

Лесоведение, 2022, № 5, стр. 504-519
Динамика размерной и возрастной структуры древостоев коренных сосняков Северного Предуралья
И. Н. Кутявин a, *, А. В. Манов a
a Институт биологии Коми НЦ УрО РАН
167982 Сыктывкар, ул. Коммунистическая, д. 28, Россия
* E-mail: kutjavin-ivan@rambler.ru
Поступила в редакцию 25.03.2021
После доработки 12.07.2021
Принята к публикации 02.02.2022
- EDN: ERCOJQ
- DOI: 10.31857/S0024114822040064
Аннотация
Изучена десятилетняя динамика структурной организации древостоев коренных сосняков на постоянных пробных площадях, располагающихся на территории Печоро-Илычского заповедника (Северное Предуралье). Исследования строения проводились в спонтанно развивающихся древостоях среднетаежных сосняков брусничных и черничных типов на автоморфных почвах. Древостои в них характеризуются разновозрастной структурой. Состав и возраст древостоев регулируется частотой и интенсивностью пожаров. Наблюдениями с 2007 по 2019 гг. выявлена достаточно высокая лабильность в строении древостоев. С пополнением их деревьями нового поколения в результате изменения морфометрических признаков – возраста, диаметра и высоты – в сосняках брусничных происходит увеличение вариации этих показателей, тогда как с гибелью материнских деревьев в сосняках черничных их вариабельность снижается. В пределах отдельных поколений в древостоях сосняков изменения размерных характеристик деревьев остаются примерно на одном уровне. Показано, что с отпадом угнетенных деревьев старших поколений происходит увеличение высоты среднего дерева этого поколения. Приведена модель связи варьирования возраста деревьев и диаметра их стволов в сосняках за десятилетний период. Результаты модели наглядно свидетельствуют об изменении возрастной структуры сосновых древостоев. Установлено, что старовозрастные сосняки, несмотря на активное пополнение состава молодыми особями древесных растений, характеризуются относительно стабильным объемом стволовой древесины. Показано, что при длительном отсутствии пирогенной активности в сосняках происходит формирование древостоев из разновозрастных поколений деревьев сосны, ели и кедра. Пожары слабой интенсивности способствуют скачку возобновления темнохвойных видов, которые в последующем активно внедряются в состав древостоев.
Изучение динамики структурной организации древостоев направлено на решение проблем мониторинга и моделирования развития лесных экосистем и ведения устойчивого управления лесами в конкретном регионе (Kurz et al., 2009; Березовская, Карев, 2015; Müller et al., 2015; Демаков, 2018; и др.). В последние десятилетия в связи с изменением климата отмечается увеличение катастрофических явлений (ветровалы, пожары, вспышки энтомовредителей), приводящих к трансформации структуры лесов (Проскуряков, 2015; Швиденко и др., 2017; Жила и др., 2019; Kulha et al., 2020). Следовательно, важна оценка изменений структуры фитоценозов лесных экосистем под воздействием различных экзогенных факторов (Мишко и др., 2018). Пройденные пожаром или ветровалом сосняки на автоморфных почвах по сравнению с другими хвойными формациями в большинстве случаев нарушаются частично и сукцессионные смены в них выражены слабо (Листов, 1986; Кутявин, 2018; Жила с соавт., 2019; и др.). С изменением климата происходит интенсивная экспансия верхних границ леса в горы, тогда как нижние предгорные массивы становятся более продуктивными и устойчивыми (Kullman, 2005; Sumichrast et al., 2020). Однако резкие климатические скачки, как показали наблюдения 2009–2010 гг., снижают устойчивость и продуктивность лесов (Bastos et al., 2014).
При определении влияния климатических условий на динамические процессы лесных фитоценозов существует ряд подходов: использование базы данных лесоустройства (Kurz et al., 2009; Henttonen et al., 2017); применение космических снимков разных лет (Kulha et al., 2020); описание возрастных рядов древостоев, развивающихся на различных этапах их формирования (Нагимов и др., 2010). К этим методам добавляется рубка всех деревьев с последующим их обмером с целью детального анализа роста древостоя по поколениям (Верхунов, 1976; Зябченко, 1984); наиболее точный метод – проведение многолетних исследований на лесных стационарах с закладкой постоянных пробных площадей (ППП) (Рысин и др., 1981; Манов, 2017; Sumichrast et al., 2020; и др.).
Исследования структуры и продуктивности сосновых лесов Европейского Северо-Востока России достаточно освещены в научных работах (Осипов, Бобкова, 2016; Кутявин, 2018; Манов, Кутявин, 2019). Материалы, характеризующие динамику строения древостоев сосняков и ельников в условиях средней тайги Республики Коми, единичны (Осипов, Бобкова, 2016; Манов, 2017). Следовательно, накопление знаний пространственно-временных изменений структуры древостоев, эдификаторов сосновых экосистем, в процессе их развития представляет большой научный интерес.
