданием трофических и топических условий, благоприятных для функционирования дождевых червей – активных переработчиков опада в хвойно-широколиственных лесах. Широкомасштабная оценка взаимосвязей между растительностью, почвенной биотой и экосистемными процессами, формирующими функцию регулирования цикла углерода лесами России, является актуальной научной задачей в связи с проблемами изменения климата. На основе оценки эффективности выполнения различных экосистемных функций лесами разных типов, идентифицированных в России с использованием классификаций растительности, может быть создана функциональная классификация лесов.
Развитие классификации типов лесов, которая обладает прогностическим потенциалом, является важнейшей задачей лесоведения. Существующие подходы к типизации лесов основываются на доминирующих (Сукачев, Дылис, 1964) и/или индикаторных (Каяндер, 1933) видах растений, характеризующих условия произрастания лесов, а также на типах условий местопроизрастания (Алексеев, 1928; Погребняк, 1928, 1955). Для лесов Европейской России разработана унифицированная классификации растительности, которая базируется на доминантном подходе, дополненном эколого-ценотическим и флористическим анализом (Заугольнова, 2008; Браславская, Заугольнова, 2010). Л.Б. Заугольновой и В.Б. Мартыненко создан электронный Определитель типов леса Европейской России (http://cepl.rssi.ru/bio/forest/index.htm). Определитель позволяет охарактеризовать типологическое разнообразие лесов на основе традиций отечественной лесной фитоценологии и зарубежных подходов к классификации растительности. В зоне хвойно-широколиственных лесов европейской части России по материалам Определителя типов леса Европейской (http://cepl. rs-si.ru/bio/forest/index.htm) России можно выделить около 160 типов леса.
В соответствии с российской классификацией почв (Классификация, 2004) в лесах выделяют более 50 типов почв, из которых не менее 15 типов формируются в зоне хвойно-широколиственных лесов. Свойства почв и почвенные процессы, на основе которых проводится классификация почв, важны для реализации экосистемных функций лесов. Однако соответствие между типами леса и типами почв установить сложно, поскольку не только один и тот же тип почв идентифицируется в разных типах леса, но и в одном и том же типе леса идентифицируются почвы разных типов.
Между тем, оценка функциональных взаимосвязей между растительностью и почвой имеет важнейшее научное и практическое значение. В последние годы активно развивается концепция мультифункциональности лесов (Manning et al., 2015; Тебенькова и др., 2019), согласно которой леса выполняют одновременно все четыре категории экосистемных функций/услуг: обеспечивающие (древесина, волокна), регулирующие (регулирование климата, гидрологического режима), поддерживающие (сохранение и формирование местообитаний, естественного плодородия почв) и культурные (рекреация, оздоровление и др.) (Millennium …, 2005).
В связи с высокой актуальностью оценки экосистемных функций и услуг лесов и взаимосвязей между ними возникает необходимость разработки функциональной классификации лесных экосистем. Подходы к выделению функциональных групп/типов растений (Смирнова и др., 2004; Cornelissen et al., 2007; Salemaa et al., 2008) и почвенной биоты (Lavelle et al., 2006) развиваются, однако задача создания функциональной классификации лесов пока не решается. Хотя все леса мультифункциональны, однако полнота реализации той или иной функции в разных типах леса различается. На основе оценки выполнения функций лесными биогеоценозами разных типов, идентифицированных в России с использованием классификаций растительности, может быть создана функциональная классификация лесов.
Цель данной статьи – обсудить актуальность функциональной классификации лесов и предложить возможные подходы к ее разработке на примере функции регулирования цикла углерода на основе оценки взаимосвязей между растительностью, почвенными сапрофагами и индикаторами экосистемных процессов, связанных с секвестрацией углерода в почвах.
Функциональная классификация лесов должна отражать эффективность выполнения лесами, формирующимися в различных условиях комбинированного влияния природных и антропогенных факторов, экосистемных функций. Таким образом, ключевым моментом создания функциональной классификации является оценка эффективности выполнения различных функций. Это позволит учитывать взаимосвязи, то есть синергию и конфликты (компромиссы) между экосистемными функциями/услугами, без чего невозможно устойчивое управление лесами.
В данной работе мы остановимся только на возможных подходах к оценке эффективности выполнения лесами функции регулирования цикла углерода. Такая оценка может быть построена на основе интеграции информации о специфических чертах/признаках/функциональных характеристиках лесной биоты разных трофических уровней, участвующей в экосистемных процессах, взаимодействие которых формирует данную функцию.
Поскольку эффективность реализации экосистемных функций зависит как от биотических факторов и экосистемных процессов, так и от абиотических и антропогенных факторов, важно учитывать их комбинированное влияние. Остановимся вначале кратко на абиотических факторах, оказывающих влияние на цикл углерода, прежде всего, на таких как почвообразующие породы, климат, рельеф. Эффективность выполнения функции регулирования цикла углерода является результатом взаимодействия лесной биоты с комплексом этих природных абиотических факторов. Идентифицируют 3 генерализованных и 9 детализированных разностей почвообразующих пород по гранулометрическому составу: пески (рыхлый, связный, супесь), суглинок (легкий, средний, тяжелый), глина (легкая, средняя, тяжелая) (Розанов, 2004). Для разработки функциональной классификации лесов в отношении функции регулирования цикла углерода, важно учитывать и варьирование химического и минералогического состава почвообразующих пород.
Для учета влияния климатических условий целесообразно использовать существующий подход к зонированию растительности лесного пояса России: северная, средняя, южная тайга и хвойно-широколиственные, широколиственные леса. Выделяются также три позиции в ландшафтах, на которых развиваются леса: автономная, транзитная и аккумулятивная. Если учитывать 3 разности по гранулометрическому составу, 4 зоны растительности лесного пояса и 3 разности по положению лесов в ландшафте, сочетания этих трех абиотических факторов дает 36 комбинированных разностей условий, важных для реализации экосистемных функции регулирования цикла углерода. На функции лесов, связанные с биогеохимическими циклами, влияют и другие абиотические факторы, такие как дренирование почв, обусловленное как их структурой, так и степенью водопроницаемости подпочвенного слоя и уровнем грунтовых вод, определяемым расположением водоносного слоя.
К антропогенным факторам, влияющим на эффективность выполнения экосистемных функций лесов, относятся режимы лесопользования и история землепользования в целом, техногенный фактор. Важнейшим и постоянно действующим фактором динамики лесов являются пожары, которые часто имеют антропогенное происхождение; необходим учет влияния прошлых (Бобровский, 2010) и современных пожаров (Барталев, Стыценко, 2021).
Таким образом, для оценки эффективности выполнения лесами функции регулирования цикла углерода необходимо учитывать весь этот комплекс абиотических факторов. Поскольку анализ данных факторов существует в многочисленных научных трудах и отражен в различных базах данных, необходима их интеграция с использованием современных подходов и методов, развитием информационных систем на разных пространственных уровнях.
Перейдем к биотическим факторам и экосистемным процессам. Все экосистемные функции обеспечиваются биотой через формирование ею экосистемных процессов. Функция регулирования цикла углерода формируется через взаимодействие таких экосистемных процессов, как поглощение соединений углерода растительностью и почвой и его аккумуляция в этих пулах, разложение, минерализация органического вещества, эмиссия CO2 в атмосферу, вынос соединений углерода из лесных экосистем с почвенными водами. Естественно, что все представители биоты без исключения участвуют в формировании этой функции, но для решения практической задачи создания функциональной классификации лесов важно ответить на вопрос: можно ли выделить наиболее информативные, относительно несложно измеряемые индикаторы, отражающие участие представителей биоты разных трофических уровней в регулировании экосистемных процессов, формирующих цикл углерода?
