

Лесоведение, 2022, № 3, стр. 297-311
Крупные древесные остатки в муссонных тропических лесах Вьетнама
Е. А. Капица a, *, А. Н. Кузнецов d, e, С. П. Кузнецова d, e, В. О. Лопес де Гереню f, В. В. Каганов g, И. Н. Курганова f, Е. В. Кушневская a, h, Е. В. Шорохова a, b, c
a Санкт-Петербургский государственный лесотехнический университет им. С.М. Кирова
194021 Санкт-Петербург, Институтский пер., д. 5, Россия
b Институт леса КарНЦ РАН
185910 Республика Карелия, Петрозаводск, ул. Пушкинская, д. 11, Россия
c Институт природных ресурсов Финляндии (Луке)
FI-00790 Хельсинки, ул. Лаатоканкартаноонкаари, д. 4, Финляндия
d Российско-Вьетнамский Тропический научно-исследовательский и технологический центр
Ханой, Кау Зай, Нгиа До, ул. Нгуен Ван Хуен, Вьетнам
e Институт проблем экологии и эволюции имени А.Н. Северцова РАН
119071 Москва, Ленинский пр-т, д. 33, Россия
f Институт физико-химических и биологических проблем почвоведения РАН
142290 Московская область, Пущино, ул. Институтская, д. 2, стр. 2, Россия
g Центр по проблемам экологии и продуктивности лесов РАН
117997 Москва, ул. Профсоюзная, д. 84/32, стр. 14, Россия
h Санкт-Петербургский государственный университет
199034 Санкт-Петербург, Университетская наб., д. 7–9, Россия
* E-mail: kapitsa@list.ru
Поступила в редакцию 03.11.2021
После доработки 16.01.2022
Принята к публикации 02.02.2022
- EDN: CXPMDZ
- DOI: 10.31857/S0024114822030081
Аннотация
Для тропических лесов, в особенности для лесных экосистем Юго-Восточной Азии существуют лишь фрагментарные оценки пулов КДО и их динамики. В работе впервые исследовали запас и структурное разнообразие крупных древесных остатков (КДО) в лесах, не затронутых коммерческими рубками на территориях национальных парков (НП) Бидуп-Нуйба и Бу Зя Мап республики Вьетнам. В низкогорных диптерокарповых лесах НП Бу Зя Мап средний запас КДО был почти в два раза ниже среднего запаса в полидоминантных горных лесах НП Бидуп-Нуйба: 44 м3 га–1 против 78 м3 га–1. Масса КДО линейно зависела от их объема и составила в среднем 24 т га–1. Проективное покрытие КДО составляло в среднем 269 м2 га–1. Среди категорий субстрата преобладал валеж, составляя 42 и 31% в лесах НП Бидуп-Нуйба и Бу Зя Мап соответственно. Доля крупных ветвей в горных лесах НП Бидуп-Нуйба составила 8%, тогда как в низкогорных лесах НП Бу Зя Мап она достигала 33% объема КДО. В НП Бу Зя Мап отмечена очень высокая активность термитов – на 73% исследованных объектов КДО. Процент термитных ходов, частично заполненных почвой, в среднем составил 32% объема КДО. Достаточно высокое проективное покрытие КДО с разнообразными путями потери их массы – потребления беспозвоночными, микогенного ксилолиза коррозионного и деструктивного типов позволяют предположить их существенную роль в почвообразовательных процессах. Выявлены сходства в запасах и размерах КДО в исследуемых муссонных тропических лесах Южного Вьетнама и в коренных таежных ельниках европейской части России с мелкоконтурной оконной динамикой. Преобладание валежа среди категорий КДО и близкая к колоколообразной форма распределения объема КДО по классам разложения дополняют сходство пула КДО в тропических низкогорных и горных полидоминантных лесах с таежными разновозрастными ельниками. Однако высокая доля ветвей в структуре КДО, в особенности в низкогорных лиственных полидоминантных лесах, отличает КДО тропических лесов от КДО лесов таежного и умеренного пояса.
Ключевая роль крупных древесных остатков (КДО) – сухостоя, валежа, зависших стволов, пней, крупных ветвей и корней – в функционировании таежных и широколиственных лесов продемонстрирована в многочисленных работах отечественных и зарубежных исследователей (Замолодчиков, 2009; Стороженко, 2011; Stokland et al., 2012; Иванов и др., 2020). Для тропических лесов существуют лишь фрагментарные оценки пулов КДО и их динамики (Chambers et al., 2000; Clark et al., 2002; Palace et al., 2012; Giardina, 2019). Соответственно, роль КДО в формировании биоразнообразия тропических лесов и их роли в почвообразовании и круговоротах биогенных элементов остается слабоизученной. Для лесных экосистем Юго-Восточной Азии некоторая информация о КДО представлена лишь в нескольких работах (Gale, 2000; Pfeifer et al., 2015). Для лесов этих регионов оценка экологической роли КДО начинается со сбора эмпирических данных, характеризующих их объем, массу и проективное покрытие в лесах различного породного состава древостоя и находящихся в разных ландшафтно-географических условиях.
Динамика пула КДО зависит от соотношения скорости отпада в результате отмирания деревьев или их отдельных фракций и разложения (Капица и др., 2012). В свою очередь, процессы разложения КДО включают биогенный ксилолиз, приводящий к потере массы и эмиссии CO2 в атмосферу, фрагментацию и поедание беспозвоночными, а также выщелачивание. На глобальном уровне биогенный ксилолиз является основным процессом, приводящим к потере массы КДО (Russell et al., 2015). В таежных лесах длительность процесса ксилолиза зависит от климатических факторов, в основном, от температуры, а также от древесной породы, типа древесного отпада и других характеристик субстрата и варьирует от нескольких десятков до нескольких сотен лет (Shorohova, Kapitsa, 2016). In vitro, при оптимальных температуре и влажности, ксилолиз коррозионного типа, вызываемый грибом одного вида, приводит к полному разложению образца древесины (Соловьев, 1992). При деструктивном типе микогенного ксилолиза сохраняется остаток, составляющий по массе около 30% (Соловьев, 1992). Однако в природных условиях (in situ), при варьирующих условиях внешней среды и составе микоценоза, КДО разлагаются, по всей видимости, не полностью, а часть разложившегося древесного вещества переходит в почву (Magnússon et al., 2016). В северных широтах возможно также так называемое “захоронение” КДО под слоем сфагновых мхов с увеличением длительности процессов ксилолиза на неопределенно долгое время (Moroni et al., 2015).
