

Лесоведение, 2022, № 1, стр. 34-46
Мозаичность ценопопуляций черники и брусники, и динамика органического вещества почвы в сосняках Южного Подмосковья
Е. В. Зубкова a, *, П. В. Фролов a, С. С. Быховец a, М. А. Надпорожская b, Г. Г. Фролова a
a Институт физико-химических и биологических проблем почвоведения Российской академии наук,
ФИЦ ПНЦБИ РАН
142290 Пущино, Московская обл., Институтская ул., 2/2, Россия
b Санкт-Петербургский государственный университет
199034 Санкт-Петербург, Университетская наб., 7/9, Россия
* E-mail: zubkova@pbcras.ru
Поступила в редакцию 16.07.2020
После доработки 11.01.2021
Принята к публикации 21.07.2021
- EDN: XXSUNT
- DOI: 10.31857/S0024114821060127
Аннотация
Исследования пространственной неоднородности распространения ценопопуляций кустарничков показали, что доминирование черники (Vaccinium myrtillus L.) и брусники (V. vitis-idaea L.) в напочвенном покрове определяется экологическими факторами. В условиях выраженного микрорельефа при выровненной освещенности ограничивающим фактором пространственного распространения кустарничков выступает влажность почвы. Черника занимает более низкие участки микрорельефа с большей влажностью по сравнению с брусникой, при этом длительное затопление талыми водами в течение 1–2 недель приводит к гибели парциальных кустов черники. На участках перекрывания экологических ниш кустарнички успешно содоминируют, благодаря различию в морфо-физиологических особенностях этих видов. Показано, что в одноярусных сосновых лесах Южного Подмосковья без древесного подроста фактор освещенности не оказывает существенного влияния на рост кустарничков. Наличие второго древесного яруса ограничивает распространение кустарничков под кронами ели и липы: при коэффициенте пропускания фотосинтетически активной радиации (ФАР) менее 7% парциальные кусты отсутствуют; в диапазоне от 7 до 10% встречаются единичные побеги черники и брусники пониженной жизненности (высотой менее 0.1 м); при пропускании ФАР более 10% совместное покрытие черники и брусники достигает 60–80%. Вычислительные эксперименты по анализу сопряженной динамики растений травяно-кустарничкового яруса и почвы с помощью системы моделей “CAMPUS-S – EFIMOD – Romul_Hum” показали высокую точность работы системы моделей и сопоставимость результатов вычислений с натурными данными.
Одними из важнейших критериев оценки функционирования лесных экосистем являются видовое разнообразие растительности и содержание запасов органического вещества почв (ОрВП) (Morris et al., 1997). Проводимые инвентаризации лесов дают сведения об их статичном состоянии на момент исследований и не учитывают специфику сукцессионной изменчивости местообитаний (Рогозин, Разин, 2015). Для учета влияния изменений климата и антропогенных воздействий применяют методы математического моделирования (Комаров и др., 2007; Грабарник и др., 2019). Относительно хорошо разработаны вопросы моделирования главных составляющих лесных ценозов: ОрВП и древесной растительности (Омелько, 2006; Комаров и др., 2007; Tuomi et al., 2011; Seidl et al., 2012; Chertov et al., 2017a, b; Komarov et al., 2017). Перечисленные модели лесных экосистем не учитывают вклад напочвенного покрова в круговорот органического вещества. Исследования показали, что, хотя доля живого напочвенного покрова в общей биомассе леса невелика, он играет существенную роль в биологическом круговороте. Так, в сосновых лесах опад травяно-кустарничкового яруса (ТКЯ) составляет около 10% от массы общего опада, а доля азота в опаде кустарничков и трав гораздо выше – до 30% от азота всего поступающего опада (Родин, Базилевич, 1965; Emmer, 1995). В настоящее время математическое моделирование динамики живого напочвенного покрова, в частности ТКЯ, – активно развивающееся направление экосистемного моделирования (Mony et al., 2011; Oborny et al., 2012; Фролов и др., 2015; Маслов, Логофет, 2016, 2020; Фролов, Зубкова, 2017; Maslov, Logofet, 2020, Frolov et al., 2020a, b).
В разработанной нами модели CAMPUS-S (Frolov et al., 2020a, b), реализованы алгоритмы учета факторов, определяющих динамику роста кустарничков, что необходимо для получения оценки динамики ценопопуляций в условиях гетерогенности среды, в том числе по факторам: температуре и влажности почвы; содержанию в почве азота; освещенности под пологом леса в зависимости от характеристик породного и возрастного состава древостоя.
Для генерации необходимых климатических данных состояния почв используется статистический генератор климата почв Soil CLImate Statistical Simulator (SCLISS). Модель SCLISS (Быховец, Комаров, 2002; Комаров и др., 2007) предназначена для оценки средних месячных значений температуры, влажности лесной подстилки и минеральной почвы в разных типах леса по стандартным метеорологическим наблюдениям: температуре воздуха, осадкам, а также, при наличии данных, по температуре почвы под травяным покровом, измеряемой на метеорологических станциях.
Для учета влияния эдафических факторов используется модель динамики органического вещества почв Romul_Hum (Komarov et al., 2017; Chertov et al., 2017a, b). Модель описывает минерализацию и гумификацию опада и ОрВП; эмиссию углекислого газа и высвобождение азота в доступных для растений минеральных формах. Скорость минерализации и гумификации в модели определяется качеством опада, температурой и влажностью подстилки и минеральных горизонтов почвы. Отличительным свойством модели является то, что она описывает процесс трансформации ОрВП отдельно для лесной подстилки и верхних органоминеральных горизонтов почвы. Начальные значения запаса ОрВП и азота в пулах почвы считываются из входного файла модели Romul_Hum при инициализации модели CAMPUS-S. Из каждой ячейки модели CAMPUS-S в модель ROMUL_Hum передаются влажность и температура подстилки и почвы (на глубине 20 см), а также количество поступающего напочвенного и внутрипочвенного опада (листья, стебли, корневища, тонкие корни, с соответствующими содержанием азота и зольностью). Модель рассчитывает данные по почвенным пулам ОрВП и азота, а также эмиссию углекислого газа из почвы и продукцию азота в доступных для растений формах и передает данные в модель CAMPUS-S, где дальше они используются при расчетах роста растений.
