

Лесоведение, 2022, № 1, стр. 47-60
Пространственная вариабельность свойств подзолов в зависимости от растительных микрогруппировок в сосняке брусничном
О. Н. Бахмет a, *, М. В. Медведева a, Е. В. Мошкина a, Ю. Н. Ткаченко a, А. В. Мамай a, С. Г. Новиков a, С. А. Мошников a, В. В. Тимофеева a, А. Ю. Карпечко a
a Институт леса КарНЦ РАН, ФИЦ “Карельский научный центр РАН”
185910 Петрозаводск, ул. Пушкинская, 11, Россия
* E-mail: obahmet@mail.ru
Поступила в редакцию 29.05.2020
После доработки 17.02.2021
Принята к публикации 03.06.2021
- EDN: GKTLQP
- DOI: 10.31857/S002411482105003X
Аннотация
Исследования проведены на особо охраняемой природной территории в западной части Европейского Севера. На примере почв альфе-гумусового генезиса (Podzols) изучена пространственная вариабельность их свойств в зависимости от типа растительной микрогруппировки (лишайниковая, брусничная, черничная, зеленомошная). Представлена морфологическая и физико-химическая характеристика исследованных почв, дано таксационное описание древостоя, изучен видовой состав растений напочвенного покрова. В связи с неоднородностью напочвенного покрова установлены отдельные тенденции изменения эдафических условий. Наиболее выраженные изменения морфологических свойств почв отмечали в верхних органогенных горизонтах, минеральная толща более инертна к пространственной неоднородности растений напочвенного покрова. Изменение мощности лесных подстилок происходит в ряду лишайниковая < зеленомошная < черничная, брусничная. Несмотря на то, что резкого изменения кислотно-основных показателей в почвах под различными микрогруппировками не выявили, тенденция возрастания значения рН отмечена в черничной микрогруппировке. Результаты математико-статистического анализа показали, что коэффициенты вариации содержания углерода в изучаемых почвах изменяются в небольшом диапазоне. Коэффициенты вариации биогенных элементов N, P, K изменяются в более широком диапазоне и составляют от 17 до 115%. Запасы С, N, Р, K в лесной подстилке значимо ниже в лишайниковой микрогруппировке по сравнению с другими растительными группировками. Кластерный анализ показал, что по совокупности исследуемых свойств выделяется органогенный горизонт лишайниковой микрогруппировки, а также подзолистый и иллювиальный горизонты черничной микроассоциации. Полученные данные могут быть использованы при организации природоохранных мероприятий, являются основой проведения мониторинга природной среды Восточной Фенноскандии.
Лесные экосистемы Восточной Фенноскандии представлены древостоями различного породного состава и возраста, они испытывают различный уровень антропогенного воздействия. Сосняки брусничные, составляющие более 65% от общего лесного фонда – наиболее распространенный на ее территории тип леса. Высокая адаптация соснового биогеоценоза (БГЦ) к различным экологическим и эдафическим условиям, а также антропогенному воздействию определяется биологическим разнообразием слагающих его компонентов. Последнее зависит от структуры древостоя, формирования специфических условий микросреды.
Растительный покров лесных экосистем достаточно сложно дифференцирован и представляет собой мозаику из комбинаций видов. Степень мозаичности фитоценозов определяется комплексом экотопических и биотопических факторов, таких как климат, структура древостоя, особенности микрорельефа и почвенного покрова и др. (Лебедева и др., 2005; Крышень, Федорец, 2006; Ипатов и др., 2014; Ипатов, 2013; Лебедева и др., 2015). Значительное влияние на гетерогенность напочвенного покрова оказывают фитогенные поля отдельных растений или групп растений, в результате взаимодействия которых в сообществе образуется особая среда, обеспечивающая стабильность и развитие фитокомплекса (Галанин, 1980; Крышень, 2000; Ипатов и др., 2009; Журавлева и др., 2012).
Сложность структуры напочвенного покрова в лесных фитоценозах отражается в разнообразии видов травяно-кустарничкового и мохово-лишайникового ярусов и их сочетании. Последнее формирует микрогруппировки, создающие неповторимость эдафо-фитоценотических микроусловий.
В естественных ненарушенных биогеоценозах существует четкое взаимодействие в системе “почва–растение–микробиота” (Карпачевский, 1977; Лукина и др., 2010). Свойства почв определяют температуру и гидрологический режим, плодородие, содержание элементов питания растений. Почва способна сглаживать низкие температуры зимой и оказывать влияние на флуктуации температуры в жаркий период, от ее свойств зависит развитие корневых систем растений. Она является средой обитания животных, микроорганизмов, которые осуществляют трансформацию органического вещества, круговорот веществ в биогеоценозе (Загуральская, Медведева, 2006).
Морфологические, физико-химические свойства почв, а также анализ растительных сообществ сосновых древостоев систематически описаны в работах российских и зарубежных ученых. Проведенные исследования лесных почв Мурманской области, Республики Коми с учетом различных тессер и микрогруппировок позволили получить представление о взаимосвязях между составом растительных сообществ и характеристиками плодородия почв (Лукина, Никонов, 1998; Орлова и др., 2003; Лукина и др., 2005; Елькина, 2008; Лукина и др., 2010; Лаптева, 2011; Шамрикова, 2013; Dymov, Gabov, 2015). В Карелии такие работы также проводились, однако недостаточно информации о вариабельности свойств альфе-гумусовых почв в условиях таежной зоны, хотя они играют глобальную роль в биологическом разнообразии лесных экосистем. В ранее проведенных исследованиях на заповедной территории Южной Карелии установлено изменение свойств почв, сформировавшихся на ленточных глинах, в зависимости от микрорельефа (Морозова и др., 2004). В работах Н.Г. Федорец с соавт. (1998) было показано изменение кислотно-основных свойств почв, сформировавшихся у ствола, в межкроновом и подкроновом пространстве сосняка брусничного. Изменение валового содержания азота и его различных соединений в зависимости от форм микрорельефа отмечено Н.Г. Федорец (Разнообразие почв …, 2006), изменение биологической активности почв, сформировавшихся в различных микрогруппировках соснового древостоя, было установлено Л.М. Загуральской, М.В. Медведевой (2006). В работах О.Н. Бахмет (2015) обоснована необходимость проведения исследования в данном направлении именно комплексным методом, предполагая изучение морфологических и физико-химических свойств почв.
Необходимо подчеркнуть, что целью настоящих исследований являлось изучение вариабельности свойств почв в отличие от работ, посвященных педоразнообразию. Последнее предполагает изучение таксономического разнообразия почв, изменение свойств почв внутри таксона (Горячкин, 2005). Проведенные исследования позволили получить информацию, которая может быть использована при мониторинге лесных экосистем в контексте глобального изменения климата и использования лесных земель. Исследование также станет основой при формировании общетеоретического представления о функционировании ненарушенных почв и научно-обоснованном управлении лесными экосистемами Восточной Фенноскандии.