Цель работы – оценить динамику состава, строения и возрастной структуры старовозрастных древостоев брусничных и черничных сосняков Северного Приуралья.
ОБЪЕКТЫ И МЕТОДИКА
Исследования проведены в коренных сосняках на территории Печоро-Илычского биосферного заповедника и его буферной зоны Комсомольского лесничества, Верхне-Печорского участкового лесничества (61°44″40′ с.ш., 57°06″25′ в.д. и 61°56″54′ с.ш., 57°55″53′ в.д.). В фитоценозах сосняков брусничных и черничных типов предгорной и равнинной части Северного Приуралья, согласно общим нормативным указаниям и лесоводственным методикам, заложена серия постоянных пробных площадей (ППП). В сосняках брусничных на ППП 1 исследования выполнены в 2007 и 2018 гг., на ППП 12 – в 2012 и 2019 гг. В черничных типах сосняков на ППП 4 наблюдения проведены в 2009 и 2019 гг., на ППП 10 – в 2011 и 2019 гг. Первичная характеристика сосновых сообществ приведена в работе И.Н. Кутявина (2018). За период развития сосновые сообщества пройдены низовыми пожарами различной интенсивности от двух до пяти раз более 100 лет назад. В составе древостоев при господстве сосны обыкновенной (Pinus sylvestris L.) присутствуют: ель сибирская (Picea obovata Ledeb.), сосна сибирская, или кедр (Pinus sibirica Du Tour), пихта сибирская (Abies sibirica Ledeb.), береза повислая и пушистая (Betula pendula Roth, Betula pubescens Ehrh.). В составе подроста редко встречается тополь дрожащий, или осина (Populus tremula L.) (табл. 1). Сосняки произрастают на пирогенных иллювиально-железистых и пирогенных иллювиально-гумусово-железистых песчаных подзолах.
Таблица 1.
Характеристика таксационных показателей древостоев сосняков и подроста
| Год учета | Состав | Элемент леса (ярус) | Густота, шт. га–1 | Запас, м3 га–1 | Абсолютная полнота, м2 га–1 |
Подрост | |||
|---|---|---|---|---|---|---|---|---|---|
| живой | сухой | живой | сухой | состав | густота тыс. экз. га–1 | ||||
| Сосняк брусничный ППП 1 | |||||||||
| 2007 | 10C | С | 170 | 60 | 210.0 | 20.8 | 21 | 8С2Е + Ос, ед. Б | 4.2 |
| 2018 | 10C ед. Е | С | 528 | 68 | 211.8 | 26.0 | 21.3 | 9С1Е ед. К, Б, Ос | 2.58 |
| Е | 8 | – | 0.02 | – | |||||
| Итого | 536 | 68 | 211.8 | 26.0 | |||||
| Сосняк брусничный ППП 12 | |||||||||
| 2012 | 9C1Е + К, Б | С | 317 | 37 | 177.6 | 11.4 | 24.9 | 6К2Е2С + Б | 1.1 |
| Е | 253 | 10 | 26.3 | 1 | |||||
| К | 63 | – | 7 | – | |||||
| Б | 30 | – | 5.3 | – | |||||
| Итого | 663 | 47 | 216.2 | 12.4 | |||||
| 2019 | 8С2Е + К, Б | С | 307 | 57 | 184.6 | 15.2 | 26.0 | 4К4С2Е + Б ед. Пх | 7.2 |
| Е | 393 | 23 | 34.2 | 1.9 | |||||
| К | 107 | – | 10.6 | – | |||||
| Б | 30 | – | 6.3 | – | |||||
| Итого | 837 | 80 | 235.7 | 17.1 | |||||
| Сосняк черничный ППП 4 | |||||||||
| 2009 | 9C1Е + Б, ед. К, Пх | С | 243 | 28 | 315.1 | 38.6 | 32.9 | 5Е2Б2К1С + Пх | 0.9 |
| Е | 198 | 23 | 37.2 | 0.8 | |||||
| Б | 53 | – | 12 | – | |||||
| К | 20 | – | 1.6 | – | |||||
| Итого | 514 | 28 | 365.9 | 38.6 | |||||
| 2019 | 8С2Е + Б ед.К, Пх | С | 223 | 45 | 296.5 | 66.4 | 29.7 | 6К2Е2Б + С, Пх | 2.2 |
| Е | 573 | 58* | 42.5 | 7.2* | |||||
| Б | 118 | 2 | 15.4 | – | |||||
| К | 28 | – | 2.2 | – | |||||
| Пх | 10 | – | 1.1 | – | |||||
| Итого | 952 | 105 | 357.7 | 66.4 | |||||
| Сосняк черничный ППП 10 | |||||||||
| 2011 | 10C | С (I) | 473 | 120 | 262.2 | 113.9 | 24.7 | 9Е1К + С ед. Б | 1.9 |
| 8С1Е1Б | С (II) | 80 | 7.3 | 3.9 | |||||
| Е (II) | 40 | – | 1.4 | – | |||||
| Б (II) | 10 | – | 0.8 | – | |||||
| Итого | 603 | 120 | 271.7 | 113.9 | 28.6 | ||||
| 2019 | 10С | С (I) | 500 | 161* | 261.2 | 131.2* | 21.5 | 5Е4К1С ед. Б | 1.5 |
| 7Е3Б | Е (II) | 70 | – | 2.2 | – | 1.2 | |||
| Б (II) | 13 | 2 | 0.8 | 0.0 | |||||
| Итого | 583 | 163 | 264.2 | 131.2 | 22.7 | ||||
Сосняк брусничный (ППП 1) располагается на предпойменной боровой террасе р. Печора. Обследование 2018 г. показало, что за десять лет с начала наблюдений в данном сосняке произошли значительные изменения в строении древостоя. В его составе появляется поколение деревьев сосны в возрасте 35–75 лет, перешедших из подроста. Обследование 2007 г. показало, что крупный подрост сосны в фитоценозе из-за сильной вытянутости стволов часто гибнет от снеголома. За прошедшие десять лет в составе древостоя сосняка отмечено появление единичных особей ели в возрасте 50–60 лет. С переходом крупного подроста в древостой и его частичным выпадением из состава количество подроста под пологом снижается с 4.2 до 2.6 тыс. экз. га–1. Увеличивается количество мелких (до 0.5 м) и средних (0.51–1.5 м) по высоте особей сосны. В составе подроста появляется кедр (табл. 1).
