

Лесоведение, 2022, № 4, стр. 364-380
Влияние гидротермических условий на подстилочных беспозвоночных вырубок и гарей Хибин
И. В. Зенкова a, *, И. М. Штабровская a
a Институт проблем промышленной экологии Севера ФИЦ “Кольский НЦ РАН”
184209 Апатиты, мкр. Академгородок, д. 14а, Россия
* E-mail: i.zenkova@ksc.ru
Поступила в редакцию 01.11.2021
После доработки 07.12.2021
Принята к публикации 02.02.2022
- EDN: EZRORH
- DOI: 10.31857/S0024114822030123
Аннотация
С 2018 г., после присвоения Хибинам статуса национального парка, возросла актуальность задач сохранения уникальных природных объектов и эталонных комплексов хибинской горной тундры и северной тайги, экологического мониторинга и восстановления территорий, нарушенных хозяйственной деятельностью горнодобывающих компаний. Одним из последствий такой деятельности, нанесшей значительный экологический ущерб уязвимым экосистемам заполярного горного массива, стал обширный низовой пожар на месте брошенной нерасчищенной промышленной вырубки в долине реки Кунийок. С целью анализа восстановительных сукцессий почвенного покрова и населяющей его фауны в изменившихся условиях инсоляции и гидротермического режима нами был организован почвенно-экологический мониторинг горно-соснового леса и трех участков его антропогенной трансформации – горелого сосняка, вырубки и горелой вырубки. Спустя 2 (2015 г.) и 5–6 лет (2018 г.) после воздействия на всех модельных участках выполнены учеты беспозвоночных (почвенными ловушками с формалином), температуры и влажности подстилки на глубине 5 см (автономными термогидрохронами). Выявлено устойчивое повышение влажности подстилки при нарушении (горелый лес) или сведении (вырубка) древесного яруса, что соответствует представлениям о развитии процессов поверхностного гидроморфизма и заболачивания на лесных вырубках и гарях. Учтены беспозвоночные 34 таксономических групп при повышенных показателях их разнообразия и динамической плотности в более теплый вегетационный сезон 2018 г. Наибольшим числом таксонов характеризовались гари; наименьшим, но с максимальной плотностью беспозвоночных – вырубка; минимальной динамической плотностью – дважды нарушенная горелая вырубка. Обе вырубки отличались более выраженными межвидовыми связями с участием паразитических перепончатокрылых и жуков-некрофагов. В паре соседних лесных биотопов “контрольный–горелый сосняк” прослежено усиление различий в структуре населения беспозвоночных через 5 лет после обширного низового пожара по сравнению с обследованием этих участков через 2 г. Установлена прямая корреляционная зависимость межгодовых различий плотности герпетобионтов от температуры лесной подстилки (r ≥ 0.71) и обратная – от ее влажности (–0.79 ≤ r ≤ –0.82). Факторами, определяющими различия в плотности беспозвоночных между годами и участками, оказались: сумма температур (r = 0.95), минимальные температуры июля (0.89) и максимальные температуры сентября (0.94) и августа (0.92). Подтверждена важность лесной подстилки для сохранения на горно-лесных вырубках и гарях исходного комплекса фауны и поддержания гидротермического режима, благоприятного для ее восстановления. Сделан методический вывод о большем фаунистическом сходстве участков, удаленных на расстоянии 1 км, но сохранивших лесную подстилку (контрольный сосняк и вырубка), по сравнению с парами участков, заложенных в 300 м друг от друга для их сопоставления по принципу “горелый – незатронутый пожаром” (т.е. контрольный и горелый сосняки, горелая и негорелая вырубки).
История промышленного освоения Хибин в Мурманской области насчитывает почти 90 лет, и все это время воздействию со стороны горнодобывающих и перерабатывающих предприятий была подвержена преимущественно юго-восточная часть массива. Одно из последних планируемых к освоению месторождений апатит-нефелиновой руды “Партомчорр” находится на севере Хибин – в долине реки Кунийок. Летом 2012 г. здесь была организована промышленная рубка леса на площади более 100 га (Новости-24 …, 2012). Из-за конфликта интересов промышленников и экологов, выступавших против строительства обогатительной фабрики и прокладки коммуникаций в долине в связи с организацией национального парка “Хибины”, работы по рубке леса и очистке территории от порубочных остатков были прекращены. Через год на неочищенной вырубке возник обширный пожар, который охватил горно-таежный лес в долине и лесной пояс на склоне горы Путеличорр. Выгоревшая площадь составила 8 га. Преградой для распространения пожара стала проходящая по долине грунтовая дорога, которая разделила территорию на сгоревшую (выгоревшие сосняк и вырубка) и не затронутую пожаром (исходный сосняк и его вырубка) (рис. 1). Таким образом, в непосредственной близости друг от друга возникли четыре участка, выбранные нами в качестве модельных для проведения экологического мониторинга за сукцессионными изменениями нарушенного почвенного покрова и населяющей его фауны.
Рис. 1.
Модельные участки в долине реки Кунийок в Хибинах: (а) – ненарушенный сосняк (контроль), (б) – горелый сосняк, (в, г) – нерасчищенная вырубка у подножья горы Партомчорр, (д, е) – горелая вырубка у подножья горы Путеличорр.

Первичное почвенно-зоологическое обследование лесных биотопов – исходного (контрольного) и горелого сосняков мы провели в 2015 г., через 2 г. после обширного низового пожара (Зенкова, 2016). В 2018 г. объем работ расширили и охватили вырубки – сгоревшую и негорелую. На всех участках оценивали таксономическое разнообразие и обилие почвенных беспозвоночных в образцах подстилки и герпетобионтной фауны, населяющей поверхность подстилки и ярус напочвенной растительности (методом почвенных ловушек с формалином). Результаты исследования почвенной фауны показали, что, несмотря на разнонаправленные тренды изменения численности отдельных таксонов, общая численность педобионтов через 5–6 лет после всех видов антропогенного воздействия была достоверно ниже (в 1.8–3.5 раза) по сравнению с контрольным сосняком при незначимых различиях этого показателя между нарушенными участками (Зенкова и др., 2020). В настоящем сообщении приводится анализ населения герпетобионтных беспозвоночных и гидротермических показателей почв на модельных участках, спустя 6 лет после вырубки леса и 2 и 5 лет после пожара.
ОБЪЕКТЫ И МЕТОДИКА
Пары лесных участков “контрольный сосняк (С)–горелый сосняк (ГС)” и вырубок “горелая (ГВ)–незатронутая пожаром (В)” были выбраны на плоском ландшафте в межгорной долине реки Кунийок на севере Хибин, 67°50ʹ с.ш., 33°39ʹ в.д., на высотах 220–235 м над ур. м. (Зенкова и др., 2020). Расстояние между парами составляло 1 км, а между участками в каждой паре – 300 м: по 150 м от разделяющей их грунтовой дороги.
Контрольный сосняк (рис. 1а) характеризовался мощной подстилкой (9.5 ± 1.3 см) и хорошо развитым гетерогенным ярусом из кустарничков, мхов и лишайников. Древесный ярус сформирован лапландской формой сосны обыкновенной (Pinus sylvestris f. lapponica), елью (Picea obovata) и березой извилистой (Betula tortuosa); кустарничковый – вороникой (Empetrum nigrum), черникой (Vaccinium myrtillus), брусникой (V. vitis-idaea), голубикой (V. uliginosum), багульником (Ledum palustre), мхами рода Bryophyta и лишайниками рода Cladonia.