Для классификации лесов по эффективности выполнения ими функции регулирования цикла углерода, прежде всего, необходимо предложить информативные специфические черты/характеристики/индикаторы представителей биоты, как продуцирующей органическое вещество (растительность), так и участвующей в его разложении и накоплении, фиксации (стабилизации) в почвах (почвенная биота).
Известно, что для оценки эффективности процессов продуцирования органического вещества растительностью необходимо оценивать такие показатели как фитомасса и мортмасса, размеры годичной продукции и годичного опада отдельных видов, групп растений и фитоценоза в целом (Уткин, 1975; Пулы и потоки …, 2007). Густота древостоя, строение/структура крон древесных растений и полога в целом регулируют количество не только проникающего под полог солнечного света, определяющего развитие нижних ярусов растительности, но и проникающих под полог атмосферных осадков и, соответственно, влияют на интенсивность выноса соединений углерода из биогеоценоза (Лукина и др., 2018), и поэтому тоже влияют на биогеохимические циклы.
Следует подчеркнуть, что все эти показатели зависят от возраста растений и сообществ, поэтому при оценке выполнения функций лесами разных типов необходимо учитывать возрастную динамику видов растений, их групп и сообществ. Для оценки и прогнозов не менее важно учитывать сукцессионный статус/стадии сукцессии лесов (Аккумуляция …, 2018): почвы хранят органический углерод, который формируется на протяжении смен поколений лесных сообществ. Для оценки бюджета углерода важнейшей задачей является измерение внутригодичной, сезонной и суточной динамики экосистемных процессов, таких как разложение опада и эмиссия парниковых газов (Курганова, 2010), поступление соединений углерода с атмосферными выпадениями и их вынос с почвенными водами (см. http://icp-forests.net), обмен парниковыми газами между лесными биогеоценозами и атмосферой (Kurbatova et al., 2013). Все эти индикаторы экосистемных процессов, отражающие как многолетнюю, так и годичную, сезонную и суточную динамику соединений углерода, важны и должны оцениваться для развития функциональной классификации лесов. Существуют уже накопленные данные, полученные на основе как научного, так и ведомственного мониторинга, и необходима их постоянная актуализация. Однако при первом приближении к решению непростой задачи построения функциональной классификации лесов необходимо предложить относительно несложные подходы, позволяющие оценивать относительную эффективность выполнения лесными биогеоценозами разных типов/стадий сукцессий функции регулирования циклов углерода. Такие подходы могут развиваться на основе оценки взаимосвязей между представителями биоты разных трофических уровней и индикаторами экосистемных процессов, связанных с секвестрацией углерода в почвах, где депонируется значительная часть экосистемного углерода (Framstad et al., 2013).
Звеном, связывающим растительность, почву и почвенную биоту, является опад. Качество опада оказывает влияние на скорость его разложения (Berg, McClaugherty, 2020) и, соответственно, на биогеохимические циклы. Характеристиками качества опада являются содержания в нем элементов питания (азот, фосфор, калий, кальций, магний и др.) и вторичных метаболитов (полифенолы, лигнины, др.), стехиометрические соотношения C : N, лигнин : N и др. (Cadish, Giller, 1997; Perez-Harguindeguy, 2000; Berg, McClaugherty, 2020). Скорость разложения опада высокого качества, отличающегося высоким содержанием азота и других элементов питания, узким соотношением C : N и лигнин : N, выше, чем опада, содержащего меньше элементов питания и больше вторичных метаболитов. Качество опада обусловливает содержание доступных для биоты питательных веществ в почве и регулирует скорость разложения растительных остатков – основного источника питания сапрофагов (Krishna, 2017).
К почвенным сапрофагам относятся микроорганизмы и беспозвоночные животные, которые являются переработчиками опада. Если разложение опада осуществляется только микроорганизмами и микрофауной, эмиссия углекислого газа довольно интенсивна, что приводит к потерям почвенного углерода. Снижает эти потери почвенная макрофауна, в особенности такие макросапрофаги, как дождевые черви, которые способствуют закреплению углерода в почвах, как за счет обеспечения процессов гумусообразования, так и за счет переноса органического вещества почвы в более глубокие горизонты в результате роющей деятельности (Frouz et al., 2013). В то же время дождевые черви за счет ферментов кишечного тракта активизируют почвенные микроорганизмы (Spurgeon et al., 2013; Kurakov et al., 2016; Cao et al., 2018).
Дождевые черви и другие почвенные беспозвоночные широко используются для зоологической диагностики почв (Гиляров, 1975; Mordokovich, 2014). Критерии для индикаторных групп выбираются в соответствии с целями и задачами исследования, например, для индикации техногенных нарушений и мониторинга природных местообитаний выбираются разные группы почвенных беспозвоночных в связи с разным составом почвенной фауны на исследуемых территориях. В целом, выбор определенной группы или вида беспозвоночных в качестве индикатора почвенных условий должен основываться на его доминировании в естественных местообитаниях (Бабенко, 2013). Выбор именно почвенной макрофауны из многочисленных представителей почвенной фауны для оценки экосистемных процессов, связанных с регулированием цикла углерода в лесах, можно обосновать тем, что представители почвенной макрофауны являются истинными педобионтами, влияют на другую почвенную биоту и на формирование почв и отражают происходящие в ней изменения. Трофическая группа макросапрофагов представляет особый интерес как индикаторная, поскольку, с одной стороны, она отражает основные физико-химические свойства почв (гранулометрический состав, влажность, кислотность) и чувствительна к качеству опада растительности (Staaf, 1987; Huang et al., 2020), с другой стороны, сама оказывает влияние на содержание С, N, Ca, отношение C : N, кислотность и другие почвенные характеристики, т.е. выступает и как отклик на комбинированное влияние абиотических факторов, почвенных свойств и растений, и может выступать как предиктор почвенных изменений. Среди почвенных макросапрофагов в лесных экосистемах европейской части России часто доминируют дождевые черви, которые составляют до 90% биомассы почвенных беспозвоночных (Чернов, 1975). В подстилке численность моллюсков, двупарноногих многоножек, личинок двукрылых может быть высокой, но их вклад в биомассу значительно ниже, чем дождевых червей.
Дождевые черви, в первую очередь, весьма чувствительны к гранулометрическому составу почв. Для дождевых червей более благоприятны легкие суглинистые и среднесуглинистые почвы, в почвах тяжелого гранулометрического состава, испытывающих затопление (тяжелые суглинки), а также легкого гранулометрического состава (пески), которые часто пересыхают, они могут отсутствовать (Перель, 1979; Холхоева, 1993; Zhang et al., 2007). Значимым фактором для успешного функционирования червей является качество опада, от которого зависят биомасса и состав морфо-экологических групп дождевых червей. Дождевые черви предпочитают быстро разлагаемый лиственный опад (липа, клен, ясень, береза) трудно разлагаемому лиственному (дуб, осина, бук) или хвойному (ель, пихта, сосна) (Перель, 1979; Schelfhout et al., 2017). Показано, что при схожем составе древостоя видовое разнообразие и биомасса дождевых червей во многом определяются ярусом кустарников и напочвенным покровом. Например, в буковых или темнохвойных лесах, древостой которых формирует трудноразлагаемый опад, разными группами дождевых червей более плотно заселены мелкотравные, щитовниково-мелкотравные, разнотравные и лещиново-разнотравные сообщества, менее заселены – мертвопокровные, зеленомошные и рододендровые (Geraskina, Shevchenko, 2021).