Коренной сосняк брусничный (ППП 12) развивается на флювиогляциальном повышении вдоль верхового болота. В составе подроста за десятилетний период увеличивается число деревьев преимущественно темнохвойных видов. Сосняк черничный (ППП 4), развивающийся на предпойменной боровой террасе р. Печора, по составу древостоя, как и на ППП 12, активно пополняется еловым элементом (табл. 1). Сосняк черничный (ППП 10) расположен между боровой террасой р. Печора и старым руслом реки (старицей). При первой и повторной инвентаризации ценоза отмечено незначительное увеличение количества подроста. Как и в сосняке черничном (ППП 4), размерные характеристики подроста сосны на ППП 10 сохраняются почти на одном уровне. За прошедшие восемь лет с начала наблюдений выявлен резкий скачок возобновления кедра с 10 до 40%. Подрост ели оставляет прочные количественные позиции (табл. 1).
В 2018–2019 гг. с целью получения более точных биометрических показателей деревьев использовалось современное таксационное оборудование Haglof. При сплошной перечислительной таксации на ППП к древостою отнесены деревья, имеющие на высоте 1.3 м диаметр ≥6 см, к подросту – деревья с диаметром <6 см. У каждого дерева в древостое проводилось измерение диаметра на высоте 1.3 м. Для оценки возрастной структуры древостоев у более половины деревьев на ППП отбирались керны. Пожарная история древостоев установлена согласно дендрохронологическим методам с использованием кернов и древесных спилов с живой и мертвой древесины, имеющей пожарные подсушины (Madany et al., 1982). Датировку древесных образцов и пожарной активности в сосняках проводили с использованием прибора Lintab и компьютерных программ Tsap – Win (Rinn, 1996) и COFECHA (Holmes, 1983).
Обработку измерений таксационных показателей древостоев сосняков, полученных на ППП, проводили с использованием нормативов (Войнов и др., 2012) и компьютерных программ, предназначенных для статистической обработки и построения моделей регрессионного анализа. Оценка степени дифференциации биометрических показателей деревьев проведена по математико-статистическим методикам (Гусев, 2002). Классификация возрастной структуры древостоев дана по Г.Е. Комину и И.В. Семечкину (1970), которые предлагают выделять шесть типов возрастной структуры: абсолютно-одновозрастные, одновозрастные, условно-разновозрастные, ступенчато-разновозрастные, циклично-разновозрастные и абсолютно-разновозрастные. Также дополнен еще один относительно разновозрастный тип возрастного строения, находящегося на стадии перестройки или демутации возраста, выделенного С.А. Дыренковым (1984).