ОБЪЕКТЫ И МЕТОДИКА
Методология исследования предполагала выполнение следующих основных условий. При закладке пробной площади учитывалось следующее: 1) участок длительное время не испытывал антропогенного воздействия; 2) тип леса был широко распространенным на территории Восточной Фенноскандии; 3) тип почвы и характер почвообразующей породы также являлись распространенными на данной территории; 4) морфометрические параметры и данные гранулометрического состава почвообразующих пород были однородны, что свидетельствовало бы об однородности исследуемых почв в ряду экотопа; 5) выполнение анализов почв проводили по апробированным методикам. Выполнение этих условий позволило получить ряд данных, результаты которых можно было корректно экстраполировать на аналогичные территории Фенноскандии.
Исследования проводили на одной из особо охраняемых природных территории Карелии – в государственном природном заповеднике “Кивач” (62°17′20.0 с.ш., 34°01′04.7″ в.д.). Закладку пробной площади размером 50 × 80 м (0.4 га) осуществляли в соответствии с общепринятыми в таксации правилами. На пробной площади производили сплошной перечет растущей части древостоя. Определение высот деревьев, возраста деревьев, оценку естественного возобновления производили на основании общепринятых принципов таксации (Анучин, 1982). Объем деревьев рассчитывали в соответствии с “Лесотаксационными таблицами” (1976), относительную полноту и класс бонитета – по справочнику (Третьяков и др., 1965).
В рассматриваемом фрагменте соснового древостоя описывали флору и определяли общее проективное покрытие травяно-кустарничкового и мохово-лишайникового ярусов, а также проективное покрытие каждого вида растения живого напочвенного покрова в отдельности. В пределах пробной площади по доминирующим видам, были выделены четыре микрогруппировки: лишайниковая, брусничная, черничная, зеленомошная. В пределах каждой микрогруппировки на квадратах площадью 1 × 1 м проводили геоботанические описания. Проективное покрытие видов сосудистых растений, мохообразных и лишайников оценивалось в процентах. Полученные данные приведены в табл. 1. Оценка покрытия каждого вида определялась как средняя величина от суммы всех описаний.
Таблица 1.
Состав и проективное покрытие видов живого напочвенного покрова в сосняке чернично-брусничном
| Таксоны | Проективное покрытие микрогруппировки, % | |||
|---|---|---|---|---|
| лишайниковая | брусничная | черничная | зеленомошная | |
| Calluna vulgaris (L.) Hull | 2 | – | + | + |
| Convallaria majalis L. | + | – | – | – |
| Hypopitys monotropa Crantz | – | + | – | – |
| Melampyrum pratense L. | + | – | + | – |
| Vaccinium myrtillus L. | 40 | 10 | 40 | 15 |
| V. vitis-idaea L. | + | 10 | 2 | 5 |
| Травяно-кустарничковый ярус | 40 | 20 | 40 | 15 |
| Dicranum cf. scoparium Hedw. | 2 | + | 5 | 10 |
| Hylocomium splendens (Hedw.) Schimp. in B.S.G. | + | – | 5 | 3 |
| Pleurozium schreberi (Bird.) Mitt. | 50 | 80 | 90 | 80 |
| Ptilium crista-castrensis (Hedw.) De Not. | + | – | 1 | – |
| Cetraria islandica (L.) Ach. | – | – | 2 | – |
| Cladonia arbuscula(Wallr.) Flot. | + | – | – | – |
| Cladonia rangiferina (L.) F. H. Wigg. | 10 | 7 | + | + |
| Мохово-лишайниковый ярус | 60 | 85 | 80 | 80 |
| Общее проективное покрытие | 90 | 85 | 90 | 85 |
На пробной площади в пределах выделенных растительных микрогруппировок закладывали почвенные разрезы. Морфологическое описание почв проводили с использованием общепринятой методики. Название почвы и индексы горизонтов даны согласно Классификации почв России (Шишов и др., 2004). Отбор почвенных образцов для физико-химического анализа производили из каждого генетического горизонта, смешанный образец брали из трех стенок разреза. Также в каждой микрогруппировке делали серию прикопок (N = 9), из которых также производили отбор образцов по горизонтам.
В отобранных образцах почв определяли гранулометрический состав и физико-химические показатели. В предварительно высушенных до воздушно-сухого состояния и просеянных через сито 2 мм образцах определяли кислотно-основные показатели, содержание органического вещества и элементов минерального питания (Аринушкина, 1970). Общее количество углерода определяли на СNH-анализаторе. Содержание подвижных соединений фосфора и калия определяли в одной вытяжке по методу Кирсанова: окончание анализа (K2О) – на атомно-абсорбционном спектрофотометре АА-7000 (Shimadzu, Япония), (Р2О5) – на фотоэлектроколориметре. Исследования выполнены на научном оборудовании Центра коллективного пользования ФИЦ “Карельский научный центр РАН”.
Статистическая обработка данных проведена с использованием пакетов программ Statistica 10, Microsoft Excel 2007. Взаимосвязи между растительностью, почвенными физическими и химическими свойствами определяли на основе корреляционного анализа. Кластерный анализ применяли для определения степени близости отдельных кластеров, последовательности их объединения/разделения.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Исследуемый древостой согласно лесной типологии В.Н. Сукачева (1972) относится к типу леса сосняк брусничный. Насаждение одноярусное, чистое по составу (10С). Возраст основной части деревьев составляет 190 лет, также произрастают деревья возрастом 220 лет. Сумма площадей сечений деревьев не превышает 28 м2 га–1, относительная полнота – 0.79. Численность растущих деревьев на пробной площади составляла 375 экз. на 1 га, их средний диаметр – 31.0 см, средняя высота – 23.1 м, при этом отдельные деревья достигали 50 см по диаметру и 26–27 м в высоту. Класс бонитета III.4. Согласно таксационной характеристике древостоя запас стволовой древесины составляет 297 м3 га–1.
Все исследованные растительные микрогруппировки отличаются бедностью видового состава: в травяно-кустарничковом ярусе выявлены 6 видов, в мохово-лишайниковом – 7 видов, что является характерным для данного типа леса. В составе травяно-кустарничкового яруса во всех микрогруппировках преобладают черника и брусника, покрытие остальных видов сосудистых растений вне зависимости от типа микрогруппировки очень мало. Мохово-лишайниковый ярус в микрогруппировках развит более равномерно и имеет высокий показатель обилия.
Профиль исследуемых почв представляет собой систему горизонтов О-Е-BF-BF2-ВС-С. Почвы диагностированы как подзолы иллювиально-железистые песчаные на водно-ледниковых отложениях. Данные морфологических и физических свойств репрезентативных профилей почв, сформировавшихся на песчаных отложениях, представлены в табл. 2 и табл. 3. Ведущими почвообразовательными процессами на исследуемом участке являются подстилконакопление, подзолообразование, иллювиирование. Верхний диагностический органогенный горизонт почв – лесная подстилка (Organic), формируется в лесных экосистемах в результате медленной минерализации органического вещества, имеет небольшую мощность. Диагностическими горизонтами являются подзолистый (Podzol) и иллювиальный (Spodic), мощность которых небольшая и составляет до 3 и 5 см соответственно. Выявленная комбинация диагностических горизонтов почв позволяет их отнести к подзолам иллювиально-железистым песчаным.
