

Лесоведение, 2022, № 1, стр. 21-33
Цикличность радиального прироста ствола и жизненное состояние дуба черешчатого в нагорной и пойменной дубравах южной лесостепи
Н. Ф. Каплина *
Институт лесоведения РАН
143030 п/о Успенское, Московская обл., Россия
* E-mail: kaplina@inbox.ru
Поступила в редакцию 02.04.2021
После доработки 20.06.2021
Принята к публикации 21.07.2021
- EDN: VJHBKD
- DOI: 10.31857/S0024114821060061
Аннотация
На примере дуба черешчатого в двух контрастных экотопах обосновано представление о жизненном состоянии дерева как совокупности долговременной, средневременной и кратковременной составляющих (по длительности восстановления скелетных осей кроны, осей ветвления, облиственных побегов). Показано, что индикаторами жизненного состояния могут служить одноименные составляющие прироста ранней древесины ствола. Составляющие прироста поздней древесины характеризуют потенциал жизненного состояния. Долговременная составляющая радиального прироста описана суммой тренда и синусоиды, средневременная составляющая – суммой двух синусоид. Тренд определяется конкуренцией в древостое; ее снижение вследствие массового усыхания дуба улучшило долговременную составляющую жизненного состояния и радиального прироста выживших деревьев. Цикличность радиального прироста в лесостепи обусловлена воздействием засух и филлофагов. Период колебаний долговременной составляющей прироста равен: ранней древесины – 31–35 лет в нагорной дубраве и 40–44 года в пойменной, поздней древесины – 28–30 лет в нагорной и 51–58 лет в пойменной. Периоды основной и дополнительной синусоид средневременной составляющей в обеих дубравах сходны, составляя соответственно: ранней древесины 12–13 и 7–9 лет, поздней древесины – 14–16 и 9–11 лет. Риск ухудшения жизненного состояния дуба повышается в годы наложения минимумов циклов долговременной и средневременной составляющих прироста ранней древесины ствола. Риск усыхания дуба повышается при снижении прироста поздней древесины до 0.5 мм, что наблюдалось в пойменной дубраве в период массового усыхания дуба, а в нагорной дубраве – в начале 2000-х годов и ожидается в конце 2020-х годов у деревьев дуба среднего и худшего развития.
Ослабление и периодическое отмирание дуба в Европе наблюдается более 100 лет, ожидается его новая волна на больших площадях (Бугаев и др., 2013).
Исследованиями с середины 1940-х годов в Теллермановском опытном лесничестве Института лесоведения РАН установлено, что засухи и насекомые-филлофаги вызывают ухудшение состояния и повышение отпада дуба (Дубравы …, 1975; Состояние …, 1989; Экосистемы …, 2004).
Понятие “жизненное состояние” или “жизненность” как способность дерева к выживанию, росту и развитию применяется в различных научных направлениях. В ценопопуляционных исследованиях жизненное состояние оценивается в первую очередь по показателям формы и роста растения (Злобин, 2009), что близко к классификации роста и развития деревьев по Крафту. Лесопатологическая оценка учитывает главным образом густоту кроны, показатели листьев и побегов, наличие усохших ветвей (Алексеев, 1989; Eichhorn et al., 2016; Правила …, 2020).
Возможность индикации состояния дуба черешчатого по радиальному приросту подтверждается многочисленными исследованиями, в том числе по дубу (Воронцов, 1978; Ловелиус, 1979; Демаков, 2002; Рубцов, Уткина, 2008; Gričar, 2012; Rybníček et al., 2015; Dobbertin, Neumann, 2016). Наиболее изучено влияние дефолиации дуба на его прирост и усыхание (Рубцов, Уткина, 2008; Лямцев, 2018).
Ведущий фактор радиального прироста и состояния деревьев и древостоев – изменчивость климата под влиянием солнечной активности; цикличность радиального прироста открывает возможности его прогнозирования (Douglass, 1919; Воронцов, 1978). В XX в. депрессии твердолиственных лесов повторялись с периодичностью 10–12 лет, а особенно сильные – 25–30 лет (Состояние …, 1989).
Ранее на примере нагорной снытево-осоковой дубравы нами показано соответствие числа рядов сосудов и ширины прироста ранней древесины ствола типу развития кроны дуба (Каплина, 2019) и выделены 12-летние циклы прироста ранней (РД) и поздней (ПД) древесины ствола (Каплина, 2020), в минимумах которых ухудшается жизненное состояние дуба.
Цель статьи – выделение циклов радиального прироста РД и ПД ствола в дубравах двух контрастных экотопов южной лесостепи, анализ их сопряженности с циклами жизненного состояния деревьев, выявление периодов повышенного риска усыхания дуба.
ОБЪЕКТЫ И МЕТОДИКА
Изучены дубравы на постоянных пробных площадях (пр. пл.) Теллермановского опытного лесничества. С начала 1980-х годов здесь исследовались причины и последствия массового усыхания дуба и динамика восстановления крон выживших деревьев (Состояние …, 1989; Ильюшенко, Романовский, 2000).
Нагорная дубрава снытево-осокового типа (кв. 6, пр. пл. 1–5, 7 общей площадью 0.55 га, 51°20′53″ с.ш., 41°58′35″ в.д.), характеризуется периодическими почвенными засухами. Древостой создан посевом дуба поздней фенологической формы по вырубке. Показатели первого яруса древостоя в 2012 г.: состав – 10Д + Я (где Д – дуб, Я – ясень), возраст – 80 лет, среднеквадратический диаметр – 31.4 см, средняя высота – 27.4 м, класс бонитета – I, число стволов – 360 шт. га–1, сумма площадей сечений – 27.9 м2 га–1. В первой половине 1970-х годов здесь не наблюдалось массового усыхания дуба, как и в большинстве дубрав моложе 60 лет. Во второй половине 1990–начале 2000 гг. отмечено ухудшение жизненного состояния и повышенный отпад дуба (Каплина, 2006, 2019; Каплина, Селочник, 2015).
Пойменная дубрава – ландышево-ежевичного типа (кв. 47, пр. пл. 1, площадь 0.436 га, 51°19′28″ с.ш., 41°58′23″ в.д.). Весеннее затопление здесь может сменяться летним иссушением (Состояние …, 1989). Древостой естественного, в основном семенного происхождения, дуб – ранней фенологической формы. Показатели первого яруса древостоя в 2011 г.: состав – 10Д, возраст – 120 лет, среднеквадратический диаметр – 40.9 см, средняя высота – 23.9 м, класс бонитета – II, число стволов – 179 шт. га–1, сумма площадей сечений – 23.5 м2 га–1. Во второй половине 1960-х–первой половине 1970-х гг. древостой пострадал от массового усыхания дуба (Состояние …, 1989; Каплина, Селочник, 2015).