Режим освещенности под пологом леса учитывается на основании данных из модели древесного полога EFIMOD (Komarov et al., 2003; Комаров и др., 2007; Shanin, et al., 2020): динамически передаваемой карты ФАР под пологом (моль м–2 мес.–1) в разрешении 0.25 м2 и координат стволов деревьев; области, соответствующие этим координатам считаются абсолютно недоступной территорией для растений ТКЯ.
Цель работы – изучить экологических факторы, определяющие мозаику расположения ценопопуляций черники и брусники, влияние абиотических и биотических условий на продуктивность ценопопуляций кустарничков, а также с применением вычислительных экспериментов оценить динамику органического вещества почвы в сосняках.
В ходе исследования решались задачи: 1) количественно оценить границы диапазонов толерантности кустарничков по освещенности и почвенным условиям (влажность, запасы азота в минеральной почве и лесной подстилке); 2) провести серию вычислительных экспериментов для анализа динамики ОрВП с применением моделей CAMPUS-S (Frolov et al., 2020a, b) и ROMUL-Hum (Komarov et al., 2017; Chertov et al., 2017a, b). От точности и согласованности полевых измерений в значительной степени зависят результаты прогнозов математических моделей и принимаемые на их основе стратегии природопользования. Для получения комплексных данных применены авторские методы полевых исследований и обработки результатов, поэтому методика работ описана подробно. Полученные сведения о распространении кустарничков при разной освещенности и влажности почв являются уникальными и позволяют проводить моделирование роста кустарничков в исследованных климатических и почвенных условиях. Подобные вычислительные эксперименты проведены впервые.
ОБЪЕКТЫ И МЕТОДИКА
Ключевые участки сосняков с парцеллами брусники и черники расположены на террасах р. Оки, сложенных плотными известняками каменноугольного периода, перекрытыми аллювиально-флювиогляциальными кварцевыми песками днепровской морены (Лидов, 1949). Исследование почвенных условий и освещенности проводилось на двух близко расположенных территориях: 1) в 80-летнем (Надпорожская и др., 2018) сосняке кустарничково-зеленомошном (9С1Б) Отрадинского участкового лесничества (ОУЛ) опытного лесного хозяйства “Русский лес”; 2) в 140-летнем (Шанин и др., 2018) сосняке сложном (4С4Б2Е + Лп) с елью (Picea abies (L.) Karst.) и липой (Tilia cordata Mill.) во втором ярусе (6Е2Лп1Д1Б) на территории Приокско-Террасного биосферного заповедника (ПТЗ). Территория в ОУЛ имеет слабоволнистый (дюнный) рельеф с мощностью песчаных отложений от 0.75 до 2 м (Лидов, 1949). Исследования в ОУЛ проводили на трех катенах (54°54′10″ с.ш., 37°39′56″ в.д.; 54°54′09″ с.ш., 37°39′57″ в.д.; 54°54′09″ с.ш., 37°40′02″ в.д.) с повторяющимся доминантами ТКЯ: вершины дюн – парцеллы с монодоминирование брусники, на склонах – содоминирование черники и брусники, нижняя часть склонов – парцеллы с монодоминированием черники. В ПТЗ была выбрана территория (54°53′17″ с.ш., 37°33′47″ в.д.) с выравненным рельефом, но с контрастными условиями освещенности вследствие неоднородности древесного полога. Под кронами елей и липы травяно-кустарничковый и моховой ярусы отсутствовали, в окнах лесного полога содоминировали черника и брусника. Детальное описание участков ОУЛ приведено в работе (Надпорожская и др., 2018), описание ключевого участка ПТЗ – в работе (Шанин и др., 2018). Названия лесных сообществ даны по (Ценофонд …, 2020).
В ОУЛ на всех трех катенах для каждой парцеллы заложены базовые почвенные разрезы по общепринятым методикам (Розанов, 2004), проведен отбор проб из почвенных генетических горизонтов. Отбор проб лесной подстилки и органоминеральных горизонтов почвы в каждом разрезе для каждого горизонта проводили в пятикратной повторности. Плотность сложения лесных подстилок определяли с помощью послойного отбора проб по всей мощности органогенных подгоризонтов с площади 0.25 × 0.25 м. Плотность сложения органоминеральных горизонтов оценивали с помощью режущего цилиндра (100 см3). Определены гигроскопическая влажность и потери при прокаливании (Аринушкина, 1970; Растворова 1983). Содержания валовых углерода и азота в образцах минеральной почвы (органоминеральный горизонт), лесной подстилки и органах кустарничков получены на анализаторе CHNS-932 фирмы LECO (США). Запасы ОрВП и валового азота для минеральных горизонтов рассчитывали по аналитическим определениям углерода и азота, плотности сложения и мощности почвенных горизонтов.
Для получения данных о режиме влажности корнеобитаемого слоя почвы для каждой синузии кустарничков на всех трех катенах измеряли влажность лесной подстилки и нижележащего минерального горизонта влагомером грунта МГ-44Э (Влагомер …, 2009). В центре каждой парцеллы был установлен деревянный колышек с номером, от него по направлениям на север, юг, запад и восток на расстоянии 0.2 и 0.4 м определяли влажность подстилки (на глубине 0.03–0.05 м) и органоминерального горизонта (на глубине 0.1–0.4 м). Измерения проводили при разных метеорологических условиях. Данные были пересчитаны в показатели объемной влажности, используемые в системе моделей SCLISS-CAMPUS.