Рассмотрим возможность оценки эффективности выполнения лесами функции регулирования цикла углерода на основе разрабатываемого нами подхода к оценке взаимосвязей между растительностью, почвенными макросапрофагами и индикаторами экосистемных процессов, обеспечивающих секвестрацию углерода в почвах, на примере девяти доминирующих типов хвойно-широколиственных лесов европейской части России. Оценка данной экосистемной функции лесов приобретает особенно высокое значение в современный период климатических изменений.
Для апробации предложенного подхода были выбраны объекты, представляющие стадии послерубочных сукцессий хвойно-широколиственных лесов европейской части России в автоморфных позициях ландшафтов, формирующиеся в разных климатических условиях на почвообразующих породах различного гранулометрического состава (табл. 1). На суглинистых почвообразующих породах проводили исследования в трех типах лесов Северо-Западного Кавказа (СЗК) и трех типах лесов Москворецко-Окской равнины (МОР), на песчаных – в трех типах лесов Брянского полесья (БП). В 2016 г. в каждом типе леса заложено по три постоянных пробных площади размером 0.25 га; всего заложено 27 пробных площадей.
Подробное физико-географическое, геоботаническое описание всех объектов исследования, а также детальная характеристика макросапрофагов и почв, геоботанические, зоологические методы и методы химического анализа почв изложены в ряде недавних работ (Аккумуляция …, 2018; Горнов и др., 2018; Казакова и др., 2018, Шевченко и др., 2019; Кузнецова и др., 2019; Гераськина, 2020).
К информативным индикаторам участия растительности в формировании лесами функции регулирования цикла углерода можно отнести проективное покрытие функциональных групп растений, различающихся по качеству опада, и, соответственно, по скорости его разложения. Качество опада учитывается через выделение трех функциональных групп по преобладанию в проективном покрытии в ярусе А древесных растений: (1) хвойные; лиственные с быстро- (2) и медленноразлагаемым (3) опадом. В качестве индикаторов выбрана доля этих групп растений. Оценивали также проективное покрытие разных функциональных групп деревьев и кустарников из яруса В: хвойных и лиственных. Растения напочвенного покрова (ярус С) могут также вносить существенный вклад в регулирование цикла углерода. К индикаторам первой группы растений этого яруса отнесено проективное покрытие кустарничков, кустарников и деревьев (КС) с низким качеством опада, к индикаторам второй группы – проективное покрытие травянистых, т.е. злаков, осок, всех трав (ТС) с высоким качеством опада. Всего в каждом типе леса сделано по 11 геоботанических описаний (20 × 20 м), на основе которых и выявлено участие растений с разным качеством опада.
Доля лиственных растений с быстроразлагаемым опадом в покрытии яруса А лесов на песчаных почвообразующих породах (БП) варьирует в среднем незначительно – от 19 до 21%, тогда как в лесах на суглинках вариации существенно больше: МОР – от 17 до 97%, СЗК – от 0.4 до 86% (табл. 1). Доля лиственных растений с медленно разлагаемым опадом в покрытии яруса А лесов БП варьирует значительно, от 2 до 31%, на суглинках МОР – от 3 до 11%, СЗК – от 15 до 59%. Доля хвойных в покрытии яруса А лесов БП варьирует от 48 до 83%, на суглинках МОР – от 0 до 73%, СЗК – от 0 до 41%. Таким образом, в целом различия в участии растений с опадом разного качества между разными типами леса в трех регионах весьма велики, но для формирования экосистемных процессов, связанных с циклом углерода, важно учитывать распределение растений с разным качеством опада на одних и тех же объектах.
Таблица 1.
Содержание глинистых частиц в почвообразующих породах и индикаторы растительности, почвенных макросапрофагов и экосистемных процессов
| Северо-Западный Кавказ | Москворецко-Окская равнина | Брянское полесье | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Показатели | осиново-грабовые мелкотравные, 50–60 лет* |
пихто-буково-грабовые мелкотравные, 80–110 лет |
пихто-
буковые мертво-покровные, >450 лет |
липово-березовые неморально-травные, 60–70 лет |
липовые неморально-травные, 90–110 лет |
дубово-еловые бореально-неморально-травные, >100 лет |
сосняки кустарничково-зелено-мошные, 40–60 лет |
сосняки сложные неморально- бореальнотравные, 70–120 лет |
полидоми-нантные с елью неморально-травные, >120 |
| Содержание глинистых частиц в почвообразующих породах | |||||||||
| <0.002 мм, % | 25.5 ± 3.4 | 25.9 ± 4.8 | 17.9 ± 1.2 | 26.3 ± 1.3 | 19.4 ± 0.2 | 19.5 ± 0.4 | 1.1 ± 0.4 | 2.1 ± 0.4 | 1.7 ± 0.1 |
| Индикаторы растительности | |||||||||
| ЛбА, % | 85.5 ± 0.5 | 49.9 ± 11.1 | 0.4 ± 0.4 | 94.3 ± 2.5 | 96.8 ± 2.1 | 16.7 ± 8 | 19.3 ± 3.9 | 21.2 ± 9.6 | 21.2 ± 4.8 |
| ЛмА, % | 14.5 ± 0.5 | 25.9 ± 7.5 | 59 ± 1.7 | 4.3 ± 1.4 | 3.2 ± 2.1 | 10.7 ± 1.8 | 2.2 ± 2.2 | 13.8 ± 2.2 | 31.1 ± 1.7 |
| ХвА, % | 0 | 24.1 ± 3.9 | 40.6 ± 1.5 | 1.4 ± 1.1 | 0 | 72.6 ± 9.8 | 83 ± 5.8 | 64.9 ± 7.5 | 47.7 ± 4.6 |
| ЛВ, % | 7.8 ± 2.3 | 15.5 ± 4.5 | 4.2 ± 3 | 34.2 ± 6.8 | 55 ± 6.1 | 46.4 ± 1.1 | 23 ± 11.1 | 40.3 ± 2.2 | 35 ± 5.5 |
| ХвВ , % | 10 ± 5.1 | 42.8 ± 7.1 | 10.8 ± 6.4 | 0.8 ± 0.7 | 0 | 3.6 ± 1.1 | 1.7 ± 1 | 5.8 ± 2.6 | 1.1 ± 0.6 |
| ТС, % | 19.5 ± 2.4 | 32.6 ± 2.9 | 0.7 ± 0.3 | 78.2 ± 1.7 | 77.6 ± 4.1 | 64 ± 4.3 | 16.4 ± 2.7 | 33.4 ± 9.1 | 57.2 ± 0.9 |
| КС, % | 19.9 ± 3 | 27.4 ± 2 | 4.6 ± 0.2 | 3 ± 0.7 | 2.4 ± 0 | 6 ± 2.3 | 28.6 ± 5.1 | 16.6 ± 1.4 | 10.6 ± 0.7 |
| Индикаторы макросапрофагов | |||||||||
| БмП, г м–2 | 2.2 ± 0.8 | 1.7 ± 1.1 | 0.4 ± 0.3 | 0 ± 0 | 1.6 ± 1.3 | 1.8 ± 0.4 | 0 ± 0 | 3.8 ± 0.3 | 5 ± 1.4 |
| БмПП, г м–2 | 0.6 ± 0.6 | 0 ± 0 | 0 ± 0 | 4.3 ± 2.9 | 2.3 ± 0.8 | 10.5 ± 4.1 | 0 ± 0 | 1.3 ± 1.3 | 0 ± 0 |
| БмСП, г м–2 | 6 ± 2.1 | 6.4 ± 0.7 | 9.9 ± 4.3 | 11.8 ± 3.5 | 5.3 ± 3.1 | 3.6 ± 1.3 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| БмН, г м–2 | 0 ± 0 | 1.2 ± 0.7 | 7.5 ± 3 | 0 ± 0 | 6.7 ± 4.6 | 4.3 ± 1.4 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| Индикаторы экосистемных процессов (L-подгоризонт подcтилки) | |||||||||
| C : N | 23 ± 1 | 25 ± 0 | 25 ± 1 | 27 ± 1 | 26 ± 0 | 21 ± 2 | 32 ± 2 | 22 ± 1 | 19 ± 1 |
| СН, % | 94 ± 0 | 92 ± 0 | 91 ± 1 | 92 ± 0 | 91 ± 0 | 91 ± 1 | 77 ± 1 | 86 ± 1 | 91 ± 1 |