Идентификацию типов развития кроны дуба с 1985 г. проводили с использованием оригинальной классификации (Каплина, Селочник, 2009, 2015). Она разработана на основе материалов многолетнего визуального мониторинга формы кроны дуба по морфологическим признакам еe развития, деградации и повреждения, проведенного Н.Н. Селочник (2015). Раскидистый тип кроны (Р-тип) характерен для деревьев I и частично II классов роста и развития по Крафту, с мощными нижними ветвями, формирующимися в период быстрого роста дерева; зонтиковидный тип (З-тип) – частично II, чаще III классов по Крафту, без нижних крупных ветвей, утраченных в процессе роста в условиях различных неблагоприятных факторов; узкокронный тип (У-тип) – обычно IV и V классов, в основном утративших первичные ветви кроны, с заменой их неразвитыми вторичными ветвями. Типы развития кроны являются также стадиями деградации, повреждения и восстановления (формирования вторичной кроны) кроны. Классификация может применяться в условиях различных естественных и антропогенных факторов, а также для отдельно стоящих деревьев (не требует оценки социального статуса дерева).
Идентификацию типа развития кроны до 1985 г. (начало визуального мониторинга) и подтверждение визуальных оценок проводили по числу рядов сосудов и ширине прироста РД ствола, сглаженных 5-летним скользящим средним: Р-тип образует преимущественно 3 ряда сосудов общей шириной 0.8–1.2 мм, З-тип –2 ряда сосудов шириной 0.4–0.8 мм, У-тип – 1 ряд сосудов шириной 0.2–0.4 мм (Каплина, 2019). Последний ряд учитывался, если он был заполнен сосудами более чем на 20% своей длины. Мы склонны считать данный способ более объективным, чем идентификация по морфологическим признакам кроны.
Керны стволов взяты на высоте 1.3 м в основном с юго-восточной стороны. Керны в сырорастущем состоянии либо после выдерживания в воде не менее 1 ч (до постоянных размеров радиального прироста) обрабатывали лезвием и сканировали при разрешении 1200 dpi. Показатели приростов РД и ПД измеряли с помощью компьютерной программы GetData Graph Digitizer 2.24 с точностью не менее ±0.02 мм. Приросты анализировали с 36-летнего возраста деревьев, то есть с начала формирования зрелой древесины (Helińska-Raczkowska, 1994). Перекрестное датирование проводили визуально, что не представляет затруднений благодаря синхронности радиального прироста ПД у деревьев всех типов развития кроны.
Получены временные ряды приростов: в нагорной дубраве – для большинства деревьев на пробных площадях (146 шт. в 2013 и 2014 г.), в пойменной дубраве – для всех деревьев (78 шт. в 2014 и 2016 г.). Из них в каждом древостое выбрано по 3 группы (по 10 временных рядов в каждой группе), наиболее характерные для различных типов развития кроны и свойственной им динамики. В нагорной дубраве: 1-я группа – деревья раскидистого типа весь период исследования (Р-Р); 2-я группа – деревья раскидистого типа в 1960-е годы, перешедшие в зонтиковидные в 1990-е (Р-З); 3-я группа – деревья зонтиковидные в 1960-е годы, перешедшие в узкокронные в 1990-е (З-У). В пойменной дубраве: 1-я группа – по 5 деревьев раскидистого и зонтиковидного типа до массового усыхания дуба в 1960–1970-е годы, развившие раскидистые кроны в 1980-е (РЗ-Р); 2-я группа – деревья зонтиковидного типа весь период исследования (З-З); 3-я группа – деревья, изначально узкокронного типа, развившие зонтиковидные кроны в 1980-е годы (У-З). Временные ряды приростов РД и ПД получали как средние для группы деревьев.
Массу листьев дуба оценивали по опаду, собранному на площадках размером 0.5 × 0.5 м2: в снытево-осоковой дубраве – в 8–9-кратной повторности, в пойменной – в 3–4-кратной. Учтены потери на гетеротрофное дыхание опада при длительном хранении в размере 26% (Каплина, Жиренко, 2012).
Различия величин оценивали с помощью дисперсионного анализа на уровне значимости α ≤ 0.05. Коэффициенты уравнений трендов временных рядов получали с помощью программы MS EXEL. Спектральный анализ Фурье и вычисление коэффициентов тригонометрических уравнений проводили с использованием пакета STATISTICA.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
На взаимосвязь развития кроны и состояния деревьев указывали различные исследователи. Жизнеспособность дуба в настоящем и будущем определяется развитием и морфоструктурой кроны (Лохматов, 1999). Социальный статус – важный показатель при изучении состояния кроны и прироста деревьев (Eichhorn et al., 2016).
Признаки жизнестойкости как способности поддержания и восстановления жизнеспособности (Демаков, 2002) наиболее выражены в условиях неблагоприятных факторов среды. Наибольшую информативность для прогнозирования имеет состояние ветвей верхней части кроны, что объясняется длительностью их восстановления (Алексеев, 1989). Ветви восстанавливаются в рамках архитектурной модели кроны (Костина, 2008). В дубравах Теллермановского леса наблюдалось успешное восстановление крон из водяных побегов после массового усыхания дуба (Состояние …, 1989; Ильюшенко, Романовский, 2000).
При оценке состояния дерева необходимо учитывать не только влияние стрессовых факторов, но и их кумулятивные проявления (Алексеев, 1989). Долговременная динамика роста и жизненности может не совпадать с кратковременными изменениями (Dobbertin, 2005).
В данной работе, развивая предыдущие представления (Каплина, Селочник, 2015), жизненное состояние дерева мы рассматриваем как совокупность составляющих, характеризующихся длительностью восстановления соответствующих морфофункциональных частей кроны: долговременной составляющей (скелетные оси кроны), средневременной (оси ветвления кроны) и кратковременной, или текущей (облиственные побеги и листья). Долговременная составляющая может быть оценена по типу развития кроны дуба, среде- и кратковременная – по густоте ветвления и облиственности кроны, соответственно. Для индикации составляющих жизненного состояния необходимо выделение соответствующих составляющих радиального прироста.