В тропических лесах с влажным и теплым климатом не менее важная, по сравнению с грибами, роль в разложении КДО принадлежит термитам (Lopes de Gerenyu et al., 2015; Shorohova et al., 2021), деятельность которых может приводить к потере массы КДО на 70 и более процентов (Shorohova et al., 2021). Первыми шагами в изучении динамики КДО в муссонных тропических лесах можно считать оценку распределения их объема по классам разложения и категориям (сухостой, валеж, зависшие стволы, пни, ветви), а также диагностику путей потери массы и объема КДО.
Цель настоящего исследования состояла в определении вариабельности запасов и структурного разнообразия КДО в низкогорных лиственных полидоминантных и горных хвойно-широколиственных лесах Южного и Центрального Вьетнама. Задачи исследования включали оценку: (1) объема, массы, проективного покрытия и размеров КДО; (2) распределения КДО по классам разложения и категориям; (3) количественного соотношения путей потери массы КДО в результате ксилолиза коррозионного и деструктивного типов и потребления беспозвоночными, в первую очередь, термитами.
ОБЪЕКТЫ И МЕТОДИКА
Полевые и лабораторные работы. Исследование выполнено на территории национальных парков Бидуп-Нуйба (Bidoup Nui Ba) и Бу Зя Мап (Bu Gia Map) Социалистической Республики Вьетнам в лесах, не затронутых коммерческими рубками (табл. 1). Все КДО образованы в результате естественного отпада отдельных деревьев или небольших групп в результате усыхания или реже ветровала и бурелома, а также отпада ветвей.
Таблица 1.
Характеристика районов исследования (Горбунов, 2019; Кузнецов, 2003)
| Национальный парк | Географические координаты районов исследований | Среднегодовая температура, °C | Диапазон высот над уровнем моря, м | Среднегодовое количество осадков, мм |
|---|---|---|---|---|
| Бидуп-Нуйба | 12°10.885′–11.235′ с.ш., 108°40.469′–41.406′ в.д. | +18.2 | 1470–1570 | 1860 |
| Бу Зя Мап | 12°11.788′–12.149′ с.ш., 107°12.248′–12.433′ в.д. | +26.2 | 387–737 | 2469 |
В ходе геоботанических описаний 14-ти растительных сообществ в НП Бидуп-Нуйба и пяти растительных сообществ в НП Бу Зя Мап был определен состав древесного яруса с определением принадлежности древесных пород с точностью до рода, и, где было возможно, до вида. Запасы стволовой древесины растущих деревьев были рассчитаны по данным сплошной перечислительной таксации на пробных площадях размером 0.1–0.125 га. К перечету принимались деревья с диаметром стволов 8 и более см на высоте 1.3 м. После группировки по породной принадлежности (с точностью до вида, рода или семейства) выполнялось измерение высоты деревьев с последующим построением графика высот для каждой из групп. Объем древесины вычислялся по формуле:
где М – объемный запас древесины, м3 · га–1; ΣG – площадь или сумма площадей сечения, м2 · га–1; Hср – средняя или текущая высота породы, м; Fср – расчетное или среднее видовое число дерева или породной группы. Для определения видовых чисел проводили измерения диаметров стволов на половине высоты деревьев с последующим расчетом видовых чисел по формуле Шиффеля.
Запасы и структурное разнообразие КДО в лесных биогеоценозах (БГЦ) анализировали по результатам учетов на трансектах длиной 50 и шириной 4 м, заложенных в 2018–2019 гг. в направлениях с севера на юг и с запада на восток. В инвентаризацию были включены все объекты КДО более 6 см. В национальном парке Бидуп-Нуйба заложена 41 трансекта в полидоминантных горных лесах, образованных: 1) только покрытосеменными древесными породами семейств: буковых (Fagaceae), элеокарповых (Elaeocarpaceae), магнолиевых (Magnoliaceae), чайных (Theaceae), миртовых (Myrtaceae), сапиндовых (Sapindaceae), молочайных (Euphorbiaceae), березовых (Betulaceae), анноновых (Annonaceae), ореховых (Juglandaceae), гамамелисовых (Hamamelidaceae), сапотовых (Sapotaceae), алтингиевые (Altingiaceae), бобовых (Fabaceae), лимонниковых (Schisandraceae), стираксовых (Styracaceae), кизиловых (Cornaceae), эбеновых (Ebenaceae), пентафилаксовых (Pentaphylacaceae), протейных (Proteaceae), мальвовых (Malvaceae), мареновых (Rubiaceae), пальмовых (Palmae); 2) покрытосеменными с участием голосеменных: сосны алеппской (Pinus dalatensis), дакрикарпуса (Dacrycarpus imbricatus), дакридиума кипарисового (Dacrydium elatum) вышеперечисленных видов и сосны Кремпфа (Pinus krempfii) и фокиении (Fokienia hodginsii); 3) а также в лесах, образованных сосной кесия (Pinus kesiya). В горном массиве сформировались полидоминантные смешанные леса со сложной (4 подъяруса) и среднесложной (3 подъяруса) вертикальной структурой древостоев – на выположенных участках и пологих склонах, и с простой (1–2 подъяруса) структурой – на гребнях хребтов, вершинах и крутых склонах (Горбунов и др., 2018). Лиственные леса расположены в поймах рек и на пологих склонах на высотах 850–1600 м над уровнем моря. Смешанные хвойно-широколиственные леса приурочены к высотам 1500–1700 м. Горные леса, образованные древостоями сосны кесия, расположены на вершинах и открытых гребнях горных хребтов на высотах 1500–1700 м. На высотах 1700– 2000 м над у. м. произрастают моховые облачные леса и криволесья.