| Индикаторы экосистемных процессов (FH-подгоризонт подcтилки) | |||||||||
| C : N | 18 ± 1 | 20 ± 1 | 23 ± 0 | 20 ± 0 | 18 ± 0 | 19 ± 0 | 29 ± 2 | 20 ± 1 | 16 ± 1 |
| СН, % | 97 ± 0 | 96 ± 0 | 95 ± 0 | 96 ± 0 | 96 ± 0 | 94 ± 0 | 67 ± 6 | 88 ± 2 | 94 ± 2 |
| Индикаторы экосистемных процессов (гумусовый горизонт А) | |||||||||
| C : N | 12 ± 0 | 12 ± 0 | 11 ± 0 | 11 ± 1 | 11 ± 0 | 13 ± 0 | 20 ± 1 | 16 ± 1 | 13 ± 1 |
| СН, % | 76 ± 3 | 71 ± 9 | 71 ± 9 | 89 ± 2 | 88 ± 0 | 76 ± 8 | 31 ± 3 | 71 ± 16 | 88 ± 4 |
| Запас почвенного углерода | |||||||||
| FH, C запас, т га–1 | 1.3 ± 0.3 | 1.9 ± 0.1 | 1.5 ± 0.1 | 0.1 ± 0 | 0.1 ± 0.1 | 0.8 ± 0.1 | 9.7 ± 0.7 | 3.1 ± 0.2 | 3.5 ± 0.2 |
| 0–50 см, C запас, т га–1 | 82.5 ± 4.5 | 82.7 ± 0.8 | 74.9 ± 4.3 | 82.8 ± 7.5 | 67.6 ± 1.1 | 76.2 ± 3.2 | 39 ± 4.7 | 58.2 ± 8.7 | 45 ± 5.9 |
Примечание. ЛбА – вклад проективного покрытия лиственных с быстроразлагаемым опадом в ярусе А; ЛмА – вклад проективного покрытия лиственных с медленноразлагаемым опадом в ярусе А; ХвА – вклад проективного покрытия хвойных в ярусе А; ЛВ – проективное покрытие лиственных в ярусе В; ХвВ – проективное покрытие хвойных в ярусе В; ТС – проективное покрытие травянистых растений в ярусе С; КС – проективное покрытие древесных растений в ярусе С; БмП – биомасса подстилочной функциональной группы дождевых червей; БмПП – биомасса почвенно-подстилочной функциональной группы дождевых червей; БмСП – биомасса собственно-подстилочной функциональной группы дождевых червей; БмН – биомасса норной функциональной группы дождевых червей; СН – степень насыщенности основаниями; * – возраст древостоя.
На объектах БП наиболее равномерное распределение в ярусе А древесных растений с разным качеством опада наблюдается в сосняках сложных и полидоминантных широколиственных лесах. При этом в обоих типах леса в ярусе В доминируют лиственные растения и хорошо развит травяной покров. Напротив, сосняки кустарничково-зеленомошные отличаются ярким доминированием в ярусе А сосны (вклад 83%), подрост лиственных не развит, в ярусе С доминируют древесные, кустарники и кустарнички.
В липовых лесах МОР с выраженным доминированием липы во всех ярусах представлены в основном растения с быстроразлагаемым опадом, тогда как в дубово-еловых лесах растения с разным качеством опада распределены более равномерно. Липово-березовые леса с доминированием березы по качеству опада занимают промежуточное положение. Отношение лигнин : N в стареющих листьях березы достигает 21, тогда как в стареющих листьях липы не превышает 13 (Jabiol et al., 2019), и поэтому по скорости разложения листья березы существенно уступают листьям липы, которые, как правило, не успевают сформировать выраженный подгоризонт опада из-за быстрой переработки почвенной биотой.
На объектах СЗК в осиново-грабовых лесах практически отсутствуют хвойные и доминируют лиственные растения с быстроразлагаемым опадом листьев граба (86%), который отличается узким отношением лигнин : N – 9 (Jabiol et al., 2019). В пихтово-буково-грабовых лесах растения с разным качеством опада распределены более равномерно. В пихтово-буковых лесах в ярусе А доминируют лиственные (59%) и хвойные (41%) с медленно разлагаемым опадом. В осиново-грабовых и пихтово-буково-грабовых лесах развит травяной покров, тогда как в темных пихтово-буковых лесах травяной покров не развит. Таким образом, во всех трех регионах функционируют типы леса с разным соотношением функциональных групп растений с быстро- и медленноразлагаемым опадом.
К информативным индикаторам участия почвенной биоты в формировании хвойно-широколиственными лесами функции регулирования цикла углерода можно отнести состав и биомассу функциональных групп доминирующих по биомассе почвенных макросапрофагов – дождевых червей. На исследуемых объектах равнинных и горных лесов отобрано около 300 проб: БП – 90, МОР – 60, СЗК – 144 пробы размерами 25 × 25 см, глубиной 30 см. Полевые работы выполнены в весенне-летние периоды 2016 и 2018 гг. Проведен количественный учет макросапрофагов и измерена их биомасса. Морфо-экологические (функциональные) группы дождевых червей приведены согласно Т.С. Перель (1979): подстилочная, почвенно-подстилочная, собственно почвенная и норная группы.
Биомасса дождевых червей в лесах БП изменяется в среднем от 0 до 5 г м–2 , на суглинках МОР – от 16 до 20 г м–2, на суглинках СЗК – от 9 до 18 г м–2. Несмотря на благоприятный гранулометрический состав для дождевых червей в почвах СЗК, показатель биомассы в лесах этого региона широко варьирует и его наименьшие значения близки к биомассе червей в лесах БП, что, прежде всего, связано с редким, но существенно подрывающим популяции дождевых червей явлением – пересыханием почвы в летний сезон (Geraskina, Shevchenko, 2021). При этом в лесах БП представлена в основном функциональная группа подстилочных червей, что объясняется неблагоприятным для развития других групп гранулометрическим (песчаным) составом почвообразующих пород, тогда как в лесах МОР и СЗК, формирующихся на суглинках, представлены все четыре функциональные группы.
Для разработки классификации лесов по эффективности выполнения ими функции регулирования цикла углерода целесообразно предложить интегральные индикаторы экосистемных процессов, связанных с секвестрацией углерода в почвах, отражающих участие биоты в формировании этих процессов. К таким информативным и просто измеряемым индикаторам экосистемных процессов, протекающих в почве, можно отнести отношение C : N в аккумулятивных горизонтах почв (подгоризонты L и FH подстилки и гумусовый горизонт А) и степень насыщенности их основаниями. Чем меньше отношение C : N в аккумулятивных горизонтах и чем выше степень их насыщенности основаниями, тем быстрее идут процессы разложения органического вещества и, соответственно, это может приводить к более интенсивной эмиссии углекислого газа и миграции соединений углерода с почвенными водами в нижние минеральные горизонты и их выносу за пределы биогеоценоза. Результат взаимодействия всех этих процессов и многолетней секвестрации соединений углерода может оцениваться на основе запаса углерода в подстилке и минеральных слоях почв. Для оценки этих индикаторов в каждом типе леса отобрано по 36 образцов подстилки и гумусового горизонта А.