Временные ряды и тренды приростов РД и ПД. Прирост РД (рис. 1а, 1б) в обеих дубравах соответствует типу развития кроны и динамике его изменения. Коэффициент вариации (CV) прироста РД невысок – 15–30%, причем он наименьший в группах деревьев, не изменивших тип развития кроны: Р-Р в нагорной дубраве и З-З – в пойменной (табл. 1). Взаимосвязь прироста РД с развитием кроны объясняется его жизненно важной ролью в снабжении кроны пасокой, что обеспечивается его формированием одновременно с распусканием листьев за счет запасных веществ предыдущих лет.
Рис. 1.
Временные ряды и тренды приростов РД (а, б) и ПД (в, г) и их соотношения (д, е) в нагорной (а, в, д) (Каплина, 2019) и пойменной (б, г, е) дубравах групп деревьев лучшего (1), среднего (2) и худшего (3) развития кроны.
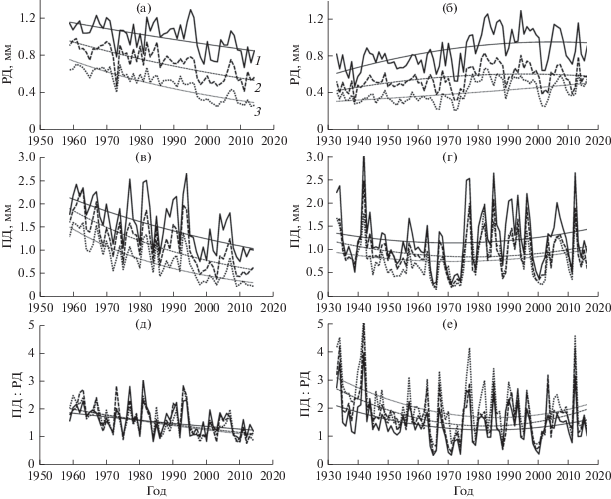
Таблица 1.
Коэффициенты вариации (CV, %) радиального прироста и его составляющих в нагорной (1959–2014 гг.) и пойменной (1933–2016 гг.) дубравах
| Нагорная дубрава | Пойменная дубрава | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| группа | прирост | тренд | Д + С | К | группа | прирост | тренд | Д + С | К |
| Ранняя древесина | |||||||||
| Р-Р | 15 | 9 | 7 | 10 | РЗ-Р | 22 | 11 | 12 | 15 |
| Р-З | 22 | 17 | 8 | 11 | З-З | 22 | 8 | 12 | 16 |
| З-У | 29 | 26 | 8 | 11 | У-З | 28 | 15 | 16 | 18 |
| Поздняя древесина | |||||||||
| Р-Р | 34 | 19 | 16 | 23 | РЗ-Р | 50 | 2 | 27 | 42 |
| Р-З | 44 | 31 | 19 | 24 | З-З | 48 | 3 | 24 | 41 |
| З-У | 52 | 41 | 19 | 25 | У-З | 52 | 2 | 29 | 42 |
Известно, что прирост ПД гораздо более изменчив по годам, его CV в обеих дубравах примерно в 2 раза выше, чем прироста РД. В отличие от РД, ПД формируется за счет ассимилятов текущего года и менее тесно коррелирует с приростом РД текущего года, чем следующего (Царалунга, Гарнага, 2004; Каплина, 2019). В слабо пострадавшей от усыхания нагорной дубраве прирост ПД составлял в 1960–1970-е годы в среднем около 1 мм и выше (в зависимости от типа развития кроны), снижаясь в засушливые годы (рис. 1в, 1г). В сильно пострадавшей пойменной дубраве он снизился уже с 1950-х годов в среднем ниже 1 мм, а с середины 1960-х годов – ниже 0.5 мм. Снижение прироста ПД и соотношения приростов ПД : РД свидетельствует об ослаблении дуба (Состояние …, 1989; Царалунга, Гарнага, 2004; Рубцов, Уткина, 2008). Согласно приведенным данным, это утверждение полностью справедливо только для прироста ПД. Он тесно связан с продукционным состоянием дерева, тем самым определяя и потенциал его жизненного состояния. Соотношение приростов ПД : РД (рис. 1д, 1е) сходно по динамике с приростом ПД в пределах группы деревьев. Однако, в отличие от прироста ПД, оно не отражает наблюдаемой более низкой облиственности крон в группах худшего развития кроны (Каплина, Селочник, 2015).
В нагорной дубраве наблюдается тренд снижения приростов РД и ПД и их соотношения, характерный для сомкнутых дубрав (рис. 1). В пойменной дубраве, в условиях сильного разреживания полога в результате массового усыхания дуба, приросты РД и ПД вернулись к наблюдавшимся до усыхания величинам уже во второй половине 1970-х годов. Затем прирост РД достиг величин, характерных для деревьев более развитого типа кроны (в 1й и 3й группах деревьев). Можно заключить, что в обеих дубравах тренд определяется в основном не возрастом, а конкурентными отношениями.
Тренды вычисляли: в нагорной дубраве – согласно уравнению экспоненты (1), в пойменной дубраве – в соответствии с уравнением параболы 2-го порядка (2):
где y – радиальный прирост, x – календарный год, a, b, c – коэффициенты.Коэффициент детерминации трендов в нагорной дубраве (r2) составляет для приростов РД и ПД соответственно 0.32–0.75 и 0.35–0.66, увеличиваясь от группы наиболее развитых к группе наименее развитых деревьев, т.е. с усилением конкуренции, а также в группах, изменивших тип развития дерева. В пойменной дубраве r2 составляет для прироста РД 0.20–0.30 и не значим для прироста ПД. Таким образом, тренд объясняет 1/3–2/3 дисперсии радиального прироста в нагорной дубраве (большие величины – у деревьев худшего развития) и менее 1/3 – в пойменной. Соответственно этому изменяется и CV тренда (табл. 1).
Выделение циклических составляющих приростов РД и ПД. Согласно эмпирической модели при объедании филлофагами 2 г. подряд по 75% листьев или 3 г. подряд по 50% листьев число усохших деревьев превысит 10%, что можно считать началом ухудшения долговременной составляющей жизненного состояния древостоя; прирост по объему ствола уменьшится соответственно примерно на 37 и 25% в год (Лямцев, 2018). По данным исследований в Теллермановском лесу в нагорной дубраве зависимость потерь прироста от дефолиации заметно усиливается уже на 2-й год, в то время как в пойменной – лишь на 3-й год, причем эта связь теснее у деревьев худшего развития. В годы с лучшими погодными условиями потери прироста меньше (Рубцов, Уткина, 2008).