Определение освещенности под лесным пологом проведено на пике вегетационной активности в июле. Для оценки светового режима применяли метод сверхширокоугольной фотографии пленочной камерой “Зенит ЕТ” с объективом “Зенитар” (фокусное расстояние 16 мм, поле зрения 180° по диагонали кадра). Съемку выполняли в зенит с высоты 0.15 м (т.е. со среднего уровня расположения фотосинтетического аппарата кустарничков) в 3-кратном повторении для каждой парцеллы (на тех же участках, что и измерения влажности). Пленку (Kodak чувствительностью 200 ед.) сканировали с разрешением 9600 точек на дюйм. Из изображения для дальнейшего анализа выделялся синий слой, дающий наилучший контраст между кронами и небом. Фотографии анализировали с использованием разработанной нами программы CaImAn (CAnopy IMage ANalyzer), позволяющей оценивать относительное пропускание кронами деревьев ФАР с учетом положения траектории движения Солнца по небосводу и соотношения прямой и рассеянной радиации. Для нахождения требовательности черники и брусники к условиям освещенности по такой же методике были обследованы участки напочвенного покрова в сосняке сложном в ПТЗ: а) с хорошо развитыми парциальными кустами (ПК) черники и брусники в окнах полога; б) в зоне единичных ПК на границе крон елей; в) под кронами группы елей, где ПК отсутствуют. Оба вида являются длиннокорневищными многолетними кустарничками (Полянская и др., 2000; Прокопьева и др., 2000) с приростами корневищ 0.1–0.2 м год–1, их ценопопуляции занимают исследуемую территорию длительный период времени (более 40 лет), следовательно, отсутствие ПК на участках сильного затенения говорит о непригодности там условий для их развития. Обследование вырубок показало, что в условиях полной освещенности гибели парциальных кустов черники и брусники не происходит.
Для подготовки входных данных дополнительно на одной из катен территории ОУЛ была заложена постоянная пробная площадь 20 × 25 м. Разметка проводилась с помощью буссоли и рулетки. Пробная площадь была отграничена по двум длинным сторонам стропами, натянутыми параллельно друг другу. Выравнивали стропы в единой плоскости при помощи лазерного дальномера-уклономера Bosch GLM 100C. Перпендикулярно основным стропам натягивали “плавающую” стропу. Все стропы размечали с шагом 0.2 м. В процессе исследований расстояние от “плавающей” стропы до уровня почвы на каждой отметке измеряли рейкой. Одновременно по той же решетке определяли доминирующие виды напочвенного покрова. Затем “плавающую” стропу перемещали на следующую отметку по “стационарным” стропам. С интервалом в 1 м измеряли влажность лесной подстилки влагомером на глубине 0.03–0.05 м и минерального горизонта на глубине 0.1–0.4 м (с учетом мощности лесной подстилки). На основании проведенных измерений была составлена план-схема рельефа, влажности и распределения ТКЯ на пробной площади (Frolov et al., 2020b).
Учет надземной и подземной биомассы кустарничков проведен на пробной площади весовым методом по монолитам (0.25 × 0.25 м × мощность корнеобитаемого горизонта). Монолиты отбирали вдоль границы пробной площади по всем элементам микрорельефа с шагом 1 м. На поверхности каждого монолита определено проективное покрытие кустарничков, проведен учет числа парциальных образований (побегов, кустов) каждого онтогенетического состояния. По четырем граням извлеченного монолита измеряли мощность лесной подстилки по подгоризонтам: L – слаборазложенный опад, F – ферментативный, H – гумусированный. Надземные части кустарничков состригали. Подземные органы отбирали из почвы, отмывали в проточной воде. Фракции биомассы разбирали по органам (листья, стебли, корневища, корни), высушивали в помещении до воздушно-сухого состояния, затем в сушильном шкафу при температуре 60°С.
Вычислительные эксперименты. При проведении вычислительных экспериментов в модель CAMPUS-S в качестве входных параметров экологических условий были введены: 1) карта микрорельефа (Frolov et al., 2020b), 2) динамическая карта освещенности, помесячно рассчитываемая в новой версии модели EFIMOD (Shanin et al., 2020), 3) ряды климатических данных (температура и влажность подстилки и поверхностных минеральных горизонтов почвы), полученные с помощью модели SCLISS (Быховец, Комаров, 2002; Комаров и др., 2007). Ранее на основе этих данных корректность работы модели была проверена по 7 параметрам: плотность ценопопуляций (шт. м–2), проективное покрытие, онтогенетические спектры ценопопуляций, локации особей, биомасса ценопопуляций (кг м–2), соотношение массы надземных и подземных органов, содержание углерода (Сорг) и азота (Nобщ) в подгоризонтах лесной подстилки и зоне максимальной плотности корней в минеральной почве (Frolov et al., 2020b).
Длительность вычислительных экспериментов составляла 80 лет. Имитировали случайное начальное распределение семян кустарничков обоих видов. Варьирование температуры и влажности на моделируемой территории задавали в диапазонах 5°C и 10% объемной влажности соответственно. Площадь моделируемой территории составляла 20 × 25 м. В вычислительном эксперименте оценивали динамику ОрВП в подстилке и минеральной почве в сосняке 1) с доминированием брусники, 2) черники и 3) содоминировании черники и брусники. Исходные почвенные запасы Сорг, Nобщ взяты по типичной для сосновых лесов ПТЗ почве (Фридланд, 1955) для слоя 0–0.15 м.
Для составления климатических сценариев были использованы данные по температуре воздуха, осадкам и температуре почвы за 1950–2017 гг. метеорологической станции Коломна (Булыгина и др., 2020; Шерстюков, 2020) и станции фонового мониторинга ПТЗ (Курганова и др., 2017).