Отношение C : N варьирует в среднем от 23 до 25 в подгоризонте подстилки L, от 18 до 23 в подгоризонте подстилки FH и от 11 до 12 в горизонте А почв лесов СЗК; от 21 до 27 в L, от 18 до 20 в FH и от 11 до 13 в горизонте А почв лесов МОР; от 19 до 32 в L, от 15 до 29 в FH и от 13 до 20 в горизонте А почв лесов БП. По этому отношению выделяются сосновые леса БП, где и в подстилке, и в гумусовом горизонте оно значительно больше. В других типах леса БП отношение C : N в подстилке сопоставимо с лесами на суглинках, но в гумусовом горизонте А оно во всех типах леса БП больше, чем в лесах на суглинках.
Степень насыщенности основаниями варьирует в среднем от 91 до 94% в L подгоризонте, от 95 до 97% в FH и от 71 до 76% в горизонте А почв лесов СЗК; от 91 до 92% в L, от 94 до 96% в FH и от 76 до 89% в горизонте А почв лесов МОР; от 77 до 91% в L, от 67 до 94% в FH и от 31 до 88% в горизонте А почв лесов БП. Таким образом, по степени насыщенности основаниями также выделяются сосновые леса БП, где этот показатель и в подстилке, и в гумусовом горизонте значительно меньше.
Запасы почвенного углерода изменяются в среднем от 1 до 2 т га–1 в подстилке (FH) и от 75 до 83 т га–1 в слое 0–50 см в почвах лесов СЗК, от 0.1 до 1 т га–1 в подстилке (FH) и от 68 до 83 т га–1 в слое 0–50 см в почвах лесов МОР, и от 3 до 10 т га–1 в подстилке (FH) и от 39 до 59 т га–1 в слое 0–50 см в почвах лесов БП. При этом запасы углерода в древесной биомассе увеличивались с возрастом древостоев особенно значительно на объектах СЗК: от 109 т га–1 в самых молодых осиново-грабовых лесах до 127 т га–1 в пихтово-буково-грабовых и 482 т га–1 в пихтово-буковых лесах. На объектах БП изменения запасов углерода в древесной биомассе с возрастом древостоя также были весьма значительны: от 77 т га–1 в сосняках кустарничково-зеленомошных до 209 т га–1 в полидоминантных широколиственных лесах. Гораздо менее выраженное увеличение запаса углерода в древесной биомассе с возрастом древостоя демонстрируют леса МОР: от 159 до185 т га–1.
Таким образом, самые высокие запасы углерода в подстилке демонстрируют леса БП, которые, напротив, отличаются небольшими запасами углерода в минеральном слое 0–50 см. Во всех регионах наименьшие запасы углерода в этом слое почвы обнаруживаются в типах леса с выраженным доминированием одной функциональной группы: или с быстро-, или с медленноразлагаемым опадом. Среди объектов МОР менее значительные запасы углерода в минеральном слое 0–50 см обнаруживаются в лесах с доминированием липы, среди объектов БП – в лесах с доминированием сосны (сосняки кустарничково-зеленомошные), а на объектах СЗК – в пихтово-буковых лесах с доминированием пихты и бука, отличающихся низким качеством опада, хотя запас древесины и количество древесного опада здесь отличаются самыми высокими значениями (Аккумуляция …., 2018).
Запасы углерода в почвах, в отличие от его запаса в древесной биомассе, не связаны с возрастом древостоев. Доля почвенного углерода от суммарного запаса углерода в древесной биомассе и почвах, включая подстилку, снижалась с возрастом древостоев от 43 до 14% на объектах СЗК и от 39 до 19% на объектах БП и составляла 29–30% на всех объектах МОР. В пихтово-буковых лесах СЗК и в полидоминантных широколиственных лесах БП, формируемых самыми высоковозрастными древостоями, запасы почвенного углерода оказались относительно невысокими. В пихтово-буковых лесах, возраст древостоя которых достигает 450 лет и более, это можно объяснить комбинированным действием нескольких факторов: (1) значительный объем осадков (2000 мм), т.е. климатический фактор, (2) хорошо известный низкий уровень транспирации деревьями бука (биотический фактор), что вместе с первым фактором приводит к развитию процессов оглеения, промывания почвенного профиля и выносу соединений углерода из почв, и (3) медленноразлагаемый опад бука и пихты (биотический фактор). О высокой влагонасыщенности минерального профиля в этих лесах свидетельствуют и большая численность и биомасса норных червей, функционирующих в глубоких почвенных слоях (Гераськина, 2018).
В полидоминантных широколиственных лесах БП значительная мощность элювиальных горизонтов, достигающая 50 см, также свидетельствует о периодическом промывании почвенного профиля и выносе соединений углерода из почв, которые не компенсируются действием быстроразлагаемых компонентов опада лиственных древесных растений и трав. Таким образом, результаты данных исследований подтверждают, что водный режим, наряду с биотическими факторами и другими абиотическими факторами, может играть значительную роль в формировании почвенного пула углерода лесов и регулировании цикла углерода в лесах.
Для оценки вклада предикторов – индикаторов растительности, дождевых червей и экосистемных процессов, а также почвообразующих пород в варьирование запасов почвенного углерода выбран метод иерархического разложения (Chevan, Sutherland, 1991). Данный метод основан на разложении R2 согласно иерархии всех возможных линейных моделей, которые можно построить с данным набором предикторов. Иерархическое разложение не ставит целью нахождение лучшей модели или некоторого оптимального набора предикторов, но позволяет оценить относительные вклады предикторов в вариацию отклика. Существенным свойством метода, важным и в настоящей работе, является его способность нивелировать проблему мультиколлинеарности предикторов. Для каждого предиктора рассчитываются два вида вкладов: 1) независимый и 2) совместный, проявляемый в присутствии других предикторов. Последний может принимать отрицательные значения, если среди предикторов есть “супрессоры” (Chevan, Sutherland, 1991). На рис. 1–2 даны два вида вкладов и R2 для полной модели (в скобках значение R2, скорректированного на число предикторов). Проценты по оси ординат рассчитаны от дисперсии, объясненной моделью, а не от общей вариации в данных. Анализ выполнен с помощью пакета hier.part (Mac Nally, Walch, 2004) в среде статистического программирования R (R Core Team, 2020).
Рис. 1.
Вклад основных предикторов в дисперсию биомассы дождевых червей. Предикторы: 0.002 – содержание глинистых частиц, ЛбА – вклад проективного покрытия лиственных с быстроразлагаемым опадом в ярусе А, ЛВ – проективное покрытие лиственных в ярусе В, ТС – проективное покрытие травянистых растений в ярусе С.
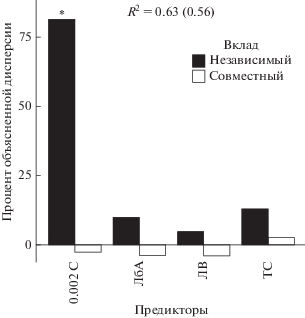
Рис. 2.