Почвенный покров НП Бидуп-Нуйба представлен двумя основными типами почв: зональными желтоземами (желтые феррсиаллитные, Haplic (или Umbric) Ferralsols согласно WRB, 2014) и интразональными аллювиальными почвами (Fluvisols) в пойме реки (Лебедев и др., 2019). В зависимости от почвообразующей (материнской) породы, которая в значительной степени зависит от положения в рельефе, желтоземы могут иметь различную мощность, минералогический и гранулометрический состав, а также отличаться по запасам органического углерода и других биогенных элементов. Так, на поверхности гребня структурной гряды под сосново-широколиственным лесом формируются желтоземы типичные маломощные легкосуглинистые на глинистой коре выветривания дацитов, а в верхней части склонов северной экспозиции под среднесомкнутым широколиственным лесом с развитым травяным ярусом формируются желтоземы лессивированные среднемощные супесчаные на щебнисто-каменном делювии дацитов (Лебедев и др., 2019). В средней части склона структурной гряды под среднесомкнутым широколиственным лесом с тремя древесными подъярусами и хорошо развитым травяным ярусом встречаются желтоземы типичные маломощные среднесуглинистые на щебнисто-каменном делювии дацитов. На холодных склонах северной экспозиции, где скорость биологического круговорота значительно ниже, чем на южных, происходит формирование более мощной подстилки с возможным формированием оторфованного горизонта (Лебедев и др., 2019).
На вершине гряды был заложен неполнопрофильный разрез с целью характеристики основных почвенных свойств в верхней 50-сантиметровой толще. Образцы почв отбирали послойно на глубинах: 0–5, 5–10, 10–20, 20–30 и 30–50 см, анализы проводили с использованием общепринятых в почвоведении методов22. Согласно классификации Качинского (Шеин, 2005), текстура почвы в слое 0–5 см соответствовала легкосуглинистой, а в слое 20–50 см постепенно становилась тяжелосуглинистой (табл. 2). Во всей 50-сантиметровой толще плотность почв невысока и варьировала от 0.84 до 1.01 г/см3. Верхний 10-сантиметровый слой содержал значительное количество органического углерода (54–83 г С/кг почвы), которое резко убывало с глубиной. Отношение C/N в верхней части профиля варьировало от 24 до 27, что свидетельствует о невысокой степени гумификации органического вещества (ОВ). Наиболее обогащенный углеродом слой 0–5 см отличался также повышенной потенциальной кислотностью (pH (KCl) = 3.2), с глубиной значения рН повышались на 0.4–0.6 единицы.
Таблица 2.
Характеристика основных свойств почв в национальных парках Бу Зя Мап и Бидуп-Нуйба
| Глубина, см | Содержание фракций (%) | Плотность, г/см3 | рH(КCl) | C | N | C/N | ||
|---|---|---|---|---|---|---|---|---|
| песок1 | глина2 | ил3 | г/кг почвы | |||||
| Национальный парк Бидуп-Нуйба (хвойно-лиственный лес, желтозем) | ||||||||
| 0–5 | 74.6 | 25.4 | 9.48 | 0.84 | 3.23 | 82.7 | 3.1 | 26.5 |
| 5–10 | 61.8 | 38.2 | 20.28 | 0.92 | 3.60 | 54.2 | 2.3 | 23.6 |
| 10–20 | 57.6 | 42.4 | 22.56 | 1.01 | 3.81 | 21.5 | 1.1 | 18.7 |
| 20–30 | 52.68 | 47.32 | 28.56 | 1.07 | 3.86 | 11.7 | 0.8 | 15.5 |
| 30–50 | 54.24 | 45.76 | 22.6 | 1.01 | 3.81 | 9.8 | 0.6 | 16.0 |
| Национальный парк Бу Зя Мап (Диптерокарпусовый древостой, краснозем) | ||||||||
| 0–5 | 22.84 | 77.16 | 48.20 | 0.99 | 3.82 | 39.8 | 3.2 | 12.4 |
| 5–10 | 18.24 | 81.76 | 54.40 | 0.93 | 3.84 | 33.3 | 2.8 | 12.0 |
| 10–20 | 8.89 | 91.11 | 60.27 | 0.97 | 3.88 | 27.1 | 2.3 | 12.0 |
| 20–30 | 5.90 | 94.10 | 64.27 | 0.92 | 3.88 | 23.6 | 1.8 | 12.9 |
| 30–40 | 6.26 | 93.74 | 68.63 | 0.87 | 4.02 | 19.9 | 1.4 | 14.3 |
| 40–50 | 7.34 | 92.66 | 67.78 | 0.91 | 4.08 | 16.9 | 1.4 | 12.4 |
| Национальный парк Бу Зя Мап (Лагерстремиевый древостой, краснозем) | ||||||||
| 0–5 | 52.72 | 47.28 | 19.32 | 0.84 | 4.24 | 58.1 | 4.5 | 13.0 |
| 5–10 | 43.68 | 56.32 | 25.84 | 0.86 | 3.74 | 38.0 | 3.5 | 10.7 |
| 10–20 | 40.18 | 59.82 | 29.31 | 0.99 | 3.62 | 30.1 | 2.8 | 10.9 |
| 20–30 | 26.86 | 73.14 | 40.19 | 1.10 | 3.58 | 22.6 | 2.1 | 10.6 |
| 30–40 | 24.11 | 75.89 | 45.23 | 1.10 | 3.57 | 21.4 | 2.0 | 10.9 |
| 40–50 | 17.99 | 82.01 | 49.83 | 1.10 | 3.62 | 18.9 | 1.9 | 9.7 |
В национальном парке Бу Зя Мап в низкогорных высокоствольных диптерокарповых лесах на склонах и на пойменных террасах заложено 23 трансекты. Отдельные участки характеризовались преобладанием Dipterocarpus costatus, D. turbinatus, D. alatus, Hopea odorata, Lagerstroemia calyculata, Swintonia floribunda.
