

Лесоведение, 2022, № 2, стр. 144-156
Формирование молодняков после рубок и пожаров в темнохвойных осоково-папоротниковых лесах Южного Сихотэ-Алиня
С. Г. Глушко a, *, Т. А. Комарова b, Н. Б. Прохоренко c
a Казанский государственный аграрный университет
420015 Казань, ул. К. Маркса, Республика Татарстан, Россия
b Федеральный научный центр биоразнообразия наземной биоты Восточной Азии ДВО РАН
690022 Владивосток, пр-т 100-летия Владивостока, 159/1, Россия
c Казанский (Приволжский) федеральный университет
420008 Казань, ул. Кремлевская, 18, Республика Татарстан, Россия
* E-mail: glushkosg@mail.ru
Поступила в редакцию 28.03.2021
После доработки 30.06.2021
Принята к публикации 01.12.2021
- EDN: XRVBHE
- DOI: 10.31857/S002411482202005X
Аннотация
Проведен сравнительный анализ формирования и естественного хода развития молодого поколения древесных пород после разных способов рубки и устойчивого низового пожара в зоне контакта пихтово-еловых и кедрово-широколиственных лесов Южного Сихотэ-Алиня. Рассмотрены особенности естественного возобновления, роста, развития, самоизреживания и динамики численности молодого поколения древесных пород в течение 33 лет на трех различных вырубках и на гари кедрово-темнохвойных осоково-папоротниковых лесов. Установлено, что при проведении рубок леса разными способами в кедрово-темнохвойных осоково-папоротниковых лесах Южного Сихотэ-Алиня наиболее быстрый и успешный лесовосстановительный процесс без смены коренных пород осуществлялся после зимней выборочной рубки. При сплошных рубках леса и низовых пожарах восстановление лесов идет через смену хвойных пород на быстрорастущие серийные лиственные породы. Наиболее длительный процесс восстановления кедрово-темнохвойных лесов происходит после сплошных рубок с использованием тяжелой техники, нарушающей напочвенный покров и подстилку, а также повреждающей подрост хвойных пород. Успешность лесовозобновительного процесса на обследованных участках связана с незначительной площадью вырубок и гари (каждая менее 1 га), сохранностью в той или иной степени подроста предварительной генерации, наличием на площадях семенников хвойных пород и близостью массивов коренных лесов. Практика проведения выборочных рубок в лесах Южного Сихотэ-Алиня при условии сохранения подроста предварительной генерации обеспечивает успешное формирование молодняков. Материалами наших исследований подтверждается успешный характер формирования молодняков на участках, пройденных выборочными рубками.
Лесообразовательный процесс в горных районах Приморского края протекает под влиянием периодически возникающих пожаров и возрастающих рубок главного пользования. После запрета рубок главного пользования в лесах, образованных сосной корейской, или кедром корейским (Pinus koraiensis Siebold & Zucc.), главным объектом рубок в Приморском крае стали темнохвойные пихтово-еловые леса. Лесные пожары и промышленные рубки привели к значительному уменьшению площади пихтово-еловых лесов. К настоящему времени все промышленно и технически значимые лесные ресурсы сильно истощены, а неосвоенные лесные массивы незначительны, приурочены к крутосклонам и сосредоточены на севере края (Ковалев, Алексеенко, 2018).
В связи с интенсивным процессом лесозаготовок и часто возникающими пожарами вопросы воспроизводства лесов и сохранения биоразнообразия лесных экосистем остаются актуальными в настоящее время (Современное состояние лесов …, 2009; Лукина и др., 2015; Ковалев и др., 2019; Kuuluvainen et al., 2019; и др.).
Сведения о естественном возобновлении пихтово-еловых лесов на Дальнем Востоке после рубок и пожаров содержатся во многих публикациях (Соловьев, 1963; Манько, Ворошилов,1967; Ворошилов, 1972; Ворошилов, 1975; Чумин и др., 1981; и др.). Несмотря на общую разработанность естественного хода восстановления лесов Дальнего Востока, недостаточно изучены особенности формирования и последовательного хода развития молодого поколения древесных пород после разных способов рубки и пожаров.
Цель наших исследований – провести сравнительный анализ характера и темпов восстановления древесных молодняков в первые 32–33 г. их развития после разных способов рубки и воздействия пожара в кедрово-темнохвойных осоково-папоротниковых лесах Южного Сихотэ-Алиня.
ОБЪЕКТЫ И МЕТОДИКА
Сообщества кедрово-темнохвойных осоково-папоротниковых лесов распространены в Южном и Среднем Сихотэ-Алине в зоне контакта широколиственно-кедровых и темнохвойных пихтово-еловых лесов. Встречаются они на пологих и среднекрутых склонах, а также на плоских водоразделах в пределах высотных отметок от 600 до 1000 м над ур. м. Произрастают на влажных горно-таежных иллювиально-гумусовых почвах, для которых характерна глубокая гумусированность всего почвенного профиля, высокая кислотность (рH 4.2–5.2) и насыщенность основаниями. На поверхности почвы накапливается достаточно мощный слой грубогумусной подстилки, что свидетельствует о замедленном темпе круговорота веществ в этих лесах.
Коренные насаждения кедрово-темнохвойного осоково-папоротникового типа леса отвечают природе южно-таежных темнохвойных лесов с участием сосны корейской и характеризуются простым строением и однообразным составом во всех ярусах. В коренных древостоях обычно доминируют ель аянская (Picea ajanensis Fisch. Ex Carr.) и пихта белокорая (Abies nephrolepis (Trautv.) Maxim.). Единичные, но крупные деревья сосны корейской в коренных лесах мало уступают по запасам древесины темнохвойным породам. Незначительное участие в древостое принимают широколиственные породы – липа Таке, клены желтый и зеленокорый (Tilia taquetii C.K. Schneid., Acer ukurunduense Trautv. et Mey., A. tegmentosum Maxim.). Древостои обычно развиваются по III классу бонитета, имеют относительную полноту в среднем 0.9–1.0 и достигают запасов древесины до 450 м3 га–1.
В естественном возобновлении древесных пород обычно преобладает подрост ели аянской и пихты белокорой, на долю которых приходится от 40 до 80%.
Кустарниковый ярус в коренных лесах преимущественно сформирован слабо, и общее покрытие его не превышает 30%. В нем встречаются единичные экземпляры бересклета большекрылого (Euonymus macroptera Rupr.), жимолости Максимовича (Lonicera maximowiczii (Rupr.) Regel), спиреи березолистной (Spiraea betulifolia Pall.) и других видов. В первые 5–7 лет после рубок и пожаров обильно разрастается бузина кистистая (Sambucus racemosa L.). Сравнительно успешно развиваются растения деревянистой лианы актинидии коломикта (Actinidia kolomikta Maxim.), образующие густые заросли в первые 20–30 лет после пожара. В дальнейшем большая часть скелетных осей актинидии не превышает 50–100 см длины, а проективное покрытие их надземных частей достигает 3–5%.