В горелом сосняке (рис. 1б) подрост ели и березы, напочвенная растительность и валежник выгорели. Лесная подстилка протлела. Высота нагара на стволах сосен достигала 2–3 м, их большая часть лишилась крон. Через два года поверхность почвы имела темную окраску, территория очагово зарастала вегетативно возобновляющейся черникой, что характерно для послепожарного восстановления травяно-кустарничкового яруса в хвойных лесах Кольского п-ова (Баккал и др., 2006). Спустя пять лет зола с поверхности почвы заметно вымылась, покров из черники и злаков стал более сомкнутым, во влажных микропонижениях росла брусника, встречался иван-чай и редкий подрост березы высотой до 30–40 см.
На 6-летней вырубке (рис. 1в, 1г), сохранившей гетерогенный ярус из кустарничков, мхов и лишайников, шло активное зарастание березой; неочищенная территория завалена стволами и ветками, труднопроходима; подстилка мощная, темноокрашенная, с признаками оторфовывания в нижней части.
Горелая вырубка, испытавшая воздействие два года подряд, и через 6 лет выглядела наиболее нарушенной (рис. 1д, 1е): рубка уничтожила древесный ярус, а пожар – кустарничковый ярус и подстилку, обнажив минеральную массу почвы. Между поваленными обугленными стволами сосен встречались куртины злаков, отдельные растения иван-чая и пятна печеночных мхов; слабый подрост березы отмечался в основном вдоль обочины дороги.
На всех участках в 2015 и 2018 гг., с июля до середины сентября (всего 75 суток), отлавливали беспозвоночных пластиковыми стаканами-ловушками (500 мл) с формалином, вкопанными в почву до верхней кромки. Повторность на каждом участке была 30-кратной: 3 линии (l–lll) по 10 ловушек с расстоянием между линиями и ловушками по 10 м, таким образом, площадь облова составила по 0.18 га. В лабораторных условиях содержимое ловушек разбирали вручную, число особей пересчитывали в экз. на 100 ловушко-суток (показатель динамической плотности). Общее число лов.-сут. равнялось 2250, число беспозвоночных, учтенных в двух сосняках в 2015 г. – 4227 экз., отловленных на 4 участках в 2018 г., – 19776 экз.
Автоматическими термогидрохронами ТРВ-2 (Регистраторы …, https://gigrotermon.ru) каждые 2 ч регистрировали температуру (Т, °С) и влажность (V, %) почвенного воздуха на глубине 5 см. Данная глубина оказалась максимальной во фрагментах подстилки на горелой вырубке с эродированным почвенным покровом. Вычисляли среднесуточные, среднемесячные и средние за 75 сут значения. Число показаний Т и V составило на каждом участке по 1800, общий объем измерений – 10 800.
Достоверность различий средних значений измеренных показателей подстилки и населения беспозвоночных между участками оценивали по t-крит. Стьюдента, различий в динамике Т и V – методом ANOVA с F-крит. Фишера. Зависимость плотности беспозвоночных от эдафических факторов определяли методом корреляционного анализа. Сходство-различие участков по таксономическому составу и динамической плотности беспозвоночных выявляли методом кластерного анализа. Результаты обрабатывали в программах Exсel, Statistica-12. Структуру доминирования характеризовали по соотношению доминантов, субдоминантов, малочисленных и редких беспозвоночных с соответствующими долями плотности: ≥10, <10, <5 и <1% (Southwood, 1978).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Гидротермические показатели почв
Влажность воздуха в 5 см слое подстилки контрольного С не опускалась ниже 97% в прохладный и дождливый вегетационный сезон 2015 г. и 76% в более теплый 2018 г. На нарушенных участках ГС и В с сохранившейся подстилкой влажность постоянно была близка к 100% (CV < 1%), что свидетельствовало об изменении водного режима почвы в направлении гидроморфизма (рис. 2а). На В это подтверждалось оторфовыванием нижней части мощной подстилки. В ГС достоверное повышение V подстилки по сравнению с соседним контрольным С отмечалось уже через 2 г. после пожара. Лишенная напочвенной растительности и органогенного горизонта ГВ отличалась наибольшей сезонной вариабельностью влажности (CV = 32%) за счет как минимальных (V = = 30%, в погожие дни “бабьего лета” 8–9/IХ), так и максимальных среди участков (до 118%, 22/VII) среднесуточных значений.
Рис. 2.
Гидротермические показатели подстилки на глубине 5 см на модельных участках в июле–сентябре 2015 и 2018 гг.: (а), (б) – динамика влажности и температуры, (в) – среднемесячные температуры. С–15, ГС–15 – контрольный и горелый сосняки, обследованные в 2015 г.
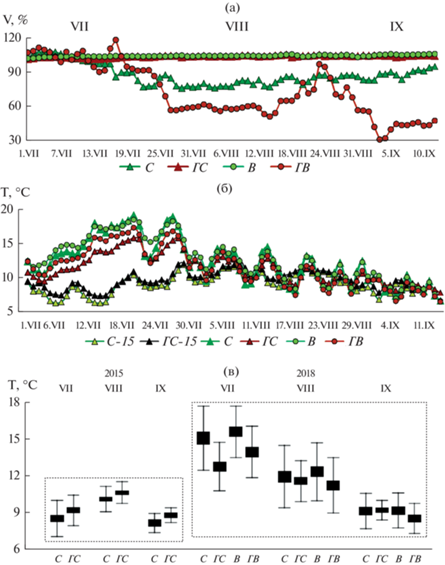
Причиной устойчивого повышения влажности сохранившейся подстилки нарушенных ГС и В можно считать уничтожение древесного яруса. Кроны деревьев, перехватывая атмосферные осадки, снижают их поступление в почву, а корни выполняют мощную водороегулирующую функцию в почве. Значительная часть почвенной влаги расходуется растениями древесного и кустарникового яруса на транспирацию (Карпечко, 2008, 2016, Архипова, Исаченкова, 2013). На фоне увеличения поступления осадков на поверхность нарушенных территорий происходит сдвиг расхода почвенной влаги с физиологического процесса ее транспирации растениями на физический процесс испарения с поверхности почвы (Краснощеков, 1994; Евдокименко, 1996; Поляков, 2010; Тарасов и др., 2011). В Хибинах, в условиях высокой влажности горного климата (Яковлев, 1961), непродолжительного периода с положительными температурами воздуха и расположения исследуемых участков в межгорной речной долине, принимающей водные стоки с окружающих склонов, расход почвенной влаги на испарение представляется нам несущественным в сравнении с ее приходом. Следовательно, дисбаланс этих процессов может быть реальной причиной гидроморфизма почв на нарушенных участках – ГС и В.
Заболачиванию почв после сведения лесов в разных широтных районах посвящена обширная литература (Морозова, 2004; Исаченкова, Герасимова, 2007; Архипова, Исаченкова, 2013). В средней тайге переувлажнение подзолистых почв в первые 5–10 лет послерубочной сукцессии проявляется даже на автоморфных позициях ландшафта (Лаптева и др., 2015). Изменение гидрологического режима почв по пути заболачивания считается основным последствием сплошных рубок на начальных стадиях восстановления и в горных районах (Дмитриев, 1950; Морозова, 2004; Мерзляков, 2008; Дымов, Милановский, 2014; Дымов, 2017). Развитию процесса поверхностного гидроморфизма способствует увеличение мощности и влагозапасов снежного покрова на вырубках по сравнению с хвойными лесами, полог которых задерживает до 30–40% снеговых осадков (Лебедев, 1982). На вырубках горно-таежных лесов запасы воды в снеге на 20–30 мм (или 15–20%) выше, чем под пологом леса (Буренина и др., 2013; Онучин и др., 2014). В условиях повышенной влажности горного климата и возросшего после сведения леса поступления атмосферных осадков на вырубках активно развивается моховой покров, который, имея высокую влагоемкость, снижает интенсивность газообмена между приземным слоем воздуха и почвой, ухудшая ее аэрацию и создавая дополнительные условия для поверхностного переувлажнения (Дымов и др., 2012; Лопатовская, 2018).