Очевидно, для дуба наиболее опасна цикличность прироста с продолжительностью его минимальных значений не менее 2-х лет, что соответствует периодам колебаний не менее 6 лет. В дубравах лесостепи и степи этому критерию соответствуют 11-летние циклы вспышек насекомых-филлофагов (Лямцев, 2018; Рубцов, Уткина, 2019) и площади листовой поверхности древостоя (Каплина, Жиренко, 2012), а также циклы радиального прироста ствола: 6–8- и 13–17-летние (Акимов, 1996), 10–11-, 13-, 16–17-, 22-, 35-летние (Маликов, 2000), 7, 11, 22, 40–50 и 100-летние (Миленин, 2012).
Выделение циклических составляющих временных рядов в объектах исследования проведено после вычитания трендов. С помощью анализа Фурье в нагорной дубраве для приростов РД и ПД выявлена максимальная спектральная плотность в диапазоне периодов (в порядке убывания мощности): 10–15 и 25–30 лет. В пойменной дубраве для прироста РД выявлена повышенная спектральная плотность в диапазоне: для прироста РД – 30–50 и 10–15 лет; для прироста ПД – 7–8, 30–50 и 10–15 лет. С учетом этого предложена аддитивная модель циклических составляющих, описывающих три диапазона периодов колебаний (3):
(3)
$\begin{gathered} y = {{A}_{1}}\sin [\left( {\frac{{2\pi \left( {x + {{c}_{1}}} \right)}}{{{{T}_{1}}}}]} \right) + {{A}_{2}}\sin [\left( {\frac{{2\pi \left( {x + {{c}_{2}}} \right)}}{{{{T}_{2}}}}]} \right) + \\ + \,\,{{A}_{3}}\sin [\left( {\frac{{2\pi \left( {x + {{c}_{3}}} \right)}}{{{{T}_{3}}}}]} \right), \\ \end{gathered} $Выделение Д-колебаний независимыми способами – как разности 13-летнего скользящего среднего (отнесенного к середине интервала) и тренда и согласно уравнению (3) приводит к сходным результатам (рис. 2). Также сходны результаты выделения С-колебаний: как разности 5- и 13‑летних скользящих средних и 2) как суммы С1 и С2 по уравнению (3) (рис. 3). Таким образом, сглаживание временных рядов методом скользящей средней позволяет выявить основные черты Д- и С-колебаний радиального прироста, несмотря на эффект ложных колебаний Слуцкого–Юла.
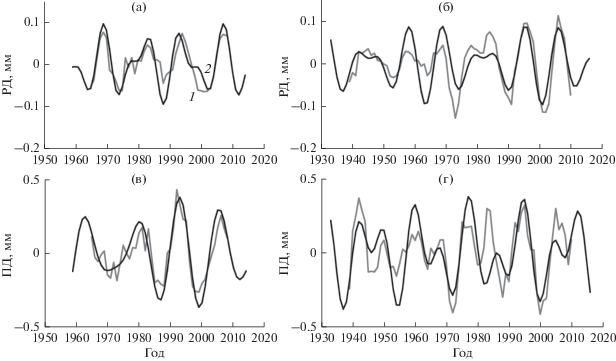
Рис. 2.
Д-циклы прироста РД (а, б) и ПД (в, г) в нагорной (а, в) и пойменной (б, г) дубравах, выделенные с помощью: 1 – скользящего среднего и 2 – уравнения (3), на примере группы деревьев среднего развития.
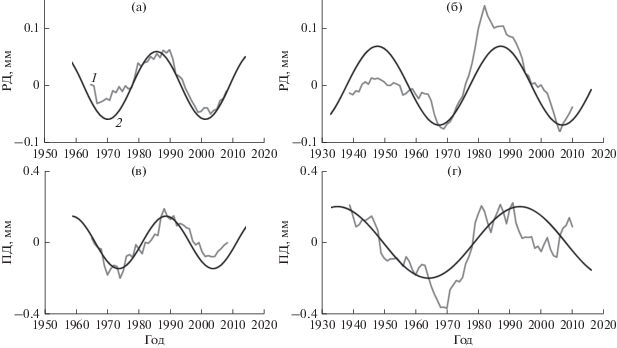
Рис. 3.
С-циклы прироста РД (а, б) и ПД (в, г) в нагорной (а, в) и пойменной (б, г) дубравах, выделенные с помощью: 1 – скользящего среднего и 2 – уравнения (3), на примере группы деревьев среднего развития.
Коэффициенты уравнений (3) в основном значимы при α ≤ 0.05, кроме трех коэффициентов в группе Р-Р нагорной дубравы, значимых при α ≤ 0.1. Для всех уравнений r2 находится в диапазоне: в нагорной дубраве 0.31 ≤ r2 ≤ 0.41, в пойменной дубраве 0.26 ≤ r2 ≤ 0.45 (α ≤ 0.05). То есть от 1/4 до почти 1/2 дисперсии детрендированных временных рядов объясняется уравнением (3). Соответственно этому, CV К-колебаний несколько больше, чем CV суммы Д- и С-циклов (табл. 1). Таким образом, в нагорной дубраве больший вклад в изменчивость приростов РД и ПД вносят тренд и Д- и С-циклы. В пойменной дубраве те же величины изменчивости достигаются для прироста РД примерно равным вкладом К-колебаний и остальных составляющих, а для прироста ПД – большим вкладом К-колебаний.
Периоды всех циклических составляющих, полученные по уравнению (3), не зависят от типа развития кроны (табл. 2). Абсолютная величина их амплитуд снижается, а их отношение к величине тренда, наоборот, повышается с ухудшением развития кроны (табл. 3).
Таблица 2.
Периоды (лет) колебаний радиального прироста, вычисленные по уравнению (3)
| Нагорная дубрава | Пойменная дубрава | ||||||
|---|---|---|---|---|---|---|---|
| группа | Д | С1 | С2 | группа | Д | С1 | С2 |
| Ранняя древесина | |||||||
| Р-Р | 31.0 | 12.7 | 7.6 | РЗ-Р | 44.3 | 12.4 | 9.1 |
| Р-З | 30.9 | 12.7 | 7.6 | З-З | 39.3 | 12.5 | 9.3 |
| З-У | 34.9 | 13.2 | 7.6 | У-З | 43.7 | 12.1 | 9.2 |
| Поздняя древесина | |||||||
| Р-Р | 29.9 | 14.1 | 10.8 | РЗ-Р | 56.0 | 15.8 | 8.9 |
| Р-З | 27.9 | 14.5 | 11.4 | З-З | 58.2 | 16.0 | 8.9 |
| З-У | 29.4 | 14.1 | 11.1 | У-З | 50.7 | 15.6 | 9.0 |
Таблица 3.