Кустарничково-травяной ярус хорошо развит, в коренных сообществах общее проективное покрытие его составляет в среднем 80–90%. В нем преобладают мезогигрофитные папоротники (Leptorumohra amurensis (Christ) Tzvel., Dryopteris expansa (C.Presl) Fras и др.) и осока мечевидная (Carex xуphium Kom.). Моховой покров редкий, образован небольшими пятнами из мезогигрофитных зеленых мхов (Rhytidiadelphus triquetrus (Hedw.) Warnst., Pleurozium schreberi (Brid.) Mitt. и др.), приуроченных к приствольным микроповышениям и полуразложившемуся валежу.
Исследования проводили на территории Верхнеуссурийского биогеоценотического стационара, образованного в 1973 г. сотрудниками Отдела леса Биолого-почвенного института (БПИ) ДВНЦ АН СССР. Стационар занимает площадь около 4.5 тыс. га и расположен в бассейне р. Правая Соколовка (приток р. Уссури) на западном макросклоне Южного Сихотэ-Алиня в пределах высотных отметок от 440 до 1100 м над ур. м (44°03′55″ с.ш., 134°20′29″ в.д.).
Для сравнительного анализа особенностей развития и темпов роста древесных молодняков после разных способов рубки и воздействия пожара в кедрово-темнохвойных лесах нами были использованы материалы, собранные на четырех пробных площадях (пр. пл.), лесовосстановительный процесс на которых осуществлялся в течение 32–33 лет.
Согласно “Правилам рубок главного пользования” (1970) в конце двадцатого столетия на территории Приморского края основными способами рубок главного пользования были приняты подневольно-выборочные и условно-сплошные. До организации Верхнеуссурийского стационара на этой территории активно проводились подневольно-выборочные рубки с удалением крупномерных стволов хвойных деревьев. Для максимального сбережения хвойного подроста в процессе лесозаготовок применялась Приморская узкопасечная технология, позволяющая сохранять подрост до 80% при зимних и 60% – при летних лесозаготовках. На узких волоках (шириной 25–35 м) вырубались все деревья, и по ним проводились трелевочные работы с помощью тракторов или лебедок. Между трелевочными волоками с помощью бензопил вырубались крупномерные деревья. В период зимних заготовок травмирование почвенного покрова и площадь минерализованных участков на волоках была минимальна.
Такая зимняя подневольно-выборочная рубка кедрово-темнохвойного осоково-папоротникового леса была проведена в 1970 г. на пологой надпойменной террасе ручья Медвежьего на высоте 650 м над ур. м. Всего здесь на площади 1 га было удалено 48 деревьев ели аянской от 60 до 85 см диаметром в основании ствола, а также 12 стволов сосны корейской 52–110 см диаметром и 4 ствола березы желтой, или ребристой (Betula costata Trautv.) 96–120 см диаметром. В центральной части заложенной здесь в 1987 г. пр. пл. 54-1987 размером 50 × 50 м2 сохранились следы трелевочного волока, по которому удалялись стволы срубленных деревьев. Последующие ревизии на этом участке были проведены в 2003 и 2007 гг.
Лесовосстановительный процесс изучался также после условно-сплошной рубки кедрово-темнохвойного осоково-папоротникового леса, проведенной в 1973 г. на плоском водоразделе, на высоте 860 м над ур. м. Здесь в 1988 г. была заложена пр. пл. 5-1988 размером 50 × 50 м2, которая соответствовала возрасту 15-летней вырубки. Во время рубки на площади 0.25 га сохранили жизнедеятельность 3 дерева ели аянской, 1 дерево сосны корейской и 7 деревьев пихты белокорой. Ревизии на этом участке были проведены нами в 2006 и 2011 гг.
Кроме того, процесс лесовосстановления в рассматриваемом типе леса изучался после сплошной экспериментальной рубки на площади 0.39 га в бассейне ключа Елового в верхней части северо-западного склона 10°–20° крутизной и на высоте 750–770 м над ур. м. Экспериментальная рубка была проведена летом 1976 г. сотрудниками Отдела леса БПИ ДВНЦ АН СССР без применения тяжелой техники. Валка деревьев производилась, начиная с 12 см ступени толщины, бензопилами “Дружба”, раскряжевка и обрубка сучьев проводилась на месте. Все операции выполнялись с максимальной предосторожностью для сохранения тонкомера и подроста хвойных пород от повреждений. Здесь была заложена постоянная пр. пл. 10-1976 размером 50 × 50 м2. В дальнейшем мы проводили ревизии в течение 7 лет ежегодно, а затем через различные интервалы. При обсуждении результатов исследования использованы материалы 2008 г., на 32 год после рубки.
Для сравнительного анализа характера развития и хода роста древесного молодняка, формирующегося не только на вырубках, но и после воздействия пожара в насаждениях рассматриваемого типа леса, нами были использованы материалы, собранные на послепожарном участке, образовавшемся после устойчивого низового пожара летом 1973 г. в верхней сравнительно плоской части в верховьях ручья Березового. По характеру и степени повреждения огнем образовавшаяся гарь соответствовала типу мелких гарей (Шешуков, 1967) и относилась к типу гарей с отмершими нижними ярусами и частично сохранившимся жизнедеятельным древостоем, полнота которого ниже 0.4 (Соловьев, Солодухин, 1975). На заложенной здесь постоянной пр. пл. 11-1975 площадью 0.25 га в древостое сохранили жизнедеятельность около 18% деревьев, главным образом ели аянской, пихты белокорой и липы Таке. Мелкий и средний подрост полностью сгорел, но сохранились 1.16 тыс. экз. га–1 растений крупного подроста, в котором преобладали растения пихты белокорой, ели аянской и липы Таке. Ревизии на пр.пл. 11-1975 проводили ежегодно в течение первых 14 лет, а затем продолжали с разными интервалами. Для сравнительного анализа были использованы материалы ревизии 2006 г., который соответствовал 33 г. после пожара.