В ГС, наряду с протлевшим слоем мха, аккумулирующим влагу, ее удерживанию в лесной подстилке и верхних минеральных горизонтах способствует пирогенный уголь. Обладая высокопористой структурой, уголь способен длительное время сорбировать большое количество влаги на единицу массы (Брянин и др., 2019). Влагонасыщение горелых лесных подстилок неоднократно объяснялось их пирогенным уплотнением (Киселева, 1978; Краснощеков, Сорокин, 1988; Бескоровайная и др., 2005; Тарасов и др., 2008; Назаркина, 2009; Тер-Мисакянц, 2013; Дымов и др., 2015). Сгорание рыхлой подстилки и образование тяжелых компонентов – сажи, частиц угля и золы уменьшает порозность почвы и изменяет ее водно-физические свойства. Продукты сгорания заполняют почвенные поры, снижают фильтрационные характеристики и водоотдачу почвы, повышают ее влагозапасы. Дополнительно водоотталкивающие свойства почве придают гидрофобные ароматические соединения – органические продукты пиролиза, которые образуются в таежных лесах при сгорании лигнинов и смолистых компонентов древесины хвойных пород (Цибарт, Геннадиев, 2011).
Прогрев подстилки в лесных биотопах в 2015 и 2018 гг. носил противоположный характер. Через 2 г. после пожара темноокрашенная подстилка ГС прогревалась достоверно сильнее по сравнению с соседним контрольным С (рис. 2б, 2в). Средняя за сезон Т на глубине 5 см составила +9.8 ± 0.2°С против +9.2 ± 0.2°С; максимальная, в первой декаде июля, +12.6°С против +12.0°С; сумма температур за 75 сут была на 43°С выше: 685 против 642°С. Минимальная за сезон среднесуточная Т также была выше в горелой подстилке: +7.4°С против +6.4°С в С. Активные среднесуточные Т ≥ ≥ +10°C регистрировались в подстилке ГС на протяжении 30 из 75 сут (или в 43% случаев), а в контрольном С – только 21 день (30%). Дни с эффективными Т ≥ +5°C (но < +10°C), напротив, преобладали в подстилке С: 49 сут против 40 в ГС или в 70% суток против 57%. Динамика Т в подстилке контрольного С была достоверно более вариабельна, чем в ГС (F = 7.0, р = 0.009) и, несмотря на наличие развитого мохово-лишайниково-кустарничкового яруса, в большей степени зависела от Т атмосферного воздуха (коэф. корр. r = 0.83 для С и 0.80 для ГС).
В более теплый вегетационный сезон 2018 г. температура подстилки была достоверно выше на участках с напочвенной растительностью – в контрольном сосняке и на вырубке. Средняя за июль–сентябрь Т на глубине 5 см составила на этих участках +12.4 и +12.7°С соответственно, а на горелых не превысила +11.5°С (рис. 2б). Сумма температур за 75 сут равнялась 954°С на В и 927°С в С, а на гарях оказалась на 90–100°С меньше, чем на В, и на 60–70°С ниже, чем в контрольном С. Число дней с активными среднесуточными Т ≥ ≥ +10°C увеличивалось в ряду: ГВ (48 сут)–ГС (54)– С (55)–В (59), т.е. для лесных биотопов было сопоставимо, в отличие от 2015 г.
В 2018 г. самым теплым месяцем, достоверно дифференцирующим прогрев участков, был июль (F ≥ 8.0, р ≤ 0.007). С первых чисел августа различия между участками в величине и динамике среднесуточных Т нивелировались, и с 9/IХ, а на ГВ раньше – с 27/VIII, температура на глубине 5 см на всех участках была устойчиво ниже +10°С (рис. 2б, 2в). В более холодный вегетационный сезон 2015 г. подстилка в лесных биотопах максимально прогрелась только к августу, но до меньших Т, чем в августе 2018 г.
Население беспозвоночных животных
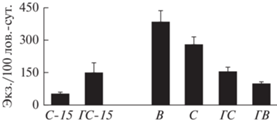
С июля по сентябрь 2015 г. в контрольном и горелом сосняках отловлены представители 28 таксономических групп при сопоставимом числе таксонов и трехкратном преобладании их общей плотности в ГС. В 2018 г. на 4-х модельных участках учтено 34 таксона при наибольшем разнообразии в горелом сосняке, наименьшем разнообразии, но максимальной плотности на вырубке и минимальной динамической плотности на дважды нарушенной горелой вырубке (табл. 1, рис. 3).
Таблица 1.
Население беспозвоночных на модельных участках в 2015 и 2018 гг. (в порядке убывания относительной плотности в ловушках контрольного сосняка, %)
| Таксономическая группа | 2015 | 2018 | |||||
|---|---|---|---|---|---|---|---|
| С–15 | ГС–15 | С | ГС | В | ГВ | ||
| Муравьи (Formicidae) | ++++ | ++++ | ++++ | ++++ | ++++ | ++ | |
| Пауки (Aranei) | ++++ | ++ | ++++ | ++++ | ++++ | ++++ | |
| Долгоносики (Curculionidae) | +++ | ++ | ++ | + | + | + | |
| Перепончатокрылые (Hymenoptera)* | ++ | + | ++ | ++ | + | +++ | |
| Стафилиниды (Staphylinidae) | ++ | + | ++ | ++ | + | ++ | |
| Жужелицы (Carabidae) | ++ | ++ | + | +++ | + | ++++ | |
| Цикадовые (Cicadoidea) | ++ | + | + | + | ++ | ++++ | |
| Многоножки (Lithobiidae) | ++ | + | + | ++ | + | ++ | |
| Мягкотелки (Cantharidae) | ++ | + | + | + | + | + | |
| Клещи-краснотелки (Trombidiidae) | ++ | – | + | + | – | + | |
| Клопы (Heteroptera) | + | + | ++ | + | + | ++ | |
| Двукрылые (Diptera) | + | + | + | + | + | ++ | |
| Щелкуны (Elateridae) | + | + | + | + | + | + | |
| Совки (Noctuidae) | + | + | + | + | + | + | |
| Coleoptera sp. | + | + | + | ++ | + | – | |
| Моллюски (Gastropoda) | + | + | + | + | + | – | |
| Короеды (Scolytidae) | + | + | + | + | – | – | |
| Усачи (Cerambycidae) | + | + | + | – | + | + | |
| Пилюльщики (Вyrrhidae) | + | + | ̶ | + | ̶ | ++ | |
| Чернотелки (Tenebrionidae) | + | – | ++ | ++ | + | +++ | |
| Пилильщики (Tenthredinoidea) | + | – | + | ++ | + | + | |
| Шмели (Bombidae) | + | – | + | – | + | – | |
| Гладкотелки (Leiodidae) | + | – | + | + | – | + | |
| Листоблошки (Psyllidoidеа) | + | – | ̶ | + | – | + | |
| Тли (Aphidoidea) | – | + | + | + | + | + | |
| Дождевые черви (Lumbricidae) | – | ̶ | + | + | + | + | |
| Сеноеды (Psocoptera) | – | ̶ | + | + | + | – | |
| Сенокосцы (Opiliones) | – | ++ | + | ++++ | – | + | |
| Могильщики (Nicrophorus) | – | ̶ | + | + | – | + | |
| Пяденицы (Geometridae) | – | ̶ | + | – | – | – | |
| Тараканы (Blattodea) | – | + | – | + | + | + | |
| Божьи коровки (Coccinellidae) | – | + | – | ̶ | – | + | |
| Энхитреиды (Enchytraeidae) | – | – | – | + | – | + | |
| Листоеды (Chrysomelidae) | – | – | – | + | – | – | |
| Личинки Insecta | – | – | – | – | + | + | |
| Трипсы (Thysanoptera) | – | – | – | – | – | + | |
| Всего: | Таксонов | 24 | 22 | 28 | 30 | 24 | 29 |
| экз./100 лов.–сут. | 50 | 149 | 283 | 153 | 383 | 105 | |
Рис. 3.
