

Лесоведение, 2022, № 3, стр. 250-261
Пространственная неоднородность химического состава лесных подстилок сосновых насаждений средней тайги Восточной Фенноскандии
Г. В. Ахметова *
Институт леса КарНЦ РАН
185910 Республика Карелия, Петрозаводск, ул. Пушкинская, д. 11, Россия
* E-mail: akhmetovagv@gmail.com
Поступила в редакцию 15.10.2021
После доработки 16.01.2022
Принята к публикации 02.02.2022
- EDN: CLSODN
- DOI: 10.31857/S0024114822030020
Аннотация
Лесная подстилка является важным компонентом таежных экосистем, и изучение трансформации ее характеристик, в том числе химического состава, представляет собой большой научный интерес в связи с ее высокой пространственной изменчивостью этого почвенного горизонта даже в пределах одного типа леса. Цель работы – выявление особенностей содержания макро- и микроэлементов в лесных подстилках и закономерностей изменения их химического состава на уровнях элементарных геохимических ландшафтов (ЭГХЛ) и растительных микрогруппировок (РМГ). Данное исследование проведено в среднетаежной подзоне Восточной Фенноскандии (республика Карелия) в Государственном заповеднике “Кивач”. В сосновом местообитании, сформированном на песчаных флювиогляциальных отложениях, заложена ландшафтно-геохимическая катена на пологом склоне озовой гряды. Проанализировано валовое содержание и содержание в подвижной форме макро- (K, Mg, Ca) и микроэлементов (Mn, Zn, Cu) в подгоризонтах лесной подстилки. Результат исследования показал, что содержание K, Mn, Zn и Cu подвержено сильному варьированию на всех изучаемых уровнях структурно-функциональной организации территории. Концентрация Mg и Ca варьирует менее значительно. Дисперсионный анализ полученных данных показал статистически значимое влияние (р ≤ 0.05) выделенных факторов (ЭГХЛ и РМГ) на изменение содержания большинства макро- и микроэлементов. Динамика большинства изучаемых элементов с высокой степенью достоверности (R2) описывается полиноминальной кривой второго порядка. Только для калия была выявлена обратная линейная зависимость от расположения в системе ЭГХЛ. Лесные подстилки водораздельных участков (элювиальный ЭГХЛ) и верхних частей склонов (трансэлювиальный ЭГХЛ) характеризуются накоплением калия, магния, цинка и меди по сравнению с подчиненными ЭГХЛ (трансаккумулятивный и аккумулятивный ЭГХЛ). Для кальция и марганца отмечается иная закономерность – максимальные их значения характерны для лесных подстилок почв, формирующихся на склонах в транзитных условиях, и высокое содержание данных элементов больше связано с высокими их концентрациями в доминирующем здесь виде растений – чернике.
Напочвенный растительный покров и лесная подстилка являются чрезвычайно важными элементами лесных биогеоценозов (Богатырев, 1990, 1997, 2013; Базилевич и др., 1986; Sayer, 2006; Hilli et al., 2010; Орлова и др., 2011, 2015; Титлянова, Шибарева, 2012). Подстилка является особым компонентом экосистемы, промежуточным звеном между растительностью, опадом и почвой (Богатырев, 1997), источником и хранилищем элементов минерального питания, средой обитания живых организмов, основой первичного почвообразования и растительным субстратом (Лукина и др., 2010; Hilli et al., 2010; Телеснина и др., 2018; Lukina et al., 2019; Kuznetsova et al., 2021). Очень важно оценить направленность воздействия растительного покрова на почву, которое наиболее интенсивно проявляется в поверхностных органогенных горизонтах (Orlova et al., 2013; Семенюк и др., 2020). Как известно, неоднородность растительного покрова во многом определяет мозаичность и мелкоконтурность почвенного покрова, что наиболее четко отражается на свойствах их верхних горизонтов, формирующихся из растительного опада (Богатырев, Фомина, 1990; Vesterdal, Raulund-Rasmussen, 1998; Tamminen, 2004; Nilsson, Wardle, 2005; Артемкина и др., 2018; Noreika et al., 2019). Роль живого напочвенного покрова и подстилки особенно значимы в таежных бореальных лесах, где фитомасса растительности подчиненных ярусов и ее ежегодный отпад сопоставимы с этими показатели древесного яруса (Hilli et al., 2010).
Подстилка служит интегральным показателем биологического круговорота, отражает основные направления функционирования экосистем, в своем строении и свойствах подчиняется биоклиматической зональности (Богатырев, 2013). Поэтому информация о различных характеристиках лесной подстилки также могут служить индикатором системы состояния лесных экосистем, что особенно значимо в условиях нарастания антропогенного влияния (Богатырев, 1990; Телеснина и др., 2018). Органогенные горизонты почвы, по сравнению с почвой, являются более лабильными компонентами таежных экосистем, варьирование их свойств на всех уровнях структурно-функциональной организации экосистемы чрезвычайно высоки, это создает определенные сложности при проведении исследований лесных почв (Семенюк и др., 2020). В последнее время увеличилось количество работ, направленных на выявление взаимосвязей растительного покрова и характеристик лесных подстилок на различных уровнях ее организации, начиная с элементарных биогеоценозов, так как именно на этом уровне происходит взаимодействие напочвенного покрова, опада и лесных подстилок (Лукина, 2010; Орлова, 2013; Орлова и др., 2015; Noreika et al., 2019; Семенюк и др., 2020). Большинство работ направлено на изучение морфологических признаков подстилок и особенностей накопления в ней органического вещества и азота, продуцирование СО2; исследований, касающихся содержания других химических элементов недостаточно.