Сценарии поступления опада растений отражают рост чистых сосновых древостоев (начиная с возраста 20 лет) для трех вариантов живого напочвенного покрова, типичных для значительной территории ПТЗ на момент начала заповедного режима (Оценка …, 2000). Начальный возраст древостоя выбран с учетом среднего возраста сосняков ПТЗ по данным лесной таксации 1940-х годов. Данные по динамике поступления фракций опада древостоя (с учетом их массы и содержания в них азота) получены в вычислительных экспериментах роста древостоев в модели EFIMOD (Комаров и др., 2007). Начальные таксационные характеристики древостоя взяты из региональных таблиц хода роста (ТХР) (Швиденко и др., 2008), соответственно: для сосняков – ТХР модальных сосновых древостоев Европейской части России (экорегионы южной тайги, зон смешанных, лиственных лесов и лесостепи), II бонитет, полнота 0.8, начальная густота 2000 шт. га–1. Видоспецифичное содержание азота в разных фракциях опада (хвоя или листья, ветви, тонкие корни, ствол, структурные корни) рассчитывает модель EFIMOD с учетом возраста древостоя и уровня трофности почв (Комаров и др., 2007). Количественные оценки поступления опада кустарничкового яруса выполнены с использованием модели CAMPUS-S. Аналогично EFIMOD, в модели CAMPUS-S учтены видоспецифичные различия в содержании азота для разных фракций опада рассматриваемых кустарничков. Динамику ОрВП оценивали в вычислительных экспериментах в модели Romul_Hum.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Оценка эдафических условий
На ключевом участке ОУЛ почвенный покров представлен сочетанием подзолов иллювиально-железистых (вершины и пологие склоны бугристо-дюнного рельефа, около 95% площади) с торфяно-подзолами иллювиально-гумусовыми глееватыми (понижения между буграми-дюнами) на аллювиально-флювиогляциальных песчано-супесчаных и щебнистых отложениях (табл. 1). Перепады высот между вершинами бугров и днищами понижений составляет 1.0–1.5 м.
Таблица 1.
Плотность сложения, запасы органического вещества и валового азота в почвах соснового леса ОУЛ на участках кустарничковых парцелл
| Горизонт | Мощность, м (n > 20) | Зольность, % (n = 3) |
Плотность сложения, кг м–3 | Запасы, кг м–2 | С : N (n = 3) |
Биомасса кустарничков, кг м–2 | |
|---|---|---|---|---|---|---|---|
| ОрВП | N вал. | ||||||
| Брусника. Почва: подзол иллювиально-железистый | |||||||
| L' | 0.016 а | 33.7 а | 50 а | 0.65 | 0.012 | 27 а | 0.026а |
| F | 0.017 а | 42.9 а | 80 б | 0.97 | 0.019 | 26 а | |
| H' | 0.017 а | 44.1 а | 100 в | 1.19 | 0.023 | 25 а | |
| Epir | 0.090 а' | 97.7 а' | 1350 а' | 5.10 | 0.17 | 15 а' | |
| Брусника-черника. Почва: подзол иллювиально-железистый | |||||||
| L | 0.024 а | 27.9 а | 50 а | 1.39 | 0.027 | 26 а | 0.150б |
| F | 0.029 б | 28.4 а | 80 б | 2.34 | 0.044 | 27 а | |
| H | 0.015 а, б | 46.1 а | 100 в | 1.52 | 0.029 | 26 а | |
| Epir | 0.11 б' | 97.0 а' | 1230 а' | 6.28 | 0.190 | 16 a' | |
| Черника. Почва: торфяно-подзол иллювиально-гумусовый глеевый | |||||||
| T' | 0.048 в | 27.4 а | 80 б | 2.88 | 0.060 | 24 а | 0.090в |
| T'' | 0.046 в | 33.8 а | 80 б | 2.92 | 0.061 | 24 а | |
| T''' | 0.039 в | 60.0 б | 270 г | 4.78 | 0.107 | 23 а | |
| Epir | 0.16 в' | 96.9 а' | 1490 а' | 4.43 | 0.19 | 12 б' | |
Примечания. Идентичные буквы обозначают отсутствие достоверных различий между средними значениями (критерий Стьюдента-Ньюмена-Келса. P < 0.05). Статистическая обработка проведена по двум группам данных: а) оценивали достоверность различий между мощностями органических подгоризонтов (буквы а, б, в, г); б) оценивали достоверность различий по суммарной мощности органических горизонтов (буквы с символом “ ' ”). Запасы ОрВП и N вал. – средние расчетные величины, их использовали как базовые для проведения вычислительных экспериментов. Варьирование входных параметров в группе моделей ROMUL производится методом Монте-Карло.
Бугристо-дюнный микрорельеф препятствовал сельскохозяйственному использованию этой территории, следов распашки в почвенных профилях нет. Сероватый цвет верхней части подзолистых горизонтов Еpir обусловлен наличием древесных углей, оставшихся после лесных пожаров. Лесные подстилки всех трех типов парцелл по зольности, содержанию валовых углерода, азота и их отношению (C : N) существенных различий не имеют. По общей мощности органических горизонтов в изученных парцеллах достоверно от других отличается чернично-сфагновая; некоторое различие по мощности подгоризонтов подстилок выявлено между брусничной и бруснично-черничной. При этом биомассы кустарничков по всем трем парцеллам различаются существенно. Минимальная биомасса кустарничков (0.026 кг м–2), определена для брусничной парцеллы (табл. 1). Биомасса кустарничков черничной парцеллы в 3.5 раз больше – 0.09 кг м–2. Максимальная биомасса кустарничков в бруснично-черничной парцелле – 0.15 кг м–2, что в 1.7 и 5.8 раз больше биомассы кустарничков брусничной и черничной парцелл соответственно. Проведенный корреляционный анализ (Надпорожская и др., 2018) показал отсутствие связи между биомассой кустарничков и мощностью лесных подстилок. Следовательно, интенсивность развития кустарничков имеет более сложные зависимости от других факторов среды. Расчетные данные по запасам ОрВП приведены по подгоризонтам лесных подстилок и для верхнего слоя минеральной части профиля почвы (табл. 1): для брусники – 0.09 м, брусника-черника – 0.11 м, черника – 0.16 м, по глубине максимального количества тонких корней.