Динамическая плотность беспозвоночных на модельных участках через 2 (С-15, ГС-15) и 5–6 лет (В, С, ГС, ГВ) после антропогенного воздействия.
Для сравнения, в образцах подстилки, отобранных в день установки почвенных ловушек в начале июля, выявлено 20 таксонов почвенной фауны (от 10 до 15 на модельных участках), общими из которых были лишь 6 таксонов, доминирующих в подстилке северотаежных лесов Мурманской области: активно подвижные хищные пауки, многоножки-костянки, муравьи, жуки мягкотелки, стафилиниды и сапрофильные личинки жуков-щелкунов (Зенкова и др., 2016, 2020).
Следует отметить, что расположение модельных участков в северной части Хибин и освобождение ведущей к ним грунтовой дороги от снежников не ранее начала июля не позволило организовать полевые работы в мае–июне, в период размножения и наибольшей активности беспозвоночных, что, вероятно, привело к недоучету их реального разнообразия и обилия.
Лесные биотопы
В контрольном сосняке в оба вегетационных сезона в ловушках преобладали муравьи, пауки и жуки долгоносики с суммарной долей 82–89%. Многочисленными были типичные для лесной подстилки хищные краснотелковые клещи и жесткокрылые (жужелицы, стафилиниды, мягкотелки); малочисленными – многоножки-костянки и жуки чернотелки, растительноядные клопы, цикадки и слизни; единичными – большинство насекомых-фитофагов: шмели, гусеницы чешуекрылых, ложногусеницы пилильщиков, щелкуны, короеды и пилюльщики, личинки жуков-усачей (табл. 1).
В более теплый сезон 2018 г. число таксонов увеличилось с 24 до 28, в том числе за счет насекомых–фитофагов: сеноедов, тлей, гусениц пядениц. Уловистость представителей 9 таксонов возросла в 2–13 раз. Плотность подстилочных хищников (многоножек, жужелиц, мягкотелок, клещей-краснотелок) и растительноядных моллюсков, цикадок, гусениц совок и жуков-усачей сохранилась на уровне 2015 г. Наряду с муравьями, пауками и долгоносиками массовыми в ловушках (более чем по 100 экз.) были стафилиниды, клопы и разнородная группа перепончатокрылых, однако возросшее обилие муравьев (74% всех беспозвоночных против 59% в 2015 г.) определило более олигодоминантную структуру населения в 2018 г. (табл. 1). Пространственное распределение беспозвоночных в С, более равномерное, чем в ГС и на В (рис. 4), также зависело от распределения муравьев (r = 0.99). В целом у половины из 28 таксонов, выявленных в контрольном сосняке в 2018 г., динамическая плотность оказалась максимальной по сравнению с нарушенными участками (табл. 2).
Рис. 4.
Схема пространственного распределения беспозвоночных на модельных участках в 2018 г. (общая плотность, экз./100 лов.-сут.). 1–10 – почвенные ловушки, I–III – ловушко–линии (в направлении от грунтовой дороги вглубь участков), в скобках – коэф. пространственного варьирования плотности, %.
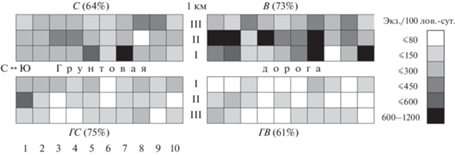
Таблица 2.
Плотность беспозвоночных на модельных участках в 2018 г. (экз./100 лов.–сут.)
| Таксон | С | В | ГС | ГВ | ||||
|---|---|---|---|---|---|---|---|---|
| М ± m | min–max | М ± m | min–max | М ± m | min–max | М ± m | min–max | |
| Formicidae | 210 ± 33bdc | 38–920 | 320 ± 51acd | 0–1141 | 58 ± 20d | 3–527 | 8 ± 0.4b | 0–9 |
| Aranei | 31 ± 2.4c | 16–77 | 38 ± 6.2c | 3–168 | 17 ± 2.9 | 0–74 | 47 ± 0.8ac | 7–217 |
| Curculionidae | 9.0 ± 0.8bcd | 3–23 | 1.2 ± 0.3acd | 0–4 | 0.2 ± 0.1 | 0–1 | 0.2 ± 0.1 | 0–2 |
| Staphylinidae | 6.2 ± 0.1bd | 0–21 | 1.5 ± 0,5a | 0–14 | 6.1 ± 2,2b | 0–63 | 2.2 ± 0.9 | 0–23 |
| Hymenoptera | 5.7 ± 1.0b | 0–27 | 1.0 ± 0,2a | 0–4 | 4.3 ± 0.9b | 0–21 | 2.7 ± 1.1b | 0–8 |
| Heteroptera | 4.2 ± 0.9bcd | 0–17 | 2.5 ± 0.8acd | 0–17 | 0.7 ± 0.2 | 0–4 | 1.8 ± 0.4c | 0–10 |
| Carabidae | 2.3 ± 0.6b | 0–17 | 0.7 ± 0.2a | 0–4 | 11 ± 1.4ab | 3–28 | 12.7 ± 2bd | 0–36 |
| Tenthredinoidea | 0.9 ± 0.2bd | 0–2 | 0.4 ± 0.2ad | 0–6 | 2.3 ± 0.5abd | 0–9 | ед. | 0–1 |
| Cantharidae | 0.9 ± 0.2bcd | 0–2 | ед. | 0–2 | 0.5 | 0–1 | ед. | 0–1 |
| Trombidiidae | 0.9 ± 0.4bd | 0–12 | – | – | 0.5 ± 0.2bd | 0–4 | ед. | 0–2 |
| Lithobiidae | 0.5 ± 0.1 | 0–2 | 0.9 ± 0.3a | 0–6 | 3.0 ± 0.5abd | 0–8 | 1.7 ± 0.4b | 0–5 |
| Aphidoidea | 0.5 ± 0.2bcd | 0–2 | ед. | 0–1 | 0.2 ± 0.1 | 0–2 | 0.2 ± 0.1 | 0–2 |
| Lepidoptera | 0,4 ± 0,1bcd | 0–2 | ед. | 0–1 | 0.9 ± 0.2bd | 0–5 | ед. | 0–1 |
| Gastropoda | 0.4 ± 0.1d | 0–2 | 0.6 ± 0.2ad | 0–4 | 0.3 ± 0.2ad | 0–4 | – | – |
| Psocoptera | 0.4 ± 0.2d | 0–4 | ед. | 0–1 | ед. | 0–1 | – | – |
| Nicrophorinae | 0.2 ± 0.1 | 0–4 | – | – | 1.3 ± 0.9ad | 0–23 | 0.5 ± 0.3d | 0–6 |
| Cicadoidea | ед. | 0–2 | 13 ± 2.3ac | 0–44 | 1.4 ± 0.4a | 0–6 | 15 ± 1.6ac | 0–31 |
| Diptera | ед. | 0–1 | ед. | 0–1 | 0.8 ± 0.3 | 0–6 | 1.7 ± 1.5 | 0–44 |
| Cerambycidae | ед. | 0–1 | 0.2 ± 0.1 | 0–2 | – | – | 0.2 ± 0.1 | 0–1 |
| Bombidae | ед. | 0–1 | 0.7 ± 0.3a | 0–6 | – | – | – | – |
| Lumbricidae | ед. | 0–1 | ед. | 0 –1 | ед. | 0–1 | ед. | 0–1 |
| Elateridae | ед. | 0–1 | ед. | 0–1 | ед. | 0–1 | ед. | 0–1 |
| Scolytidae | 0.3 | 0–2 | – | – | 0.6 ± 0.3 | 0–8 | – | – |
| Opiliones | ед. | 0–1 | – | – | 35 ± 4.4abd | 4.1–82 | ед. | 0–1 |
| Blattodea | – | – | 1.0 ± 0.3acd | 0–6 | ед. | 0–1 | ед. | 0–2 |
| Psyllidae | – | – | – | – | ед. | 0–1 | ед. | 0–1 |
| Сhrysomielidae | – | – | – | – | ед. | 0–1 | – | – |
| Enchytraeidae | – | – | – | – | 0.2 ± 0.1 | 0–2 | ед. | 0–1 |
| Byrrhidae | – | – | – | – | 0.4 ± 0.2 | 0–4 | 1.0 ± 0.3c | 0–5 |
| Coccinellida | – | – | – | – | – | – | 0.8 ± 0.2 | 0–2 |