Амплитуды (мм) колебаний радиального прироста, вычисленные по уравнению (3)
| Нагорная дубрава | Пойменная дубрава | ||||||
|---|---|---|---|---|---|---|---|
| группа | Д | С1 | С2 | группа | Д | С1 | С2 |
| Ранняя древесина | |||||||
| Р-Р | 0.068 | 0.086 | 0.058 | РЗ-Р | 0.100 | 0.076 | 0.057 |
| Р-З | 0.059 | 0.058 | 0.037 | З-З | 0.069 | 0.055 | 0.042 |
| З-У | 0.039 | 0.038 | 0.023 | У-З | 0.070 | 0.044 | 0.033 |
| Поздняя древесина | |||||||
| Р-Р | 0.15 | 0.30 | 0.15 | РЗ-Р | 0.31 | 0.27 | 0.26 |
| Р-З | 0.15 | 0.24 | 0.14 | З-З | 0.20 | 0.19 | 0.19 |
| З-У | 0.10 | 0.13 | 0.13 | У-З | 0.25 | 0.16 | 0.18 |
Д-циклы радиального прироста и жизненного состояния. Период Д-циклов прироста в пределах древостоя не различается значимо между РД и ПД, составляя в среднем для древостоев: в нагорной дубраве – 33 г. для РД и 29 лет – для ПД, в пойменной дубраве – соответственно 42 и 54 г. То есть долговременная динамика развития кроны и продукции ствола зависит от одних и тех же факторов. В пойменной дубраве период колебаний значимо больше, чем в нагорной (α ≤ 0.05), что, возможно, объясняется нарушением в ней полога в результате массового усыхания дуба.
Минимум Д-цикла прироста РД в нагорной дубраве приходится на первую половину 1970-х годов, а в пойменной – на вторую половину 1960-х годов, что согласуется с зафиксированным более ранним началом усыхания дуба в пойме (Состояние …, 1989).
Максимумы прироста РД в обеих дубравах пришлись на середину – вторую половину 1980-х годов, что совпадает со стабилизацией развития крон дуба в нагорной дубраве и улучшением – в пойменной. Так, из деревьев Р-типа развития, по учету 1983 г., к 1997 г. сохранили Р-тип в нагорной и пойменной дубравах соответственно 86 и 97%, усохло – 7 и 0%. В то же время в пойменной дубраве 30% деревьев У-типа перешли в З-тип (Каплина, Селочник, 2015).
Следующий минимум пришелся на конец 1990-х–первую половину 2000-х гг. в нагорной дубраве и на 2000-е годы в пойменной. Развитие крон дуба ухудшилось в нагорной дубраве и стало разнонаправленным в пойменной. Из деревьев Р‑типа развития, по учету 1998 г., к 2012 г. сохранили Р-тип в нагорной и пойменной дубравах соответственно лишь 32 и 53%, усохло соответственно 8 и 24%. В то же время в пойменной дубраве 30% деревьев З-типа перешло в Р-тип и почти половина деревьев У-типа перешла в З-тип (Каплина, Селочник, 2015).
Амплитуда Д-циклов прироста РД в нагорной дубраве – 0.04–0.07 мм значимо меньше (α ≤ 0.1), чем в пойменной – 0.07–0.10 мм. Это составляет относительно ширины одного ряда сосудов РД в нагорной дубраве до 18–20%, в пойменной – до 25–35% (большие абсолютные и меньшие относительные величины зафиксированы у деревьев лучшего развития). Следовательно, снижение прироста РД только за счет Д-колебаний могло привести лишь к небольшому ухудшению развития крон, чувствительному лишь для наименее развитых деревьев. Амплитуда Д-циклов прироста ПД в нагорной дубраве в 2 раза меньше, чем в пойменной, что указывает на большее снижение потенциала жизненного состояния дуба в годы минимумов в пойменной дубраве.
Наблюдения за массовым усыханием нагорных дубрав Теллермановского леса подтверждают, что засухи являются первичным фактором ухудшения жизненного состояния. Так, площадь листовой поверхности древостоев сокращалась в дубравах, как поврежденных, так и не поврежденных филлофагами. Другие лесообразующие виды снизили площадь листовой поверхности одновременно с дубом. Известно, что пойменные дубравы Теллермановского опытного лесничества пребывают в ритме обычного для них периодического отпада, согласованного с водным режимом р. Хопер (Экосистемы …, 2004). Волны массового отмирания дуба 1927–1946 и 1964–1983 гг. (Бугаев и др., 2013) совпадают с наиболее засушливыми периодами. Кросс-спектральный анализ показал зависимую от осадков и температуры цикличность радиального прироста сосны: 7–8-, 11-, 20–26- и 41-летнюю (Матвеев, Тимащук, 2019).
Вместе с тем, длинноволновые колебания прироста отражают колебания плотности древостоев (Демаков, 2015). Период влияния повреждения полога на прирост – более 21 г. (Nowacki, Abrams, 1997). В Германии широко распространен тренд снижения прироста дуба, вызванный конкуренцией, и затем примерно 30-летнее его повышение после рубок 1930–1940 гг., в отличие от отдельно стоящих дубов (Scharnweber et al., 2019). Засухи вызывают не только сокращение роста, но и его увеличение в течение 11 лет, что интерпретируется как снижение конкуренции (Rubio-Cuadrado et al., 2018).
Таким образом, долговременная составляющая жизненного состояния характеризуется суммой тренда и Д-циклов прироста РД. В нагорной дубраве вклад тренда превалирует (табл. 1), следовательно, снижение ее густоты рубками ухода было недостаточным. Массовое усыхание дуба в пойме способствовало улучшению развития крон выживших деревьев, чем вызвало увеличение амплитуды Д-цикла прироста.
Продолжительность восходящей ветви Д-цикла прироста РД (16–22 г) достаточна для восстановления скелетных осей, что можно считать признаком адаптации морфоструктуры кроны дуба к засухам.