На территории ОУЛ отмечено статистически значимое отличие отношений C : N на участках с доминированием черники от участков с содоминированием черники и брусники и доминированием брусники, не различавшихся между собой по этому параметру (рис. 1).
Рис. 1.
Соотношение содержания азота и углерода на участках с доминированием кустарничков: V_v-i – брусника , V_m – черника, V_v-i + V_m – брусника и черника, P_a –участки без растений ТКЯ под елью, T_c – участки без растений ТКЯ под липой. Подгоризонты лесной подстилки: L – слаборазложенный, F – ферментативный и H – гумусированный; горизонты минеральной почвы: Еpir – подзолистый, АУ – серогумусовый. На диаграмме размахов C : N высота прямоугольника равна интерквартильному размаху (IQR = Q3– Q1). Отрезок внутри этого прямоугольника обозначает медианное значение C : N на соответствующем участке. Концы отрезков, отходящих от торцов прямоугольника, имеют следующие координаты: верхний отрезок min(max x, Q3+ 1.5 × IQR), нижний отрезок min(min x, Q1– 1.5 × IQR). Значения, лежащие вне ограниченного этими отрезками интервала (выбросы), изображены в виде отдельных точек.
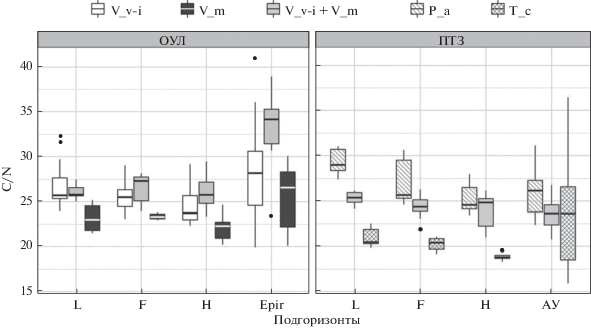
Полевое исследование влияния фитоценотических и эдафических факторов на распределение кустарничков в сосняках показало, что неоднородность напочвенного покрова формируется под воздействием комплекса факторов, динамически изменяющихся и взаимодействующих друг с другом.
Грубогумусные лесные подстилки, формирующиеся под сосновыми лесами на кварцевых песках, эвтрофируют почвы, что делает их более благоприятными для роста растений (Иванов, Шадриков, 2010). Наряду с близостью занимаемых экологических ниш по фактору почвенного богатства (Mäkipää, 1999) наши исследования подтвердили данные экологических шкал Д.Н. Цыганова (1983) и H. Ellenberg с соавторами (1991) о большей требовательности черники (рис. 1) к содержанию азота в субстрате (определенному с учетом мощности подстилки). Это свойство черники можно связать, с тем, что кустарничек сбрасывает листья на зиму, в отличие от брусники, длительность жизни листьев которой составляет 3–4 г. Экономия в расходовании пластических веществ позволяет бруснике занимать наиболее бедные участки вершин песчаных дюн.
Проведенные исследования почв (C : N) показали более бедные условия в минеральном горизонте ОУЛ по сравнению с ПТЗ (рис. 1). По нашему мнению, это является следствием как породного состава полога леса, так и влияния рекреации, приводящей к более частым низовым пожарам на территории ОУЛ (Надпорожская и др., 2018). Сравнение почв в ПТЗ под елью и липой показало более богатые условия под липой, что хорошо согласуется с данными о большем поступлении азота в почву с ее опадом в сравнении с елью (Ремезов и др., 1959); при этом под кустарничками значения были промежуточными (рис. 1). Анализ бруснично-черничных парцелл сосняка сложного (ПТЗ) и сосняка кустарничкового (ОУЛ) выявил сходные показатели валовых значений углерода и азота (рис. 1) в лесных подстилках, что является уникальной информацией и говорит об определяющем влиянии растений травяно-кустарничкового яруса на почвы.
Оценка влияния влажности почв
Объемная влажность, измеренная влагомером в разные сроки вегетационного периода, обобщена по 3 катенам (рис. 2), что позволило, несмотря на значительный разброс индивидуальных измерений, свести стандартную ошибку средних значений влажности к 1–2%. Ни в один из сроков измерения не выявлено значимых различий между участками пробных площадей “Брусника” и “Черника и брусника”, сезонный ход влажности здесь также был выражен слабо. На участках пробных площадей “Черника” значения влажности достоверно выше, и сезонные изменения влажности были существенны.
Рис. 2.
Сезонная динамика влажности органических и минеральных горизонтов почвы соснового леса ОУЛ на участках “Черника и брусника”, “Черника”, “Брусника” в 2016 г. (по измерениям влагомером).
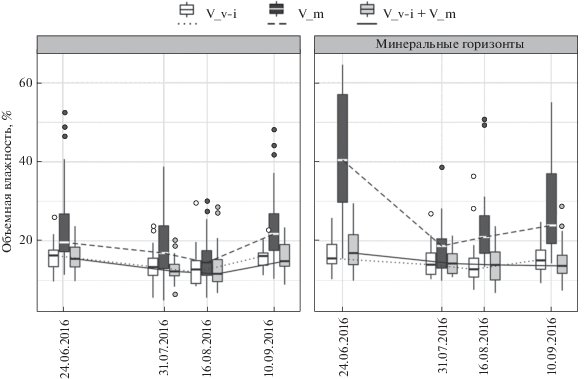
Исследование влияния влажности почв на рост кустарничков, проведенное в условиях дюнно-бугристого микрорельефа, выявило большую чувствительность брусники к повышению влажности, что можно объяснить более глубоким залеганием ее корневищ в минеральной почве по сравнению с черникой, корневища которой расположены преимущественно в нижних подгоризонтах лесной подстилки и лучше переносят более высокую влажность почвы. В полевых наблюдениях также было выявлено, что в понижениях рельефа, где весной застаиваются в течение 1–2 недель талые воды, происходит гибель парциальных кустов и корневищ черники.