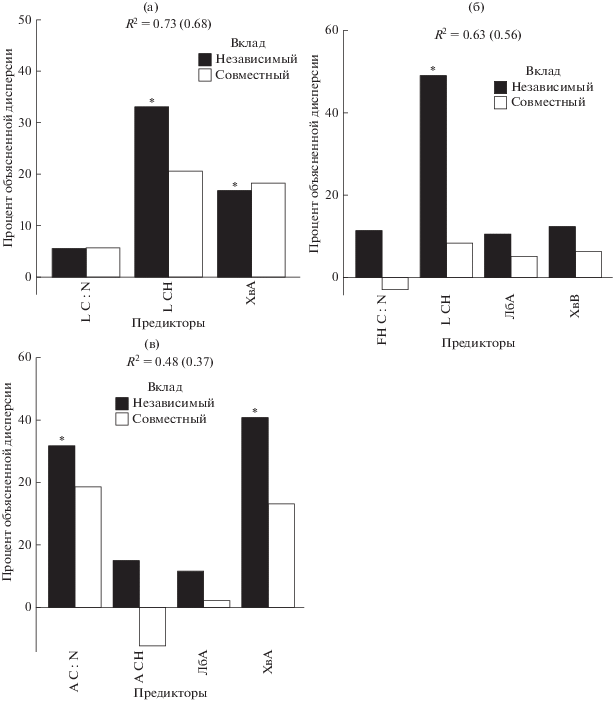
Вклад основных предикторов в дисперсию запаса углерода в почвах. Предикторы: LСН – степень насыщенности основаниями в подгоризонте L подстилки, L C : N – отношение углерода к азоту в подгоризонте L подстилки, ХвА – вклад проективного покрытия хвойных в ярусе А, FH C : N – отношение углерода к азоту в подгоризонте FH подстилки, ЛбА – вклад проективного покрытия лиственных с быстроразлагаемым опадом в ярусе А, ХвВ – проективное покрытие хвойных в ярусе В, A CH – отношение углерода к азоту в горизонте A, А СН – степень насыщенности основаниями в горизонте А. Отклики: а – запасы углерода подстилки, б – запасы углерода в минеральном слое 0–50 см, в – запасы углерода в минеральном слое 15–50 см.
В качестве отклика выбран запас углерода в подстилке и минеральных слоях почв. В качестве предикторов оценивали долю частиц меньше 0.002 мм в почвообразующих породах как характеристику гранулометрического состава, индикаторы функционального разнообразия растительности (ярусы А, В и С), позволяющие учитывать как количество (проективное покрытие), так и качество опада, а также отношение C : N, степень насыщенности основаниями аккумулятивных горизонтов почв (подстилки и гумусового горизонта А).
Биомасса и функциональное разнообразие дождевых червей также рассматривались как отклик, поскольку их функционирование лимитируется гранулометрическим составом почвообразующих пород и обусловлено качеством опада. С другой стороны, биомасса червей рассматривается как предиктор, поскольку черви значительно влияют на реализацию рассматриваемых экосистемных процессов и, соответственно, накопление углерода в почвах. К переменным применялись необходимые преобразования для выполнения статистических требований общей линейной модели.
Наиболее значимые предикторы секвестрации углерода в почвах выявлялись на основе иерархического анализа вкладов потенциальных предикторов. Анализ предикторов биомассы червей как отклика с учетом объектов всех трех регионов показал, что, как и следовало ожидать, доля глинистых частиц в почвообразующей породе вносит наибольший вклад в дисперсию биомассы червей (около 80%), а совокупный индивидуальный вклад растений с быстроразлагаемым опадом из всех трех ярусов составляет около 20% (рис. 1). При этом связь биомассы червей со всеми этими предикторами положительная.
Существенный вклад (от 30 до 50%) в дисперсию запасов углерода и в FH подгоризонте подстилки, и в минеральном слое 0–50 см (рис. 2а, 2б) вносит качество опада (L подгоризонт подстилки), а именно, степень его насыщенности основаниями: чем выше степень насыщенности опада основаниями, тем меньше запас подстилки и, соответственно, запас углерода в ней и, напротив, тем больше запас углерода в минеральных горизонтах почв, в которые мигрируют соединения углерода из богатого основаниями быстро разлагающегося опада. Вклад функциональных групп растений с опадом разного качества в дисперсию запасов углерода может достигать от 18% в подстилке до 25% (совокупный вклад) в минеральном полуметровом слое. Связь вариации запасов углерода в подстилке со вкладом в общее проективное покрытие хвойных растений с медленноразлагаемым опадом положительная. Запасы в минеральном слое 0–50 см положительно связаны со вкладом растений с быстроразлагаемым опадом и отрицательно – со вкладом растений с медленно разлагаемым опадом.
Обнаруживается также связь между отношением C : N в гумусовом горизонте А и запасами почвенного углерода: чем меньше отношение C : N в гумусовом горизонте А, тем больше запас углерода в нижележащих минеральных горизонтах почв (слой 15–50 см) (рис. 2в). При этом между отношением C : N в подгоризонтах L и FH подстилки и в гумусовом горизонте А существует тесная положительная связь (r > 0.5–0.7). Это подтверждает, что отношение C : N в нижних подгоризонтах подстилки (FH) и в гумусовом горизонте A наследуется из опада (L).
Таким образом, качество опада разных функциональных групп вносит существенный вклад в аккумуляцию почвенного углерода. Можно было ожидать, что чем интенсивнее поток органического вещества из опада и подстилки, обеспечиваемый в первую очередь растениями с быстроразлагаемым опадом высокого качества, тем больше углерода накапливается в минеральных слоях почвы. Однако наибольший уровень аккумуляции почвенного углерода обнаруживается в лесах с более равномерным распределением растений с разным качеством опада. Объяснить это можно следующим образом. Дождевые черви, являющиеся активными переработчиками опада, влияющими и на функционирование грибов, формируют функциональные группы (подстилочные, почвенно-подстилочные, собственно почвенные и норные), связанные как с подстилкой, так и с минеральными горизонтами почв. Подстилка для части из них является не только источником питания, но и местообитанием, и для оптимального функционирования червей сохранение подстилки имеет ключевое значение. Показано, что для дождевых червей более благоприятен смешанный хвойно-лиственный опад, формируемый растительностью разных ярусов, так как за счет трудноразлагаемых фракций сохранится подстилка, которая служит местообитанием дождевым червям, а легкоразлагаемые фракции – трофическим ресурсом высокого качества (Sariyildiz, 2008; Sariyildiz, Küçük, 2008; Гераськина, 2020). При высокой скорости разложения быстроразлагаемого опада уровень эмиссии углекислого газа может быть высоким, но благодаря деятельности дождевых червей соединения углерода закрепляются в почвах за счет обеспечения процессов гумусообразования и переноса органического вещества почвы в более глубокие горизонты в результате роющей деятельности (Frouz et al., 2013). Поэтому дождевые черви играют ключевую роль в экосистемных процессах, способствующих накоплению углерода в экосистемах. В лесах с более равномерным распределением растений с разным качеством опада для таких активных переработчиков опада, как дождевые черви, складываются более благоприятные условия для функционирования, что в итоге приводит к повышению эффективности экосистемных процессов, обеспечивающих секвестрацию почвенного углерода в этих лесах.
Широкомасштабная оценка взаимосвязей между растительностью, почвенной биотой и всеми экосистемными процессами, связанными с продуцированием и разложением органического вещества, позволит классифицировать леса по эффективности выполнения ими функции регулирования цикла углерода, что является актуальной научной задачей в связи с изменениями климата.
Важнейшей проблемой является поиск информативных показателей экосистемных процессов, обеспечивающих и другие экосистемные функции. Эти оценки позволят создать функциональную классификацию лесов с учетом взаимосвязей (синергии и компромиссы) между всеми функциями.