Изучение особенностей взаимосвязей в системе – “растение–лесная подстилка” в настоящее время также актуально в связи с тенденциями в изменении климатических показателей (Второй …, 2014; Kuemmerle et al., 2015; Climate change …, 2021) и растительного покрова (Epstein et al., 2004; Голубятников, Денисенко, 2009; Zellweger et al., 2020; Липка и др., 2021). Как известно, при повышении температуры ускоряются процессы разложения подстилки, происходит снижение их запасов (Попова, 2018), круговорот элементов-биогенов меняется – быстро освобождаются биогенные элементы и более активно вымывается калий в условиях промывного режима почв. Показано, что в зависимости от изменений климатических показателей будут меняться абсолютные величины аккумуляции и выщелачивания элементов (Шибарева, 2004).
Цель данной работы заключалась в выявлении особенностей содержания и закономерностей изменения химического состава лесных подстилок в зависимости от градиента расположения в ряду сопряженных почв и характера напочвенного покрова.
ОБЪЕКТЫ И МЕТОДИКА
Исследования проводились в Республике Карелия, в Государственном заповеднике “Кивач”, который расположен в юго-восточной части Балтийского кристаллического щита в среднетаежной подзоне Восточной Фенноскандии. Кристаллическое основание и покрывающая его морена перекрыты мощными гравийно-галечными песчаными флювиогляциальными наносами, рельеф формируют высокие песчано-галечные гряды – озы, высота которых составляет 20 м. В понижениях рельефа формируются верховые и переходные болота и небольшие заболоченные лесные озера. Данный тип местности широко распространен на территории Карелии, он отличается средним и низким классом бонитета сосновых лесов (лишайниковых и зеленомошных), а также бедностью напочвенного растительного покрова.
Пробные площади заложены в ряду сопряженных элементарных геохимических ландшафтов (катене) – от элювиальных – на водоразделе, через транзитные (трансэлювиальные и трансэлювиально-аккумулятивные) – на склоне к подчиненным (аккумулятивные) элементам рельефа (рис. 1). На автоморфных позициях в элювиальных и трансэлювиальных ЭГХЛ под сосняками брусничными формируются подзолы иллювиально-железистые песчаные. Данные почвы широко распространены на территории Карелии и считаются фоновыми, имеют легкий гранулометрический состав, хорошо дренированы и имеют высокую степень аэрации. В нижней части склона оза неширокой полосой в периферийной части заболоченного участка леса под чернично-сфагновыми микрогруппировками подзолы иллювиально-железистые сменяются полугидроморфными почвами – торфяно-подзолами иллювиально-железистыми глеевыми. В подчиненных элементах рельефа в условиях заболоченного сосняка багульнико-сфагнового формируются торфяные и торфяно-глеевые почвы переходного типа. Более подробные описания почвенных условий даны в ранее вышедших работах (Ахметова, 2019, 2021).
Рис. 1.
Расположение пробных площадей в сопряженном ряду элементарных геохимических ландшафтов (1 – лишайниковая РМГ, 2 – зеленомошная РМГ, 3 – брусничная РМГ, 4 – черничная РМГ, 5 – черничная влажная РМГ, 6 – чернично-сфагновая РМГ, 7 – багульниково-сфагновая РМГ, 8 – сфагновая РМГ).
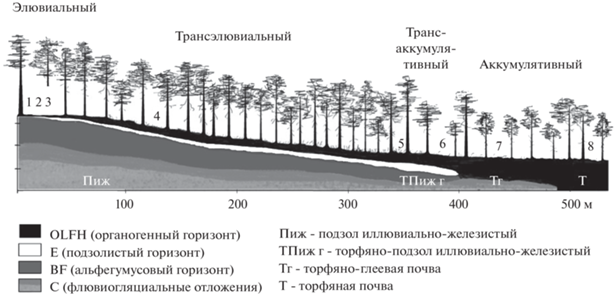
Древостой представлен чистым сосновым насаждением (10С), возраст – 190 лет (до 220 лет), класс бонитета – III,4. Напочвенный покров отличается бедностью видового состава напочвенного покрова, что является характерным для песчаных флювиогляциальных ландшафтов (Крышень и др., 2006). Лишайниковые и брусничные растительные микрогруппировки отличаются обилием и разнообразием лишайников, преобладают ягельные виды – кладония оленья (Сladonia rangiferina), цетрария исландская (Cetraria islandica). Среди зеленых мхов доминирует плевроциум Шребера (Pleurozium schreberi), реже встречаются гилокомиум блестящий (Hylocomium splendens) и птилиум гребенчатый (Ptilium crista-castrensis). В кустарничковом ярусе брусничных и черничных РМГ преобладают черника (Vaccinium myrtillus L.) и брусника (V. vitis-idaea L.). На заболоченных участках широко распространены торфообразующие мхи – сфагнум магелланский (Sphagnum magellanicum) и политрихум обыкновенный (Polytrichum commune). Среди кустарничков, помимо черники и брусники, встречается багульник (Lédum palústre), реже голубика (Vaccínium uliginósum).
В элювиальном ЭГХЛ исследования проводились на трех выделенных растительных микрогруппировках (РМГ) – лишайниковой, зеленомошной, брусничной. В элювиально-транзитном изучались наиболее распространенные черничные микрогруппировки. В трансаккумулятивном ЭГХЛ выделены две растительные микрогруппировки – черничная влажная и чернично-сфагновая, в аккумулятивном – багульниково-сфагновая и сфагновая.