Наиболее распространенным типом почв в пределах исследуемой территории НП Бу Зя Мап являлись красные ферраллитные почвы (Feralsols, согласно WRB, 2014). В разных типах древостоев (диптерокарповом – на вершине холма и лагерстремиевом – на относительно пологом участке) нами были заложены неполнопрофильные разрезы, и из верхних 0–50 см послойно отобраны почвенные образцы: с шагом 0–5 см в пределах первых 10 см, а далее каждые 10 см до глубины 50 см (табл. 2). Гранулометрический состав почв изменялся от тяжелосуглинистого до глинистого в пределах верхнего 5-сантиметрового слоя и затем постепенно утяжелялся вниз по профилю, свидетельствуя о процессах лессиважа (выноса илистой фракции из верхних слоев в нижние). Несмотря на тяжелый гранулометрический состав, плотность почв невысока: <1 г/см3 в диптерокарпусовом насаждении и увеличивается от 0.84 до 1.10 г/см3 – в лагерстремиевом, что, по-видимому, может быть связано с характерной псевдопесчаной структурой почв. Значения потенциальной кислотности почв в НП Бу Зя Мап оказались чуть выше, чем в почвах НП Бидуп Нуйба. и изменялись от 3.57 до 4.4 ед. рН. Верхний 5-сантиметровый слой почвы под лагерстремией содержал углерода и азота в 1.5 раз больше, чем тот же слой почвы в диптерокарпусовом насаждении (табл. 2). За исключением верхнего слоя, отношение С/N в почве лагерстремиевого насаждения несколько меньше по величине, чем в диптерокарпусовом, что свидетельствует о более глубокой гумификации ОВ в этих почвах.
Для валежа, зависших деревьев и крупных ветвей учитывали класс разложения (Shorohova et al., 2021, 2022) (табл. 3) и измеряли с помощью сантиметровой ленты окружность в месте пересечения ходовой линии. Использовали метод учета валежа на линейных трансектах (Грабовский, Замолодчиков, 2012).
Таблица 3.
Характеристика классов ксилолиза КДО на основании степени разложения древесины, определяемой по мягкости (глубине проникновения ножа) и влажности (Shorohova et al., 2021, 2022)
| Класс разложения | Характеристика класса разложения | |
|---|---|---|
| Национальный парк Бидуп-Нуйба | Национальный парк Бу Зя Мап | |
| 1 | Объем разложившейся древесины составляет 0–10%; остальная древесина не затронута ксилолизом. Кора и ветви чаще всего присутствуют, плодовые тела дереворазрушающих грибов не наблюдаются, могут встречаться лишайники. Лезвие ножа проникает внутрь на несколько миллиметров |
Объем разложившейся древесины составляет 0–10%; остальная древесина не затронута ксилолизом. Древесина относительно сухая. Лезвие ножа проникает внутрь на несколько миллиметров. Туннели термитов занимают не более 10% от объема ствола |
| Среднее значение плотности древесины, г см–3 | ||
| 0.56 | 0.628 | |
| 2 | Объем сильно разложившейся древесины составляет 0–10%; остальная древесина или не затронута ксилолизом вообще, или слегка размягчена. Кора и ветви присутствуют частично или полностью, появляются первые признаки заселения дереворазрушающих грибов (гифы) и мхов. Лезвие ножа проникает внутрь менее чем на 2 см | Древесина слегка разложившаяся. Лезвие ножа проникает внутрь менее чем на 2 см. Доля термитных ходов в КДО варьирует от 0 до 30% от объема ствола |
| Среднее значение плотности древесины, г см–3 | ||
| 0.490 | 0.501 | |
| 3 | Разложением затронуто от 11 до 100% древесины. Кора и ветви присутствуют частично, наблюдается активное освоение субстрата дереворазрушающими грибами и мхами, которые покрывают ствол частично или полностью | Древесина сильно разложившаяся, легко “режется” ножом, ее объем составляет 10–100%. В большинстве случаев древесина относительно влажная. Лезвие ножа проникает насквозь. Доля термитных ходов в КДО варьирует от 31 до 60% |
| Среднее значение плотности древесины, г см–3 | ||
| 0.370 | 0.332 | |
| 4 | Вся древесина сильно разложилась. Образцы древесины дробятся на отдельные волокна и могут быть измельчены пальцами. Кора и ветви отсутствуют, мхи покрывают ствол полностью | Вся древесина сильно разложилась. Образцы древесины дробятся на отдельные волокна и могут быть измельчены пальцами. Древесина очень влажная. Доля термитных ходов в КДО варьирует от 61 до 80% |
| Среднее значение плотности древесины, г см–3 | ||
| 0.280 | 0.169 | |
| 5 | Тип и границы гнилей трудно различимы. Активно идет процесс гумификации. Кора и ветви отсутствуют, мхи покрывают ствол полностью | Тип и границы гнилей трудно различимы. Активно идет процесс гумификации. Форма стволов сильно изменена. Туннели термитов занимают более 80% от объема ствола |
| Среднее значение плотности древесины, г см–3 | ||
| 0.150 | 0.071 | |
Объем КДО классам разложения без учета объема древесины, потребленной термитами, рассчитывали по формуле (Stahl et al., 2001):
где V – объем КДО данного класса разложения, di – диаметр i-го объекта в месте пересечения ходовой линии, рассчитанный на основании измерения длины окружности объекта, Lj – длина j-й ходовой линии, S – площадь, в данном случае равная 1 га.Для сухостоя и пней диаметром более 4 см регистрировали класс разложения, диаметр основания и вершины (или диаметр на высоте 1.3 м) и высоту. Объем пней рассчитывали по формуле усеченного конуса. Объем сухостойных деревьев рассчитывали умножением площади сечения на высоте 1.3 м на видовое число и высоту. Видовое число рассчитывали по формуле Шиффеля для каждой древесной породы по данным таксационных измерений на постоянных пробных площадях. Для каждого дерева измеряли диаметр на высоте 1.3 м, на половине высоты дерева и высоту. При неизвестности видовых чисел исследуемых древесных пород использовали видовые числа для осины, или, для голосеменных – сосны (Тетюхин и др., 2004).
Площадь проективного покрытия валежа, зависших деревьев и крупных ветвей рассчитывали для каждой трансекты делением их суммарного объема на средний диаметр. Для пней и сухостоя проективное покрытие считали равным площади сечения ствола.