Таблица 2.
Морфологическое строение альфе-гумусовых подзолов, сформировавшихся в различных условиях фитоценотической среды
| Горизонт почв |
Глубина, см |
Гранулометрический состав/степень разложения | Цвет | Плотность сложения | Особенности |
|---|---|---|---|---|---|
| Лишайниковая микрогруппировка | |||||
| OL | 0–1 | н | 5YR7/6 | р | Темно-бурого цвета, рыхлая |
| OF | 1–2 | с | 5YR4/1 | р | Мало корней |
| OH | 2–2.5(3) | х | 5YR2/1 | р | Встречаются угли и небольшая минеральная примесь |
| E | 2.5(3)–4(5) | п | 10YR8/1 | р | В верхней части потеки гумуса, встречаются угли |
| BF | 4(5)–18(22) | п | 7.5YR3/4 | у | – |
| B2 | 18(22)–30 | п | 7.5YR4/3 | у | Основная масса корней сосредоточена в верхней части |
| В3 | 30–42 | п | 7.5YR7/1 | у | – |
| В4 | 42–60 | п | 7.5YR7/2 | п | По всему горизонту ржавые пятна |
| ВС | 60–90 | с | 7.5YR8/3 | п | В средней части ржавые пятна |
| С | 90–120 | с | 7.5YR4/6 | п | Ржавые и черные пятна по всему горизонту |
| Брусничная микрогруппировка | |||||
| OL | 0–3 | н | 5YR7/6 | р | – |
| OF | 3–5 | с | 5YR4/1 | р | Мало корней |
| OH | 5–6 | х | 5YR2/1 | р | Угли, небольшая минеральная примесь |
| E | 6–8(13) | п | 10YR8/1 | р | В верхней части угли, мало корней |
| BF | 8(13)– 14(30) | п | 5YR4/2 | р | Густо пронизан корнями |
| B2 | 14(30)–30 | п | 5YR5/3 | у | – |
| В3 | 30–37(42) | п | 5YR6/1 | р | – |
| В4 | 37(42)–57 | п | 5YR7/1 | р | По всему горизонту ржавые вертикальные потеки |
| ВС | 57–94 | п | 5YR7/2 | р | Ржавые и черные вертикальные полосы по всему горизонту |
| С | 94–150 | п | 5YR8/2 | р | Ржавые пятна по всему горизонту |
| Черничная микрогруппировка | |||||
| OL | 0–2 | н | 5YR7/6 | р | – |
| OF | 2–5 | с | 5YR4/1 | р | Мало корней |
| OH | 5–6(8) | х | 5YR2/1 | р | Угли, небольшая минеральная примесь |
| E | 6(8)–7(9) | п | 10YR8/1 | у | В верхней части встречаются угли |
| BF | 7(9)–14(16) | п | 7.5YR3/4 | у | – |
| B2 | 14(16)–27(30) | п | 7.5YR5/4 | п | – |
| В3 | 27(30)–36(44) | п | 7.5YR7/1 | п | – |
| В4 | 36(44)–52(56) | п | 7.5YR6/2 | п | В верхней части небольшое количество корней, ржавые пятна по всему горизонту |
| ВС | 52(56)–80 | п | 7.5YR8/2 | у | Ржавые пятна по всему горизонту |
| С | 80–115 | п | 7.5YR8/1 | у | Ржавые вертикальные полосы по всему горизонту |
| Зеленомошная микрогруппировка | |||||
| OL | 0–1 | н | 5YR7/6 | р | – |
| OF | 1–2.5 | с | 5YR4/1 | р | Мало корней |
| OH | 2.5–3 | х | 5YR2/1 | р | Угли, небольшая минеральная примесь |
| E | 3–5(9) | п | 10YR8/1 | р | В верхней части встречаются угли |
| BF | 5(9)–10(15) | п | 7.5YR3/3 | у | – |
| B2 | 10(15)–20 | п | 7.5YR4/3 | р | – |
| В3 | 20–40 | п | 7.5YR5/4 | у | В верхней части встречаются корни |
| В4 | 40–60(65) | п | 7.5YR8/2 | р | – |
| ВС | 60(65)–110 | с | 7.5YR8/1 | у | Встречается мелкий гравий |
| С | 110–185 | п | 7.5YR7/2 | р | Плохо сортированный песок, в нижней части ржавые полосы |
Таблица 3.
Показатели биогеоценозной вариабельности физико-химических показателей альфе-гумусовых почв
| Гори- зонт |
Показатель | pH | V | C | N | C : N | K2O | P2O5 | |
|---|---|---|---|---|---|---|---|---|---|
| H2O | KCl | % | мг × 100 г–1 | ||||||
| Лишайниковая микрогруппировка | |||||||||
| О | Min–max | 4.0–4.5 | 3.0–3.4 | 0.2–13.7 | 19.5–48.2 | 0.5–1.1 | 34.0–57.5 | 34.0–94.1 | 9.4–28.6 |
| X ± SD | 4.3 ± 0.2 а | 3.2 ± 0.1 а | 5.9 ± 5.6 а | 40.5 ± 8.8 а | 0.8 ± 0.2 а | 41.4 ± 7.0 а | 81.9 ± 18.7 а | 18.5 ± 8.7 а | |
| CV, % | 4 | 3 | – | 22 | 22 | 17 | 23 | 47 | |
| Е | Min–max | 3.8–4.7 | 3.0–4.8 | 2.4–11.6 | 0.7–1.4 | 0.02–0.05 | 14.9–36.7 | 1.7–3.8 | 0.8–5.39 |
| X ± SD | 4.2 ± 0.4 а' | 3.7 ± 0.3 а' | 7.21 ± 2.8 а' | 1.0 ± 0.2 а' | 0.03 ± 0.01 а' | 26.1+8.0 а' | 2.4 ± 0.6 а' | 3.1 ± 1.4 b' | |
| CV, % | 8 | 15 | – | 23 | 30 | 31 | 25 | 45 | |