| Thysanoptera | – | – | – | – | – | – | ед. | 0–1 |
| экз./100 лов.-сут. | 283 ± 33bс | 81–973 | 383 ± 52сd | 77–1212 | 153 ± 22d | 22–568 | 105 ± 11 | 29–283 |
В горелом сосняке через 2 г. после низового пожара обитали беспозвоночные 22 таксонов. Общими с контрольным участком, удаленным на 300 м, были 18 групп (табл. 1). Из них у представителей 8-ми таксонов динамическая плотность была ниже контрольных величин (пауки, мягкотелки, перепончатокрылые, короеды, цикадки, гусеницы совок, моллюски, личинки двукрылых), у 7-ми таксонов – выше (многоножки, муравьи, жужелицы, стафилиниды, усачи, долгоносики, клопы); у растительноядных щелкунов и пилюльщиков – сопоставима с контролем. В ловушках ГС не оказалось жуков гладкотелок и чернотелок, клещей-краснотелок и фитофагов – пилильщиков, шмелей и листоблошек. Напротив, появились обитатели открытых пространств – сенокосцы, тараканы и божьи коровки, не выявленные в контрольном С. В целом из 28 таксонов, отловленных на двух лесных участках в 2015 г., представители 14 таксонов (или 50%) имели большую плотность в контрольном С, представители 11 таксонов (39%) – в ГС (табл. 3). Пятикратно возросшее в сравнении с контролем обилие муравьев (северного лесного Formica aquilonia) сделало их абсолютными доминантами в ГС: 89% от общей плотности беспозвоночных. Заселение муравьями сосняков, нарушенных низовым пожаром, и активное формирование муравьиных семей в течение 3-х первых лет развития гарей было прослежено в Окском заповеднике и объяснялось высокой численностью насекомых-ксилофагов и слабым задернением почвы (Аверина, 2002).
Таблица 3.
Структурные показатели населения беспозвоночных в лесных биотопах через 2 и 5 лет после пожара
| Число таксономических групп | 2015 г. | 2018 г. |
|---|---|---|
| В контрольном сосняке С | 24 | 28 |
| В горелом сосняке ГС | 22 | 30 |
| Суммарно в С и ГС | 28 | 34 |
| Общих для С и ГС, из них: | 18 (или 64%) | 25 (или 74%) |
| С большей плотностью в С | 8 (44%) | 10 (40%) |
| С большей плотностью в ГС | 7 (39%) | 11 (44%) |
| Со сходной плотностью в С и ГС | 3 (17%) | 4 (16%) |
| Выявленных только в С | 6 | 3 |
| Выявленных только в ГС | 4 | 5 |
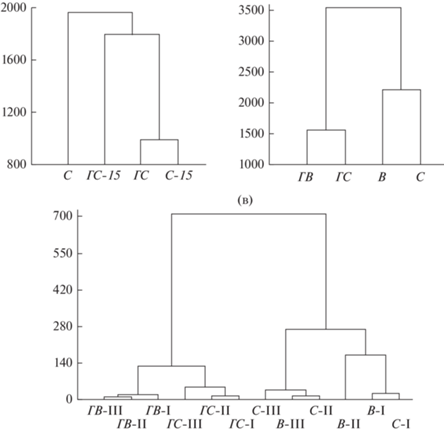
Через 5 лет после пожара структура населения беспозвоночных в ГС претерпела значительные изменения как по отношению к контрольному С, так и в сравнении с 2015 г., что отразилось на дендрограмме кластеризации этих участков (рис. 5а). В более теплый вегетационный сезон 2018 г. в обоих лесных биотопах, С и ГС, увеличилось число таксонов в ловушках (с 22–24 до 28–30) и возросла динамическая плотность большинства таксонов: у муравьев (на порядок), долгоносиков и клопов более выражено – в контрольном С; у большинства фитофагов (цикадки, гусеницы чешуекрылых, ложногусеницы пилильщиков, пилюльщики, короеды) и личинок двукрылых – в ГС, превысив контрольные показатели; у прочих перепончатокрылых, пауков, стафилинид, чернотелок и тлей – в обоих лесных биотопах. Сравнялась с контролем плотность стафилинид, гладкотелок и слизней. В обоих лесных биотопах появились сапрофильные дождевые черви, сеноеды и жуки-могильщики. Плотность хищных пауков, клещей-краснотелок, перепончатокрылых и мягкотелок в ГС оставалась ниже контрольных величин в оба года, а у муравьев, клопов, щелкунов, долгоносиков и усачей стала уступать контрольному С в 2018 г. Доля муравьев снизилась до 38%, и они делили доминирование с сенокосцами (23%) и пауками (11%). Таксонами, дифференцирующими лесные биотопы в оба года, оказались шмели, отмеченные только в ловушках контрольного С, и тараканы, выявленные только в ГС.
Рис. 5.
Кластеризация модельных участков по таксономическому составу и динамической плотности беспозвоночных: (а) – контрольный и горелый сосняки в 2015 и 2018 гг. (single linkage method, Euclidean distances), (б) – лесные участки и вырубки в 2018 г. (средние для каждого участка показатели по 30-ти ловушкам, Ward`s method), (в) – средние значения по ловушко-линиям.
В результате разнонаправленных изменений обилия таксонов общая динамическая плотность беспозвоночных достоверно, в 5.7 раза, увеличилась только в контрольном С (табл. 1, рис. 3). Для сравнения в почвенных пробах общая численность фауны двукратно возросла в обоих лесных биотопах, однако все доминирующие таксоны оставались менее обильными в ГС, в итоге трехкратная разница в общей численности, выявленная между С и ГС в 2015 г., сохранилась и в 2018 г. (Зенкова и др., 2020).
В ряде публикаций, анализирующих начальные стадии послепожарных сукцессий, отмечается повышенная численность насекомых-вредителей. На разновозрастных гарях Печоро-Илычского заповедника в Республике Коми это были долгоносики (Hylobius abietis), привлекаемые ослабленными соснами (Савельева, Долгин, 2008, 2009), в сосняках Окского заповедника, в зависимости от их спелости, – жуки-пилюльщики, щелкуны или листоеды вида Adoxus obscurus, трофически связанного с иван-чаем, разрастающимся на гарях в первые годы после пожаров (Потапова, 2002). В сосновых лесах соседних регионов – центральной Швеции и Республики Карелия – в первые два года после пожаров на зарастающих гарях более многочисленными, чем в негорелых лесах, были сосущие фитофаги – тли, цикадовые, трипсы. Комплекс хищных беспозвоночных из-за недостатка пищи восстанавливался медленно: через 6 лет после пожара их биомасса составляла лишь 40–60% от контрольных показателей (Gongalsky, Persson, 2013).