При характеристике растительных сообществ были использованы общепринятые геоботанические и лесоводственные методики (Сукачев, Зонн, 1961; Корчагин, 1976; Мелехов, 1980; и др.). На всех сравниваемых участках проводили детальные геоботанические описания, устанавливали видовой состав и определяли количественное участие у каждого вида. Для оценки древостоя проводили сплошной перечет деревьев на всей площади по одно- двух- и четырехсантиметровым ступеням толщины. Определение высоты, возраста и хода роста древесных растений осуществляли по моделям разных ступеней толщины за пределами пробных площадей.
Учет подроста проводили на двух ленточных площадках (50 × 4 м), расположенных по диагоналям пробных площадей, по различным ступеням высоты: до 50 см (мелкий), от 51 до 150 (средний) и от 151 до 200 см (крупный). Всходы учитывались отдельно. Деревья высотой более 2 м и диаметром менее 12 см относили к тонкомеру, а диаметром более 12 см – к крупным деревьям.
Расчет таксационных показателей выполнен с использованием Справочника лесоустроителя Дальнего Востока (1973).
Для краткого обозначения древесных растений нами были использованы следующие обозначения: Бж – береза желтая, или ребристая, Бп – береза плосколистная, Бш – береза шерстистая, Вм – вишня Максимовича, Еа – ель аянская, Ивк – ива козья, Ивп – ива поронайская, Ил – ильм лопастный, К – сосна корейская, или кедр, Клж– клен желтый, Клз – клен зеленокорый, Клм – клен мелколистный, Лт – липа Таке, Ос – осина, П – пихта белокорая, Рс – рябина сибирская, Тс – тис остроконечный, Тк – тополь корейский, Чм – черемуха Маака, Я – ясень маньчжурский.
Названия растений приведены по сводке “Сосудистые растения Советского Дальнего Востока” (1985–1996).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Лесовосстановительный процесс после рубок и пожаров зависит от степени нарушения растительности и почвы, площади вырубки и гари, а также определяется численностью сохранившихся деревьев, подроста и близостью обсеменителей. Состав формирующихся сообществ определяется видовым составом исходных фитоценозов, поступлением диаспор с соседних участков и наличием жизнеспособных семян в почве, на что указывали многие исследователи (Соловьев, 1963; Манько, Ворошилов, 1967; Комарова и др., 2017; и др.).
В формировании разных видов берез важное место принадлежит всхожим семенам, покоящимся в почве и выполняющим роль резерва, который активизируется при нарушении растительного покрова и подстилки. Cогласно ранее проведенным исследованиям по проращиванию всхожих семян в почвенных образцах, взятых в кедрово-темнохвойном осоково-папоротниковом лесу в возрасте около 200 лет, было отмечено на 1 м2 100 всхожих семян березы шерстистой (Betula lanata (Regel) V.Vassil.), 59 семян березы желтой и 28 всхожих семян березы плосколистной (Комарова и др., 2017, с. 336).
Виды растений с ускоренным развитием на первых этапах онтогенеза мы относим вслед за Ф. Клементсом (Clements, 1928) к группе серийных видов. Высокие темпы роста серийных видов и быстрое достижение генеративного периода обеспечивают их первому поколению устойчивые позиции и возможность доминировать в производных сообществах. Последующие поколения не развиваются активно без отсутствия очередных нарушений в растительном покрове и присутствуют только в виде примеси. Среди серийных видов обычно первыми заселяют освободившиеся участки после рубки и пожаров разные виды берез. Березы желтая и шерстистая, отличаясь большей теневыносливостью по сравнению с березой плосколистной и другими серийными видами, обладают способностью возобновляться семенным путем под пологом древостоя преимущественно на разлагающемся валеже, но основная их часть погибает на 2–4 годы развития. Быстро заселяют гари и вырубки представители семейства Ивовые (осина обыкновенная Populus tremula L., тополь корейский Populus koreana Rehder, ива козья Salix caprea L., ива тарайкинская Salix taraikensis Kimura и др.). благодаря массовому распространению мелких и легких семян с помощью ветра и быстрому прорастанию. При наличии достаточной влаги в почве семена могут прорастать уже на 2-й день.
Анализ годичных приростов по высоте у молодых растений серийных и климаксовых (хвойных и лиственных) пород в первые годы их развития на гари (пр. пл. 11-1975) и экспериментальной вырубке (пр. пл. 10-1976) показал значительные различия по темпам роста (рис. 1).
Рис. 1.
Изменение среднего годичного прироста в высоту у молодых древесных растений в первые 6 лет их развития на экспериментальной вырубке (пр. пл. 10-1976; а) и после устойчивого низового пожара (пр. пл. 11-1975; б) кедрово-темнохвойного осоково-папоротникового леса.
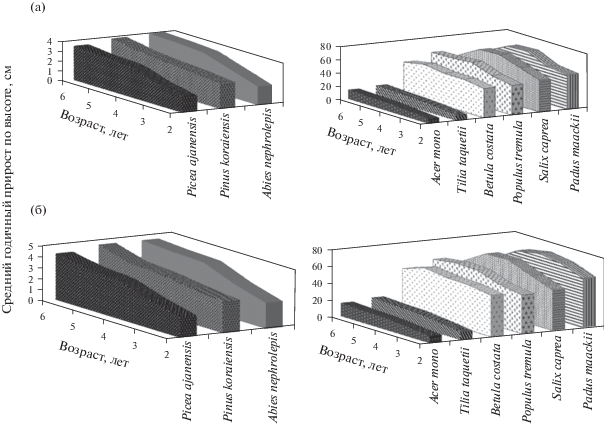
Средний прирост по высоте у растений серийных видов (береза желтая, осина, ива козья, черемуха Маака Padus maackii (Rupr.) Kom. & Aliss.) в первые 6 лет их развития составил на послепожарном участке от 46 до 72 см год–1, а на – вырубке – от 43 до 67 см год–1. У семенных растений климаксовых широколиственных пород (липа Таке и клен мелколистный) годичный прирост на гари составил 9–18 см год–1, а на вырубке – 7–15 см год–1. У хвойных растений (ель аянская, пихта белокорая и сосна корейская) отмечались самые низкие темпы роста – от 1.8 до 4.5 см год–1 на послепожарном участке и от 1.6 до 4.0 см год–1 на вырубке. Среди хвойных видов наиболее высокие показатели годичного прироста были у растений сосны корейской, а самые низкие – у ели аянской как на гари, так и на вырубке. При этом пихта белокорая превышает ель аянскую не только в приросте по высоте, но и по радиальному приросту до начала генеративного периода (Трофимова, 2012).