При оценке типов возрастной структуры применяли распределение числа деревьев на ППП с выделением в них поколений. За период формирования поколения принимали временной отрезок или “скачок” от начала накопления деревьев в древостое до его затухания, возникающего при определенных сложившихся условиях под пологом леса вследствие воздействия как экзо- (естественное изреживание), так и эндогенных (пожары) факторов. Такое выделение поколений в древостое наиболее приемлемо с точки зрения оценки возрастных смен (Гавриков, 1983). Так, по мнению данного автора, “поколение – это группа деревьев, реализовавшая общую экологическую нишу” (с. 22). Дополнительно нами приняты за поколения единичные старые материнские деревья, некогда занимавшие основное пространство древостоя. Хотя на данный момент они не оказывают значительного влияния на структуру древостоя, однако показывают его историю развития в пространственно-временном интервале.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Пожарная история исследуемых сосняков. За период развития сосняк брусничный (ППП 1) был подвержен пяти низовым пожарам, пик которых приходился на XVIII в. Последний раз рассматриваемое сообщество подвергалось пирогенному воздействию 130 лет назад (1887 г.). При этом пожары с 1805 по 1728 гг. не вызывали разрывов в распределении деревьев по возрасту в первом поколении, а наоборот способствовали скачкам возобновления сосны (рис. 1а). Древостой на ППП 12 за период его развития был подвержен пирогенному воздействию три раза: в 1911, 1906 и 1869 гг. По пожарным подсушинам с сухостойных деревьев и пней в сосняке установлены еще два старых пожара 1841 и 1772 гг., где пожар 1841 г. способствовал формированию первого (151–173 лет) поколения сосны (рис. 2б). Такая же дата пожара нами установлена на ППП 4 (рис. 1в). Однако эти два сосняка находятся на противоположных сторонах реки на расстоянии 2 км друг от друга. По-видимому, текущий год был засушливым и способствовал большому распространению огня на территории. В сосняке черничном (ППП 4) установлены два пожара, последний из которых прошел 178 лет назад в 1841 г. (рис. 1в). Сосна в древостое представлена пятью поколениями с преобладанием двух условно-разновозрастных, состоящих из старых деревьев второго поколения (290–360 лет) и деревьев пятого поколения (92–164 лет), сформировавшихся после пожара. Остальные три поколения деревьев сосны представлены единичными особями и состоят из остатков некогда преобладающего первого 400-летнего и двух 189- и 260-летних поколений деревьев, выживших при пожарах. Фитоценоз сосняка черничного на ППП 10 был охвачен двумя пожарами, прошедшими в 1905 и 1868 гг. Из рис. 1г видно, что пожар 1868 г. вызвал “скачок” возобновления, тем самым сформировав третье поколение сосны. За прошедшие 114 лет после второго пожара возобновление сосны практически отсутствовало.
Рис. 1.
Распределения деревьев сосны по возрасту в сосняках: а, б – брусничных (ППП 1, 12); в, г – черничных (ППП 4, 10). (1) 2007 г. – а, 2012 г. наблюдений – б, 2009 г. – в, 2011 г. – г и (2) 2018 г. – а, 2019 г. – б, в, г. Стрелками указаны даты пожаров.
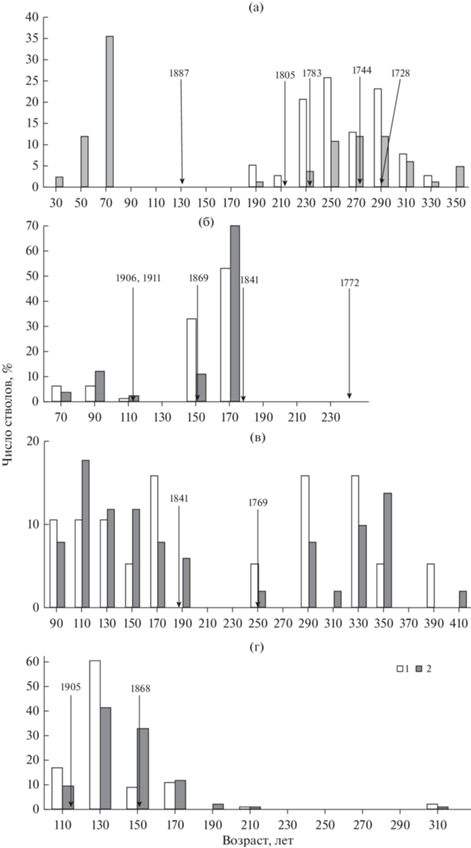
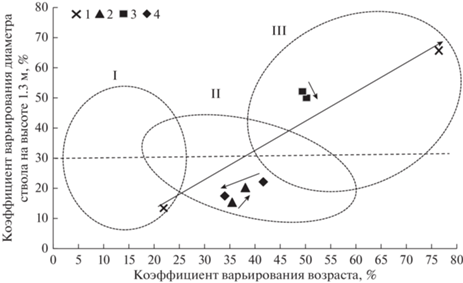
Рис. 2.
Изменение коэффициентов вариации возраста и диаметра деревьев сосны в сосняках: брусничных 1 (ППП 1) и 2 (ППП 12); черничных 3 (ППП 4) и 4 (ППП 12). Овалами показаны области группирования типов возрастной структуры, полученных ранее см. по тексту. Стрелками показаны изменения коэффициентов вариации за исследуемый период.