Заключение. Разработка функциональной классификации лесов является актуальной проблемой. Такая классификация может быть разработана на основе оценки эффективности выполнения различных экосистемных функций лесами разных типов/стадий сукцессий. Одним из подходов к оценке эффективности функции регулирования цикла углерода в лесах является изучение взаимосвязей между представителями биоты разных трофических уровней, отвечающих как за продуцирование органического вещества, так и его разложение и секвестрацию в почвах, где депонируется значительная часть экосистемного углерода. Проведенные исследования в хвойно-широколиственных лесах демонстрируют, что самым низким уровнем аккумуляции почвенного углерода отличаются леса на песчаных почвах, характеризующиеся небольшим функциональным разнообразием и низкой биомассой дождевых червей. Высоким запасом углерода в почвах, формирующихся как на суглинках, так и на песках, отличаются хвойно-широколиственные леса с более равномерно распределенным составом растений с разным качеством опада. Это объясняется благоприятными трофическими и топическими условиями для функционирования почвенной биоты, прежде всего дождевых червей, которые являются активными переработчиками опада в этих лесах.
Широкомасштабные исследования взаимосвязей между растительностью, почвенной биотой и экосистемными процессами как в хвойно-широколиственных, так и в доминирующих в России бореальных лесах, открывают перспективу развития классификации лесов по эффективности выполнения ими функции регулирования цикла углерода. Участие лесов в экосистемных процессах, связанных с формированием цикла углерода, может оцениваться на основе несложно измеряемых индикаторов. Для таких оценок могут использоваться существующие и новые данные геоботанических описаний и исследований почвенной биоты в лесах, а также анализ связей между ними. При этом необходимо учитывать абиотические факторы, к которым относятся климатические условия, почвообразующие породы, рельеф, водный режим.
Разработка подходов к оценке эффективности выполнения различных экосистемных функций лесами разных типов, идентифицированных в России на основе классификаций растительности, позволит создать функциональную классификацию лесов.
Список литературы
Аккумуляция углерода в лесных почвах и сукцессионный статус лесов / Под ред. Лукиной Н.В. М.: Товарищество научных изданий КМК, 2018. 232 с.
Алексеев Е.В. Типы Украинского леса. Правобережье. Киев, 1928. 120 с.
Бабенко А.C. Почвенные беспозвоночные как индикаторы состояния территории // Геохимия живого вещества: материалы Международной молодежной школы-семинара (2–5 июня 2013 г., г. Томск). Томск, 2013. С. 40–42.
Барталев С.А., Стыценко Ф.В. Спутниковая оценка гибели древостоев от пожаров по данным о сезонном распределении пройденной огнем площади // Лесоведение. 2021. № 2. С. 115–122.
Бобровский М.В. Лесные почвы Европейской России: биотические и антропогенные факторы формирования. М.: Товарищество научных изданий КМК, 2010. 359 с.
Браславская Т.Ю., Заугольнова Л.Б. Классификация и типология лесной растительности как инструмент исследования лесного покрова // Методические подходы к экологической оценке лесного покрова в бассейне малой реки. М.: Товарищество научных изданий КМК, 2010. С. 56–71.
Гераськина А.П. Преобразования комплекса дождевых червей в ходе послерубочных сукцессий в лесах Северо-Западного Кавказа // Вопросы лесной науки. 2018. № 1. С. 1–14. https://doi.org/10.31509/2658-607x-2018-1-1-1-14
Гераськина А.П. Влияние дождевых червей разных морфо-экологических групп на аккумуляцию углерода в лесных почвах // Вопросы лесной науки. 2020. Т. 3. № 2. С. 1–20. https://doi.org/10.31509/2658-607x-2020-3-2-1-20
Гиляров М.С. Учет крупных беспозвоночных (мезофауны) // Методы почвенно-зоологических исследований. М.: Наука, 1975. С. 12–29.
Горнов А.В., Горнова М.В., Тихонова Е.В., Шевченко Н.Е., Кузнецова А.И., Ручинская Е.В., Тебенькова Д.Н. Оценка сукцессионного статуса хвойно-широколиственных лесов европейской части России на основе популяционного подхода // Лесоведение. 2018. № 4. С. 243–257. https://doi.org/10.1134/S0024114818040083
Заугольнова Л.Б. Подходы к оценке типологического разнообразия лесного покрова // Мониторинг биологического разнообразия лесов России: методология и методы. М.: Наука, 2008. С. 36–58.
Заугольнова Л.Б., Мартыненко В.Б. Определитель типов леса Европейской России URL: http://cepl.rssi.ru/ bio/forest/index.htm (дата обращения 01.05.2021).
Казакова А.И., Семиколенных А.А., Горнов А.В., Горнова М.В., Лукина Н.В. Влияние растительности на лабильные характеристики лесных почв зандровых местностей заповедника “Брянский лес” // Вестник Московского университета. Серия 17: Почвоведение. 2018. № 3. С. 9–15.
Каяндер А.К. Сущность и значение типов леса. М.: Гослестехиздат, 1933. 36 с.
Классификация и диагностика почв России / Сост. Шишов Л.Л., Тонконогов В.Д., Лебедева И.И., Герасимова М.И. Смоленск: Ойкумена, 2004. 342 с.
Кузнецова А.И., Лукина Н.В., Тихонова Е.В., Горнов А.В., Горнова М.В., Смирнов В.Э., Гераськина А.П., Шевченко Н.Е., Тебенькова Д.Н., Чумаченко С.И. Аккумуляция углерода в песчаных и суглинистых почвах равнинных хвойно-широколиственных лесов в ходе восстановительных сукцессий // Почвоведение. 2019. № 7. С. 803–816. https://doi.org/10.1134/S0032180X19070086
Курганова И.Н. Эмиссия и баланс диоксида углерода в наземных экосистемах России: дис. … д-ра биол. наук: 03.00.27; 03.00.16. М.: МГУ, 2010. 325 с.
Лукина Н.В., Ершов В.В., Горбачева Т.Т., Орлова М.А. (Данилова М.А.), Исаева Л.Г., Тебенькова Д.Н. Оценка состава почвенных вод северо-таежных хвойных лесов фоновых территорий индустриально развитого региона // Почвоведение. 2018. № 3. С. 284–296. https://doi.org/10.7868/S0032180X18030036
Перель Т.С. Распространение и закономерности распределения дождевых червей фауны СССР. М.: Наука, 1979. 272 с.
Погребняк П.С. Лесорастительные условия левобережного Полесья // Труды по опытному лесному делу Украины. 1928. № 10. С. 46–78.
Погребняк П.С. Основы лесной типологии. К.: Изд-во АН УССР, 1955. 452 с.
Пулы и потоки углерода в наземных экосистемах России / Под ред. Г.А. Заварзина. М.: Наука, 2007. 315 с.
Розанов Б.Г. Морфология почв. М.: Академический проект, 2004. 432 с.
Смирнова О.В., Ханина Л.Г., Смирнов В.Э. Эколого-ценотические группы в растительном покрове лесного пояса Восточной Европы // Восточноевропейские леса: история в голоцене и современность. Кн. 1. М.: Товарищество научных изданий КМК, 2004. С. 165–175.
Сукачев В.Н., Дылис Н.В. Основы лесной биогеоценологии. М.: Наука, 1964. 574 с.