Все молодые древесные растения, произраставшие на гари, имели более высокие показатели роста по сравнению с растениями, росшими на вырубках, что было связано с обогащением почвы зольными веществами после сгорания растений, валежа и подстилки, а также увеличением pH и концентрации минеральных и органических соединений (Algrren, Algrren, 1960; Viro, 1974; Сапожников и др., 2001; и др.). На вырубках же изъятие значительной части зеленой массы и стволовой древесины, наоборот, снижает поступление органических веществ в почву.
Семенное возобновление древесных растений активно происходит преимущественно в первые 2 г. после рубки и пожаров (Манько, Ворошилов, 1967; Ворошилов, 1975; Комарова, 1986; и др.). Вместе с тем, на прорастание семян ели и пихты неблагоприятное влияние оказывает высокая зольность почвы, возникающая сразу же после пожаров (Сысоев, 1959; Данилик, 1965; Комарова, Ильина, 2002; и др.). Согласно исследованиям Е. Сысоева (1959) всхожесть семян ели сохраняется при содержании золы в почве не более 2 кг на 1 м2. Согласно нашим исследованиям (Комарова, Ильина, 2002), на однолетней гари темнохвойно-кедрового леса не сформировалось ни одного проростка ели аянской и пихты белокорой, но на второй год после пожара началось массовое и обильное их прорастание.
На второй год зарастания гари (пр. пл. 11-1975) численность всходов у быстрорастущих деревьев березы шерстистой составила 130 экземпляров, у березы желтой – 2079, а у березы плосколистной – 2132 на площади 50 м2 (Комарова, 1986). Молодые растения быстрорастущих серийных видов к 5–7 гг. после пожара сформировали сравнительно сомкнутые биогруппы, в которых преобладали растения березы желтой. При этом 60% молодых растений этого вида (из общего числа 11.8 тыс. экз. га–1) принадлежали группе замедленного роста, высота которых не превышала 1.5 м, в то время как растения усиленного роста, составлявшие 20%, достигали высоты 3.0–3.5 м. У других серийных видов также преобладали растения с замедленным ростом.
По мере смыкания крон древесного молодняка серийных видов усиливалась интенсивность конкуренции между растениями за жизненное пространство – свет, влагу, элементы минерального питания. Слабые растения, отстававшие в росте, подавлялись более крупными и погибали. Процессы самоизреживания древесных молодняков изучали многие исследователи (Гулисашвили, 1974; Мелехов,1980; и др.), однако большая часть их данных связана с отпадом молодых деревьев, начиная с 20–40-летнего возраста насаждений. Нами проводился учет численности древесных растений за 32-летний период на опытной вырубке кедрово-темнохвойного леса (пр. пл. 10-1976, рис. 2а) и в течение первых 33 лет на послепожарном участке (пр. пл. 11-1975, рис. 2б).
Рис. 2.
Изменение численности популяций хвойных климаксовых (1–3), широколиственных климаксовых (4–6) и серийных (7–9) древесных видов за 32-летний период после экспериментальной рубки (пр. пл. 10-1976; а) и после устойчивого низового пожара за 33-летний период (11–1975; б) лесовосстановительного процесса в кедрово-темнохвойных осоково-папоротниковых лесах. 1– Picea ajanensis, 2 – Abies nephrolepis, 3 – Pinus koraiensis, 4 – Tilia taquetii, 5 – Acer ukurunduense, 6 – Acer tegmentosum, 7– Betula costata, 8 – Salix caprea, 9 – Padus maackii.
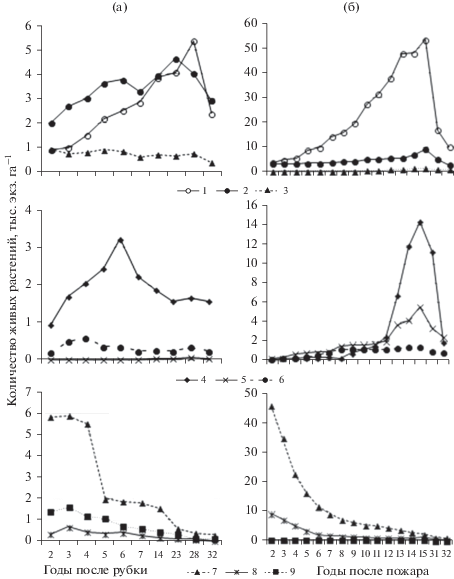
По мере роста серийных видов на гари происходило значительное снижение численности молодых растений, и на 15-й год после пожара сохранили жизнедеятельность только 23.5% молодых растений черемухи Маака, 13.6% растений ивы козьей и 5.0% деревьев березы желтой от количества растений, установленных на второй год после пожара. У остальных серийных видов также значительно снизилась численность растений.
В то же время количество растений климаксовых видов, напротив, возрастало, и на 15-й год после пожара численность молодого поколения липы Таке семенного и вегетативного происхождения возросла в 54 раза, клена желтого – в 29 раз, сосны корейской – в 26 раз, ели аянской – в 15 раз, а у пихты белокорой увеличилась в 3 раза по сравнению с количеством их на двухлетней гари. Постоянному нарастанию численности хвойных и широколиственных климаксовых видов способствовали сохранившие жизнедеятельность после пожара деревья и примыкающие с трех сторон к этой гари ненарушенные участки кедрово-темнохвойного леса.
На экспериментальной вырубке (пр. пл. 10-1976) отмечалась иная динамика численности древесных растений. Сохранившиеся здесь напочвенный покров и подстилка на большей части территории препятствовали активному заселению вырубки инициальными видами травянистых растений и кустарников, а также прорастанию всхожих семян берез из почвенных банков. На второй год после экспериментальной рубки было отмечено всего 7.8 тыс. га–1 молодых растений берез в отличие от 68.7 тыс. га–1 берез на двухлетней гари.
Успешному лесовозобновительному процессу на этой вырубке способствовал сохранившийся тонкомер и подрост хвойных пород предварительной генерации. До рубки здесь преобладал мелкий и средний подрост пихты белокорой в возрасте от 5 до 40 лет, максимум их приходился на группу возраста 16–20 лет. Мелкого и среднего подроста ели аянской было в три раза меньше, чем пихты, и средний их возраст составлял 15 лет. Еще меньше было подроста сосны корейской со средним возрастом 17 лет.
На второй год после рубки в мелком и среднем подросте с общей численностью 15.5 тыс. экз. га–1 преобладали растения березы желтой (5.3 тыс. экз. га–1) и пихты белокорой (2.0 тыс. экз. га–1). Средний подрост пихты полностью состоял из растений предварительной генерации. В крупном подросте (1.3 тыс. экз. га–1) преобладали сохранившиеся растения клена желтого (1.0 тыс. экз. га–1), а пихта и ель отсутствовали.