В Хибинах в горелом сосняке выраженным увеличением плотности через два года после пожара отличались активно подвижные хищники, привлекаемые разреженным и лучше прогреваемым горелым лесом (сенокосцы, жужелицы) и его более теплой и влажной подстилкой (многоножки-костянки, стафилиниды), а также разрушители древесины – муравьи и долгоносики. Изменение общей плотности герпетобионтов по годам и участкам (трехкратное преобладание в ГС через 2 г. после пожара и почти двукратное отставание от контрольного С через 5 лет) также было скоррелировано преимущественно с динамикой подстилочных хищников – муравьев (r = 0.92), пауков (0.82), перепончатокрылых (0.78), стафилинид (0.74) и фитофагов долгоносиков (0.63). Четкую тенденцию к увеличению численности в ГС через два и особенно через 5 лет после пожара проявили три группы зоофагов – многоножки, сенокосцы и жужелицы. Для многоножек, чувствительных к кислотности почвы, дополнительным благоприятным фактором могла быть повышенная зольность протлевшей подстилки.
Стволовые вредители – усачи и короеды, способные заселять ослабленные, усыхающие и мертвые деревья, а также привлекаемые территориями с большим количеством поваленных стволов, в оба года были единичны в ловушках как контрольного, так и горелого сосняков. По наблюдениям энтомологов, в Восточной Фенноскандии, несмотря на наличие старовозрастных лесов, частые пожары и отсутствие должного лесопатологического ухода, вспышек массового размножения этих ксилофагов не зафиксировано (Яковлев, 1996; Yakovlev et al., 2000).
Для лесных участков выявлена прямая зависимость межгодовых различий плотности герпетобионтов от температуры подстилки (r ≥ 0.71) и обратная – от ее влажности (–0.79 ≤ r ≤ –0.82). Показателями, достоверно определяющими различия в плотности беспозвоночных между годами и участками, оказались: сумма температур июля (r = 0.95), Тmax сентября (0.94) и августа (0.92) и Тmin июля (0.89), которые в вегетационный сезон 2018 г. были выше в подстилке контрольного С (рис. 2б, 2в). Корреляция плотности беспозвоночных с температурой и запасами влаги в подстилке была выявлена при сравнении нарушенных участков с разными древесными породами в процессе лесовосстановления в южной тайге Центральной Сибири (Безкоровайная, Егунова, 2011).
Вырубки
Структура населения беспозвоночных шестилетней вырубки отражала два сукцессионных процесса: смену лесной фауны на луговую и усиление гидроморфизма почвы. Несмотря на сохранение мощной лесной подстилки, плотность типичных подстилочных хищников – стафилинид, жужелиц, чернотелок, личинок мягкотелок и перепончатокрылых была в 3–10 раз ниже, чем в контрольном С. Как и в ГС, исчезли краснотелковые клещи, появились божьи коровки. Плотность влаголюбивых муравьев, пауков, многоножек, дождевых червей, моллюсков и личинок двукрылых, напротив, в 1.2–3 раза превысила таковую в С, что соответствовало высокой влажности подстилки на В при сходной степени прогрева этих участков (рис. 2), а также обилию пищевых ресурсов для сапрофильных беспозвоночных и ксилофагов в виде многочисленных поваленных стволов и веток деревьев.
Несмотря на активный подрост березы, в комплексе фитофагов В, в отличие от контрольного С, не отмечены гусеницы пядениц, единичны гусеницы совок, в 1.7–11 раз сократилась плотность клопов, тлей, сеноедов и насекомых, трофически связанных с древесиной и корнями живых сосен – жуков-короедов, долгоносиков и ложногусениц пилильщиков. Уловистость личинок жуков-усачей, развитие которых проходит в мертвой древесине, напротив, увеличилась в 4 раза, как и луговых фитофагов: цикадок (на 2 порядка), влаголюбивых тараканов, опылителей – шмелей. Тараканы и шмели исчислялись в ловушках десятками особей, цикадки – сотнями. В целом 4 группы беспозвоночных (муравьи, моллюски, шмели и тараканы) достигали на В наибольшей плотности среди участков. Этот список дополняли личинки усачей и цикадки с оговоркой, что сходную плотность они имели и на второй вырубке – горелой. В результате при наименьшем числе таксонов (24) лесная вырубка отличалась самой высокой динамической плотностью беспозвоночных (рис. 3, табл. 1, 2) и их наиболее неравномерным пространственным распределением из-за гетерогенности среды обитания в виде скопления–разброса порубочного материала (рис. 4). Максимальные локальные скопления формировали муравьи, пауки и цикадки.
В сравнении с контрольным С на В плотность более половины таксонов (13 из 24) снизилась, у 9 таксонов – увеличилась; представители 6 таксонов не были выявлены, как и в ГС появились тараканы. Из-за обилия муравьев структура населения была более олигодоминантной: муравьи составляли 86% улова (против 75% в контрольном С), пауки – 10, цикадки – 3, суммарная доля представителей остальных таксонов не превысила 1% (табл. 1).
Структура населения дважды нарушенной горелой вырубки наиболее отличалась от контрольного С и была сходна частично с ГС, частично – с соседней В. Динамическая плотность беспозвоночных была здесь достоверно меньшей: в 1.5 раза в сравнении с ГС и в 3–4 раза в сравнении участками С и В, сохранившими подстилку и кустарничковый ярус (табл. 1, 2, рис. 3). Обилие муравьев, преобладающих на остальных участках, сократилось на два порядка; плотность типичных обитателей лесной подстилки – стафилинид и мягкотелок – на порядок; моллюски и сеноеды отсутствовали.
В то же время представители 6 таксонов достигали на ГВ наибольшей плотности: обитатели открытых пространств – божьи коровки и активные хищники – пауки и жужелицы, предпочитающие в Хибинах открытые ландшафты лесотундры и горной тундры, и жуки-чернотелки. Последние проявили себя как некрофаги: вместе с жуками-мертвоедами массово шли в ловушки с попавшими в них мышами–полевками, также привлекаемыми горелой вырубкой со сложным лабиринтом из поваленных стволов деревьев. Еще двумя группами с максимальной уловистостью на ГВ были цикадки (собрано 333 экз.) и бриофаги пилюльщики (два десятка имаго), развивающиеся на редких пятнах мохового покрова. Наряду с цикадками среди фитофагов преобладали клопы, присутствовали тараканы и развивающиеся в стволах мертвых деревьев личинки жуков-усачей. Как и на В, комплекс фитофагов ГВ имел луговой облик, а растительноядные группы, типичные для контрольного С, были здесь единичны (щелкуны, долгоносики, гусеницы совок, тли, трипсы) или отсутствовали (листоеды, короеды, гусеницы пядениц, ложногусеницы пилильщиков). Слабо зарастающая ГВ также не привлекала шмелей, обычных на соседней В. Уловистость многовидовой группы перепончатокрылых была высокой, но уступала контрольному С, а плотность многоножек – ГС (табл. 2).
Из-за снижения обилия муравьев – основного доминанта в уловах других участков, только ГВ отличалась полидоминантной структурой населения беспозвоночных и их более равномерным пространственным распределением (рис. 4). Преобладали пауки – 47, цикадки – 16, жужелицы – 13, чернотелки и перепончатокрылые – 5–6%. Доля представителей каждого из остальных 22-х таксонов не превысила 2% (табл. 1).
На примере обеих вырубок, как антропогенно нарушенных горных биотопов, впервые за 10-летний период почвенно-зоологических исследований Хибин, нами установлен факт массового заражения сосущих фитофагов – цикадок их специализированными паразитоидами – мелкими жалящими осами-дриинидами рода Gonatopus (Hymenoptera, Aculeata, Dryinidae). Биология представителей небольшого семейства дриинид изучена недостаточно (Gugliemino, 2002), но известно, что бескрылые самки захватывают и временно парализуют личинок цикадок для откладки яиц в их тельца, что нарушает естественный метаморфоз цикадок. По мере развития паразитической личинки на теле хозяина разрастается шаровидная капсула. Большинство цикадок, отловленных на вырубках Хибин, имели такие капсулы различной локализации, что свидетельствовало о массовом заражении их популяции; попадались в почвенные ловушки и сами осы-паразиты.