Оценка влияния условий освещенности
Полученные в ОУЛ оценки относительной освещенности под пологом соснового леса показали отсутствие статистически значимого различия этого показателя для парцелл кустарничков. По исследованиям в ПТЗ выявлены следующие условия освещенности по встречаемости надземных побегов: при коэффициенте пропускания ФАР менее 7% ПК отсутствуют; в диапазоне от 7 до 10% встречаются единичные ПК черники и брусники пониженной жизненности (высотой менее 0.1 м); при пропускании ФАР более 10% совместное покрытие черники и брусники достигает 60–80%. Проведенные исследования показали сходность требовательности кустарничков к освещенности.
Таким образом, при составлении прогнозов развития лесных сообществ необходимо учитывать биологические особенности видов и их проявления в условиях пространственной неоднородности под пологом леса по освещенности и влажности почв.
Запасы органического вещества в почве при доминировании разных видов кустарничков. По результатам вычислительных экспериментов к 80 годам биомасса брусники на участках с монодоминированием составила 0.025 кг м–2, на участках с содоминированием – 0.12 кг м–2; биомасса черники на участках с монодоминированием составила 0.085 кг м–2, на участках с содоминированием – 0.024 кг м–2 (рис. 3).
Рис. 3.
Динамика биомассы ценопопуляций черники (V_m) и брусники (V_v-i) на участках с моно- и содоминированием.
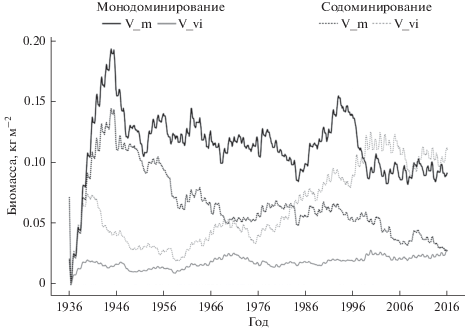
Демонстрируемое различие в динамике биомассы кустарничков при моно- и содоминировании объясняется тем, что экологические условия на большей части территории с содоминированием кустарничков оказываются ближе к оптимуму для брусники. Прирост биомассы черники в первые годы моделирования более, чем вдвое превышает прирост биомассы брусники, и это может быть связано с более высокой интенсивностью фотосинтеза при высокой доступности ФАР (в молодых сосняках).
Динамика ОрВП, вычисленная в ходе вычислительного эксперимента, приведена на рис. 4. Отношение запасов ОрВП, полученных в ходе вычислительных экспериментов, к измеренным составило 1.19 для участка с доминированием брусники, 0.94 для участка с доминированием черники и 0.93 для участка с содоминированием черники и брусники.
Рис. 4.
Динамика органического вещества почвы в органических и минеральных горизонтах на участках с монодоминированием черники, брусники и содоминированием обоих видов.
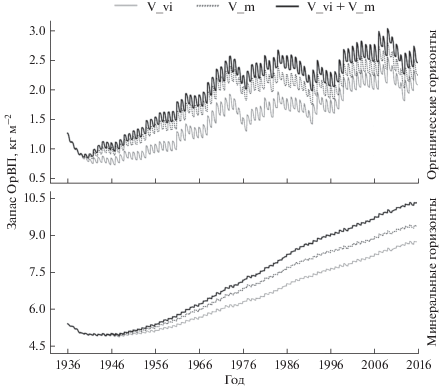
Так как в данном вычислительном эксперименте минимальный по количеству и концентрации в нем азота опад кустарничков был на участке с монодоминированием брусники, накопление органического вещества как в органических, так и в минеральных горизонтах почвы на этом участке происходило медленнее, чем на двух других. На участке с содоминированием черники и брусники количество опада было существенно выше, однако в связи с преимущественным вкладом брусники в суммарный опад концентрация азота в нем незначительно выше, чем на брусничном участке. Из-за этого происходит существенное накопление органического вещества в подстилке, однако в минеральной почве накопление усиливается незначительно по сравнению с брусничным участком. На участке с доминированием черники происходит существенное накопление органического вещества как в подстилке, так и в минеральной почве. Это можно объяснить лучшей водообеспеченностью, а также ускорением динамики ОрВП за счет поступления черничного опада, богатого азотом.
Результаты вычислительных экспериментов хорошо согласуются с полученными экспериментальными данными, что говорит о корректной работе и параметризации системы моделей “CAMPUS-S – EFIMOD – Romul_Hum”.
Заключение. Исследования, проведенные на территории ОУЛ, показали, что доминирование разных видов кустарничков определяется условиями среды обитания. Мозаика рельефа песчаных дюн, создающая контрастные условия влажности почв, становится причиной пространственной неоднородности распространения ценопопуляций черники и брусники. На участках перекрывания экологических ниш, кустарнички успешно содоминируют, благодаря разнице в морфо-физиологических особенностях строения и функционирования этих видов.
Также показано, что в одноярусных сосновых лесах без древесного подроста фактор освещенности не оказывает существенного влияния на рост кустарничков. Наличие в сосновых лесах ПТЗ второго древесного яруса ограничивает распространение кустарничков под кронами ели и липы, лимитирующих количество доступной ФАР. В условиях выраженного микрорельефа ОУЛ при выровненной освещенности ограничивающим фактором пространственного распространения кустарничков выступает влажность почвы. Брусника занимает более дренированные участки микрорельефа по сравнению с черникой.
Вычислительные эксперименты по анализу сопряженной динамики растений травяно-кустарничкового яруса и почвы с помощью системы моделей “CAMPUS-S – EFIMOD – Romul_Hum” показали высокую точность работы системы моделей и сопоставимость результатов ее вычислений с натурными данными.
* * *
Авторы благодарят руководство лесничества “Русский лес” и Приокско-Террасного заповедника за предоставленную возможность проводить исследования, а также коллег, оказавших помощь в работе: канд. техн. наук. В.О. Лопеса де Гереню, канд. биол. наук Т.Н. Лебедеву, канд. биол. наук В.Н. Шанина. Также авторы благодарят рецензентов за ценные замечания и рекомендации, которые существенно улучшили публикацию.