У серийных видов вслед за интенсивным ростом в первые 5–7 лет происходило активное естественное самоизреживание в густых биогруппах. На 7-й год после рубки сформировался одноярусный древесный молодняк 2.5–3 м высотой и 2–4 см диаметром, имеющий состав 8(Бш + Бж + + Бп)1Ос1Чм + Ивк.
Хвойные растения последующей генерации развивались медленно, и к 7-му году развития они стали преобладать в мелком подросте. При этом мелкий хвойный подрост предварительной генерации переместился в группу среднего подроста, а из среднего хвойного подроста образовался крупный подрост. К 7-му году у всех растений хвойных и широколиственных климаксовых видов возросла численность: ели – на 31.6%, липы Таке – на 41.4%, пихты – на 61%, клена желтого – на 72%, клена зеленокорого – на 75.4% по сравнению с количеством их на 2-летней вырубке.
На 15 год после экспериментальной рубки сформировавшийся древесный молодняк, в составе которого присутствовали растения 15 видов, имел состав по запасу: 2Бж2Клж2П2Чм1Бш1Лт + К, Еа, Бп, Ивк, Ос ед. Клз, Рп, Тс, Вм – при общей численности живых стволов – 5445 экз. га–1 и средних для преобладающих пород размеров стволов – 7.5 м высотой и 4.6 см диаметром, сумма площадей сечения составляла 8.17 м2, а запасы древесины – 35.5 м3.
В то же время древесный молодняк, сформировавшийся на 15 год после условно-сплошной рубки (пр. пл. 5-1988), включающий также 15 древесных видов, имел состав по запасу: 4Бш2Ивк1Бж1Бп1Еа1Чм + П, Тк, Клж ед. Ивп, Клм, К, Лт, Рп, Ос при общей численности живых стволов 4220 экз. га–1 и средних их показателей для преобладающих пород – 8.1 м высотой и 6.0 см диаметром, сумма площадей сечения составляла 4.15 м2, а запас – 24.2 м3.
Следовательно, у молодняка, сформировавшегося после условно-сплошной рубки, в отличие от древесного молодняка, образовавшегося на экспериментальной вырубке, были значительно ниже показатели численности стволов, суммы площадей сечения и запасов древесины, что, вероятно, было связано с активным заселением этой вырубки инициальными травянистыми и кустарниковыми видами и дальнейшим их мощным разрастанием. Все это препятствовало активному естественному возобновлению древесных видов. Успевшие сформироваться в первые годы после условно-сплошной рубки немногочисленные растения быстрорастущих серийных видов быстро развивались в более разреженном древостое.
В составе и структуре сравниваемых древесных молодняков на трех различных вырубках и послепожарном участке, образовавшихся в течение 32–33 лет на месте кедрово-темнохвойных осоково-папоротниковых лесов, отмечались как общие черты, так и существенные различия.
К общим чертам относится незначительная площадь (менее 1 га) у всех четырех нарушенных рубкой и пожаром участков, окруженных стенами коренных лесов. Это определило во всех сравниваемых древесных молодняках доминирование ели аянской и пихты белокорой во всех категориях подроста и тонкомера (табл. 1). При этом максимальной численностью (7–20 тыс. экз. га–1) отличался мелкий подрост, а наименьшее количество экземпляров (0.4–0.8 тыс. экз. га–1) было представлено в крупном подросте. Для всех насаждений был характерен сравнительно близкий состав всходов, в котором доминировали разные виды берез, произрастающие, как правило, на гниющей древесине валежа и реже пней. Меньшей численностью были представлены всходы широколиственных пород (липы Таке, кленов желтого и зеленокорого), большая часть которых также была приурочена к гниющему валежу. Среди крупных деревьев доминировали березы шерстистая и желтая. Исключение составили пр. пл. 54-1987, где была проведена подневольно-выборочная рубка с помощью узкопасечного способа, а также пр. пл. 11-1975, где после пожара среди подроста, тонкомера и даже крупных деревьев (с диаметром ствола более 12 см) по численности преобладали растения ели и пихты (табл. 1).
Таблица 1.
Количество (тыс. экз. га–1) и состав (по числу стволов) всходов, мелкого, среднего и крупного подроста, а также тонкомерных и крупных деревьев в насаждениях, сформированных в течение 32–33 лет после разных способов рубок и низового пожара
| Категории крупности | № пр. пл | n, тыс. экз. га–1 | Породный состав (по числу стволов) |
|---|---|---|---|
| Всходы | 54-1987 | 4.90 | 4Б(ж + пл)3Клж1Лт1К1Кз ед. П, Еа, Км, Я |
| 5-1988 | 6.00 | 4Б(ж + ш)3Клж1Лт1К1Кз + Еа, П | |
| 10-1976 | 3.10 | 5Б(ж + ш)2Клж1Лт1Кз + Еа, П | |
| 11-1975 | 6.30 | 4Б(ж + ш)2Клж1Еа1Клз ед. П, К, Км, Я, Ив(к + тр.), Вм | |
| Мелкий подрост | 54-1987 | 19.60 | 5П2Б(ж + ш)1Еа1Клж1Клз + К, Лт, Я, Км |
| 5-1988 | 6.40 | 3П3Еа 2Б(ж + ш)1Кз1Клж ед. К, Лт | |
| 10-1976 | 6.60 | 3Еа2Кз2Б(ж + ш)2Клж1Лт | |
| 11-1975 | 11,50 | 4Еа2П1Б(ж + ш)1Лт1Клж1Клз ед.К, И(к + тр.), Ос, Вм, Рс | |
| Средний подрост | 54-1987 | 2.90 | 6Еа2П1Лт1Клз ед. К, Клж.Ос |
| 5-1988 | 3.40 | 6Еа2П1Лт1Кз ед. К, Бж, Клж | |
| 10-1976 | 2.60 | 3Еа3П2Лт2Клж | |
| 11-1975 | 6.90 | 7Еа1П1Лт1К ед. Клз, Клж, Вм | |
| Крупный подрост | 54-1987 | 0.70 | 4П2Еа2Лт1Клз1Вм + Ос |
| 5-1988 | 0.40 | 3Еа3Лт2П1Кз1Ос | |
| 10-1976 | 0.40 | 4Еа3П2Клж1Кз | |
| 11-1975 | 0.80 | 9Еа1П ед.К, КлжКлз, К | |
| Тонкомерные деревья | 54-1987 | 0.42 | 3Еа3П2Кж1Бж1Лт ед. К, Км, Кз |
| 5-1988 | 2.40 | 4Бш2Еа2П1Клж1Бж + К, Ос, Чм, Тк, Рс, Ик, Ит, Я, Ил, Кз | |
| 10-1976 | 4.87 | 3П3Клж1Бш1Лт1Еа1Бж ед. К, Ик, Клз, Бпл, Ос, Вм | |
| 11-1975 | 3.10 | 5П3Еа1Клж1Лт + К ед. Б(ж + ш), Клз | |
| Крупные деревья | 54-1987 | 0.50 | 5Еа2К2П1Б(ш + ж) |
| 5-1988 | 0.40 | 4Бш1Бж1Тк1П1Бпл1Ос1Чм + Еа, Кж, Ик, Ит, Чм | |
| 10-1976 | 0.21 | 5Бш2Бж1Бпл1Чм1Пед.К | |
| 11-1975 | 1.16 | 4Еа4П1Б(ш + ж)1К + Лт |
Анализ состава и структуры древостоя после подневольно-выборочной рубки (пр. пл. 54-1987) показал, что за 33-летний период насаждение здесь почти полностью восстановилось без смены пород и без существенных изменений в составе и структуре древостоя. После изъятия крупномерных стволов ели аянской и сосны корейской, имевших возраст 170–190 лет, средний возраст деревьев верхнего полога стал 110–130 лет. Древостой в количестве 8 видов, как и до рубки, состоял из трех подъярусов, в-которых преобладали темнохвойные породы (табл. 2).