Отбор образцов лесной подстилки проводился в подкроновых позициях в 4–6-кратной повторности. Разделение подстилки на слои проводилось в зависимости от ее морфологии (Богатырев, Фомина, 1990; Tamminen et al., 2004; Орлова и др., 2015), фиксировались мощность каждого слоя и ее вес. Для оценки запасов подстилки использовали почвенный бур диаметром 10 см. В лабораторных условиях было проведено определение абсолютной влажности образцов и рассчитан поправочный коэффициент. Содержание химических элементов (кальция, калия, магния, марганца, цинка, меди) определяли методом атомно-абсорбционной спектрофотометрии (AAnalyst 800 spectrometer). Определение содержания подвижных форм элементов проводили методом атомно-адсорбционной спектрометрии с извлечением ацетатно-аммонийным буферным раствором с рН 4.8 (ААБ).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
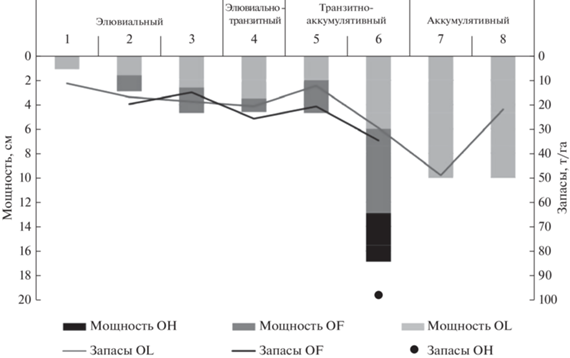
Лесная подстилка – один из наиболее динамичных элементов таежных экосистем, ее свойства подвержены существенным трансформациям в связи с изменениями условий произрастания фитоценозов (Богатырев, 1986; Орлова и др., 2011, 2015; Сергеева и др., 2020). Основные морфологические характеристики лесных подстилок: строение, мощность, запасы – существенно изменяются в соответствии с местоположением в системе ЭГХЛ и РМГ (рис. 2). Под лишайниковой растительностью формируется маломощная подстилка (0.5–1.5 см), и разделить ее на подгоризонты по степени гумификации не представлялось возможным. Для зеленомошно-кустарничковых РМГ характерна более мощная подстилка, она делилась на две части – опадочный OL и ферментативный OF. Гумусированный слой ОН не выделялся из-за его дискретности и значительного количества включений минеральных частиц. В полугидроморфных позициях транзитного ландшафта формируется мощная оторфованная подстилка, где отчетливо выделяются OL, OF и OH слои. Торфяные почвы аккумулятивного ландшафта представляют собой мощный слой (40–60 см) слабо разложенного сфагнового мха, в процессе данного исследования расчеты велись для верхней 10-сантиметровой толщи торфа. Запасы лесной подстилки сильно варьируют в зависимости от растительного покрова и местоположения в рельефе. Наименьшие значения рассчитаны для подстилок, сформированных под лишайниковой растительностью – 6–16 т/га, в кустарничково-зеленомошных РМГ запасы подстилки в 2–3 раза выше. Максимальные значения характерны для полугидроморфных местообитаний – трансаккумулятивной ЭГХЛ (до 150 т/га).
Рис. 2.
Изменение мощности и запасов лесных подстилок, формирующихся в сопряженных элементарных ландшафтах под различными РМГ: 1 – лишайниковая РМГ, 2 – зеленомошная РМГ, 3 – брусничная РМГ, 4 – черничная РМГ, 5 – черничная влажная РМГ, 6 – чернично-сфагновая РМГ, 7 – багульниково-сфагновая РМГ, 8 – сфагновая РМГ.
Химический состав органогенных горизонтов исследуемых почв таежных экосистем характеризуется широким диапазоном данных. Рассчитаны статистические показатели содержания макро- и микроэлементов в лесных подстилках на разных уровнях организации исследуемой территории – от катены в целом и элементарных ландшафтов до изучения изменчивости внутри каждой выделенной РМГ (табл. 1).
Таблица 1.
Статистические показатели содержания макро- и микроэлементов в лесных подстилках почв на уровнях ЭГХЛ и РМГ
| Уровень | Форма | Показатель | K | Mg | Ca | Mn | Zn | Cu |
|---|---|---|---|---|---|---|---|---|
| Катена | Вал. | ср ± ош.ср | 680 ± 70 | 440 ± 22 | 3000 ± 250 | 234 ± 57 | 29.12 ± 3.1 | 4.7 ± 0.6 |
| CV, % | 70 | 21 | 38 | 99 | 47 | 61 | ||
| Подв. | ср ± ош.ср | 240 ± 44 | 131 ± 7 | 640 ± 34 | 44.1 ± 8.7 | 3.77 ± 0.44 | 0.42 ± 0.02 | |
| CV, % | 91 | 25 | 27 | 98 | 59 | 29 | ||
| Элювиальный ЭГХЛ |
Вал. | ср ± ош.ср | 860 ± 87 | 470 ± 33 | 3384 ± 272 | 291 ± 54 | 34.69 ± 5.3 | 5.61 ± 1.21 |
| CV, % | 31 | 21 | 24 | 56 | 47 | 66 | ||
| Подв. | ср ± ош.ср | 307 ± 73 | 146 ± 8 | 674 ± 47 | 105.7 ± 17.3 | 5.26 ± 0.55 | 0.42 ± 0.03 | |
| CV, % | 83 | 18 | 24 | 57 | 36 | 26 | ||
| Лишайни-ковая РМГ | Вал. | ср ± ош.ср | 860 ± 121 | 430 ± 19 | 3200 ± 400 | 180 ± 8 | 42.7 ± 12 | 5.1 ± 1.8 |