Тебенькова Д.Н., Лукина Н.В., Данилова М.А., Кузнецова А.И., Горнов А.В., Шевченко Н.Е., Катаев А.Д., Гагарин Ю.Н., Чумаченко С.И. Мультифункциональность и биоразнообразие лесных экосистем // Лесоведение. 2019. № 5. С. 341–356. https://doi.org/10.1134/S0024114819050115
Уткин А.И. Биологическая продуктивность лесов (методы изучения и результаты) // Лесоведение и лесоводство. 1975. Т. 1. С. 9–190.
Холхоева Л.С. Некоторые особенности распространения дождевых червей по типам почв Левобережной Украины // Известия Харьковского энтомологического общества. 1993. Т. 2. № 1. С. 125–130.
Ценофонд лесов Европейской России. URL: http://cepl. rssi.ru/bio/flora/main.htm (дата обращения 28.06.2021).
Чернов Ю.И. Природная зональность и животный мир суши. М.: Наука, 1975. 225 с.
Шевченко Н.Е., Кузнецова А.И., Тебенькова Д.Н., Смирнов В.Э., Гераськина А.П., Горнов А.В., Тихонова Е.В., Лукина Н.В. Сукцессионная динамика запасов почвенного углерода и растительности хвойно-широколиственных лесов Северо-Западного Кавказа // Лесоведение. 2019. № 3. С. 163–176. https://doi.org/10.1134/S0024114819030082
Berg B., McClaugherty C. Plant Litter. Decomposition, Humus Formation, Carbon Sequestration. Springer Cham, 2020. 332 p. https://doi.org/10.1007/978-3-030-59631-6
Cadisch G., Giller K.E. Driven by Nature: Plant Litter Quality and Decomposition. Wallingford: CAB International, 1997. 432 p.
Cao J., Wang C., Dou Z., Liu M., Ji D. Hyphospheric impacts of earthworms and arbuscular mycorrhizal fungus on soil bacterial community to promote oxytetracycline degradation // J. Hazard Mater. 2018. V. 341. P. 346–354. https://doi.org/10.1016/j.jhazmat.2017.07.038
Chevan A., Sutherland M. Hierarchical partitioning // The American Statistician. 1991. V. 45. P. 90–96.
Cornelissen J.H., Lang S.I., Soudzilovskaia N.A., During H.J. Comparative cryptogam ecology: A review of bryophyte and lichen traits that drive biogeochemistry // Annals of Botany. 2007. V. 99. P. 987–1001.
Framstad E., Wit H., Mäkipää R., Larjavaara M., Vesterdal L., Karltun E. Biodiversity, Carbon Storage and Dynamics of Old Northern Forests. Copenhagen, Denmark. 2013. https://doi.org/10.6027/TN2013-507
Frouz J., Livečková M., Albrechtová J., Chroňáková A., Cajthaml T., Pižl V., Háněl L., Starý J., Baldrian P., Lhotáková Z., Šimáčková H., Cepáková Š. Is the effect of trees on soil properties mediated by soil fauna? A case study from post-mining sites // Forest Ecology and Management. 2013. V. 309. P. 87–95. https://doi.org/10.1016/j.foreco.2013.02.013
Geraskina A., Shevchenko N. Distribution of epi-endogeic and endogeic earthworm species (Oligochaeta: Lumbricidae) in the forest belt of the Northwest Caucasus // Zootaxa. 2021. V. 4975. № 3. P. 561–573. https://doi.org/10.11646/zootaxa.4975.3.6
Huang W., Gonzalez G., Zou X. Earthworm abundance and functional group diversity regulate plant litter decay and soil organic carbon level: A global meta-analysis // Applied Soil Ecology. 2020. V. 150. P. 1–15. https://doi.org/10.1016/j.apsoil.2019.103473
Jabiol J., Lecerf A., Lamothe S., Gessner M., Chauvet E. Litter quality modulates effects of dissolved nitrogen on leaf decomposition by stream microbial communities // Microbial Ecology. 2019. V. 77. № 4. P. 959–966. 1007/s00248-019-01353-3
Krishna M.P., Mohan M. Litter decomposition in forest ecosystems: a review // Energy, Ecology and Environment. 2017. V. 2. № 4. P. 236–249.
Kurakov A.V., Kharin S.A., Byzov B.A. Changes in the composition and physiological and biochemical properties of fungi during passage through the digestive tract of earthworms // Biological Bulletin. 2016. V. 43. P. 290–299. https://doi.org/10.1134/S1062359016040075
Kurbatova J., Tatarinov F., Molchanov A., Varlagin A., Avilov V., Kozlov D., Valentini R. Partitioning of ecosystem respiration in a paludified shallowpeat spruce forest in the southern taiga of European Russia // Environmental Research Letters. 2013. V. 8. № 4. P. 045028.
Lavelle P., Decaëns T., Aubert M., Barot S., Blouin M., Bureau F., Rossi J.P. Soil invertebrates and ecosystem service // European Journal of Soil Biology. 2006. V. 42. P. 3–15.
Mac Nally R., Walsh C. Hierarchical partitioning public-domain software // Biodiversity and Conservation. 2004. V. 13. № 3. P. 659–660.
Manning P., Taylor G., Hanley M.E. Bioenergy, food production and biodiversity – an unlikely alliance? // Global Change Biology Bioenergy. 2015. V. 7. № 4. P. 570–576. https://doi.org/10.1111/gcbb.12173
Millennium Ecosystem Assessment. Ecosystems and Human Wellbeing: Synthesis. Washington, DC: Island Press. 2005. URL: http://www.millenniumassessment.org/en/ Reports.aspx# (дата обращения 28.06.2021).
Mordkovich V.G. Zoological diagnostics of soils: Imperatives, purposes, and place within soil zoology and pedology // Biology Bulletin Reviews. 2014. V. 4. № 5. P. 404–411.
Pérez-Harguindeguy N. Chemistry and toughness predict leaf litter decomposition rates over a wide spectrum of functional types and taxa in central Argentina // Plant and Soil. 2000. V. 218. № 1. P. 21–30.
R Core Team R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria. 2020. URL: https://www.R-project.org/
Salemaa M., Derome J., Nojd P. Response of boreal forest vegetation to the fertility status of the organic layer along a climatic gradient // Boreal Environment Research. 2008. V. 13. Suppl. B. P. 48–66.
Sariyildiz T. Effects of tree canopy on litter decomposition rates of Abies nordmanniana, Picea orientalis and Pinus sylvestris // Scandinavian J. Forest Research. 2008. V. 23. № 4. P. 330–338. https://doi.org/10.1080/02827580802275816
Sariyildiz T., Küçük M. Litter mass loss rates in deciduous and coniferous trees in Artvin, northeast Turkey: Relationships with litter quality, microclimate, and soil characteristics // Turkish J. Agriculture and Forestry. 2008. V. 32. № 6. P. 547–559.
Schelfhout S., Mertens J., Verheyen K., Vesterdal L., Baeten L., Muys B., De Schrijver A. Tree species identity shapes earthworm communities // Forests. 2017. V. 8. № 3. P. 85.
Spurgeon D.J., Keith A.M., Schmidt O., Lammertsma D.R., Faber J.H. Land-use and land-management change: relationships with earthworm and fungi communities and soil structural properties // BMC Ecology. 2013. V. 13. № 1. P. 46. https://doi.org/10.1186/1472-6785-13-46
Staaf H. Foliage litter turnover and earthworm populations in three beech forests of contrasting soil and vegetation types // Oecologia. 1987. V. 72. № 1. P. 58–64.
Zhang W., Chen D., Zhao C. Functions of earthworm in ecosystem // Biodiversity Science. 2007. V. 15. № 2. P. 142–153. https://doi.org/10.1360/biodiv.060294
Дополнительные материалы отсутствуют.