Динамика возрастной и размерной структуры. За десятилетний период наблюдений в сосняках брусничных отмечается увеличение густоты деревьев на ППП 1 со 170 до 528, на ППП 12 – с 316 до 500 экз. га–1. Следует отметить, что на ППП 1 пополнение деревьев идет за счет соснового подроста, тогда как на ППП 12 в основном за счет крупных особей подроста ели и кедра. На ППП 1, несмотря на существенное увеличение численности особей молодого поколения сосны, наблюдается небольшая тенденция увеличения на 1.8 м3 га–1 объема стволовой древесины. Невысокое увеличение запаса стволов в сосновом фитоценозе объясняется постепенным отпадом в нем деревьев старого материнского поколения. Сформированное новое поколение древостоя, состоящее из тонкомерных деревьев диаметром от 6 до 15 см, пока не оказывает существенного влияния на рост древесины стволов. Из табл. 1 видно, что, несмотря на снижение густоты сосны в древостое сосняка (ППП 12), происходит увеличение запаса стволовой древесины со 178 до 185 м3 га–1. Такая тенденция накопления объема древесины объясняется наличием в древостое относительно большого числа деревьев старшего поколения в возрасте 150–173 лет, способных еще накапливать древесину. Проанализированные древесные керны показали, что за период наблюдений максимальные значения прироста по радиусу отмечались у деревьев сосны в возрасте 160 лет и составили в среднем 3.3 см. Количество отпавших деревьев сосны и ели в древостое исследуемого сосняка увеличивается с 47 до 80 экз. га–1 с пополнением объема мертвой стволовой древесины на 5 м3 га–1 (табл. 1). Выявлено усыхание единичных деревьев сосны, а также ветровальных и сломанных снегом особей сосны.
С образованием нового разновозрастного поколения на ППП 1 и пополнением его новыми деревьями сосны из подроста на ППП 12 изменяется амплитуда колебания возрастных и морфометрических показателей древостоев (табл. 2). С появлением нового поколения в составе древостоя сосняка на ППП 1 отмечается резкое изменение коэффициентов вариации всех наблюдаемых признаков с их увеличением на 30–55%, тогда как на ППП 12 данные показатели изменяются от 1 до 4%.
Таблица 2.
Изменения статистических показателей древостоев сосняков
| Древесная порода (год наблюдения) | Поко-ление | MA ± mMA, (Amin–A max) | CVA, АsА, ЕА | MD ± mMD, (Dmin–Dmax) | CVD, АsD, ED | MH ± mMH, (Hmin–Hmax) | CVH, AsH, EH |
|---|---|---|---|---|---|---|---|
| Брусничный (ППП 1) | |||||||
| С (2007) | I | 264 ± 6, (181–344) | 13.3, 0.4, 0.5 | 39.1 ± 1.6 (20.0–62.0) | 21.9, –0.2, 3.5 | 20.2 ± 0.6 (12.0–24.5) | 11.7, –0.1, –0.8 |
| С (2018) | I–II | 170 ± 12, (34–355) | 65.6, 0.2, –1.8 | 18.2 ± 1.2 (6.1–62.0) | 76.4, 0.9, –0.7 | 13.0 ± 0.5 (4.8–25.1) | 40.8, 0.6, –0.8 |
| (2018) | I | 279 ± 6, (192–355) | 12.7, 0.2, 0.1 | 35.4 ± 1.2 (21.2–62.0) | 23.5, –0.2, –1.3 | 19.3 ± 0.4 (12.4–25.0) | 15.6, –0.2, 0.1 |
| (2018) | II | 63 ± 1, (34–75) |
14.7, –1.3, 1.7 | 8.7 ± 0.3 (6.1–14.9) |
25.8, 0.6, –0.6 | 9.6 ± 0.2 (4.8–15.0) |
22.4, 0.1, –0.5 |
| Е (2018) | 50–61 | – | 6.2–6.7 | – | 5.2–6.9 | – | |
| Брусничный (ППП 12) | |||||||
| С (2012) | I–II | 152 ± 3 (96–165) | 15.3, –2.5, 4.8 | 26.6 ± 0.9 (6.0–45.9) | 35.5, 0.14, –0.5 | 17.0 ± 0.5 (5.0–22.5) | 22.5, –0.8, 0.1 |
| С (2019) | I–II | 154 ± 3 (72–173) | 19.8, –1.8, 1.4 | 26.8 ± 1.0 (6.0–48.1) | 36.4, –0.2, –0.4 | 17.5 ± 0.4 (7.3–23.6) | 23.3, –0.8, 0.3 |
| (2012) | I | 159 ± 1 (143–165) | 3.7, –1.7, 1.9 | 28.4 ± 1.0 (14.0–45.9) | 27.7, 0.2, –0.6 | 17.7 ± 0.4 (10.5–22.5) | 16.8, –0.5, –0.4 |