С-циклы радиального прироста и жизненного состояния. Основной вклад в С-колебания прироста РД вносят С1-циклы: их амплитуда в нагорной дубраве равна амплитуде Д-циклов, в пойменной – немного меньше, чем Д-циклов (табл. 3). Период С1-циклов прироста РД в обеих дубравах одинаков – около 12–13 лет, что указывает на сходные экзогенные факторы этих колебаний, такие как засухи и филлофаги. В Теллермановском опытном лесничестве вспышки насекомых-филлофагов проходили с 11-летней периодичностью, причем в последние 30 лет уменьшается вредоносность одних видов филлофагов и увеличивается – других (Рубцов, Уткина, 2019). Обращает на себя внимание независимость периода С1-циклов радиального прироста РД от видового состава филлофагов, а также способность восстановления осей ветвления кроны дуба за 6–7 лет на восходящей ветви С1-циклов. Вместе со способностью восстановления скелетных осей кроны на восходящей ветви Д-цикла, это свидетельствует в пользу адаптации морфоструктуры кроны дуба к цикличности засух и повреждения филлофагами.
В обоих древостоях период С1-циклов прироста РД меньше чем ПД на 1–3 г. (α ≤ 0.05), видимо, из-за меньшей инерции колебаний облиственности кроны по сравнению с продукцией ствола. Колебания прироста ПД в нагорной дубраве в большей мере определяются С1-циклами, в меньшей – Д- и С2-циклами. В пойменной дубраве колебания прироста ПД более сложные, с примерно равным вкладом этих трех циклических составляющих.
Вклад С2 в С-колебания заключается в корректировке асимметрии, сдвига и амплитуды экстремумов. В приросте ПД пойменной дубравы С2‑циклы наиболее заметны, описывая всплески прироста между основными максимумами. Период С2-циклов прироста РД в нагорной дубраве несколько меньше, чем в пойменной, соответственно 7.6 против 9.2 лет. В нагорной дубраве для прироста РД он меньше, чем для ПД (как и период С1) на 3–4 г. (α ≤ 0.05), а в пойменной дубраве – одинаков для РД и ПД.
С-колебания прироста РД в обеих дубравах в основном синхронны. Так, их минимумы в обеих дубравах пришлись на середину 1960-х, середину 1970-х, конец 1980-х, конец 1990–начало 2000, начало 2010 гг., а максимумы – на конец 1960-х, середину 1980-х, 1990-х, 2000-х гг. (рис. 3). Также довольно синхронны С-колебания приростов РД и ПД в нагорной дубраве, в то время как в пойменной дубраве синхронность нарушается из-за значительного вклада С2 в прирост ПД.
В нагорной дубраве прирост РД в среднем за три года минимума (2011–2013 гг.) был ниже, чем за три года максимума (2006–2008 гг.): на 0.13–0.24 мм, или в 1.3–1.5 раза (большие абсолютные и меньшие относительные величины – у деревьев лучшего развития), а масса листового опада снизилась на 0.9 т га–1 или в 1.4 раза. В пойменной дубраве прирост РД в среднем за три года минимума (2010–2012 гг.) был ниже, чем за три года максимума (2005–2007 гг.): на 0.16–0.34 мм, или в 1.4–1.5 раза. Масса листового опада в пойме в годы минимума была того же уровня, что и в нагорной дубраве – соответственно 2.1 и 1.9 т га–1. По нашим наблюдениям, следующий максимум облиственности пришелся на 2017 г. во всех типах дубрав Теллермановского леса.
В обеих дубравах в 1960-х–1970-х гг. на минимум Д-цикла прироста РД наложились минимумы С-циклов, усилив его снижение (рис. 4). Прирост РД был ниже тренда: в нагорной дубраве на 0.08–0.15 мм, в пойменной дубраве – на 0.15–0.23 мм. Относительно ширины одного ряда сосудов РД это составляет: в нагорной дубраве до 37–40%, в пойменной – до 55–75% (большие абсолютные и меньшие относительные величины – у деревьев лучшего развития). Это объясняет ухудшение развития крон и жизненного состояния дуба в нагорной дубраве и массовое его усыхание в пойменной. Следующее наложение минимумов Д- и С-циклов прироста РД пришлось в обеих дубравах на начало 2000-х годов и начало 2010 гг. В этот период прирост РД был ниже тренда: в нагорной дубраве – на 0.08–0.13 мм; в пойменной – на 0.12–0.16 мм, что составило от снижения в 1960–1970-е годы соответственно 87–100 и 70–80%.
Рис 4.
Долговременная составляющая (пунктирная линия) и сумма долговременной и средневременной составляющих (сплошная линия) прироста РД (а, б) и ПД (в, г) в нагорной (а, в) и пойменной (б, г) дубравах групп деревьев лучшего (1), среднего (2) и худшего (3) развития кроны.
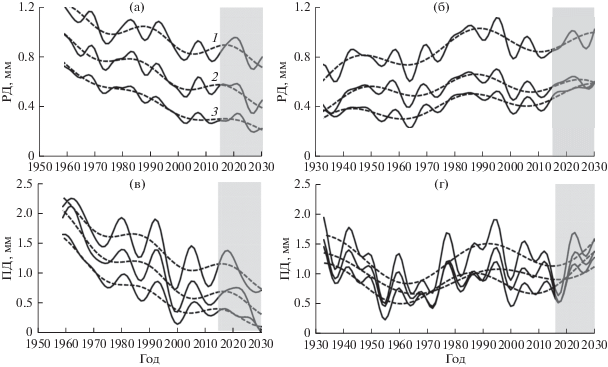
Уменьшение суммы тренда и Д- и С-циклов прироста ПД ниже 0.5 мм может служить признаком снижения потенциала жизненного состояния. Такое снижение наблюдалось у выживших деревьев в пойменной дубраве до и в период массового усыхания дуба, а также в обеих дубравах в начале 2000-х и 2010-х гг. у деревьев У-типа развития и, в некоторых случаях, З-типа (рис. 4).
Согласно исследованиям ослабления и усыхания дуба для окончательной гибели деревьев всегда необходим внешний толчок (Демаков, 2002). После экстремально засушливого 1972 г. в аридных условиях произрастания деревья дуба всех рангов развития ухудшили прирост до 8 лет, что вызвано повреждением различных органов дуба (Сапанов, 2019). Массовые размножения насекомых начинаются лишь после достаточно сильного падения прироста (Лямцев, 2018), вызванного другими факторами. Пики ослабления и усыхания дуба приходятся на годы резкого снижения радиального прироста как вследствие климатических флуктуаций, так и в результате вспышек численности листогрызущих насекомых (Царалунга, Гарнага, 2004).