Таблица 2.
Характеристика древостоев в сообществах кедрово-темнохвойного осоково-папоротникового леса, сформированных в течение 32–33 лет после разных способов рубок и низового пожара
| № пр. пл.-год закладки | Подъярус (высотные пределы, м) |
Породный состав (по запасу) |
Живых стволов, шт. га–1 |
Полнота относительная |
Площадь сечения, м2 га–1 |
Запас древесины, м3 га–1 |
Средние для преобладающей породы | |
|---|---|---|---|---|---|---|---|---|
| высота, м | диаметр, см | |||||||
| 54-1987 | I (20.1–27.0) | 7Еа2К1П + Лт, Бп | 128 | 1.0 | 17.7 | 194.1 | 23.0 | 40.4 |
| II (15.1–21.0) | 5Еа4П1Лт + Ос, Бп, Клз | 216 | 12.0 | 112.0 | 19.2 | 28.2 | ||
| III (5.0–15.0) | 5Еа3П1Лт1Клж + К | 340 | 2.8 | 16.9 | 10.0 | 11.2 | ||
| 5-1988 | I (14.1–24.0) | 3Бш3Тк1П1Ос1Бп1Бж ед. Еа, К | 348 | 0.76 | 9.9 | 81.4 | 17.3 | 16.9 |
| II (2.1–14.0) | 5Бш2Бж1Еа1П1Чм + Ивк, Итр ед. Кж, К, Ос | 2356 | 7.9 | 55.5 | 8.1 | 5.7 | ||
| 10-1976 | I (14.1–23.0) | 6Бш3Бж1Бп + Ос | 620 | 1.1 | 8.09 | 61.8 | 16.0 | 13.2 |
| II (2.1–13.0) | 3П3Клж1Лт1К1Бп1Чм + Еа, Ивк, ед. Клз, Бж | 4178 | 13.1 | 53.88 | 6.0 | 6.1 | ||
| 11-1975 | I (14.1–24.0) | 3Б(ш + ж)2Бп2Еа2П1Лт + Ивк, Итр, К | 426 | 0.83 | 14.6 | 100.0 | 19.7 | 19.5 |
| II (2.1–14.0) | 5Б(ш + ж)3П1Еа1(Ивк + Итр) + Клж ед.Бп, Лт, К | 2812 | 8.3 | 46.5 | 9.0 | 7.0 | ||
На остальных участках произошла смена господствующих пород, и среди крупных деревьев по численности и запасу древесины доминировали березы шерстистая, желтая и плосколистная. В древостое, сформировавшемся к 32 г. на экспериментальной вырубке (пр. пл. 10-1976), было отмечено 12 видов деревьев и выделялись два подъяруса. В первом, 14.1–23.0 м высотой, 16–22 см диаметром и сомкнутостью крон 0.4–0.5, доминировали березы всех трех видов (табл. 2). При этом береза плосколистная превышала по высоте (21.0–23.0 м) березы шерстистую и желтую, средняя высота которых составляла 16.0–18.0 м. В составе второго подъяруса, 2.0–14.0 м высотой, 3.0–16.0 см диаметром и сомкнутостью крон 0.8–0.9, по численности преобладали тонкомерные деревья пихты белокорой (1.23 тыс. экз. га–1). Ее растения опережали в росте ель аянскую благодаря более высоким значениям прироста по высоте (см. рис. 1) и значительному преобладанию ее в подросте предварительной генерации. В результате этого к 32 г. после рубки большая часть растений пихты белокорой перешла в категорию тонкомера, тогда как у ели основная часть растений находилась в категории крупного подроста.
Отдельные деревья пихты, сформировавшиеся из подроста предварительной генерации и достигшие 12–13 м высоты, перешли в возрастное состояние молодых генеративных растений и приступили к семеношению. О важности и необходимости сохранения предварительного подроста для успешного лесовозобновительного процесса отмечали многие исследователи (Ворошилов, 1970; Солодухин, 1965; и др.). Согласно К.П. Соловьеву и Е.Д. Солодухину, сохранение предварительного возобновления ели и пихты на рубках главного пользования сокращает сроки восстановления исходных древостоев на 30–40 лет.
К 32 году после экспериментальной рубки образовались полностью сомкнутые биогруппы хвойного молодняка с преобладанием растений пихты. В результате сильного затенения под их пологом исчезли почти все растения у представителей нижних ярусов, резко сократилось количество всходов, подроста и в целом произошло резкое снижение численности популяций у всех древесных видов (см. рис. 2). К этому времени остались жизнеспособными только 11.3% растений березы шерстистой, 8.8 – черемухи Маака, 5.1 – березы желтой и 4.9% растений ивы козьей от численности их на второй год после рубки. Снизилась численность и у хвойных пород: пихты – в 1.6 раз, ели и сосны корейской – в 1.7 раз по сравнению с достигавшими их максимумами на 15 год после рубки.
Видовое разнообразие в данном 32-летнем сообществе после экспериментальной рубки насчитывало 37 видов. В составе нижних ярусов к этому времени полностью исчезли растения инициальных видов, характерные для начальных стадий сукцессий. В кустарниковом ярусе, состоящем из 8 видов и имевшем общее проективное покрытие 10–15%, преобладали низкорослые (50–80 см длиной) лианы актинидии коломикта. Редкий осоково-папоротниковый травяной покров, состоящий из 18 видов и имевший общее проективное покрытие около 20%, отличался сильной мозаичностью.