Обобщая данные по таксономическому составу и обилию беспозвоночных в оба периода исследований, можно сделать методический вывод. Из 4-х модельных участков, изначально выбранных для парного сравнения по принципу “горелый – незатронутый пожаром” (С–ГС и В–ГВ), наиболее сходными по разнообразию, плотности и структуре доминирования населения беспозвоночных–герпетобионтов, оказались не эти пары участков, заложенные в 300 м друг от друга, а контрольный сосняк (С) и вырубка (В), удаленные на расстоянии 1 км, но имеющие лесную подстилку и развитый мохово-кустарничковый ярус. Аналогичный вывод был сделан нами по результатам сравнительных исследований почвенной фауны в образцах подстилки с этих модельных участков (Зенкова и др., 2020).
Высокая плотность в контрольном С и на В основного доминанта – муравьев (210–320 против 8–58 экз./100 лов.-сут. на гарях), сходная плотность пауков (30–40 экз./100 лов.-сут.), клопов и присутствие сеноедов и шмелей повлияли на обособление этих участков с лесной подстилкой в подкластер, противопоставленный подкластеру горелых участков – ГС и ГВ (рис. 5б). Горелые сосняк и вырубка отличались от негорелых участков присутствием энхитреид, пилюльщиков, листоблошек и божьих коровок, повышенной плотностью жужелиц (11–13 против 1–2 экз./100 лов.-сут.), многоножек, сенокосцев, личинок двукрылых. Еще отчетливее обособление горелых участков и, напротив, сходство контрольного сосняка и вырубки, проявилось при кластеризации данных по ловушко-линиям (рис. 5в).
Многочисленность жужелиц (вида Cicindela sylvatica) на начальных стадиях послепожарной сукцессии отмечена на разновозрастных гарях в Республике Коми (Савельева, Долгин, 2008, 2009). Повышенные показатели уловистости и разнообразия этих жуков по сравнению с ненарушенным лесом и выборочной рубкой выявлены после экспериментального выжигания вырубки в юго-восточной Норвегии (Gongalsky et al., 2006). Нами в горелом сосняке и на горелой вырубке за полевые сезоны 2015 и 2018 гг. учтено 19 видов жужелиц (тогда как за 10 лет исследования природных биотопов Хибин суммарно – 30 видов), из них 8 видов, принадлежащих родам Amara, Bembidion, Dicheirotrichus, Harpalus, Pterostichus, отловлены только на этих нарушенных участках (Zenkova, Filippov, 2019).
ЗАКЛЮЧЕНИЕ
В результате обширной промышленной рубки в 2012 г. и последующего низового пожара в 2013 г. в речной долине на севере Хибин образовались три варианта антропогенной трансформации горно-таежного леса (горелый лес, вырубка и горелая вырубка), послужившие модельными биотопами для мониторинга восстановительных сукцессий почвенного покрова и населяющей его фауны в условиях заполярного горного массива. Во всех биотопах герпетобионтные беспозвоночные, учтенные почвенными ловушками, оказались более разнообразным и многочисленным компонентом по сравнению с фауной в образцах лесной подстилки, исследованной в эти же сезоны (Зенкова и др., 2020). Структура их населения через 5–6 лет после воздействия соответствовала степени трансформации горных биотопов, которые можно расположить в следующем порядке: лесная вырубка (сохранение лесной подстилки и мохово-кустарничкового яруса, активное возобновление березы) – горелый сосняк (мертвый древостой, протлевшая подстилка, зарастание черникой) – горелая вырубка (полное уничтожение лесного фитоценоза и лесной подстилки, эродированная до минеральной массы почва, фрагментарное зарастание территории). Учитывая близкое расположение модельных участков, позволяющее беспозвоночным активно мигрировать на их территории, очевидно, что различия в разнообразии и динамической плотности герпетобионтов определялись наличием-отсутствием лесной подстилки и ее гидротермическими свойствами, что подтверждено результатами корреляционного и кластерного анализов. На вырубке, несмотря на формирование лугового комплекса беспозвоночных в первые годы появления березового подроста, наличие подстилки и яруса напочвенной растительности имеет первостепенное значение для поддержания гидротермического режима, благоприятного для сохранения и восстановления горно-лесного комплекса беспозвоночных даже после полного уничтожения древесного полога.
Список литературы
Аверина И.А. Динамика численности и размещение на гари Окского заповедника муравьев рода Formica // Мониторинг сообществ на гарях и управление пожарами в заповедниках. М.: ВНИИ природы, 2002. С. 66–69.
Архипова М.В., Исаченкова Л.Б. Динамика почвенно-растительного покрова экосистемы сосново-еловых лесов (на примере юго-западного Подмосковья) // Геоэкология. Инженерная геология. Гидрогеология. Геокриология. 2013. № 6. С. 491–501.
Баккал И.Ю., Горшков В.В., Пазуха А.В. Послепожарное восстановление напочвенного покрова еловых лесов Кольского полуострова // Бюлл. Ботанич. сада Саратовского гос. ун-та. 2006. В. 5. С. 74–77.
Безкоровайная И.Н., Егунова М.Н. Формирование комплексов почвенных беспозвоночных в процессе лесовосстановления (южная тайга, Центральная Сибирь) // Ульяновский мед.-биол. журн. 2011. № 4. С. 117–129.
Безкоровайная И.Н., Иванова Г.А., Тарасов П.А., Сорокин Н.Д., Богородская А.В., Иванов В.А., Конард С.Г., Макрае Д.Дж. Пирогенная трансформация почв сосняков средней тайги Красноярского края // Сиб. экол. журн. 2005. Т. 12. № 1. С. 143–152.
Брянин С.В., Суслопарова Е.С., Молчанов Д.А., Абрамова Е.Р. Постпирогенный уголь как фактор почвообразования в таёжных лесах Дальнего Востока // Лесные почвы и функционирование лесных экосистем: матер. VII Всерос. научн. конф. с межд. уч. М.: ЦЭПЛ РАН, 2019. С. 98–101.
Буренина Т.А., Шишикин А.С., Онучин А.А., Борисов А.Н. Снежный покров на вырубках разных лет в пихтово-кедровых лесах Енисейского кряжа // Лесоведение. 2013. № 6. С. 26–36.
Дмитриев А.С. Заболачивание и разболачивание концентрированных вырубок в борах- черничниках в бассейне Сысолы (Коми АССР). Автореф. дис. … канд. с-х. наук. Сыктывкар, 1950. 16 с.
Дымов А.А., Габов Д.Н., Дубровский Ю.А., Жангуров Е.В., Низовцев Н.А. Влияние пожара в северотаежном ельнике на органическое вещество почвы // Лесоведение. 2015. № 1. С. 52–62.
Дымов А.А. Влияние сплошных рубок в бореальных лесах России на почвы (обзор) // Почвоведение. 2017. № 7. С. 787–798. https://doi.org/10.7868/S0032180X17070024
Дымов А.А., Лаптева Е.М., Милановский Е.Ю. Изменение почв и почвенного органического вещества в процессе естественного лесовозобновления после рубки сосняка бруснично-зеленомошного // Вестн. Московского гос. ун-та леса (Лесной вестник). М. 2012. Т. 2. № 85. С. 67–71.
Дымов А.А., Милановский Е.Ю. Изменение органического вещества таежных почв в процессе естественного лесовозобновления растительности после рубок (средняя тайга Республики Коми) // Почвоведение. 2014. № 1. С. 39–47. https://doi.org/10.7868/S0032180X14010043
Евдокименко М.Д. Послепожарная динамика микроклимата и гидротермического режима мерзлотных почв в лиственничниках Станового хребта // Сибирский экол. журн. 1996. № 1. С. 73–79.