Полученные нами и приведенные в литературе результаты позволяют полагать, что первичный фактор цикличности жизненного состояния и риска усыхания дуба в исследуемых дубравах – это цикличность засух. По классификации факторов P.D. Manion (Экосистемы …, 2004), минимальный уровень Д-цикла влагообеспеченности и сопутствующая активизация филлофагов являются предрасполагающими факторами, наложение минимумов Д- и С-циклов влагообеспеченности – инициирующим, а последующая вспышка филлофагов – усугубляющим фактором.
О динамике жизненного состояния дуба в ближайшие годы можно судить по долговременной составляющей приростов РД и ПД (сумме тренда и Д-циклов), а о периодах повышенного риска усыхания дуба – по сумме их долго- и средневременных составляющих (сумме тренда, Д- и С-циклов) (рис. 4). В нагорной дубраве Д-цикличность определяет ступенчатое снижение прироста и жизненного состояния деревьев. В середине 2020-х годов ожидается очередное их снижение с переходом части деревьев Р-, З- и У-типа соответственно в З-, У-тип и отпад, а в первой половине 2030-х гг. – стабилизация. Уменьшение прироста ПД ниже 0.5 мм в нагорной дубраве с конца 1990-х годов у деревьев У- и части З-типа указывает на снижение потенциала их жизненного состояния и риск усыхания. В пойменной дубраве прогноз оптимистичнее: в 2020-е годы ожидается стабилизация прироста РД и развития крон всех групп деревьев на уровне максимума Д-цикла, возможно увеличение прироста ПД. Наши выводы не противоречат прогнозу о сохранении уровня радиального прироста дуба до 2024 г., учитывающему 11-летнюю цикличность прироста и его зависимость от климатических параметров (Matveev et al., 2018).
Выводы. 1. Обосновано представление о жизненном состоянии дерева как совокупности долговременной, средневременной и кратковременной составляющих (по длительности восстановления морфофункциональных частей кроны: скелетных осей, осей ветвления, облиственных побегов).
2. Двумя независимыми способами выделены циклы долго- и средневременных составляющих радиального прироста ранней и поздней древесины ствола. Показано, что сглаживание временных рядов методом скользящей средней позволяет выявить основные черты цикличности, несмотря на эффект Слуцкого–Юла.
3. Долго- и средневременная составляющие прироста ранней древесины могут служить индикаторами одноименных составляющих жизненного состояния дуба, а составляющие прироста поздней древесины – его потенциала. Долговременная составляющая этих показателей описана суммой тренда и синусоиды: для ранней древесины – с периодом 31–35 лет в нагорной дубраве и 40–44 г. в пойменной, для поздней древесины – 28–30 лет в нагорной и 51–58 лет в пойменной. Средневременная составляющая описана суммой основной и дополнительной синусоид, периоды которых в обеих дубравах сходны и равны соответственно: для ранней древесины – 12–13 и 7–9 лет, для поздней древесины – 14–16 и 9–11 лет.
4. Тренд радиального прироста определяется конкуренцией в древостое, её снижение позволяет улучшить долговременную составляющую жизненного состояния дуба. Массовое усыхание дуба, как и рубки ухода, способствует улучшению развития крон выживших деревьев и тем самым адаптации дуба к повторяющимся засухам.
5. Продолжительность восходящей ветви цикла долговременной составляющей прироста ранней древесины (16–22 г.) достаточна для восстановления скелетных осей, а средневременной составляющей (6–7 лет) – осей ветвления кроны. Это свидетельствует об адаптации структуры кроны дуба к цикличности засух и повреждению филлофагами.
6. Можно полагать, что первичный фактор цикличности радиального прироста, жизненного состояния и риска усыхания дуба в объектах исследования – это цикличность засух. Причем минимум долговременной составляющей обеспеченности влагой и сопутствующая активизация филлофагов являются предрасполагающими факторами, наложение минимумов долго- и средневременных составляющих – инициирующим, а последующая вспышка филлофагов – усугубляющим фактором. Ближайший период повышенного риска ожидается в конце 2020-х гг. в нагорной дубраве.
Список литературы
Акимов П.В. Многолетняя динамика прироста дубовых древостоев в пойменных биогеоценозах низовья Северского Донца: Автореф. дис. … канд. с.-х. наук (спец. 06.03.02). Воронеж: Изд-во Воронежской гос. лесотех. академии, 1996. 20 с.
Алексеев В.А. Диагностика жизненного состояния деревьев и древостоев // Лесоведение. 1989. № 4. С. 51–57.
Бугаев В.А., Мусиевский А.Л., Царалунга В.В. Дубравы лесостепи. Воронеж: Изд-во Воронежской гос. лесотех. академии, 2013. 247 с.
Воронцов А.И. Патология леса. М.: Лесная пром-сть, 1978. 272 с.
Демаков Ю.П. Защита растений. Жизнеспособность и жизнестойкость древесных растений: Учеб. пособие. Йошкар-Ола: Изд-во Марийского гос. тех. университета, 2002. 74 с.
Демаков Ю.П., Исаев А.В. Закономерности динамики радиального прироста деревьев дуба в пойменных лесах заповедника // Научные труды государственного природного заповедника “Большая Кокшага”. Йошкар-Ола: Изд-во Поволжского гос. тех. университета, 2015. Вып. 7. С. 139–156.
Дубравы лесостепи в биогеоценотическом освещении / Под ред. А.А. Молчанова. М.: Наука, 1975. 374 с.
Злобин Ю.А. Популяционная экология растений: современное состояние, точки роста. Сумы: Университетская книга, 2009. 263 с.
Ильюшенко А.Ф., Романовский М.Г. Формирование вторичной кроны дуба и ее роль в динамике состояния древостоев // Лесоведение. 2000. № 3. С. 65–72.
Каплина Н.Ф. Динамика прироста деревьев в нагорных антропогенных дубравах южной лесостепи // Там же. 2006. № 4. С. 3–11.
Каплина Н.Ф. Влияние развития кроны на радиальный прирост ранней и поздней древесины ствола дуба черешчатого // Вестник Поволжского государственного технологического университета. Серия: Лес. Экология. Природопользование. 2019. № 2(42). С. 17–25.
Каплина Н.Ф. Составляющие динамики прироста ранней и поздней древесины ствола дуба черешчатого в нагорной дубраве южной лесостепи // Известия вузов. Лесной журн. 2020. № 5. С. 51–63.