Иной характер в составе и сложении сообществ отмечался в 33-летнем насаждении, сформированном после условно-сплошной рубки (пр. пл. 5-1988).
Как уже отмечалось ранее, сильное разрушение подстилки и трансформация почвенного покрова тяжелой техникой во время условно-сплошной рубки обусловили прорастание многочисленных хранившихся в почве всхожих семян берез и внедрение травянистых и кустарниковых растений инициальных видов. На 33 год после условно-сплошной рубки общий флористический состав производного сообщества включал 75 видов высших растений в отличие от 37 видов, отмеченных на 32 г. после экспериментальной рубки (пр. пл. 10-1976). Значительную примесь к осоково-папоротниковому покрову на пр. пл. 5-1988 составили инициальные виды (кипрей узколистный Chamaenerion angustifolium (L.) Scop., недотрога обыкновенная Impatiens noli-tangere L. и др.) и мезогигрофитное крупнотравье (чемерица Veratrum sp., какалия копьевидная Cacalia hastata L., лабазник камчатский Filipendula camtschatica (Pall.) Maxim. и др.). В кустарниковом ярусе также активно развивались инициальные виды (бузина красная Sambucus racemosa L., аралия высокая Aralia elata (Miq.) Seem., шиповник иглистый Rosa acicularis Lindl.).
В древостое на этой 33-летней вырубке, состоящем из двух подъярусов и образованном 11 видами деревьев, доминировали разные виды берез. В верхнем подъярусе, сформированном березами с ускоренным ростом, в качестве примеси присутствовали быстрорастущие деревья тополя корейского и осины, а также сохранившиеся во время рубки деревья хвойных пород (табл. 2). Во втором подъярусе доминировали деревья берез с замедленным ростом с примесью пихты белокорой и ели аянской, которые сформировались из тонкомера и крупного подроста, сохранившегося по периферии вырубки. Древостой на этой вырубке был более разреженный, с невысокой относительной полнотой (0.8), в отличие от древостоя на пр. пл. 10-1976, имевшем относительную полноту 1.0.
В то же время невысокая полнота древостоя, а также сохранившиеся семенники хвойных деревьев и примыкающие с двух сторон коренные кедрово-темнохвойные леса способствовали успешному формированию и развитию подроста темнохвойных пород всех высотных категорий к 33-му году после условно-сплошной рубки.
На гари кедрово-темнохвойного леса (пр. пл. 11-1975), как было отмечено ранее, в течение первых 15 лет после пожара происходило постоянное нарастание численности хвойных пород. По мере роста молодых растений ели и пихты происходило смыкание их крон, и к 15 г. после пожара сформировались сомкнутые биогруппы высотой 1.5–3.0 м, под пологом которых началось интенсивное изреживание подроста древесных растений, в основном за счет отставших в росте экземпляров. Наиболее интенсивное самоизреживание отмечалось у многочисленных растений ели аянской, численность популяции которой снизилась с 53.4 тыс. экз. га–1, отмеченных в 15-летнем послепожарном сообществе, до 9.9 тыс. экз. га–1 экземпляров на 33 год после пожара. У всех других хвойных и лиственных климаксовых видов за этот период также произошло значительное снижение численности их растений (см. рис. 2).
На 33 г. после пожара в сформировавшемся производном сообществе флористический состав почти полностью восстановился и включал 63 вида высших сосудистых растений. Среди инициальных видов присутствовали лишь единичные низкорослые растения (высотой менее 1 м) бузины кистистой и аралии высокой. Кустарниковый и кустарничково-травяной ярусы отличались от коренных сообществ более высокой мозаичностью и меньшим проективным покрытием из-за образования плотно сомкнутых биогрупп темнохвойных пород.
Древостой был образован 10 видами деревьев, а по запасам древесины в двух сформировавшихся подъярусах доминировали быстрорастущие деревья берез (табл. 2). При этом в первом подъярусе по высоте деревьев четко выделялись два полога: первый полог 19–23 м высотой и 20–25 см диаметром образовали растения березы плосколистной и осины, а также сохранившиеся во время пожара деревья ели, сосны корейской и липы Таке. Второй полог 14–18 м высотой и 12–20 см диаметром сформировали деревья берез желтой и шерстистой с примесью ив козьей и поронайской, черемухи Маака и клена желтого. Второй подъярус (2.1–14 м высотой и 4–12 см диаметром) составили отставшие в росте деревья берез желтой и шерстистой, а также растения с ускоренным ростом пихты и ели. При этом пихта опережала в росте растения ели аянской, и к 33 г. после пожара значительная часть ее растений первой послепожарной генерации перешла в категорию тонкомера, тогда как у ели большая часть растений такого же возраста находилась в категории крупного подроста.
ЗАКЛЮЧЕНИЕ
Анализ лесовосстановительного процесса в течение 32–33 лет после разных способов рубки и устойчивого низового пожара в сообществах кедрово-темнохвойного осоково-папоротникового леса Южного Сихотэ-Алиня показал, что естественный ход возобновления и развития молодого поколения хвойных лесообразователей в целом осуществлялся удовлетворительно во всех сообществах. При этом в разных категориях подроста и в тонкомере преобладали ель аянская и пихта белокорая. Успешность лесовозобновительного процесса на исследуемых участках была связана с незначительной площадью вырубок и гари (каждая менее 1 га), сохранностью в той или иной степени подроста предварительной генерации, наличием на площадях семенников хвойных пород и близостью коренных лесов.
Наиболее быстрый и успешный лесовосстановительный процесс без смены коренных пород осуществлялся после зимней подневольно-выборочной рубки. После удаления крупномерных стволов ели и сосны корейской, имевших возраст 170–190 лет, средний возраст верхнего полога древостоя снизился до 110–130 лет, а состав и структура насаждения почти полностью восстановились к 33 г. после рубки.
После экспериментальной сплошной рубки, а также после условно-сплошной рубки и устойчивого низового пожара в сообществах кедрово-темнохвойного осоково-папоротникового леса восстановление сообществ идет через смену климаксовых хвойных пород на быстрорастущие серийные виды берез, осины, ив козьей и поронайской, черемухи Маака и др.
Наиболее длительный процесс восстановления кедрово-темнохвойных лесов происходит после сплошных рубок с использованием тяжелой агрегатной техники, нарушающих напочвенный покров, подстилку и почти весь подрост хвойных пород.