| BF | Min–max | 4.2–5.1 | 3.7–4.7 | 3.56–46.8 | 0.5–1.0 | 0.01–0.06 | 9.2–49.7 | 0.8–2.1 | 1.8–47.0 |
| X ± SD | 4.7 ± 0.3 а'' | 4.3 ± 0.3 а'' | 15.1 ± 16.6 b'' | 0.7 ± 0.2 аb'' | 0.04 ± 0.01 а'' | 17.9 ± 12.5 а'' | 1.3 ± 0.4 а'' | 21.1 ± 14.1 а'' | |
| CV, % | 6 | 7 | – | 25 | 35 | 70 | 33 | 67 | |
| Брусничная микрогруппировка | |||||||||
| О | Min–max | 4.2–4.6 | 2.9–3.5 | 0.4–13.9 | 36.5–49.3 | 0.7–1.1 | 32.3–49.1 | 88.1–158.6 | 19.4–100.9 |
| X ± SD | 4.3 ± 0.2 а | 3.1 ± 0.2 а | 7.2 ± 5.0 а | 43.7 ± 4.0 а | 0.9 ± 0.2 а | 40.9 ± 5.4 а | 126.3 ± 23.5 а | 45.6 ± 25.1 b | |
| CV, % | 3 | 5 | – | 9 | 17 | 13 | 19 | 55 | |
| Е | Min–max | 4.0–4.5 | 2.9–3.7 | 2.7–13.2 | 0.6–1.5 а' | 0.01–0.04 | 15.3–88.8 | 1.5–19.3 | 0.7–2.7 |
| X ± SD | 4.2 ± 0.2 а' | 3.2 ± 0.2 а' | 6.01 ± 4.2 а' | 0.9 ± 0.3 | 0.02 ± 0.01 а' | 38.1 ± 23.4 а' | 8.2 ± 7.6 а' | 1.1 ± 0.6 а' | |
| CV, % | 4 | 7 | – | 33 | 42 | 61 | 93 | 58 | |
| BF | Min–max | 4.7–5.0 | 4.0–4.6 | 4.4–7.1 | 0.6–1.5 аb'' | 0.01–0.04 | 13.5–49.0 | 0.8–12.0 | 10.0–29.6 |
| X ± SD | 4.8 ± 0.1 а'' | 4.3 ± 0.2 а'' | 5.6 ± 1.0 а'' | 0.6 ± 0.1 | 0.02 ± 0.01 b'' | 32.5 ± 13.0 b'' | 6.2 ± 5.0 аb'' | 19.1 ± 6.2 а'' | |
| CV, % | 3 | 4 | – | 12 | 45 | 40 | 80 | 32 | |
| Черничная микрогруппировка | |||||||||
| О | Min–max | 3.1–4.6 | 2.9–4.4 | 1.3–11.3 | 33.5–51.7 | 0.6–1.0 | 38.9–63.6 | 15.2–159.1 | 19.6–40.4 |
| X ± SD | 4.0 ± 0.6 а | 3.5 ± 0.6 а | 5.9 ± 3.0 а | 45.2 ± 6.0 а | 0.8 ± 0.02 а | 46.7 ± 7.3 а | 117.6 ± 42.3 а | 34.3 ± 6.4 b | |
| CV, % | 14 | 18 | – | 13 | 17 | 16 | 36 | 19 | |
| Е | Min–max | 3.1–4.6 | 3.0–4.3 | 1.4–6.7 | 0.7–1.2 | 0.01–0.08 | 9.5–82.7 | 1.0–104.9 | 0.6–4.5 |
| X ± SD | 4.0 ± 0.6 а' | 3.5 ± 0.6 а' | 3.9 ± 1.9 а' | 0.9 ± 0.2 а' | 0.03 ± 0.02 а' | 36.2 ± 22.4 а' | 37.7 ± 29.1 а' | 1.3 ± 1.2 а' | |
| CV, % | 16 | 17 | – | 17 | 68 | 62 | 77 | 96 | |
| BF | Min–max | 4.5–5.3 | 3.8–5.1 | 3.8–7.6 | 0.6–1.0 | 0.01–0.03 | 19.9–66.6 b'' | 8.9–23.6 | 15.1–40.9 |
| X ± SD | 4.9 ± 0.3 а'' | 4.5 ± 0.4 а'' | 5.6 ± 1.3 а'' | 0.8 ± 0.1 b'' | 0.02 ± 0.01 b'' | 35.3 ± 16.3 | 13.1 ± 4.5 b'' | 31.1 ± 8.8 а'' | |
| CV, % | 6 | 8 | – | 17 | 27 | 45 | 34 | 28 | |
| Зеленомошная микрогруппировка | |||||||||
| O | Min–max | 4.2–4.7 | 3.0–3.3 | 0.2–11.3 | 39.7–52.4 | 0.8–1.2 | 36.1–57.3 | 69.7–184.2 | 8.5–38.8 |
| X ± SD | 4.3 ± 0.2 а | 3.1 ± 0.2 а | 3.9 ± 4.0 а | 48.2 ± 7.8 а | 1.0 ± 0.2 а | 43.8 ± 5.7 а | 111.3 ± 36.6 а | 19.0 ± 9.5 а | |
| CV, % | 4 | 8 | – | 10 | 15 | 13 | 33 | 50 | |
| E | Min–max | 3.8–4.6 | 3.0–3.3 | 1.6–6.7 | 0.3–1.3 | 0.03–0.1 | 4.3–23.3 | 0.8–4.0 | 0.2–3.3 |
| X ± SD | 4.3 ± 0.2 а' | 3.1 ± 0.1 а' | 4.0 ± 1.9 а' | 0.7 ± 0.3 а' | 0.05 ± 0.02 b' | 15.1 ± 6.8 b' | 1.85 ± 1.01 а' | 0.8 ± 0.8 а' | |
| CV, % | 6 | 3 | – | 43 | 46 | 45 | 55 | 115 | |
| BF | Min–max | 4.3–5.1 | 3.2–4.8 | 2.4–9.4 | 0.2–0.8 | 0.03–0.05 | 5.9–20.2 | 0.6–1.9 | 9.2–28.5 |
| X ± SD | 4.9 ± 0.3 а'' | 4.4 ± 0.5 а'' | 4.9 ± 2.3 а'' | 0.5 ± 0.2 а'' | 0.04 ± 0.02 а'' | 12.1 ± 4.0 а'' | 1.1 ± 0.4 аb'' | 18.8+6.2 а'' | |
| CV, % | 6 | 11 | – | 31 | 16 | 33 | 35 | 33 | |
Примечание. V – степень насыщенности почв основаниями; “–” – нет сведений; min – минимальное значение; max – максимальное значение; X – среднее значение; SD – стандартное отклонение; CV – коэффициент вариации. Индексами "а”, “b” показаны статистически достоверные различия (P < 0.05) для горизонта О, “а'", "b'" – для горизонта Е, “а''”, “b''”, “аb''”” для горизонта BF.
Небольшие различия в морфологических свойствах почв отмечены в верхних корнеобитаемых горизонтах. Выявлено уменьшение мощности лесной подстилки (О горизонт) в ряду: зеленомошная < лишайниковая < черничная < брусничная микрогруппировки. Подзолистый (Е) и иллювиальный (BF) горизонты в изучаемых растительных микрогруппировках имели различную окраску, что косвенно свидетельствует о разной интенсивности почвообразовательного процесса.