Каплина Н.Ф., Жиренко Н.Г. Динамика фитомассы листьев, состояния и развития крон деревьев нагорной дубравы юго-восточной лесостепи в неблагоприятных условиях последнего десятилетия // Вестник Поволжского государственного технологического университета. Серия: Лес. Экология. Природопользование. 2012. № 2(16). С. 3–11.
Каплина Н.Ф., Селочник Н.Н. Морфология крон и состояние дуба черешчатого в средневозрастных насаждениях лесостепи // Лесоведение. 2009. № 3. С. 32–42.
Каплина Н.Ф., Селочник Н.Н. Текущее и долговременное состояние дуба черешчатого в трех контрастных типах леса южной лесостепи // Лесоведение. 2015. № 3. С. 191–201.
Костина М.В. Иерархическая классификация конструктивных единиц (модулей) древесных растений умеренной зоны // Вестник Тверского гос. университета. Серия “Биология и экология”. 2008. Вып. 9. С. 114–118.
Ловелиус Н.В. Изменчивость прироста деревьев. Дендроиндикация природных процессов и антропогенных воздействий. Л.: Наука, 1979. 232 с.
Лохматов Н.А. Развитие и возобновление степных лесных насаждений. Балаклея: “CiM”, 1999. 498 с.
Лямцев Н.И. Прогнозирование массовых размножений непарного шелкопряда, угрозы повреждения дубрав и необходимости защитных мероприятий. Пушкино: Всерос. НИИ лесоводства и механизации лес. хоз-ва, 2018. 84 с.
Маликов А.Б. Дендроклиматические аспекты состояния дубрав Воронежской области: Автореф. дис. … канд. с.-х. наук (спец. 03.00.16). Воронеж: Изд-во Воронежской гос. лесотех. академии, 2000. 20 с.
Матвеев С.М., Тимащук Д.А. Дендроклиматический анализ 200-летнего древостоя сосны обыкновенной в Воронежском биосферном заповеднике // Лесоведение. 2019. № 2. С. 93–104.
Миленин А.И. Влияние температуры воздуха на радиальный прирост дуба черешчатого в Шиповом лесу // Лесотехнический журн. 2012. № 2. С. 65–69.
Правила санитарной безопасности в лесах. Утв. Постановлением Правительства Российской Федерации от 09.12.2020 №2047. 20 с.
Рубцов В.В., Уткина И.А. Адаптационные реакции дуба на дефолиацию. М.: Институт лесоведения РАН, 2008. 302 с.
Рубцов В.В., Уткина И.А. Реакция лесных насекомых-филлофагов на современное изменение климата // Лесоведение. 2019. № 5. С. 375–384.
Сапанов М.К. Климатогенные факторы внезапного изменения хода роста дерева // Поволжский экологический журн. 2019. № 2. С. 253–263.
Селочник Н.Н. Состояние дубрав Среднерусской лесостепи и их грибные сообщества. М.: Институт лесоведения РАН, 2015. 216 с.
Состояние дубрав лесостепи / Под ред. А.Я. Орлова, В.В. Осипова. М.: Наука, 1989. 230 с.
Царалунга В.В., Гарнага В.В. Радиальный прирост деревьев дуба различных категорий состояния // Вестник Московского государственного университета леса – Лесной вестник. 2004. № 1. С. 5–9.
Экосистемы Теллермановского леса / Под ред. В.В. Осипова. М.: Наука, 2004. 340 с.
Dobbertin M. Tree Growth as Indicator of Tree Vitality and of Tree Reaction to Environmental Stress: a Review // European J. Forest Research. 2005. V. 124. P. 319–333.
Dobbertin M., Neumann M. Part V: Tree Growth. In: U-NECE ICP Forests, Programme Coordinating Centre (ed.): Manual on Methods and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects of Air Pollution on Forests. Thünen Institute of Forest Ecosystems, Eberswalde, Germany, 2016. 17 p.
Douglass A.E. Climatic Cycles and Tree-Growth: A Study of the Annual Rings of Trees in Relation to Climate and Solar Activity. Washington: Carnegie Institution of Washington, 1919. 127 p.
Eichhorn J., Roskams P., Potocic N. Timmermann V., Ferretti M., Mues V., Szepesi A., Durrant D., Seletković I., Schröck H.-W., Nevalainen S., Bussotti F., Garcia P., Wulff S. Part IV: Visual Assessment of Crown Condition and Damaging Agents. In: UNECE ICP Forests Programme Coordinating Centre (ed.): Manual on Methods and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects of Air Pollution on Forests. Thünen Institute of Forest Ecosystems, Eberswalde, Germany, 2016. 49 p.
Gričar J. Cambial Cell Production and Structure of Xylem and Phloem as an Indicator of Tree Vitality: a Review. In: Sustainable forest management: current research. Rijeka, Croatia: InTech, 2012. P. 111–134.
Helińska-Raczkowska L. Variation of Vessel Lumen Diameter in Radial Direction as an Indication of the Juvenile Wood Growth in Oak (Quercus petraea Liebl) // Annals of Forest Science. 1994. V. 51. P. 283–290.
Matveev S., Milenin A., Timashchuk D. The Effects of Limiting Climate Factors on the Increment of Native Tree Species (Pinus sylvestris L., Quercus robur L.) of the Voronezh Region // J. Forest Science. 2018. V. 64. № 10. P. 427–434.
Nowacki G.J., Abrams M.D. Radial-Growth Averaging Criteria for Reconstructing Disturbance Histories from Presettlement-Origin Oaks // Ecological Monographs. 1997. V. 67. № 2. P. 225–249.
Rubio-Cuadrado Á., Camarero J.J., del Río M. et al. Long-term impacts of drought on growth and forest dynamics in a temperate beech-oak-birch forest // Agricultural and Forest Meteorology. 2018. V. 259. P. 48–59.
Rybníček M., Čermák P., Žid T., Kolář T., Trnka M., Büntgen U. Exploring Growth Variability and Crown Vitality of Sessile Oak (Quercus petraea) in the Czech Republic // Geochronometria. 2015. V. 42. № 1. P. 17–27.
Scharnweber T, Heinze L, Cruz-Garcı ́a R, van der Maaten-Theunissen M., Wilmking M. Confessions of solitary oaks: we grow fast but we fear thedrought // Dendrochronologia. 2019. V. 55. P. 43–49.
Дополнительные материалы отсутствуют.