| (2019) | I | 167 ± 1 (151–173) |
3.5, –1.7, 1.9 | 29.7 ± 1.0 (15.3–48.1) | 26.5, –0.2, –0.6 | 19 ± 0.4 (10.8–23.6) |
16.6, –0.5, –0.4 |
| (2012) | II | 87 ± 4 (72–104) | 11.9, –0.13, 0.03 | 10.1 ± 1.1 (6.0–23.1) | 31.3, –0.3, –1.5 | 10.4 ± 0.9 (7.0–13.5) | 25.3, –0.2, –1.6 |
| (2019) | II | 89 ± 3 (72–112) | 12.5, 0.5, –0.3 | 11.7 ± 1.5 (6.0–26.1) | 46.6, 1.5, 3.2 | 10.9 ± 0.8 (7.3–15.8) | 26.7, 0.1, –1.4 |
| E (2012) | I | 136 ± 6 (96–158) | 13.9, –0.9, 1.4 | 12.9 ± 0.4 (8.0–22.0) | 28.8, 0.7, –0.2 | 9.6 ± 1.6 (5.0–19.5) |
51.9, 1.3, 0.5 |
| Е (2019) | I | 138 ± 5 (80–166) |
17.0, –0.7, 0.1 | 10.8 ± 0.4 (6.0–22.9) | 38.1, 0.8, 0.1 | 9.3 ± 0.4 (3.7–19.4) |
38.7, 0.7, –0.1 |
| К (2012) | I | 108 ± 5 (70–140) | 17.7, –0.7, 0.5 | 14.2 ± 1.3 (8.0–24.0) | 39.4, 0.3, –1.4 | 9.8 ± 1.5 (5.0–16.5) |
43.3, 0.5, –1.5 |
| К (2019) | I | 112 ± 5 (78–148) |
18.2, 0.1, –0.3 | 12.5 ± 1.1 (6.3–25.7) | 47.1, 0.9, –0.5 | 9.9 ± 0.7 (4.2–17.1) |
39.0, 0.4, –1.1 |
| Черничный (ППП 4) | |||||||
| С (2009) | I–V | 198 ± 25 (82–400) |
50.2, 0.6, –1.4 | 36.4 ± 2.0 (8.5–72.0) | 50.0, –0.04, –1.4 | 22.2 ± 0.5 (14.6–31.2) | 19.3, 0.1, –1.5 |
| С (2019) | I–V | 211 ± 14 (94–410) |
47.4, 0.5, –1.5 | 31.1 ± 1.7 (6.8–72.0) | 52.0, 0.5, –1.2 | 20.0 ± 0.5 (8.2–31.2) | 24.0, –0.1, –0.1 |
| (2009) | I | 400 | – | 60.0 | – | 28.0 | – |
| (2019) | I | 410 | – | 60.0 | – | 28.0 | – |
| (2009) | II | 321 ± 6 (280–350) |
7.3, –0.5, –0.1 | 48.3 ± 2.6 (31.4–72.0) | 22.3, 0.1, –0.9 | 27.8 ± 0.9 (22.0–31.2) | 8.2, 0.8, –1.4 |
| (2019) | II | 334 ± 4 (290–360) |
6.5, –0.6, –0.8 | 49.9 ± 2.5 (33.0–72.0) | 20.3, 0.2, –0.8 | 25.7 ± 0.7 (22.0–31.2) | 10.4, 0.4, –0.4 |
| (2009) | III | 250 | – | 38.5 | – | 20.5 | – |
| (2019) | III | 260 | – | 39.3 | – | 20.5 | – |
| (2009) | IV | 178 ± 5 (175–180) |
– | 24.7 (22.7–26.4) |
– | 22.7 (21.5–23.8) |
– |
| (2019) | IV | 188 (186–190) |
– | 26.4 (24.4–28.1) |
– | 22.7 (21.5–23.8) |
– |
| (2009) | V | 121 ± 4 (82–154) |
18.4, 0.1, –1.2 | 19.6 ± 1.1 (8.5–36.4) | 30.8, 0.8, 1.7 | 18.3 ± 0.6 (11.1–21.1) | 8.8, 0.4, 0.6 |
| (2019) | V | 132 ± 4 (94–164) |
16.6, –0.04, –1.3 | 20.8 ± 1.3 (6.8–37.4) | 33.0, 0.2, 0.4 | 18.6 ± 0.7 (8.2–25.4) | 20.3, –0.8, 1.3 |
| Е (2009) | I | 125 ± 3 (101–164) |
12.8, 0.6, –0.7 | 12.4 ± 0.8 (7.0–22.4) | 35.4, 0.5, –0.9 | 16.7 ± 3.4 (12.7–22.4) | 20.5, 0.2, 1.2 |
| Е (2019) | I | 100 ± 2 (105–174) |
12.4, 0.4, –0.3 | 10.5 ± 0.3 (6.0–28.9) | 39.9, 1.3, 0.1 | 9.9 ± 0.3 (3.0–22.8) |
42.7, 0.7, 0.1 |
| Черничный (ППП 10) | |||||||
| С (2011) | I–IV | 137 ± 3 (101–312) |
22.1, 3.9, 20.2 | 21.4 ± 0.9 (7.4–56.2) | 41.7, 1.3, 3.0 | 19.0 ± 0.5 (5.0–26.5) | 25.1, –0.6, –0.2 |
| С (2019) | I–IV | 144 ± 3 (111–316) |
17.3, 3.9, 24.5 | 22.2 ± 0.5 (7.9–58.5) | 34.0, 0.9, 2.1 | 19.3 ± 0.2 (8.4–28.5) | 17.5, –0.6, 0.4 |
| (2011) | I | 312 (308–312) |
– | 55.1 (53.9–56.2) |
– | 26.0 (25.5–26.5) |
– |
| (2019) | I | 316 | – | 58 | – | 27.4 | – |
| (2011) | II | 210 | – | 48.1 | – | 25.5 | – |
| (2019) | II | 218 | – | 49.9 | – | 26.4 | – |
| (2011) | III | 164 ± 1 (161–173) |
2.7, 3.1, 0.6 | 28.1 ± 2 (13.7–36.9) |
23.1, –1.2, 1.4 | 21.1 ± 1.4 (11.5–25.5) | 21.3, –1.2, 0.7 |
| (2019) | III | 171 ± 2 (162–181) |
3.3, 0.4, 0.2 | 28.0 ± 2 (14.5–38.7) |
26.4, –0.7, 0.4 | 21.1 (12.4–26.5) |
23.0, –0.7, –0.9 |
| (2011) | IV | 129 ± 1 (101–159) |
9.1, –0.3, 0.4 | 19.5 ± 0.7 (7.4–36.2) | 34.0, 0.3, –0.4 | 18.5 ± 0.5 (5.0–26.0) | 25.1, –0.6, –0.1 |
| (2019) | IV | 136 ± 1 (111–156) |