По гранулометрическому составу почвы отнесены к песчаным разновидностям. В результате педогенных процессов происходило накопление крупнопесчаных фракций в нижних горизонтах почв. Содержание физического песка и физической глины в почвах небольшое.
Кислотность верхних горизонтов почв высокая и составляет до 3.5, нижних – несколько выше и достигает до 5. Результаты анализа химических свойств почв показали, что резкое изменение кислотно-основных свойств почв в различных микрогруппировках отсутствует. В целом прослеживается тенденция более низких показателей кислотности в черничной микрогруппировке. Это обусловлено химическим составом опада черники, в которой содержится больше кальция, поэтому и кислотность ее по сравнению с брусникой ниже (Лукина, Никонов, 1996; Ingestad, 2006). Более высокая кислотность органогенных горизонтов почв сосновых парцелл по сравнению с межкроновыми лишайниковыми и зеленомошными связана с высоким содержанием кислого и грубого гумуса под кронами (Лукина и др., 2010).
Содержание С и N, их распределение по профилю почв является характерным для почв данного генезиса: наиболее высокое содержание в верхнем горизонте, в глубь почвенной толщи оно резко уменьшается. В иллювиальном горизонте содержание углерода и азота невысокое (<3%), что свидетельствует о невысоком плодородии почв, низкой продуктивности древостоя. Это связано, во-первых, с тем, что основу органогенного горизонта почв составляет “кислый” опад сосновых древостоев: хвоя, корни сосны, листья вечнозеленых кустарничков, мхи, а также талломы лишайников, трансформация которых происходит замедленно. Также это обусловлено тем, что органические и минеральные вещества, образуемые в процессе трансформации опада, не задерживаются в верхнем горизонте почв, а перемещаются в нижележащие. Однако в минеральной толще, имеющей песчаный состав, не происходит их аккумуляции, они быстро перехватываются корнями растений. Подтверждением этому является невысокое содержание органического вещества в верхнем горизонте почв и минеральной толще. В работах A.S. Koch и E. Matzner (1993) отмечено отсутствие видимых изменений в содержании азота и углерода в почвах в зависимости от расстояния от ствола дерева. При этом необходимо отметить, что разложение хвойного опада растений идет медленно, поэтому отношение С : N в почве широкое.
Степень насыщенности основаниями верхних и нижележащих горизонтов почв свидетельствует о невысоком выветривании литогенной основы, вовлечении элементов в природный круговорот веществ. Степень насыщенности основаниями в различных горизонтах исследованных почв под различными растительными микроассоциациями резко не меняется.
Резкого изменения в содержании доступного растениям фосфора и калия в различных микрогруппировках также не выявили, распределение их в почвах носит биогенно-аккумулятивный характер. Содержание фосфора и калия более высокое в почвах, сформировавшихся под черничной и зеленомошной микрогруппировками. Это, на наш взгляд, отражает тесную генетическую связь почв и растений и обусловлено комплексом факторов, ведущими, из которых являются химический состав растений (накопление калия, фосфора отмечается во мхах (Klavina et al., 2012)), а также ролью данных элементов в постсинтетических процессах. Известно, что калий участвует в белковом и углеводном обмене, синтезе аскорбиновой кислоты, содержание которой в чернике высокое. Также калий участвует в усвоении углекислого газа, образование которого в моховой микрогруппировке высокое.
Почвы зеленомошных и, особенно, сфагновых микрогруппировок отличаются высоким содержанием алюминия и железа в органогенных горизонтах, так как могут быть аккумуляторами этих элементов (Лукина и др., 2010).
Для анализа однородности совокупности рассчитывали коэффициент вариации. Анализ показал, что коэффициент вариации показателей физико-химических свойств почв изменяется в широком пределе (4–115%). Однако выявлено, что для кислотно-щелочных свойств изменение коэффициента вариации происходит в небольших пределах и составляет 4–17%. Коэффициент вариации содержания углерода также изменяется в небольшом диапазоне и соответствует литературным данным (Рыжова, Подвезенная, 2008; Рыжова и др., 2020). Коэффициент вариации биогенных элементов N, P, K изменяется в широком диапазоне и составляет от 17 до 115%. При этом более высокие показатели изменчивости содержания биогенных элементов в почве отмечены для подвижных форм K и Р.
На вариабельность свойств почв может оказывать влияние тип растительной микрогруппировки. Наши результаты показали, что прослеживается тенденция наименее выраженной пространственной вариабельности свойств почв в горизонте О, сформировавшемся под лишайниковой растительной микрогруппировкой. И это следовало ожидать, так как мощность подстилки в лишайниковой микрогруппировке меньше, формируются более сухие эдафические условия, отсутствуют или малочисленны корневые системы вечнозеленых кустарничков, таллом лишайников расположен на поверхности минеральной толщи и не проникает в нее. О более равномерном развитии растений мохово-лишайникового яруса отмечено выше.
Дисперсионный анализ показал, что статистически достоверные различия средних показателей отмечены по содержанию подвижного фосфора в лесной подстилке (P = 0.0006), углерода, общего азота, подвижного калия в горизонте BF для почв исследуемых микрогруппировок (P < 0.001), а также по содержанию азота (P = 0.04) и подвижного фосфора (P < 0.001) в горизонте Е. Это во многом связано с выраженностью подзолообразовательного процесса, который является ведущим процессом почвообразования в среднетаежной подзоне Карелии и в зависимости от микрогруппировок проявляется по-разному (см. морфологическое описание почв, табл. 1).
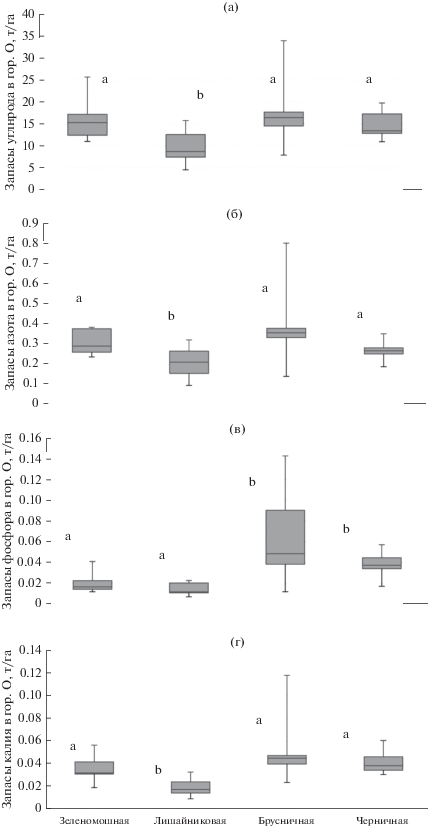
Как известно, информативным показателем продуктивности лесных почв является определение запасов биогенных элементов лесной подстилке. В этой связи делали перерасчет содержания элементов – биогенов в горизонте О с учетом его мощности и объемного веса (рис. 1). Статистический анализ показал, что запасы С, N, Р, K в лесной подстилке значимо ниже в лишайниковой микрогруппировке по сравнению с другими (P = 0.01 для С, N; P = 0.002 для Р, K). Полученные данные подтверждают специфические особенности верхней части органопрофиля почв, сформировавшегося в различных микрогруппировках.