| CV, % | 28 | 9 | 24 | 9 | 56 | 70 | ||
| Подв. | ср ± ош.ср | 469 ± 210 | 138 ± 14 | 475 ± 44 | 11.3 ± 0.7 | 2.99 ± 0.28 | 0.44 ± 0.04 | |
| CV, % | 77 | 17 | 16 | 10 | 16 | 15 | ||
| Зеленомошная РМГ | Вал. | ср ± ош.ср | 962 ± 80 | 461 ± 22 | 2870 ± 104 | 219 ± 27 | 33.8 ± 5.4 | 14.28 ± 6.2 |
| CV, % | 14 | 8 | 6,3 | 60 | 28 | 76 | ||
| Подв. | ср ± ош.ср | 428 ± 215 | 158 ± 16 | 864 ± 73 | 84.1 ± 7.7 | 6.43 ± 1.17 | 0.32 ± 0.04 | |
| CV, % | 87 | 18 | 15 | 16 | 31 | 21 | ||
| Брусничная РМГ | Вал. | ср ± ош.ср | 541 ± 142 | 443 ± 70 | 4000 ± 790 | 328 ± 75 | 33.8 ± 5.4 | 14.28 ± 6.3 |
| CV, % | 46 | 27 | 34 | 60 | 28 | 76 | ||
| Подв. | ср ± ош.ср | 354 ± 39 | 149 ± 5 | 638 ± 11 | 148.4 ± 11.8 | 5.35 ± 0.93 | 0.48 ± 0.1 | |
| CV, % | 19 | 6 | 3 | 14 | 30 | 35 | ||
| Транс-элювиальный ЭГХЛ Черничная РМГ |
Вал. | ср ± ош.ср | 470 ± 132 | 417 ± 22 | 4000 ± 350 | 372 ± 138 | 26.4 ± 6.26 | 5.32 ± 1.7 |
| CV, % | 48 | 9 | 15 | 78 | 41 | 54 | ||
| Подв. | ср ± ош.ср | 166 ± 14 | 144 ± 26 | 680 ± 58 | 147.9 ± 8.2 | 5.40 ± 1.05 | 0.48 ± 0.05 | |
| CV, % | 15 | 32 | 15 | 10 | 34 | 17 | ||
| Трансаккуму-лятивный ЭГХЛ | Вал. | ср ± ош.ср | 410 ± 170 | 381 ± 81 | 2240 ± 508 | 135 ± 72 | 24.2 ± 4.87 | 3.50 ± 0.35 |
| CV, % | 64 | 37 | 40 | 93 | 35 | 17 | ||
| Подв. | ср ± ош.ср | 177 ± 29 | 126 ± 10 | 724 ± 62 | 60.6 ± 35 | 3.28 ± 1.3 | 0.37 ± 0.1 | |
| CV, % | 26 | 28 | 28 | 93 | 63 | 36 | ||
| Черничная влажная РМГ | Вал. | ср ± ош.ср | 263 ± 18 | 350 ± 19 | 1550 ± 270 | 2.64 ± 4.3 | 17.1 ± 0.2 | 3.4 ± 0.46 |
| CV, % | 12 | 9 | 30 | 28 | 1.3 | 24 | ||
| Подв. | ср ± ош.ср | 189 ± 38 | 151 ± 18 | 904 ± 16 | 109.5 ± 15.0 | 4.84 ± 1.06 | 0.43 ± 0.08 | |
| CV, % | 35 | 21 | 3 | 24 | 38 | 30 | ||
| Чернично-сфагновая РМГ | Вал. | ср ± ош.ср | 516 ± 21 | 400 ± 99 | 2930 ± 230 | 178 ± 50 | 31.3 ± 2.4 | 3.65 ± 0.36 |
| CV, % | 24 | 57 | 14 | 28 | 11 | 17 | ||
| Подв. | ср ± ош.ср | 165 ± 9 | 101 ± 8 | 543 ± 35 | 11.6 ± 1.8 | 1.72 ± 0.17 | 0.32 ± 0.08 | |
| CV, % | 9 | 13 | 11 | 27 | 17 | 43 | ||
| Аккумулятив-ный ЭГХЛ | Вал. | ср ± ош.ср | 638 ± 116 | 460 ± 32 | 2680 ± 246 | 25.3 ± 5.,2 | 19.9 ± 4.5 | 3.2 ± 0.2 |
| CV, % | 31 | 12 | 16 | 36 | 11 | 11 | ||
| Подв. | ср ± ош.ср | 45 ± 6 | 103 ± 11 | 510 ± 34 | 2.7 ± 1.0 | 1.70 ± 0.45 | 0.43 ± 0.06 | |
| CV, % | 35 | 25 | 16 | 92 | 65 | 31 | ||
| Багульниково-сфагновая РМГ | Вал. | ср ± ош.ср | 73 ± 6 | 432 ± 3 | 2202 ± 190 | 25 ± 1 | 14.2 ± 1 | 3.3 ± 1 |
| CV, % | 29 | 20 | 18 | 10 | 16 | 12 | ||
| Подв. | ср ± ош.ср | 47 ± 12 | 87 ± 9 | 536 ± 61 | 1.9 ± 0.7 | 0.55 ± 0.22 | 0.44 ± 0.1 | |
| CV, % | 45 | 18 | 20 | 64 | 68 | 40 | ||
| Сфагновая РМГ | Вал. | ср ± ош.ср | 590 ± 150 | 471 ± 41 | 2900 ± 94 | 25.8 ± 2.6 | 22.7 ± 4.9 | 3.18 ± 0.27 |
| CV, % | 44 | 15 | 5,6 | 12 | 37 | 15 | ||
| Подв. | ср ± ош.ср | 35 ± 7 | 119 ± 15 | 483 ± 39 | 2.2 ± 0.9 | 1.82 ± 0.36 | 0.42 ± 0.07 | |
| CV, % | 35 | 22 | 14 | 69 | 34 | 28 |
Варьирование содержания макро- и микроэлементов в лесных подстилках изучаемых почв различается в зависимости от элемента и формы его нахождения. Наименьшая изменчивость выявлена для магния и кальция, для них получены наименьшие значения коэффициентов вариации (10–40%) как внутри каждой выделенной РМГ, так и катены в целом. Содержание остальных изученных элементов подвержено более значительному варьированию, очень высокие коэффициенты вариации (до 100%) рассчитаны для данных валового содержания калия и микроэлементов – марганца, меди и цинка. Как известно, в подвижной форме содержание элементов характеризуется более сильным варьированием, наши данные это подтверждают отчасти. Диапазон разброса значений концентрации элементов в подвижной форме в целом для катены более широкий по сравнению с валовым содержанием, наибольшим коэффициентом вариации характеризуется подвижный марганец. Тогда как при расчете данных для отдельных ЭГХЛ выявлено, что содержание элементов в подвижной форме характеризуется таким же, а иногда и меньшим варьированием. Это может свидетельствовать о более сильных связях с характером растительного покрова.