7.4, –0.7, 0.3 | 21.6 ± 0.7 (10.0–38.0) | 28.5, 0.3, –0.3 | 19.7 ± 0.5 (8.4–26.9) | 21.7, –0.5, –0.2 |
| Е (2011) | I | 98 ± 5 (79–104) |
10.7, –2.1, –4.6 | 10.0 ± 06 (8.0–16.0) |
25.3, 0.9, 0.03 | 7.8 ± 1.3 (6.0–11.5) |
32.2, 1.8, 3.2 |
| Е (2019) | I | 112 ± 2 (87–132) |
8.7, –0.7, 2.2 | 7.6 ± 0.3 (6.0–10.3) |
18.0, 0.3, –0.4 | 6.1 ± 0.5 (3.9–9.9) |
25.8, 1.2, 2.7 |
Примечания: A – возраст (лет), D – диаметр (см), H – высота (м). MХ ± mMХ – среднеарифметическое значение наблюдаемых признаков и его ошибка, Хmin-Х max – минимальное и максимальное значение наблюдаемых признаков, CVХ – коэффициент вариации, AsХ – асимметрия, ЕХ – эксцесс. Прочерк указывает на недостаточное количество данных для статистической оценки.
Для оценки дифференциации биометрических параметров деревьев в древостоях необходимо учитывать такие показатели, как асимметрия и эксцесс (Кузьмичев, 2013). С формированием нового поколения в древостое сосняка брусничного (ППП 1) асимметрия возраста изменяется незначительно. Эксцесс переходит в отрицательное значение –1.8 (табл. 2), указывая на наличие двух обособленных кривых или поколений в древостое (рис. 1а). Показатель асимметрии диаметра деревьев сосны в древостое коренного сосняка за десять лет изменяется с –0.2 до 0.9, свидетельствуя о появлении в нем тонких деревьев. В первом поколении асимметрия диаметра остается в пределах своих значений (–0.2). Показатель асимметрии по толщине деревьев второго поколения показывает на преобладание в древостое тонких особей от 6 до 10 см, на что указывает средняя положительная асимметрия (0.6). Показатель эксцесса деревьев сосны в древостое по диаметру переходит из положительной островершинной (3.5) в отрицательную плосковершинную кривую (–0.7). Асимметрия диаметра на современном этапе развития сосняка как в древостое, так и в поколениях отрицательная, что говорит о рассеянности диаметров деревьев сосны (табл. 2).
По мнению Е.С. Мельникова (2002), при сильной конкуренции деревьев в фитоценозах с преобладающим участием деревьев близких размеров отмечается высокая положительная величина эксцесса. По нашим данным (табл. 2), у сосны в первом поколении сосняка брусничного (ППП 1), несмотря на повышение конкуренции со стороны деревьев второго поколения, показатель эксцесса увеличивается незначительно, смещаясь в сторону нулевого значения. Такая тенденция, вероятно, связана с частичным отпадом угнетенных деревьев первого поколения и, как следствие, увеличением концентрации деревьев в кривой высот около среднего значения, тогда как по диаметру эксцесс в первом поколении снижается, переходя из островершинной (3.5) в плосковершинную кривую (–1.3).
Показатели асимметрии возраста деревьев сосны на ППП 12 остаются в пределах отрицательных значений. С пополнением тонкомерных деревьев второго поколения этот показатель смещается к нулевому значению с –2.5 до –1.8. С увеличением амплитуды колебания возраста значение эксцесса смещается в сторону нуля, изменяясь от 4.8 до 1.8. В первом поколении показатели асимметрии и эксцесса остаются в пределах своих значений. Во втором поколении асимметрия возраста смещается в сторону преобладания младших по возрасту особей сосны, изменяясь с –0.1 до 0.5, что говорит о небольшом пополнении данного поколения молодыми особями сосны из подроста. Эксцесс возраста увеличивается от 0.03 до 0.3. У ели и кедра с увеличением густоты тонкомерных молодых особей в древостое сосняка асимметрия возраста повышается, занимая правостороннюю кривую. Значение эксцесса снижается.