Для перевода объема КДО в массу со всех КДО, попадающих в учет на трансекте, отбирали образцы правильной геометрической формы c участков разных стадий разложения. Объем выборки для КДО лесов Бидуп-Нуйба и Бу Зя Мапа составил 360 и 157 объектов соответственно. Количество и процент участков, разлагающихся по типам коррозии и деструкции и стадий разложения, а также пропорции древесины, потребленной беспозвоночными, включая термитные ходы, заполненные почвой, оценивали визуально на спилах. Отобранные образцы замеряли с целью определения объема. Сильно разложившиеся фракции отбирали при помощи почвенного бура. Объем образцов определяли геометрически с использованием формул параллелепипеда, усеченного конуса и куба, для сильно разложившихся образцов объем принимали равным объему бура. Все образцы высушивали в сушильном шкафу при температуре 103°С в течение 48 ч до абсолютно сухого состояния, взвешивали, рассчитывали базисную плотность древесины (ρ, г см–3) по формуле (Полубояринов, 1976):
где: m – масса в абсолютно сухом состоянии, г, V – объем образца, см3.Базисную плотность рассчитывали для каждого образца по отдельности, а затем переводили с ее помощью в массу всего ствола с учетом процентного соотношения по объему участков разных стадий разложения, пустот и термитных ходов.
Статистический анализ данных. Связь массы, проективного покрытия и объема КДО оценивали с помощью линейной регрессионной модели. Зависимости запаса КДО от типа биогеоценоза оценивали с использованием обобщенной линейной множественной регрессионной модели в среде R (R Core Team, 2013).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Объем КДО
Объем КДО в двух изученных лесных массивах в среднем составил 66 м3 га–1. В лесах Бу Зя Мапа средний запас КДО составил 44 м3 га–1 (Min = 5 м3 га–1; Max = 144 м3 га–1) и был почти в два раза ниже среднего запаса в Бидуп-Нуйба – 78 м3 га–1 (Min = 13 м3 га–1; Max = = 226 м3 га–1) (рис. 1). Средние объемы стволов живых деревьев составили 502.9 м3 га–1 и 440.6 м3 га–1, соответственно. Ключевую роль в формировании запасов древесины в лесах НП Бу Зя Мап играют Swintonia floribunda и Lagerstroemia calyculata, для НП Бидуп-Нуйба наибольшую роль играют представители родов Syzygium, Lithocarpus и Camellia. Особенностью горных лесов НП Бидуп-Нуйба является наличие в древостоях представителей голосеменных – Fokienia hodginsii и Pinus krempfii, отличающихся долговечностью и значительными размерами. Старовозрастные экземпляры данных видов могут иметь объем ствола более 10 м3, и их наличие или отсутствие на пробных площадях и учетных трансектах оказывает существенное влияние на получаемые данные о пространственном распределении запасов стволовой древесины и крупных древесных остатков.
Доля КДО в совокупных запасах древесины обследованных лесных массивов национальных парков составила, в среднем, 8.8% и 15%. Важной отличительной чертой рассматриваемых древостоев является очень малое количество сухостойных деревьев. В НП Бу Зя Мап средний объем сухостоя составил 0.3 м3 га–1, на одной из пробных площадей сухостой отсутствует полностью. В НП Бидуп-Нуйба средний запас сухостоя имеет величину 2.1 м3 га–1. В обоих случаях доля сухостоя составляет менее 1% от общего запаса деревьев, находящихся на корню и еще меньшую долю в совокупном запасе живой и мертвой древесины на пробных площадях. Наиболее вероятной причиной этого является отмеченная ранее высокая активность термитов и дереворазрушающих грибов, использующих древесину погибших деревьев в качестве питательного субстрата. Наиболее доступные элементы деревьев – корни и комлевые части стволов разрушаются в первую очередь, в результате чего дерево теряет устойчивость и вскоре переходит в пул крупных древесных остатков. Следует отметить, что быстрому выпадению из древостоя погибших деревьев не препятствуют и хорошо развитые досковидные корни. На одной из пробных площадей в НП Бу Зя Мап крупный экземпляр Swintonia floribunda, в декабре 2018 года находившийся в составе древостоя, в марте 2019 года был отмечен, как сухостой, а уже в декабре 2019 года оказался в пуле КДО. Таким образом, различия в запасах КДО двух изучаемых лесных массивов обусловлены их географическим положением, породным составом древостоев и климатическими условиями. Более низкие температуры воздуха позволяют предположить более низкую скорость разложения КДО и, соответственно, более активное их накопление в лесах Бидуп-Нуйба, по сравнению с лесами Бу Зя Мапа.
Вариация запасов КДО в лесах Бидуп-Нуйба была обусловлена породным составом древостоя, положением в рельефе и почвенными условиями исследуемых БГЦ. Средние объемы КДО возрастали в ряду: туманные леса, расположенные на склонах на самых высоких отметках высот над уровнем моря (34 м3 га–1) > пойменные лиственные леса (61 м3 га–1) > смешанные хвойно-широколиственные леса (79 м3 га–1) > > кезиевые леса на вершинах холмов (88 м3 га–1) > > смешанные хвойно-широколиственные леса с участием фокиении (109 м3 га–1) (рис. 2). Результаты обобщенной линейной модели показали достоверные отличия запасов КДО в разных БГЦ (табл. 4).

Таблица 4.
Результаты статистического анализа данных и параметры обобщенной линейной модели для разных БГЦ в национальном парке Бидуп-Нуйба
| Средние значения коэффициентов модели с распределением Пуассона (SE) | Z(p) |
|---|---|
| Свободный член (туманные леса): 3.51 (0.09) | 40.65 (<0.001) |
| Смешанные хвойно-широколиственные леса: 0.78 (0.09) | 8.55 (<0.001) |
| Смешанные хвойно-широколиственные леса с участием фокиении: 1.28 (0.09) | 13.84 (<0.001) |
| Кезиевые леса: 0.99 (0.10) | 9.55 (<0.001) |
| Пойменные лиственные леса: 0.64 (0.10) | 6.54 (<0.001) |
В национальном парке Бу Зя Мап статистически достоверных различий в вариабельности запасов КДО в зависимости от высоты над уровнем моря и положения в рельефе не выявлено. При относительно небольшом объеме выборки мы можем лишь с определенной долей вероятности назвать факторы, регулирующие объемы КДО в изучаемых низкогорных диптерокарповых лесах. Это может быть относительно равномерная скорость отпада древостоя и отмирания ветвей, а также относительно высокая скорость их разложения при активном участии термитов. Почти в три раза более высокие средние величины запасов КДО найдены в нескольких равнинных смешанных диптерокарповых лесах острова Борнео (117 м3 га–1; Gale, 2000) и острова Суматра (116 м3 га–1; Yoneda et al., 1990). В другом исследовании для равнинных лесов острова Суматра объем КДО оценен в 28 га–1 (Meriem et al., 2016). Интересно, что при более низких высотах над уровнем моря (55–320 м над у. м. по сравнению с 387–737 м над у. м. в нашей работе) запасы КДО зависели от положения БГЦ в рельефе (Gale, 2000).