Зенкова И.В. Почва и почвенная фауна пирогенного леса: последствия низового пожара в Хибинском горном массиве // Почвоведение – продовольственной и экологической безопасности страны: тез. докл. VII съезда Об-ва почвоведов им. В.В. Докучаева. М.: Белгород, 2016. В 2-х ч. Ч. I. С. 221–222. https://doi.org/10.13140/RG.2.2.29090.63683
Зенкова И.В., Штабровская И.М., Усова Д.В. Почвенная фауна вырубок и гарей Хибин // Вестник МГТУ. 2020. Т. 23. № 2. С. 160–172. https://doi.org/10.21443/1560-9278-2020-23-2-160-172
Исаченкова Л.Б., Герасимова М.И. Пространственно-временная организация почвенного покрова антропогенно-измененных лесов юго-западного Подмосковья // Пространственно-временная организация почвенного покрова, теоретические и прикладные аспекты: матер. Междунар. науч. конф. СПб. 2007. С. 434–437.
Карпечко А.Ю. Изменение плотности и корненасыщенности почв под влиянием лесозаготовительной техники в еловых лесах Южной Карелии // Лесоведение. 2008. № 5. С. 66–70.
Карпечко А.Ю. Влияние рубок на сток с лесопокрытой части водосбора Онежского озера // Труды КарНЦ РАН. Лимнология и океанология. 2016. № 5. С.13–20.
Киселева Г.А. Изменение водно-физических свойств почв в результате послепожарного заболачивания // Изменение почвенного покрова Дальнего Востока в результате сельскохозяйственного использования и мелиорации. Уссурийск: ПримСХИ. 1978. В. 52. С. 92–94.
Краснощеков Ю.Н. Влияние пожаров на свойства горных дерново-таежных почв лиственничников Монголии // Почвоведение. 1994. № 9. С. 102–109.
Краснощеков Ю.Н., Сорокин Н.Д. Почвенно-экологические изменения на вырубках и гарях Восточного Хэнтэя (МНР) // Почвоведение. 1988. № 1. С. 117–127.
Лаптева Е.М., Втюрин Г.М., Бобкова К.С., Каверин Д.А., Дымов А.А., Симонов Г.А. Изменение почв и почвенного покрова еловых лесов после сплошнолесосечных рубок // Сибирский лесн. журн. 2015. № 5. С. 64–76. https://doi.org/10.15372/SJFS20150505
Лебедев А.В. Гидрологическая роль горных лесов Сибири. Новосибирск: Наука, 1982. 182 с.
Лопатовская О.Г., Максимова Е.Н., Попов В.В., Соловьева К.С. Влияние сплошных рубок на лесорастительные свойства почв Братского района Иркутской области // Изв. Байкальского гос. ун-та. Экология. 2018. С. 159–165.
Мерзляков О.Э. Влияние сплошных рубок на изменение лесорастительных свойств мерзлотных грубогумусных почв // Вестн. Том. гос. ун-та. Биология. 2008. № 1(2). С. 71–81.
Морозова Р.М. Влияние концентрированных рубок еловых лесов на свойства почв // Антропогенная трансформация таежных экосистем Европы. Петрозаводск. 2004. С. 182–184.
Назаркина А.В. Изменение плотности сложения и влагозапасов лесных почв долины р. Амур в результате действия пожаров // Вестн. КрасГАУ. 2009. № 4. С. 37–41.
Новости-24 от 29.08.12: В Мурманской области в Хибинах вырублено около 100 га леса // Комсомольская правда–Мурманск. URL: https://www.murmansk.kp.ru/ online/news/1234173/ (Дата обращения: 01.10.2021)
Поляков В.Л. Подпочвенное увлажнение несвязных несуффозионных грунтов в условиях интенсивного испарения // Прикладна гiдромеханiка. 2010. Т. 12. № 3. С. 54–61.
Потапова Н.А. Почвенные беспозвоночные (мезофауна) – 20 лет наблюдений в Окском заповеднике // Мониторинг сообществ на гарях и управление пожарами в заповедниках. М.: ВНИИприроды, 2002. С. 57–65.
Онучин А.А., Буренина Т.А., Зирюкина Н.В., Фарбер С.К. Лесогидрологические последствия рубок в условиях Средней Сибири // Сиб. лесн. журн. 2014. № 1. С. 110–118.
Регистраторы (логгеры) температуры и влажности / ООО “Инженерные Технологии” (Челябинск). URL: https://gigrotermon.ru (Дата обращения: 15.09.2021)
Савельева Л.Ю., Долгин М.М. Структура населения жесткокрылых (Insecta: Coleoptera) разновозрастных сосновых гарей подзоны средней тайги Республики Коми // Вестн. Поморск. ун-та. Сер. естеств. и точн. наук. 2008. № 1(13). С. 40–44.
Савельева Л.Ю., Долгин М.М. Изменение структуры населения жесткокрылых (Insecta: Coleoptera) сосновых гарей подзоны средней тайги Республики Коми // Изв. Самарск. НЦ РАН. 2009. Т. 11. С. 656–660.
Тарасов П.А., Иванов В.А., Иванова Г.А. Особенности температурного режима почв в сосняках средней тайги, пройденных низовыми пожарами // Хвойн. бореал. зоны. 2008. Т. 25. № 3–4. С. 300–304.
Тарасов П.А., Иванов В.А., Иванова Г.А., Краснощекова Е.Н. Постпирогенные изменения гидротермических параметров почв среднетаежных сосняков // Почвоведение. 2011. № 7. С. 795–803.
Тер-Мисакянц Т.А. Изменение биологических свойств дерново-карбонатных почв Кавказа после вырубки леса // Изв. вузов. Северо-Кавказ. регион. Естественные науки. 2013. № 3. С. 57–59.
Цибарт А.С., Геннадиев А.Н. Ассоциации полициклических ароматических углеводородов в пройденных пожарами почвах // Вестн. Моск. ун-та. Сер. 5. География. 2011. № 3. С. 13–19.
Яковлев Б.А. Климат Мурманской области. Мурманск: Книжн. изд-во, 1961. 200 с.
Яковлев Е.Б. К характеристике комплексов ксилофитных жеcткокрылых (Coleoptera) в лесах Карелии, не подвергавшихся лесоводственному уходу // Проблемы антропогенной трансформации лесных биогеоценозов Карелии. Петрозаводск. 1996. С. 139–166.
Gongalsky K., Midtgaard F., Overgaard H.J. Effects of prescribed forest burning on carabid beetles (Coleoptera: Carabidae): A case study in south-eastern Norway // Entomol. Fennica. 2006. V. 17. № 3. P. 325–333.
Gongalsky K.B., Persson T. Recovery of soil macrofauna after wildfires in boreal forests // Soil Biol. & Biochem. 2013. V. 57. P. 182–191. https://doi.org/10.1016/j.soilbio.2012.07.005
Gugliemino A. Dryinidae (Hymenoptera Chrysidoidea): an interesting group among the natural enemies of the Auchenorrhyncha (Hemiptera) // Denisia. 2002. V. 4. P. 549–556. URL: https://www.zobodat.at/pdf/DENISIA_0004_0549-0556.pdf
Southwood T.R.E. Ecological methods. London. Champan and Hall, 1978. 575 p.
Zenkova I.V., Filippov B.Yu. The ground beetles fauna (Coleoptera, Carabidae) of natural and anthropogenic habitats in Khibiny polar massif. // The Proceed. of 8th Intern. Symp. of Ecol. (ISEM8). Pešić V. (ed.). Budva, Montenegro, 2019. P. 121–129.
Yakovlev E.B., Scherbakov A.N., Polevoi A.V., Humala A.E. Insect fauna of the Paanajarvi National Park and proposed Kalevala National Park with particular emphasis on saproxylic Coleoptera, Diptera and Hymenoptera // Biodiversity of old-growth forests and its conservation in north-west Russia. Oulu, 2000. P. 103–158.
Дополнительные материалы отсутствуют.