В связи с тем, что при гумуфикации подстилки происходит изменение ее химического состава (Tamminen et al., 2004), слои лесных подстилок могут характеризоваться различным уровнем обеспеченности макро- и микроэлементами. Дисперсионный анализ (табл. 2 ) показал статистически значимое влияние (р ≤ 0.05) на содержание большинства изучаемых макро- и микроэлементов степени разложения подстилки. Рассчитанные в ранее проведенных исследованиях (Ахметова, 2021) коэффициенты биологического поглощения этих элементов имеют высокие значения и показывают, что они относятся к группе элементов энергичного (Mn, K, Ca) и сильного (Mg, Zn, Cu) биогенного накопления, т.е. активно включаются в круговорот. Поэтому, в той или иной степени, с увеличением гумификации наблюдается снижение содержания элементов, наиболее быстро “теряется” из подстилки калий.
На химический состав лесных подстилок почв таежных экосистем влияет множество факторов (как природных, так и антропогенных), в данном исследовании изучалось влияние особенностей напочвенного растительного покрова (уровень РМГ), сопряженного с изменением в положении в рельефе и соответственного увеличения степени гидроморфизма (уровень ЭГХЛ). Дисперсионный анализ (табл. 2 ) показал статистически значимое влияние (р ≤ 0.05) данных факторов в связи с изменением содержания большинства макро- и микроэлементов. Отмечается, что концентрация элементов в подвижной форме более чутко реагирует на смену растительного сообщества, в связи с этим рекомендуется для оценки экологической ситуации использовать доступные растениям соединения (Нечаева 1985, Lukina et al., 2019).
Таблица 2.
Дисперсионный анализ (ANOVA) влияния факторов на химический состав лесных подстилок (даны значение уровня значимости р, при р ≤ 0.05 – влияние значимо, >0.05 – не значимо)
| Фактор | Форма | K | Ca | Mg | Mn | Zn | Cu |
|---|---|---|---|---|---|---|---|
| Степень разложения | Валовая | 0.001 | 0.05 | 0.01 | 0.006 | 0.04 | 0.002 |
| Подвижная | 0.002 | 0.04 | 0.01 | 0.033 | 0.002 | 0.02 | |
| ЭГХЛ | Валовая | 0.001 | 0.1 | 0.07 | 0.0007 | 0.17 | 0.37 |
| Подвижная | 7.8 × 10–7 | 0.04 | 0.00002 | 2 × 10–6 | 7 × 10–7 | 0.33 | |
| РМГ | Валовая | 0.0009 | 0.3 | 0.4 | 0.004 | 9.8 × 10–7 | 0.3 |
| Подвижная | 3.4 × 10–11 | 1 × 10–10 | 1.4 × 10–6 | 1 × 10–14 | 2.6 × 10–7 | 0.006 |
Регрессионный анализ дополнил информацию к полученным закономерностям и показал, что изменение содержания большинства изучаемых макро- и микроэлементов с высокой степенью достоверности (R2) описывается полиноминальной кривой второго порядка (рис. 3). Только для калия была выявлена иная закономерность. Известно, что калий активно вымывается атмосферными осадками из крон древостоев, опада, лесной подстилки и верхней минеральной части почвы, его миграционная активность ослабевает на глубине почвы 20–40 см (Нечаева, 1985; Титлянова, 2014; Кожевникова и др., 2017). Анализ полученных данных показал достоверное влияние на содержание данного элемента в лесных подстилках месторасположения почвы в ряду сопряженных ЭГХЛ – с нарастанием увлажнения (от элювиального к аккумулятивному ЭГХЛ) наблюдается существенное снижение содержания данного макроэлемента. Этот тренд особенно отчетливо прослеживается для подвижной формы калия, тогда как для валового калия эта закономерность больше характерна для подгоризонта OF. В вышележащем подгоризонте OL большее влияние на содержание калия оказывает характер напочвенной растительности. Отмечается, что в наиболее распространенном виде растений в сфагновых РМГ– сфагнума магелланского содержание калия высокое (Ахметова, 2021), поэтому концентрация валового калия в торфе довольно велика, однако в нижних слоях торфа его количество резко снижается на порядок. В связи с этим тренды зависимости содержания валового калия отличаются для разных подгоризонтов подстилки (рис. 2). Для подвижного калия данной взаимосвязи не выявлено, хорошо прослеживается тренд снижения содержания калия во всех подгоризонтах лесных подстилок в соответствии с увеличением увлажнения, который аппроксимируется обратной линейной зависимостью с очень высокой величиной достоверности аппроксимации (R2 = 0.7–0.9).
Рис. 3.
Зависимость содержания (мг/кг) биогенных макро- и микроэлементов в подгоризонтах лесной подстилки от расположения в градиенте ЭГХЛ + РМГ (элювиальный ЭГХЛ: 1 – лишайниковая РМГ, 2 – зеленомошная РМГ, 3 – брусничная РМГ; трансэлювиальный: 4 – черничная РМГ; трансаккумулятивный: 5 – черничная влажная РМГ, 6 – чернично-сфагновая РМГ; аккумулятивный: 7 – багульниково-сфагновая РМГ, 8 – сфагновая РМГ).