Список литературы
Аринушкина Е.В. Руководство по химическому анализу почв. М.: Изд-во МГУ, 1970. 487 с.
Булыгина О.Н., Разуваев В.Н., Александрова Т.М. Описание массива данных суточной температуры воздуха и количества осадков на метеорологических станциях России и бывшего СССР (TTTR). Свидетельство о государственной регистрации базы данных № 2014620942. URL: http://meteo.ru/data/162-temperature-precipitation# описание-массива-данных (дата обращения 20.02.2020).
Быховец С.С., Комаров А.С. Простой статистический имитатор климата почвы с месячным шагом // Почвоведение. 2002. № 4. С. 443–452.
Влагомер грунта “МГ-44”. Техническое описание и инструкция по эксплуатации. Харьков: ООО "Весоизмеритель”, 2009. 9 с. URL: http://vlagomer.at.ua/load/ instrukcija_po_ehkspluatacii_mg_44/1-1-0-7 (дата обращения 12.05.2020).
Грабарник П.Я., Шанин В.Н., Чертов О.Г., Припутина И.В., Быховец С.С., Петропавловский Б.С., Фролов П.В., Зубкова Е.В., Шашков М.П., Фролова Г.Г. Моделирование динамики лесных экосистем как инструмент прогнозирования и управления лесами // Лесоведение. 2019. № 6. С. 488–500.
Иванов И.В., Шадриков И.Г. Коэволюция почв и растительного покрова в южной тайге (на примере Приокско-Террасного заповедника) // Почвоведение. 2010. № 11. С. 1326–1333.
Комаров А.С., Чертов О.Г., Абакумов Е.В., Андриенко Г., Андриенко Н., Аппс М., Бобровский М.В., Бхатти Дж., Быховец С.С., Грабарник П.Я., Глухова Е.М., Зубкова Е.В., Зудин С.Л., Зудина Е.В., Кубасова Т.С., Ларионова А.А., Лукьянов А.М., Мартынкин А.В., Михайлов А.В., Морен Ф., Надпорожская М.А., Припутина И.В., Смирнов В.Э., Ханина Л.Г., Шанин В.Н., Шоу С. Моделирование динамики органического вещества в лесных экосистемах. М.: Наука, 2007. 380 с.
Курганова И.Н., Лопес де Гереню В.О., Аблеева В.А., Быховец С.С. Климат южного Подмосковья: современные тренды и оценка экстремальности // Фундаментальная и прикладная климатология. 2017. № 4. С. 62–78.
Лидов В.П. Геоморфологические особенности среднего течения долины р. Оки на примере Приокско-Террасного государственного заповедника и прилегающих территорий // Окская комплексная экспедиция. Тр. НИИ географии МГУ. М.: 1949. С. 179–190.
Маслов А.А., Логофет Д.О. Анализ мелкомасштабной динамики двух видов-доминантов в сосняке чернично-бруснично-долгомошном. I. Однородная марковская цепь и показатели цикличности // Журн. общей биологии. 2016. Т. 77. №. 6. С. 423–433.
Маслов А.А., Логофет Д.О. Совместная динамика популяций черники и брусники в заповедном послепожарном сосняке зеленомошнике. Модель с осредненными вероятностями перехода // Журн. общей биологии, 2020. № 4. Т. 81. С. 243–256.
Надпорожская М.А., Зубкова Е.В., Фролов П.В, Быховец С.С., Чертов О.Г. Соподчиненность почвенных условий и растительных сообществ в сосняках как следствие действия комплекса факторов // Вестник ТвГУ. Сер. Биология и экология. 2018. № 2. С. 122–138.
Омелько А.М. Модель роста деревьев темнохвойных пород на основе L-систем // Сибирский экологический журн. 2006. Т. 13. № 2. С. 181–188.
Оценка и сохранение биоразнообразия лесного покрова в заповедниках Eвропейской России. М.: Научный мир, 2000. 196 с.
Полянская Т.А., Жукова Л.А., Шестакова Э.В. Онтогенез черники обыкновенной (Vaccinium myrtillus L.) // Онтогенетический атлас лекарственных растений. Йошкар-Ола: МарГУ, 2000. Т. II. С. 51–59.
Прокопьева Л.В., Жукова Л.А., Глотов Н.В. Онтогенез брусники обыкновенной (Vaccinium vitis-idaea L.) // Онтогенетический атлас лекарственных растений. Йошкар-Ола: МарГУ, 2000. Т. II. С. 39–46.
Растворова О.Г. Физика почв (практическое руководство). Л.: Изд-во Ленингр. университета, 1983. 194 с.
Ремезов Н.П., Быкова Л.Н., Смирнова К.М. Потребление и круговорот азота и зольных элементов в лесах Европейской части СССР. М.: Изд-во МГУ. 1959. 284 с.
Рогозин М.В., Разин Г.С. Модели динамики и моделирование развития древостоев // Сибирский лесной журн. 2015. № 2. С. 55–70.
Родин Л.Е., Базилевич Н.И. Динамика органического вещества и биологический круговорот в основных типах растительности. М., Л.: Наука, 1965. 253 с.
Розанов Б.Г. Морфология почв. М.: Академический проект, 2004. 432 с.
Фридланд В.М. Почвы Приокско-Террасного государственного заповедника // Материалы по географии и генезису почв лесной зоны Европейской территории СССР. Труды Почвенного инст. им. В.В. Докучаева. М.: Изд-во АН СССР, 1955. Т. XLVI. С. 136–190.
Фролов П.В., Зубкова Е.В. Моделирование динамики биомассы растений травяно-кустарничкового яруса лесных экосистем // Сохранение лесных экосистем: проблемы и пути их решения. Киров: Радуга-Пресс, 2017. С. 334–339.