Рис. 1.
Варьирование запасов углерода (а), общего азота (б), подвижного фосфора (в) и калия (г) в т/га в лесной подстилке альфе-гумусовых почв в зависимости от растительной микрогруппировки. Индексами “а”, “b” показаны статистически достоверные различия (P < 0.05).
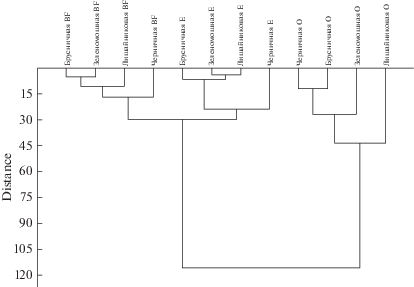
Для выделения пространственно однородных групп в парцеллярной структуре исследованного древостоя применялся кластерный анализ (рис. 2). Построенная дендрограмма показывает степень близости изучаемых объектов, а также наглядно демонстрирует в графическом виде последовательность их объединения/разделения. Визуальный анализ дендрограммы позволил установить, что наиболее высокую степень различия между собой имеют органогенный и минеральные горизонты (E, BF), образующие два отдельных кластера. В органогенном горизонте выделяется лишайниковая микрогруппировка, что подтверждает вышесказанное. В подзолистом и иллювиальном горизонтах наибольшее различие свойств проявляется между черничной и зеленомошно-бруснично-лишайниковыми растительными микрогруппировками. В целом порог сходства почвенных горизонтов различных парцелл довольно низкий.
Рис. 2.
Дендрограмма сходства физико-химических свойств почв, расположенных в различных растительных микрогруппировках (лишайниковая, черничная, брусничная, зеленомошная), сосняка брусничного. Индексами О, Е, ВF обозначены горизонты почв: О – органогенный, Е – элювиальный, ВF – иллювиальный.
Сравнительный анализ свойств почв, сформировавшихся в различных растительных микрогруппировках, позволил вывить сходства и различия. Прежде всего отмечали, что согласно таксационным показателям продуктивность исследуемого древостоя несколько выше типичного сосняка брусничного IV класса бонитета в условиях Карелии (Козлов, 1985). Возможно, это связано с пирогенным прошлым данного участка: угли, представляющие собой концентраты элементов минерального питания растений, найденные в почве (табл. 2), сосновый древостой использует на начальном этапе развития, вследствие чего его продукционная способность может увеличиваться. Это может оказывать прямо или косвенно влияние на эдафо-фитоценотические свойства всего биогеоценоза (Järvinen et аl., 1993).
В зависимости от доминирующих видов в травяно-кустарничковом и мохово-лишайниковом ярусе выделяют отдельные ценобиотические микрогруппировки, которые под воздействием природных и антропогенных факторов способны сменять друг друга (Лукина и др., 2010). В чистых лесных насаждениях весь БГЦ может состоять практически из одной растительной микрогруппировки, тогда как в смешанных насаждениях их может быть более 20 (Карпачевский, 1977). Ранее проведенные исследования лесов заповедника “Кивач” показали, что сосняки брусничные характеризуются однородностью со сравнительно высокомозаичной структурой напочвенного покрова, в которой выделено 4 типа растительных микрогруппировок (Крышень и др., 2006). На исследуемом участке мы также выделили четыре микрогруппировки. При этом граница перехода между лишайниковой и остальными (брусничной, черничной, зеленомошной) хорошо диагностировалась по растениям напочвенного покрова, мощности лесной подстилки и подзолистого горизонта Е. Расположение микрогруппировок является типичным для данного леса, когда в условиях микроповышения формируются лишайниково-брусничные локусы, в более гидроморфных условиях – чернично-зеленомошные.
Как известно, экология растений-доминантов растительных микрогруппировок различная. Лишайники, к примеру, способны длительно существовать в условиях недостатка влаги и использовать элементы питания из атмосферы, могут получить жизненно важные компоненты с помощью других микроорганизмов, присутствующих как на их слоевищах, так и в окружающей среде (Cardinale et al., 2012; Sigurbjörnsdóttir et al., 2016; Spribille et al., 2016). Мхи рода дикранум предпочитают мезоморфные условия, могут аккумулировать влагу и поэтому также способны длительно переносить неблагоприятные гидрологические условия.
Влияние микрогруппировок растений на свойства почв может быть связано и с различным перехватыванием ими элементов питания из атмосферных осадков и аэрозолей (Bergkvist et al., 1989; Levia, Frost, 2003). Можно также отметить, что растения в различных микрогруппировках по-разному могут перехватывать элементы из атмосферы. Следовательно, их сток через листву в почву также может быть неодинаковым (Levia, Frost, 2003). Напочвенный покров также способствует изменению гидротермических свойств почв (Коронатова, Миляева, 2010).
Проведенные результаты показали, что влияние растительных микрогруппировок в сосновом древостое на пространственную изменчивость почвенных свойств затрагивает только верхние горизонты и сильно ослабевает с глубиной (Разнообразие почв …, 2006). Преобладание крупнопесчаных фракций в гранулометрическом составе почв не обеспечивает задержку, закрепление и осаждение коллоидных частиц в иллювиальном горизонте, а, напротив, способствует фронтальному глубокому току почвенных вод, что в целом может нивелировать различия, которые проявляются между различными горизонтами. Это согласуется с результатами М.А. Подвезенной, И.М Рыжовой (2010, 2011) отмечавшими, что от типа растительной микрогруппировки зависят величина и пространственная изменчивость запасов подстилки, содержания и запасов углерода только в самом верхнем минеральном горизонте почвы мощностью 5 см. В более глубоких горизонтах эта зависимость не прослеживается, так как она может затушевываться совокупным действием других факторов.
Интересную гипотезу об изменении вариабельности почв и разнообразии растений предложил D. Tilman (1987). По его мнению, на почвах с высокой вариабельностью свойств создается больше оптимальных условий для растений, поэтому разнообразие их выше. Однако эффект почвенной вариабельности на рост биоразнообразия более высокий на почвах с низким содержанием элементов питания. Это связано с тем, что в условиях с высоким фоновым содержанием элементов питания отклик растений на увеличение плодородия почв проявляется слабее (Tilman, 1987).