В горных лесах национального парка Бидуп-Нуйба более высокое разнообразие состава и структуры древесного яруса обусловливают более высокую вариабельность процессов отпада. Изучение качества КДО и их динамики в годы работ на территории парка Бидуп-Нуйба позволяют предположить их более низкую скорость разложения, чем в лесах Бу Зя Мапа. Ведущая роль в процессе ксилолиза в парке Бидуп-Нуйба принадлежит дереворазрушающим грибам, активность которых, возможно, варьирует в зависимости от сезона и древесной породы КДО. В результате происходит локальное накопление КДО.
Общий объем КДО и его вариабельность в лесах обоих национальных парков сравнимы с таковыми в коренных таежных ельниках европейской части России с мелкоконтурной оконной динамикой (Shorohova et al., 2016).
Масса, проективное покрытие и размеры КДО
Масса КДО составила в среднем 24 т га–1 (29 и 15 т га–1 в лесах Бидуп-Нуйба и Бу Зя Мап соответственно) и линейно зависела от их объема (рис. 3а). Средние величины массы КДО сравнимы с таковыми в нескольких старовозрастных лесах островов Борнео (21.6 и 57.1 т га–1; Pfeifer et al., 2015) и Суматра (11 т га–1; Meriem et al., 2016; 39 т га–1; Yoneda et al., 1990). С позиций углеродного цикла важным отличием КДО коренных тропических лесов от таежных является более слабая связь массы с объемом КДО (Капица и др., 2012).
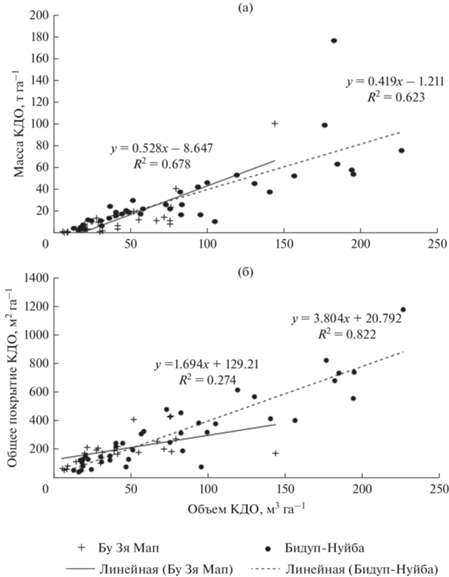
Проективное покрытие КДО изменялось от 32 до 1178 м2 га–1, в среднем составляя 269 м2 га–1 (рис. 3б). Таким образом, в изучаемых лесных экосистемах проективное покрытие КДО может достигать 12% площади поверхности, что указывает на значительную роль КДО в процессах почвообразования. Теснота связи между проективным покрытием и объемами КДО связана с существенной долей сухостоя и пней в их объеме.
Средний диаметр стволов КДО составил 26 см (30 и 24 см в лесах Бу Зя Мап и Бидуп-Нуйба соответственно). В таежной зоне средние диаметры КДО близки к таковым в муссонных тропических лесах. Средний диаметр КДО в ельниках средней тайги составил 24 см, в сосняках – 28 см, а в ельниках и сосняках северной тайги – 24 и 28 см, соответственно (Капица и др., 2012).
Структурное разнообразие КДО
Среди категорий субстрата преобладал валеж, составляя 42 и 31% в лесах Бидуп-Нуйба и Бу Зя Мап соответственно (рис. 4а). Особенностью древесного отпада в тропических лесах является отмирание отдельных ветвей живого дерева (Denslow, 1987; Кузнецов, 2003). Доля крупных ветвей в горных лесах Бидуп-Нуйба составила 8%, тогда как в низкогорных лесах Бу Зя Мап она достигла 33% объема КДО. В туманных лесах, где большинство деревьев были многоствольными, КДО могли представлять собой часть живого дерева.
Рис. 4.
Распределение объема КДО по категориям субстрата (а) и классам разложения (б). Приведены средние величины и стандартные ошибки.
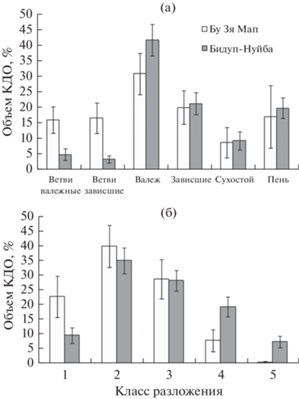
Распределение объема КДО по классам разложения характеризовалось колоколообразной формой с максимумами во втором классе разложения. Леса Бу Зя Мапа отличались от лесов Бидуп-Нуйба практически полным отсутствием КДО 5-го класса разложения и более высокой долей КДО 1-го класса разложения (23% в национальном парке Бу Зя Мап в отличие от 10% в национальном парке Бидуп-Нуйба) (рис. 4б).
Преобладание валежа среди категорий КДО и близкая к колоколообразной форма распределения объема КДО по классам разложения дополняет сходство пула КДО тропических низкогорных и горных полидоминантных лесов и таежных разновозрастных ельников. Однако высокая доля ветвей в структуре КДО, в особенности в низкогорных диптерокарповых лесах, отличает КДО тропических лесов от КДО лесов таежного и умеренного пояса. Анализ перечисленных выше сходств и различий в проявлении экологических функций КДО в разных типах тропических лесов, в особенности их роли в круговоротах веществ и биоразнообразии, требует дальнейших исследований.