Фролов П.В., Зубкова Е.В., Комаров А.С. Клеточно–автоматная модель сообщества двух видов растений разных жизненных форм // Известия Академии наук, серия биология, 2015. № 4. С. 341–349.
Ценофонд лесов Европейской России (Электронный ресурс) URL: http://cepl.rssi.ru/bio/flora/ecoscale.htm (дата обращения 12.05.2020).
Цыганов Д.Н. Фитоиндикация экологических режимов в подзоне хвойно-широколиственных лесов. М.: Наука, 1983. 196 с.
Шанин В.Н., Шашков М.П., Иванова Н.В., Быховец С.С., Грабарник П.Я. Исследование структуры древостоев и микроклиматических условий под пологом леса на постоянной пробной площади в Приокско-Террасном заповеднике // Труды Приокско-Террасного заповедника. Вып. 7. М.: Изд. Товарищество научных изданий КМК, 2018. С. 72–85.
Швиденко А.З., Щепащенко Д.Г., Нильсон С., Булуй Ю.И. Таблицы и модели хода роста и продуктивности насаждений основных лесообразующих пород Северной Евразии (нормативно-справочные материалы). М.: Рослесхоз, Международный институт прикладного системного анализа, 2008. 886 с.
Шерстюков А.Б. Описание массива суточных данных о температуре почвы на глубинах до 320 см по метеорологическим станциям Российской Федерации. URL: http://meteo.ru/data/164-soil-temperature (дата обращения 20.02.2020).
Chertov O., Komarov A., Shaw C., Bykhovets S., Frolov P., Shanin V., Grabarnik P., Priputina I., Zubkova E., Shashkov M. Romul_Hum–A model of soil organic matter formation coupling with soil biota activity. II. Parameterisation of the soil food web biota activity // Ecological Modelling. 2017a. V. 345. P. 125–139.
Chertov O., Shaw C., Shashkov M., Komarov A., Bykhovets S., Shanin V., Grabarnik P., Frolov P., Kalinina O., Priputina I., Zubkova E. Romul_Hum model of soil organic matter formation coupled with soil biota activity. III. Parameterisation of earthworm activity // Ecological Modelling, 2017b. V. 345. P. 140–149.
Ellenberg H., Weber H.E., Düll R., Wirth V., Werner W., Paulißen D. Zeigerwerte von Pflanzen in Mitteleuropa (Indicator values of plants in Central Europe) // Scripta Geobotanica. 1991. V. 18. Verlag Erich Goltze KG, Göttingen. 248 s.
Emmer I.M. Humus form and soil development during a primary succession of monoculture Pinus sylvestris on poor sandy substrates. Amsterdam: Annals of Forest Science, 1995. 135 p.
Frolov P., Shanin V., Zubkova E., Bykhovets S., Grabarnik P. CAMPUS-S – The model of ground layer vegetation populations in forest ecosystems and their contribution to the dynamics of carbon and nitrogen. I. Problem formulation and description of the model // Ecological Modelling. 2020a. V. 431. Article 109184. P. 1–12.
Frolov P., Zubkova E., Shanin V., Bykhovets S., Mäkipää R., Salemaa M. CAMPUS-S – The model of ground layer vegetation populations in forest ecosystems and their contribution to the dynamics of carbon and nitrogen. II. Parameterization, validation and simulation experiments // Ecological Modelling. 2020b. V. 431. Article 109183. P. 1–14.
Komarov A.S., Chertov O.G., Zudin S.L., Nadporozhskaya M.A., Mikhailov A.V., Bykhovets S.S., Zudina E.V., Zoubkova E.V. EFIMOD 2 – a model of growth and elements cycling of boreal forest ecosystems // Ecological Modelling. 2003. V. 170. P. 373–392.
Komarov A., Chertov O., Bykhovets S., Shaw C., Nadporozhskaya M., Frolov P., Shashkov M., Shanin V., Grabarnik P., Priputina I., Zubkova E. Romul_Hum model of soil organic matter formation coupled with soil biota activity. I. Problem formulation, model description, and testing // Ecological Modelling. 2017. V. 345. P. 113–124.
Mäkipää R. Response patterns of Vaccinium myrtillus and V. vitis-idaea along nutrient gradients in boreal forest // J. Vegetation Science, 1999. V. 10. № 1. P. 17–26.
Maslov A.A., Logofet D.O., Bilberry vs. cowberry in a Scots pine boreal forest: III Another forest, another method, and similar conclusions // Ecological Modelling. 2020. V. 431, Article 109193.
Mony C., Garbey M., Smaoui M., Benot M.L. Large scale parameter study of an individual-based model of clonal plant with volunteer computing // Ecological Modelling. 2011. V. 222. № 4. P. 935–946.
Morris D.M., Kimmins J.P., Dan I., Duckert R. The use of soil organic matter as a criterion of the relative sustainability of forest management alternatives: A modeling approach using FORECAST // Forest Ecology & Management, 1997. V. 94. № 1–3. P. 61–78.
Oborny B., Mony C., Herben T. From virtual plants to real communities: a review of modelling clonal growth // Ecological Modelling. 2012. V. 234. P. 3–19.
Seidl R., Rammer W., Scheller R.M., Spies T.A. An individual-based process model to simulate landscape-scale forest ecosystem dynamics // Ecological Modelling. 2012. V. 231. P. 87–100.
Shanin V., Grabarnik P., Shashkov M., Ivanova N., Bykhovets S., Frolov P., Stamenov M. Crown asymmetry and niche segregation as an adaptation of trees to competition for light: conclusions from simulation experiments in mixed boreal stands // Mathematical & Computational Forestry & Natural-Resource Sciences. 2020. V. 12. № 1. P. 26–49.
Tuomi M., Rasinmäki J., Repo A., Vanhala P., Liski J. Soil carbon model Yasso07 graphical user interface // Environmental Modeling & Softwar. 2011. V. 26. № 11. P. 1358–1362.
Дополнительные материалы отсутствуют.