В проведенном исследовании нами установлено, что мощность верхнего органогенного горизонта почв наибольшая именно в чернично-брусничных парцеллах, в то время как в лишайниковой и зеленомошной мощность лесной подстилки ниже. Мощность подзолистого горизонта почв, сформировавшихся в различных микрогруппировах, значимо различалась и находилась в минимуме в лишайниковой микрогруппировке. T. Hokkanen с соавт. (1995) показали, что изменение свойств почв в различных микрозонах происходит в наибольшей степени в верхних горизонтах и мало выражено в нижележащих горизонтах почв. Возможно, это связано с влиянием фитогенного поля деревьев и распределением их корневых систем. В свою очередь корненасыщенность верхних горизонтов почв может существенно различаться в зависимости от микроусловий педосреды и, следовательно, влиять на гетерогенность свойств почв (Сенов и др., 1994). Лишайники могут выступать в качестве генераторов экологических ниш для бактериальных сообществ, как внутри таллия, так и в субстрате под ним (Bjelland et al., 2011), продукты их метаболизма могут структурировать даже почвенные бактериальные сообщества, присутствующие под ними (Maier et al., 2014).
ЗАКЛЮЧЕНИЕ
На основании анализа данных были изучены особенности взаимного влияния эдафических и биотических компонентов природной среды. Установлены три основных блока, через которые осуществляется данное взаимодействие. Первый включает в себя древостой и растения напочвенного покрова, которые являются поставщиками различных субстратов в почву, оказывают стимулирующее/ингибирующее влияние на биоту. Они прямо или опосредованно участвуют в формировании органогенного горизонта почв, создают условия для адгезии клеток микроорганизмов, поддерживают пул элементов-биофилов. Вторым важным компонентом этих взаимодействий является верхний органогенный горизонт почв (лесная подстилка). Она определяет микрозональность происходящих в почве процессов, гидротермические условия развития микробиоты. Влияние органогенного горизонта на растения может быть прямым: гумусовые вещества обеспечивают питанием корневую системы, создают условия для формирования микоризообразователей, а также косвенным. Последнее происходит путем регулирования круговорота питательных веществ, а также через микробиотический компонент экотопа. Третий блок предполагает участие минеральной толщи в формировании эдафо-фитоценотических взаимодействий. В результате установления консортивных связей между микробиотой, корневыми системами и почвенными частицами формируются органоминеральные “узлы”, через которые проходят основные взаимодействия. Таким образом, при совместной работе трех главных составляющих формируется единый каркас, взаимодействия в котором происходят в горизонтальном и вертикальном направлениях, осуществляются “масса–перенос” энергии и вещества, циркуляция питательных соединений. На таком этапе развития экосистема наиболее устойчива к неблагоприятным факторам среды, обеспечивает надежный пространственно-временной континуум почв и почвенного покрова.
Механизмы влияния растений на свойства почв до конца не выяснены, поэтому их необходимо продолжать исследовать. Очевидно, что действие растений на почвы и почвы на растение взаимно. При этом можно отметить, что необходимо изучать не только изменение свойств почв в центральной части микрогруппировок, а также на границе их раздела. Последнее является необходимым именно для познания формирований причинно-следственных связей в такой сложной системе, какой является почва.
Список литературы
Анучин Н.П. Лесная таксация: Учебник для ВУЗов. 5‑е изд. М.: Лесная промышленность, 1982. 552 с.
Аринушкина Е.В. Руководство по химическому анализу почвы: Уч. пособие для ВУЗов по спец.: “Агрохимия и почвоведение”. М.: Изд-во Московского университета, 1970. 487 с.
Бахмет О.Н. Структурно-функциональная организация органопрофилей почв лесных экосистем Северо-Запада России: Автореф. дис. … докт. биол. наук: 03.02.08. Петрозаводск: Изд-во КарНЦ РАН, 2015. 49 с.
Галанин А.В. Влияние фитогенного поля на распределение растений // Экология. 1980. № 6. С. 76–78.
Горячкин С.В. Исследование структур почвенного покрова в современном почвоведении: подходы и тенденции развития // Почвоведение. 2005. № 12. С. 1461–1468.
Елькина Г.Я. Оптимизация минерального питания растений на подзолистых почвах. Екатеринбург: Изд-во УрО РАН, 2008. 280 с.
Журавлева Е.Н., Ипатов В.С., Лебедева В.Х., Тиходеева М.Ю. Изменение растительности на лугах под влиянием сосны обыкновенной (Pinus sylvestris L.) // Вестник Санкт-Петербургского университета. Серия 3: Биология. 2012. № 2. С. 3–12.
Загуральская Л.М., Медведева М.В. Микробное разнообразие почв хвойных и лиственных лесов // Разнообразие почв и биоразнообразие в лесных экосистемах средней тайги. М.: Наука, 2006. С. 228–234.
Ипатов В.С. Вновь о понятии “фитоценоз” // Ботанический журн. 2013. Т. 98. № 4. С. 481–486.
Ипатов В.С., Лебедева В.Х., Тиходеева М.Ю. О гетерогенности и квантованности растительности пробных площадей // Ботанический журн. 2014. Т. 99. № 1. С. 3–22.
Ипатов В.С., Журавлева Е.Н., Лебедева В.Х., Тиходеева М.Ю. Фитогенное поле Picea abies, P. obovata (Pinaceae) // Ботанический журн. 2009. Т. 94. № 4. С. 558–568.
Карпачевский Л.О. Пестрота почвенного покрова в лесном биогеоценозе. М.: Изд-во МГУ, 1977. 312 с.
Козлов И.Ф. Развитие и рост сосновых насаждений Карельской АССР. Петрозаводск: Изд-во Карельского филиала АН СССР, 1985. 162 с.
Коронатова Н.Г., Миляева Е.В. Влияние растительности на физические свойства молодых почв выработанных карьеров // Интерэкспо Гео-Сибирь. 2010. № 2. С. 162–166.
Крышень А.М. Фитогенное поле: теория и проявление в природе // Известия РАН. Серия Биология. 2000. № 4. С. 437–443.
Крышень A.M., Федорец Н.Г. Взаимосвязь зональных типов леса с почвами // Разнообразие почв и биоразнообразие в лесных экосистемах средней тайги. М.: Наука, 2006. С. 200–207.
Крышень А.М., Рудковская О.А., Преснухин Ю.В., Тимофеева В.В. Морфоструктура напочвенного покрова основных типов лесных сообществ заповедника “Кивач” (средняя тайга) // Труды Карельского НЦ РАН. 2006. Вып. 10. С. 54–62.
Лаптева Е.М. Отдел почвоведения: итоги и перспективы // Вестник Института биологии Коми НЦ УрО РАН. 2011. № 12. С.4–12.
Лебедева В.Х., Ипатов В.С., Тиходеева М.Ю. Неоднородность пространственной структуры живого напочвенного покрова в лесных сообществах // Вестник Санкт-Петербургского гос. унивеситета, 2015. Серия 3. Вып. 2. С. 32–46.
Лебедева В.Х., Тиходеева М.Ю., Ипатов В.С. Влияние древесного полога на виды напочвенного покрова в ельнике чернично-зеленомошном // Ботанический журн. 2005. Т. 90. № 3. С. 400–410.