Сохранение предварительного подроста и тонкомера во время экспериментальной рубки позволяет сокращать сроки восстановления коренных лесов примерно на 30–40 лет, но и в этом случае должно пройти не менее 100 лет.
После устойчивого низового пожара к 33 г. в нижних ярусах кедрово-темнохвойных осоково-папоротниковых лесов исчезают почти все инициальные виды, а в древостое доминируют быстрорастущие серийные виды. Проведенный нами ранее анализ послепожарных лесовосстановительных сукцессий исследуемых лесов показал, что после устойчивых низовых пожаров господствующая роль ели аянской и пихты белокорой наступает к 100–120 гг., а максимальные запасы древесины у сосны корейской отмечаются после 160–180 лет.
Список литературы
Ворошилов В.П., Манько Ю.И. Выживаемость хвойного подроста на сплошных вырубках в пихтово-еловых лесах // Использование и воспроизводство лесных ресурсов Дальнего Востока. Хабаровск: ДальНИИЛХ, 1972. С. 91–93.
Ворошилов В.П. Выживаемость и рост хвойного подроста после рубок в ельниках Среднего Сихотэ-Алиня // Стационарные исследования в пихтово-еловых лесах Сихотэ-Алиня. Владивосток: Изд-во ДВНЦ АН СССР, 1975. С. 123–130.
Гулисашвили В.З. Внутривидовые взаимоотношения и регулирование численности особей в растительных сообществах // Современные вопросы лесоведения и лесной биогеоценологии. М.: Изд-во Наука, 1974. С. 233–245.
Данилик В.Н. Экологические особенности возобновления ели // Физиология и экология древесных растений. Свердловск, 1965. Вып. 43. С. 209–213.
Ковалев А.П., Алексеенко А.Ю. Лесные ресурсы Дальнего Востока и перспективы неистощительного пользования // Леса России: политика, промышленность, наука, образование: мат-лы третьей Междунар. науч.-техн.конф. СПб., 2018. С. 150–152.
Ковалев А.П., Орлов А.М., Лашина Е.В., Грищенова Ю.А. Состояние и перспективы использования лесных ресурсов Приморского края // Сибирский лесной журн. 2019. № 5. С. 15–21.
Комарова Т.А. Семенное возобновление растений на свежих гарях (леса Южного Сихотэ-Алиня). Владивосток: Изд-во ДВНЦ АН СССР, 1986. 224 с.
Комарова Т.А., Ильина Т.М. Изменение всхожести семян ели аянской в зависимости от характера послепожарного субстрата // Динамика и состояние лесных ресурсов Дальнего Востока. Хабаровск: Изд-во ФГУ ДальНИИЛХ, 2002. С. 115–121.
Комарова Т.А., Прохоренко Н.Б., Глушко С.Г., Терехина Н.В. Послепожарные сукцессии в лесах Сихотэ-Алиня с участием Pinus koraiensis Siebold et Zucc. СПб.: ООО “Свое издательство”, 2017. 402 с.
Корчагин А.А. Строение растительных сообществ // Полевая геоботаника. Л.: Изд-во Наука, 1976. 313 с.
Лукина Н.В., Исаев А.С., Крышень А.М., Онучин А.А., Сирин А.А., Гагарин Ю.Н., Барталев С.А. Приоритетные направления развития лесной науки как основы устойчивого управления лесами // Лесоведение. 2015. № 4. С. 243–254.
Манько Ю.И., Ворошилов В.П. Естественное возобновление на вырубках и гарях в кедрово-широколиственных лесах Приморского края // Итоги изучения лесов Дальнего Востока. Владивосток: Изд-во ДВФ СО АН СССР, 1967. С. 197–200.
Мелехов И.С. Лесоведение. М.: Изд-во Лесн. пром-сть, 1980. 408 с.
Правила рубок главного пользования в лесах Дальнего Востока. М.: Изд-во ЦБНТИлесхоз, 1970. 44с.
Сапожников А.П., Карпачевский Л.О., Ильина Т.М. Послепожарное почвообразование в кедрово-широколиственных лесах // Лесной вестник. 2001. № 1(16). С. 132–164.
Современное состояние лесов российского Дальнего Востока и перспективы их исользования / под редакцией А.П. Ковалева. Хабаровск: ДальНИИЛХ, 2009. 470 с.
Соловьев К.П. О динамике древостоев после рубок в Приморье // Сб. тр. ДальНИИЛХ. 1963. Вып. 5. С. 29–39.
Соловьев К.П., Солодухин Е.Д. Возобновление древесных пород в лесах Дальнего Востока // Возобновление леса. М.: Изд-во Колос, 1975. С. 283–303.
Солодухин Е.Д. Лесоводственные основы хозяйства в кедровых лесах Дальнего Востока. Владивосток: Дальневост. кн. изд-во, 1965. 367 с.
Сосудистые растения советского Дальнего Востока. СПб.: Изд-во Наука, 1985–1996. Т. 1–8.
Справочник лесоустроителя Дальнего Востока. Хабаровск: Дальневосточное лесоустроительное предприятие ВО “Леспроект”, 1973. 226 с.
Сукачев В.Н., Зонн С.В. Методические указания к изучению типов леса. М.: Изд-во АН СССР, 1961. 144 с.
Сысоев Е. Лесовосстановление концентрированных вырубок путем аэросева в условиях Кировской области. Киров: Кн. изд-во, 1959. 78 с.
Трофимова А.Д. Дендроиндикация условий произрастания основных хвойных пород Южного и Среднего Сихотэ-Алиня: дис. … канд. геогр. наук: 25.00.25. СПб., 2012. 190 с.
Чумин В.Т., Ковалев А.П., Обручников С.Г. Влияние сплошных рубок с применением агрегатных машин на сохранность тонкомера и подроста // Лесное хозяйство. 1981. № 1. С. 12-14.
Шешуков М.А. О классификации лесных пожаров по величине выгоревшей площади // Лесное хозяйство. 1967. Вып. 1. С. 53–57.
Algren T.F., Algren C.E. Ecological effects of forest fires // Bot. Rev. 1960. V. 26. № 4. P. 483–533.
Clements F.E. Plant Succession and Indicators. N.Y.: Wilson; 1928. 452 p.
Kuuluvainen T., Lindberg H., Vanha-Majamaa I., Keto-Tokoi P., Punttila P. Low-level retention forestry, certification, and biodiversity: case Finland // Ecological Processes. 2019. V. 8. № 1. 13 p.
Viro P.J. Effects of forest fire on soil // Fire and Ecosystems. N.Y.: Acad. press, 1974. P. 3–46.
Дополнительные материалы отсутствуют.