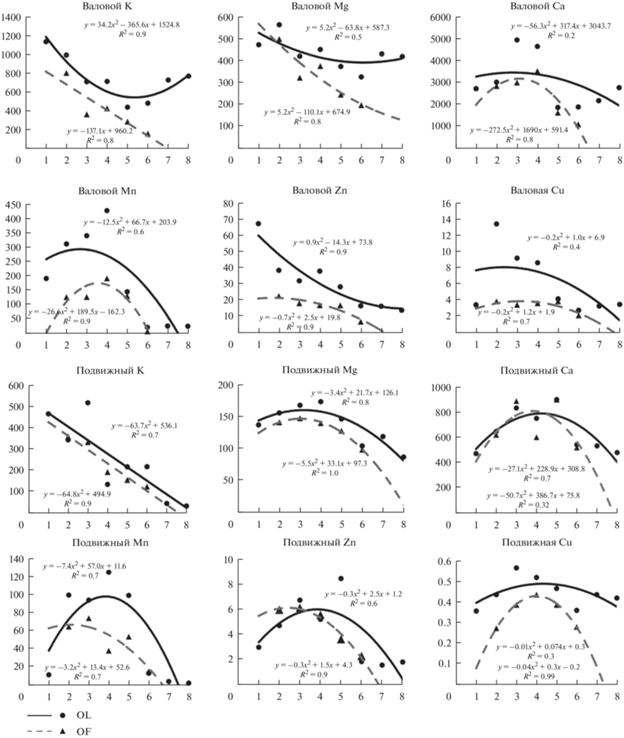
Изменчивость уровня концентрации магния и кальция в подстилках изучаемых почв, по сравнению с калием, намного ниже. Также для данных макроэлементов выявлены иные закономерности варьирования их содержания в лесных подстилках. Магний и кальций, несмотря на то, что являются довольно активными водными мигрантами в почве, более устойчивы при разложении органических остатков (Артемкина и др., 2018) по сравнению с калием и медленнее выносятся из опада, подстилки и почвы в целом. Не выявлено четких закономерностей изменения валового содержания кальция и магния при смене состава напочвенного покрова и нарастания гидроморфизма. Есть тенденция к увеличению содержания кальция в подстилках почв, сформированных под черничными и, в меньшей мере, под брусничными РМГ, что может быть связано с повышенной концентрацией кальция в кустарничках, особенно в чернике (Лукина и др., 2008). Однако наблюдается резкое снижение содержания кальция в подстилке черничной РМГ, сформированной в более влажных условиях – в трансэлювиальном ландшафте. Что касается подвижных форм рассматриваемых макроэлементов, выявлена достоверная связь изменения их концентрации, особенно магния, связанная с нарастанием степени гидроморфизма и изменения РМГ. Максимальные концентрации подвижных форм элементов отмечены в подстилках верхних склоновых участков, занятых брусничными и черничными РМГ. Выявленный тренд описывается полиномом второго порядка с высокой степенью достоверности – R2 = 0.7–0.9.
Марганец характеризуется наибольшей биогенностью среди изучаемых элементов, он является индикатором биогеохимических циклов в таежных ландшафтах (Нечаева, 1985) в связи с изменением валентного состояния в зависимости от окислительно-восстановительных условий. Выявлены достоверные различия степени накопления в подстилках, связанные с характером растительного покрова, приуроченность данного микроэлемента к кустарничкам рода Vaccinium – чернике и бруснике известна (Нечаева, 1985; Лукина и др., 2008), особенно богаты марганцем листья черники. В связи с этим лесные подстилки черничных РМГ отличаются наиболее высокой концентрацией данного микроэлемента; как валовый, так и подвижный марганец содержится в подстилках кустарничковых РМГ в 3–4 и более раз больше, чем в остальных рассматриваемых РМГ. Очень низкие значения данного микроэлемента отмечены в оторфованных подстилках торфяных почв багульниково-сфагновых и сфагновых РМГ. Изменение содержания марганца в зависимости от РМГ и степени увлажнения хорошо аппроксимируется полиномиальной функцией с R2 = 0.6–0.7.
Цинк и медь также относятся к группе с сильным биологическим накоплением, они активно участвуют в биогенном круговороте и накапливаются в подстилке, но закономерности изменения данных микроэлементов различаются.
Для цинка так же, как и для марганца, прослеживается тенденция к изменению его концентрации в подстилках в соответствии с изменением растительного сообщества, и в зависимости от степени гидроморфизма, выявленный тренд хорошо описывается полиноминальной функцией с R2 более 0.9. Максимальные значения данного микроэлемента выявлены в лишайниковой РМГ, наименьшие – в оторфованной подстилке сфагновых РМГ. Характер изменения содержания подвижного цинка иной. Отмечена приуроченность его максимального количества, так же, как и марганца, в подстилках элювиальных и трансэлювиальных позиций, сформированных под брусничными и черничными РМГ, в оторфованных подстилках (чернично-сфагновой, багульниково-сфагновой и сфагновой РМГ) его концентрация резко снижается.
Варьирование содержания меди в лесных подстилках, даже внутри каждой выделенной РМГ, довольно высокое, особенно высокие значения коэффициентов вариации отмечены для подвижной формы элемента в подстилках элювиального ЭГХЛ. Что касается роли растительного покрова и расположения в градиенте увлажнения в изменении валового содержании данного микроэлемента, достоверной закономерности выявлено не было. Для подвижной меди наблюдается связь его содержания с изменением характера растительности, выявлена наблюдаемая для кальция и марганца приуроченность наибольшей концентрации ее в подстилках черничных РМГ.
Таким образом, наблюдается отмечаемая Л.Г. Богатыревым для органического вещества подстилок таежных экосистем (Богатырев, 2013) приуроченность наибольшего содержания макро- и микроэлементов в органогенных горизонтах транзитных ландшафтах, что в значительной степени связано с характером растительного напочвенного покрова – преобладанием кустарничков рода Vaccinium, для которых характерна высокая биохимическая активность.
ЗАКЛЮЧЕНИЕ
Исследования, проведенные в сопряженном ряду почв сосновых древостоев флювиогляциальных равнин, позволили оценить особенности строения и химического состава лесных подстилок и определить закономерности изменения их свойств в зависимости от расположения в градиенте увлажнения и растительного напочвенного покрова. Основные морфологические характеристики лесных подстилок: строение, мощность, запасы – подвержены значительному варьированию, мощность подстилки и ее запасы имеют наименьшие значения в элювиальной ЭГХЛ в лишайниковой РМГ, максимальных значений эти параметры достигают в оторфованной подстилке трансаккумулятивной ЭГХЛ под чернично-сфагновой РМГ.