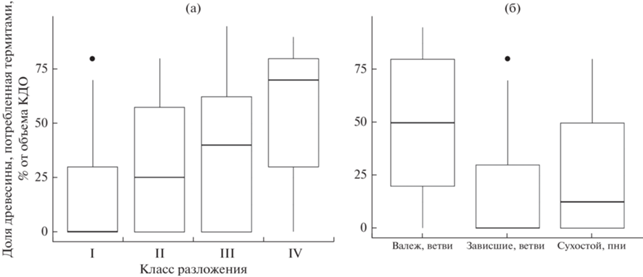
Активность термитов в национальном парке Бу Зя Мап очень высока. Согласно нашим наблюдениям, 73% всех исследованных объектов КДО подвергалось их воздействию (Shorohova et al., 2021). Процент термитных ходов, частично заполненных почвой, по объему КДО в среднем составлял 32%. Объемная доля древесины, потребляемая термитами в КДО 1-го, 2-го, 3-го и 4-го классов разложения, составила в среднем 15, 32, 38 и 54% соответственно (рис. 5а). Доля древесины, потребляемая термитами, зависела от положения КДО и составляла в среднем 17% для зависших стволов и ветвей, 28% для пней и сухостоя и 49% для валежа и валежных ветвей (рис. 5б).
Рис. 5.
Доля древесины, потребляемая термитами, в зависимости от класса разложения (а) и категории КДО (б). На графике показаны медианные, максимальные и минимальные значения.
Микогенный ксилолиз, т.е. разложение древесины, не потребленной термитами, в низкогорных высокоствольных диптерокарповых лесах Бу Зя Мапа проходил исключительно по типу коррозии, случаев деструкции не выявлено. Для изучения причин подобной закономерности необходим анализ видового состава ксилотрофных грибов и их ферментных комплексов. В полидоминантных горных лесах Бидуп-Нуйба ведущую роль в процессах биогенного ксилолиза играют, по-видимому, дереворазрушающие грибы. Активность беспозвоночных отмечена лишь на 3-х процентах исследуемых объектов, которые потребили в среднем 8% объема древесины. Преобладающая часть исследованных КДО (88%) были поражены белой гнилью, 8% – бурой и лишь у 4% встречались участки как белой, так и бурой гнили. Доминирование белой гнили подтверждает важность грибов в процессах деградации лигнина в тропических лесах (Fujii, 2014). Большинство грибов бурой гнили активно участвуют в разложении голосеменных, в то время как грибы белой гнили предпочитают разлагать покрытосеменные (Krah et al., 2018). На наших участках преобладали КДО покрытосеменных видов. Преобладание микогенного пути ксилолиза над зоогенным в лесах Бидуп-Нуйба и высокая активность термитов, потребляющих КДО, в лесах Бу Зя Мапа, по-видимому, являются еще одной причиной почти двукратных различий в запасе КДО в двух изучаемых лесных массивах. Медленно разлагающиеся КДО в лесах Бидуп-Нуйба аккумулируются на более длительное время. Полученные результаты позволяют выдвинуть гипотезу о том, что климат и породный состав древостоя определяют пути потери массы КДО в изучаемых тропических лесах Южного Вьетнама.
ЗАКЛЮЧЕНИЕ
Выявлены сходства в запасах и размерах КДО в исследуемых муссонных тропических лесах Южного Вьетнама и в коренных таежных ельниках европейской части России с мелкоконтурной оконной динамикой. Преобладание валежа среди категорий КДО и близкая к колоколообразной форма распределения объема КДО по классам разложения дополняют сходство пула КДО тропических низкогорных и горных полидоминантных лесов и таежных разновозрастных ельников. Однако высокая доля ветвей в структуре КДО, в особенности в низкогорных лиственных полидоминантных лесах, отличает КДО тропических лесов от КДО лесов таежного и умеренного пояса.
Полученные результаты создают основу будущих исследований экосистемных функций КДО в муссонных тропических лесах Южного Вьетнама. Относительно высокие запасы КДО указывают на их значительную роль в круговороте основных биогенных элементов. Высокое структурное и породное разнообразие КДО создает множество различных экологических ниш, что позволяет предположить высокое разнообразие ксилофильных сообществ (сообществ, связанных с КДО на всех этапах их разложения). Достаточно высокое проективное покрытие КДО с разнообразными путями потери их массы – потребления беспозвоночными микогенного ксилолиза коррозионного и деструктивного типов определяют их существенную роль в почвообразовательных процессах.
Список литературы
Горбунов Р.В., Горбунова Т.Ю., Кузнецов А.Н., Кузнецова С.П., Лебедев Я.О., Нгуен Д.Н., Ву М. Особенности формирования элементов радиационного баланса среднегорных тропических лесов южного Вьетнама // Труды Карадагской научной станции им. Т.И. Вяземского – Природного заповедника РАН. 2019. № 4(12). С. 3–16.
Замолодчиков Д.Г. Оценка пула углерода крупных древесных остатков в лесах России с учетом влияния пожаров и рубок // Лесоведение. 2009. № 4. С. 3–15.
Иванов А.В., Замолодчиков Д.Г., Лошаков С.Ю., Комин А.Э., Косинов Д.Е., Браун М., Грабовский В.И. Вклад крупных древесных остатков в биогенный цикл углерода хвойно-широколиственных лесов юга Дальнего Востока России. Лесоведение. 2020. № 4. С. 357–366.
Горбунов Р.В., Кузнецов А.Н., Лебедев Я.О., Горбунова Т.Ю., Котлов И.П., Хой Н.Д. О некоторых особенностях структуры и функционирования горных тропических лесных ландшафтов Центрального Вьетнама и необходимости создания ландшафтноэкологического стационара // Труды Карадагской научной станции им. Т.И. Вяземского – природного заповедника РАН. 2018. № 3(7). С. 43–67.
Грабовский В.И., Замолодчиков Д.Г. Модели оценки запасов валежа по данным учетов на трансектах // Лесоведение. 2012. № 2. С. 66–73.
Капица Е.А., Шорохова Е.В., Кузнецов А.А. Пул углерода крупных древесных остатков в коренных лесах северо-запада Русской равнины // Лесоведение. 2012. № 5. С. 36–43.
Кузнецов А.Н. Тропический диптерокарповый лес: на примере сомкнутого высокоствольного влажного равнинного диптерокарпового леса массива Ма Да, Южный Вьетнам // РАН, Совместный российско-вьетнамский тропический научно-исследовательский и технологический центр (тропический центр). М.: ГЕОС, 2003. 138 с.