Лесотаксационные таблицы. Петрозаводск: Карельское лесоустроительное предприятие; Институт леса Карельского Филиала АН СССР. 1976. 32 с.
Лукина Н.В., Никонов В.В. Биогеохимические циклы в лесах Севера в условиях аэротехногенного загрязнения. Апатиты: Изд-во Кольского НЦ РАН, 1996. Ч. 1. 213 с.
Лукина Н.В., Никонов В.В. Питательный режим лесов северной тайги: природные и техногенные аспекты. Апатиты: Изд-во Кольского НЦ РАН, 1998. 316 с.
Лукина Н.В., Орлова М.А., Исаева Л.Г. Плодородие лесных почв как основа взаимосвязи почва-растительность // Лесоведение. 2010. № 5. С. 45–56.
Лукина Н.В., Сухарева Т.А., Исаева Л.Г. Техногенные дигрессии и восстановительные сукцессии в северотаежных лесах. М.: Наука, 2005. 245 с.
Морозова Р.М., Федорец Н.Г., Бахмет О.Н. Почвы и почвенный покров Заонежья Карелии // Труды Карельского НЦ РАН. 2004. Вып. 6. С. 69–89.
Орлова М.А., Лукина Н.В., Никонов В.В. Влияние ели на пространственную изменчивость кислотности подзолов северотаежных лесов // Лесоведение. 2003. № 6. С. 3–11.
Подвезенная М.А., Рыжова И.М. Зависимость вариабельности запасов углерода в почве от пространственной структуры растительного покрова лесных биогеоценозов // Вестник Московского университета. Серия 17. Почвоведение. 2010. № 4. С. 3–9.
Подвезенная М.А., Рыжова И.М. Изменчивость содержания и запасов углерода в почвах лесных биогеоценозов южной тайги // Лесоведение. № 1. 2011. С. 52–60.
Разнообразие почв и биоразнообразие в лесных экосистемах средней тайги / Отв. ред. Н.Г. Федорец. М.: Наука, 2006. 287 с.
Рыжова И.М., Подвезенная М.А. Пространственная вариабельность запасов органического углерода в почвах лесных и степных биогеоценозов // Почвоведение. 2008. № 12. С. 1429–1437
Рыжова И.М., Телеснина В.М., Ситникова А.А. Динамика свойств почв и структуры запасов углерода в постагрогенных экосистемах в процессе естественного лесовосстановления // Почвоведение. 2020. № 2. С. 230–243.
Сенов С.Н., Банева Н.А., Смирнов А.П. Корненасыщенность и парцеллярная структура в высокопроизводительном ельнике // Лесоведение. 1994. № 2. С. 78–80.
Сукачёв В.Н. Избранные труды в трех томах / Под ред. Е.М. Лавренко. Л.: Наука, 1972. Т. 1. Основы лесной типологии и биогеоценологии. 419 с.
Третьяков Н.В., Горский П.В., Самойлович Г.Г. Справочник таксатора (таблицы для таксации леса). 2-е изд. М.: Лесная промышленность, 1965. 459 с.
Федорец Н.Г., Дьяконов В.В., Литинский П.Ю., Шильцова Г.В. Загрязнение лесной территории Карелии тяжелыми металлами и серой. Петрозаводск: Карельский НЦ РАН, 1998. 47 с.
Шамрикова Е.В. Кислотность почв таежной и тундровой зон европейского Северо-Востока России. СПб.: Наука, 2013. 160 с.
Шишов Л.Л., Тонконогов В.Д., Лебедева И.И., Герасимова М.И. Классификация и диагностика почв России. Смоленск: Ойкумена, 2004. 342 с.
Bergkvist B., Folkeson L., Berggren D. Fluxes of Cu, Zn, Pb, Cd, Cr and Ni in temperate forest ecosystems // Water Air & Soil Pollution. 1989. V. 47. P. 217–286.
Bjelland T., Grube M., Hoem S., Jørgensen S.L., Daae F.L., Thorseth I.H., Øvreås L. Microbial metacommunities in the lichen-rock habitat // Environmental Microbiology Reports, 2011. № 3. P. 434–442.
Cardinale M., Grube M., Castro J.V. Bacterial taxa associated with the lung lichen Lobaria pulmonaria are differentially shaped by geography and habitat // FEMS Microbiology Letters, 2012. № 329(2). P. 1–5.
Dymov A.A., Gabov D.N. Pyrogenic alterations of Podzols at the North-East European part of Russia: morphology, carbon pools, PAH content // Geoderma. 2015. V. 241–242. P. 230–237.
Hokkanen T.J., Jarvienen E., Kuuluvainen T. Properties of top soil and the relationship between soil and trees in a boreal Scots pine stand // Silva Fennica. 1995. V. 29. № 3. P. 189-203.
Ingestad T. Mineral nutrient requirements of Vaccinium vitis-idaea and Vaccinium myrtillus // Physiologia Plantarum, 2006. № 29. P. 239–246.
Järvinen E., Hokkanen T., Kuuluvainen T. Spatial heterogeneity and relationships of mineral soil properties in a boreal Pinus sylvestris stand // Scandinavian J. Forest Research. 1993. V. 8. P. 435–445.
Klavina L., Bikovens O., Steinberga I., Maksimova V., Eglite L. Characterization of chemical composition of some bryophytes common in Latvia // Environmental & Experimental Biology, 2012. № 10. P. 27–34.
Koch A.S., Matzner E. Heterogeneity of soil and soil solution chemistry under Norway spruce (Piceaabies Karst.) and European beech (Fagussilvatica L.) as influenced by distance from the stem basis // Plant & Soil. 1993. V. 151. P. 227–237.
Levia D.F., Frost E.E. A review and evaluation of stemflow literature in the hydrologic and biogeochemical cycles of forested and agricultural ecosystems // J. Hydrology. 2003. V. 274. P. 1–29.
Maier S., Schmidt T.S.B., Zheng L., Peer T., Wagner V., Grube M. Analyses of dryland biological soil crusts highlight lichens as an important regulator of microbial communities // Biodiversity & Conservation. 2014. V. 23. Is. 7. P. 1735–1755.
Sigurbjörnsdóttir M.A., Andrésson Ó.S., Vilhelmsson O. Nutrient scavenging activity and antagonistic factors of non-photobiont lichen-associated bacteria: a review // World J. Microbiology & Biotechnology. 2016. № 32. Article number: 68.
Spribille T., Tuovinen V., Resl P., Vanderpool D., Wolinski H., Aime M.C., Schneider K., Stabentheiner E., Toome-Heller M., Thor G., Mayrhofer H., Johannesson H., McCutcheon J.P. Basidiomycete yeasts in the cortex of ascomycete macrolichens // Science. 2016. № 353(6298). P. 488–492.
Tilman D. Secondary succession and the patterns of plant dominance along experimental nitrogen gradients // Ecological Monographs. 1987. V. 57. Is. 3. P. 189–214.
Дополнительные материалы отсутствуют.