Пространственная неоднородность содержания макро- и микроэлементов в подстилках различается в зависимости от элемента и формы его нахождения. Наименьшая изменчивость выявлена для магния и кальция, содержание остальных изученных элементов подвержено более значительному варьированию.
Дисперсионный анализ полученных данных показал статистически значимое влияние (р ≤ 0.05) выделенных факторов (ЭГХЛ и РМГ) на содержание большинства макро- и микроэлементов в органогенных горизонтах почв. Динамика содержания большинства элементов с высокой степенью достоверности (R2) описывается полиноминальной кривой второго порядка. Только для калия была выявлена обратная линейная зависимость от расположения в системе ЭГХЛ. В целом лесные подстилки водораздельных участков (элювиальный ЭГХЛ) и верхних частей склонов (трансэлювиальный ЭГХЛ) характеризуются накоплением калия, магния, цинка и меди, по сравнению с подчиненными ЭГХЛ (трансаккумулятивным и аккумулятивным ЭГХЛ). Для кальция и марганца отмечается иная закономерность – максимальные значения характерны для лесных подстилок почв, формирующихся на склонах в транзитных условиях, и высокое содержание данных элементов связано с высокими их концентрациями в доминирующем здесь виде растений – чернике.
Данные были получены с использованием оборудования ЦКП “Аналитическая лаборатория” ИЛ КарНЦ РАН.
Список литературы
Артемкина Н.А., Орлова М.А., Лукина Н.В. Микромозаика растительности и вариабельность химического состава L-горизонтов подстилки северотаежных ельников кустарничково-зеленомошных // Лесоведение. 2018. № 2. С. 97–106. https://doi.org/10.7868/S002411481802002X
Ахметова Г.В. Геохимические особенности почв волнистой озерно-ледниковой песчаной равнины среднетаежной подзоны Карелии // Бюллетень Почвенного института им. В.В. Докучаева. 2019. № 100. С. 53–82. https://doi.org/10.19047/0136-1694-2019-100-53-82
Ахметова Г.В. Особенности распределения макро- и микроэлементов в системе “почва-растение” в среднетаежных условиях Восточной Фенноскандии // Труды КарНЦ РАН. 2021. № 5. С. 5–19. https://doi.org/10.17076/eco1228
Базилевич Н.И., Гребенщиков О.С., Тишков А.А. Географические закономерности структуры и функционирования экосистем. М.: Наука, 1986. 297 с.
Богатырев Л.Г. О классификации лесных подстилок // Почвоведение. 1990. № 3. С. 118–127.
Богатырев Л.Г. О роли гетерогенности подстилок в функционировании лесных биогеоценозов // Общие проблемы биогеоценологии. М.: АН СССР. 1986. С. 227–229.
Богатырев Л.Г. Образование подстилок – один из важнейших процессов в лесных экосистемах // Почвоведение. 1997. № 4. С. 501–512.
Богатырев Л.Г., Фомина Т.В. Диагностика нижних горизонтов лесных подстилок // Вестник Московского университета. Серия 17: Почвоведение. 1990. № 1. С. 54–56.
Богатырев Л.Г. Географические аспекты функционирования лесных подстилок // Вестник Московского университета. Серия 17. Почвоведение. 2013. № 1. С. 30–36.
Второй оценочный доклад Росгидромета об изменениях климата и их последствиях на территории Российской Федерации. М.: Росгидромет, 2014. 1006 с.
Голубятников Л.Л., Денисенко Е.А. Влияние климатических изменений на растительный покров Европейской России // Известия Российской академии наук. Серия географическая. 2009. № 2. С. 57–68.
Кожевникова Н.К., Луценко Т.Н., Болдескул А.Г., Лупаков С.Ю., Шамов В.В. Водная миграция макроэлементов в хвойно-широколиственных лесах Сихотэ-Алиня // Сибирский лесной журн. 2017. № 3. С. 60–73. https://doi.org/10.15372/SJFS20170306
Крышень А.М., Рудковская О.А., Преснухин Ю.В., Тимофеева В.В. Морфоструктура напочвенного покрова основных типов лесных сообществ заповедника “Кивач” (средняя тайга) // Труды Карельского научного центра РАН. 2006. Вып. 10. С. 54–62.
Липка О.Н., Корзухин М.Д., Замолодчиков Д.Г., Добролюбов Н.Ю., Крыленко С.В., Богданович А.Ю., Семенов С.М. Роль лесов в адаптации природных систем к изменениям климата // Лесоведение. 2021. № 5. С. 531–546. https://doi.org/10.31857/S0024114821050077
Лукина Н.В., Орлова М.А., Исаева Л.Г. Плодородие лесных почв как основа взаимосвязи почва-растительность // Лесоведение. 2010. № 5. С. 45–56.
Лукина Н.В., Полянская Л.М., Орлова М.A. Питательный режим почв северотаежных лесов. М.: Наука, 2008. 342 с.
Нечаева Е.Г. Ландшафтно-геохимический анализ динамики таежных геосистем. Иркутск: Институт географии СО АН РАН, 1985. 210 с.
Орлова М.А. Элементарная единица лесного биогеоценотического покрова для оценки эко системных функций лесов // Тр. Карельского НЦ. Сер. Экологические исследования. 2013. № 6. С. 126–132.
Орлова М.А., Лукина Н.В., Камаев И.О., Смирнов В.Э., Кравченко Т.В. Мозаичность лесных биогеоценозов и плодородие почв // Лесоведение. 2011. № 6. С. 39–48.
Орлова М.А., Лукина Н.В., Смирнов В.Э. Методические подходы к отбору образцов лесной подстилки с учетом мозаичности лесных биогеоценозов // Лесоведение. 2015. № 3. С. 214–221. https://doi.org/10.31857/S0033994620010045
Попова Н.В. Типизация напочвенных органогенных горизонтов в основных почвенно-фитоценотических экосистемах и индикация их состояния по параметрам деструктивной ветви малого биологического круговорота: автореф. дис. д. биол. наук: 03.02.08. Владимир, 2018. 47 с.