Лебедев Я.О., Горбунов Р.В., Горбунова Т.Ю., Кузнецов А.Н., Кузнецова С.П., Нгуен Т.В., Бобко Н.И., Капранов С.В. Некоторые особенности геохимических миграций в условиях соподчиненных ландшафтов среднегорных тропических лесов Южного Вьетнама // Труды Карадагской научной станции им. Т.И. Вяземского – Природного заповедника РАН. 2019. № 3(11). С. 3–16.
Полубояринов О.И. Плотность древесины. М.: Лесная пром-сть, 1976. 159 с.
Пулы и потоки углерода в наземных экосистемах России / Под ред. Заварзина Г.А. М.: Наука, 2007. 315 с.
Соловьев В.А. Микогенный ксилолиз, его экологическое и технологическое значение // Научные основы устойчивости лесов к дереворазрушающим грибам. М.: Наука, 1992. С. 140–171.
Стороженко В.Г. Древесный отпад в коренных лесах Русской равнины // М.: Тов-во КМК, 2011. 122 с.
Тетюхин С.В., Минаев В.Н., Богомолова Л.П. Лесная таксация и лесоустройство. Нормативно-справочные материалы по северо-западу РФ. Санкт-Петербург: ЛТА. 2004. 369 с.
Шеин Е.В. Курс физики почв. М.: Изд-во МГУ, 2005. 432 с.
Chambers J.Q., Higuchi N., Schimel J.P., Ferreira L.V., Melack J.M. Decomposition and carbon cycling of dead trees in tropical forests of the central Amazon // Oecologia. 2000. V. 122. P. 380–388.
Clark D.B., Clark D.A., Brown S., Oberbauer S.F., Veldkamp E. Stocks and flows of coarse woody debris across a tropical rain forest nutrient and topography gradient // Forest Ecology and Management. 2002. V. 164. P. 237–248.
Denslow J.S. Tropical rainforest gaps and tree species diversity // Annual Review in Ecology and Systematics. 1987. V. 18. P. 431–451.
Fujii K. Soil acidification and adaptations of plants and microorganisms in Bornean tropical forests // Ecol Res. 2014. V. 29. P. 371–381.
Gale N. The aftermath of tree death: coarse woody debris and the topography in four tropical rain forests // Canadian J. Forest Research. 2000. V. 30. P. 1489–1493.
Giardina C.P. Advancing Our Understanding of Woody Debris in Tropical Forests. Ecosystems. 2019. V. 22. P. 1173–1175.
Krah F.S., Bässler C., Heibl C., Soghigian J., Schaefer H., Hibbett D.S. Evolutionary dynamics of host specialization in wood-decay fungi // BMC Evolutionary Biology. 2018. V. 18. 119 p.
Lopes de Gerenyu V.O., Anichkin A.E., Avilov V.K., Kuznetsov A.N., Kurganova I.N. Termites as a factor of spatial differentiation of CO2 fluxes from the soils of monsoon tropical forests in southern Vietnam. Eurasian Soil Science. 2015. V. 48. № 2. P. 208–217.
Magnússon R.I., Tietema A., Cornelissen J.H.C., Hefting M.M., Kalbitz K. Tamm Review: Sequestration of carbon from coarse woody debris in forest soils // Forest Ecology and Management. 2016. V. 377. P. 1–15.
Meriem S., Tjitrosoedirjo S., Kotowska M.M., Hertel D., Triadiati T. Carbon and nitrogen stocks in dead wood of tropical lowland forests as dependent on wood decay stages and land-use intensity. Annals of Forest Research. 2016. V. 59. № 2. P. 299–310.
Moroni M.T., Morris D.M., Shaw C., Stokland J.N., Harmon M.E., Fenton N.J., Merganicová K., Merganic J., Okabe K., Hagemann U. Buried wood: a common yet poorly documented form of deadwood // Ecosystems. 2015. V. 18. P. 605–628.
Palace M., Keller M., Hurtt G., Frolking S. A review of above ground necromass in tropical forests. Tropical forests. 2012. P. 215– 252.
Pfeifer M., Lefebvre V., Turner E., Cusack J., Khoo M., Chey V.K., Peni M., Ewers R.M. Deadwood biomass: an underestimated carbon stock in degraded tropical forests? Environmental Research Letters. 2015. V. 10. № 4. P. 044 019.
R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, Austria, 2013.
Russell M.B., Fraver S., Aakala T., Gove J.H., Woodall C.W., D’Amato A.W., Ducey M.J. Quantifying carbon stores and decomposition in dead wood: A review // Forest Ecology and Management. 2015. V. 350. P. 107–128.
Shorohova E., Kapitsa E., Kuznetsov A., Kuznetsova S., Lopes de Gerenuy V., Kaganov V., Kurganova I. Coarse woody debris density and carbon concentration by decay classes in mixed montane wet tropical forests // Biotropica. 2022 (в пeчaти).
Shorohova E., Kapitsa E. The decomposition rate of non-stem components of coarse woody debris (CWD) in European boreal forests mainly depends on site moisture and tree species // European Journal of Forest Research. 2016. V. 135. P. 593–606.
Shorohova E., Kapitsa E., Kazartsev I., Romashkin I., Polevoj A., Kushnevskaya H. Tree species traits are the predominant control on the decomposition rate of tree log bark in a mesic old-growth boreal forest // Forest Ecology and Management. 2016. V. 377. P. 36–45.
Shorohova E., Kapitsa E., Kuznetsov A., Kuznetsova S., Lopes de Gerenuy V., Kaganov V., Kurganova I. Decay classes of coarse woody debris in a lowland Dipterocarp forest: implications for volume, density, and carbon estimates // Biotropica. 2021. V. 53. № 3. P. 879–887.
Ståhl G., Ringvall A., Fridman J. Assessment of coarse woody debris – a methodological overview // Ecological Bulletins. 2001. V. 49. P. 57–70.
Stokland J.N., Siitonen J., Jonsson B.G. Biodiversity in dead wood // Cambrigde Univ. Press. 2012. 509 p.
WRB. World Reference Base for Soil Resources 2014, Update 2015. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps. World Soil Resources Reports. 2014. V. 106. Rome: FAO.
Yoneda T., Tamin R., Ogino K. Dynamics of aboveground big woody organs in a foothill dipterocarp forest, West Sumatra, Indonesia. Ecological Research. 1990. V. 5. № 1. P. 111–130.
Дополнительные материалы отсутствуют.