Семенюк О.В., Телеснина В.М., Богатырев Л.Г., Бенедиктова А.И., Кузнецов Я.Д. Оценка внутрибиогеоценозной изменчивости лесных подстилок и травяно-кустарничковой растительности в еловых насаждениях // Почвоведение. 2020. № 1. С. 31–43.
Сергеева О.В., Мухортова Л.В., Кривобоков Л.В. Распределение запасов подстилки и биомассы живого напочвенного покрова в северной тайге центральной Эвенкии в зависимости от рельефа // Сибирский лесной журн. 2020. № 1. С. 38–46. https://doi.org/10.15372/SJFS20200104
Телеснина В.М., Семенюк О.В., Богатырев Л.Г. и др. Особенности напочвенного покрова и лесных подстилок в искусственных липовых насаждениях в зависимости от характера ухода // Вестн. Моск. ун-та. Сер. 17. Почвоведение. 2018. № 2. С. 3–11.
Титлянова А.А., Шибарева С.В. Подстилки в лесных и травяных экосистемах. Новосибирск: СО РАН, 2012. 137 с.
Шибарева С.В. Запасы и элементный состав подстилок в лесных и травяных экосистемах Сибири: автореф. дис. … к. биол. наук: 03.00.27. Новосибирск, 2004. 24 с.
Choudhary D. K., Mishra Arti, Varma Ajit. Climate Change and the Microbiome: Sustenance of the Ecosphere. Springer Nature, 2021. 748 p. https://doi.org/10.1007/978-3-030-76863-8
Epstein H.E., Calef M.P., Walker M.D., Chapin F.S., Starfield A.M. Detecting changes in arctic tundra plant communities in response to warming over decadal time scales // Global Change Biology. 2004. № 10. P.1325–1334.
Hilli S., Starka S., Derome J. Litter decomposition rates in relation to litter stocks in boreal coniferous forests along climatic and soil fertility gradients // Appl. Soil Ecol. 2010. № 46. P. 200–208. https://doi.org/10.1016/j.apsoil.2010.08.012
Kuemmerle T., Kaplan J.O., Prishchepov A.V., Rylsky I., Chaskovskyy O., Tikunov V.S., Müller D. Forest transitions in Eastern Europe and their effects on carbon budgets // Global Change Biology. 2015. V. 21. № 8. P. 3049–3061. https://doi.org/10.1111/gcb.12897
Kuznetsova A.I., Geraskina A.P., Lukina N.V., Smirnov V.E., Tikhonova E.V., Shevchenko N.E., Gornov A.V., Ruchinskaya E.V., Tebenkova D.N. Linking Vegetation, Soil Carbon Stocks, and Earthworms in Upland Coniferous–Broadleaf Forests // Forests. 2021. № 12. P. 1179. https://doi.org/10.3390/f12091179
Lukina N.V., Tikhonova E.V., Danilova M.A., Bakhmet O.N., Kryshen A.M., Tebenkova D.N., Kuznetsova A.I., Smirnov V.I., Braslavskaya T.Yu., Gornov A.V., Shashkov M.P., Knyazeva S.V., Kataev A.D., Isaeva L.G., Zukert N.V. Associations between forest vegetation and the fertility of soil organic horizons in northwestern Russia // Forest Ecosystems. 2019. № 6. P. 34. https://doi.org/10.1186/s40663-019-0190-2
Nilsson M.-Ch., Wardle D.A. Understory vegetation as a forest ecosystem driver: evidence from the northern Swedish boreal forest // Front. Ecol. Environ. 2005. V. 3. № 8. P. 421–428.
Noreika N., Helm A., Öpik M., Jairus T., Vasar M., Reier Ü., Kook E., Riibak K., Kasari L., Tullus H., Tullus T., Lutter R., Oja E., Saag A., Randlane T. and Pärtel M. Forest biomass, soil and biodiversity relationships originate from biogeographic affinity and direct ecological effects // Oikos, 2019. № 128. P. 1653–1665. https://doi.org/10.1111/oik.06693
Orlova M.A., Lukina N.V., Tutubalina O.V. et al. Soil nutrient’s spatial variability in forest–tundra ecotones on the Kola Peninsula, Russia // Biogeochemistry. 2013. V. 113. № 1–3. P. 283–305.
Sayer E.J. Using experimental manipulation to assess the roles of leaf litter in the functioning of forest ecosystems // Biological Reviews. 2006. № 81. P. 1–31.
Tamminen P., Starr M., Kubin E. Element concentrations in boreal, coniferous forest humus layers in relation to moss chemistry and soil factors// Plant and soil. 2004. V. 259. № 1–2. P. 51–58.
Vesterdal L., Raulund-Rasmussen K. Forest floor chemistry under seven tree species along a soil fertility gradient // Canadian J. Forest Research. 1998. V. 28. № 11. P. 1636–1647. https://doi.org/10.1139/cjfr-28-11-1636
Zellweger F., De Frenne P., Lenoir J., Vangansbeke P., Verheyen K., Bernhardt-Römermann M., Baeten L., Hédl R., Berki I., Brunet J., Van Calster H., Chudomelová M., Decocq G., Dirnböck T., Durak T., Heinken T., Jaroszewicz B., Kopecký M., Máliš F., Macek M., Marek M., Naaf T., Nagel T.A., Ortmann-Ajkai A., Petřík P., Pielech R., Reczyńska K., Schmidt W., Standovár T., Świerkosz K., Teleki B., Vild O., Wulf M., Coomes D. Forest microclimate dynamics drive plant responses to warming // Science. 2020. V. 368. № 6492. P. 772–775. https://doi.org/10.1126/science.aba6880
Дополнительные материалы отсутствуют.