Коэффициенты асимметрии по толщине у сосны в древостое сосняка брусничного (ППП 12) за восемь лет смещаются в отрицательную сторону, о чем свидетельствует увеличение преобладающего количества деревьев выше среднего диаметра. Схожая тенденция отмечается у сосны в первом поколении. Показатели эксцесса в сосновой части древостоя и в первом поколении остаются на прежнем уровне. Во втором поколении сосны асимметрия смещается из левосторонней (–0.3) в правостороннюю кривую (1.5), свидетельствуя об увеличении тонких деревьев сосны. У деревьев сосны второго поколения эксцесс распределения деревьев по толщине смещается в положительную сторону, кривая становится более островершинной. У темнохвойных видов, участвующих в составе древостоя сосняка, отмечается схожая тенденция перестроения асимметрии по толщине в сторону положительного значения.
В сосняке черничном (ППП 4) за период наблюдений выявлено небольшое (на 2.2%) снижение запасов стволовой древесины. Основное снижение ее объема происходит в сосновой части древесного яруса в результате усыхания деревьев старых поколений и повреждения снегом тонкомерных стволов сосны. Густота соснового элемента за десять лет уменьшается с 243 до 223 экз. га–1. Количество сухостойных деревьев сосны увеличивается с 28 до 45 экз. га–1. В отличие от брусничных сосняков (ППП 1 и 12), в древостое сосняка черничного на ППП 4 за прошедший период сосновая часть ценоза пополнилась только тремя деревьями сосны, которые перешли из особей крупного подроста. За десять лет с начала наблюдений в древостое увеличивается густота деревьев ели с 198 до 573 экз. га–1 и, как следствие, отмечается повышение объема стволовой древесины на 5.3 м3 га–1.
При первой инвентаризации в древостое сосняка черничного (ППП 10) был выявлен активный отпад деревьев сосны старших поколений. Причиной их массовой гибели, вероятно, послужил крупный ветровал, прошедший в непосредственной близости от ППП 10 в 2009 г. В 2011 г. на обследуемом участке сосняка была отмечена, по визуальным наблюдениям, вспышка короеда. В сосняке присутствовали единично вываленные деревья и большое количество свежего сухостоя. Как показали последние наблюдения, распад старых поколений продолжился, что подтверждается снижением густоты сосны с 553 до 500 экз. га–1 и понижением запаса стволовой древесины сосны на 8.3 м3 га–1. При повторном перечете отмечена вызванная снеголомом гибель особей сосны мелкой категории, относящихся к четвертому поколению.
В связи с постепенным отмиранием старых деревьев сосны в сосняке (ППП 4) коэффициенты вариации по всем наблюдаемым признакам в сосновой части древостоя за десять лет наблюдений изменяются незначительно, оставаясь в пределах больших и средних значений (табл. 2). В пределах поколений величина коэффициента изменчивости показателей также остается постоянной, за исключением пятого поколения, где вариация диаметра увеличивается с 30.8 до 33.0%, высоты – с 8.8 до 20.3% (табл. 2), свидетельствуя о неравномерности роста деревьев и увеличении амплитуды колебаний этих признаков. В процессе распада отставших в росте мелких деревьев сосны в древостое на ППП 10 и увеличения высоты за десять лет сосновая часть второго яруса перешла в первый (табл. 1). Об этом свидетельствует снижение вариации возраста с 22 до 17, диаметра с 42 до 34%. С выпадением мелких деревьев значение средней высоты увеличивается незначительно, однако коэффициент изменчивости высоты становится меньше, снижаясь с 25 до 17.5% (табл. 2).
Асимметрия возраста сосны в древостое сосняка черничного (ППП 4) за наблюдаемый период остается в пределах положительных значений, указывая на преобладание деревьев младшего пятого поколения. В пределах поколений асимметрия возраста также постоянна. Отрицательная величина эксцесса в древостое и в поколениях свидетельствует о плосковершинности кривых распределения деревьев по возрасту. В исследуемом сосняке черничном в связи со снижением среднего диаметра и высоты показатели асимметрии диаметра смещаются в сторону преобладания мелких по толщине деревьев. Асимметрия распределения деревьев по высоте изменяется незначительно, смещаясь в отрицательную сторону. Показатель эксцесса рядов распределения деревьев по диаметру в сосняке смещается к нулевому значению. Несмотря на усыхание крупных деревьев второго поколения, значения асимметрии и эксцесса диаметра остаются в пределах постоянных значений. В пятом поколении с увеличением среднего диаметра сосны асимметрия смещается в сторону преобладания средних по толщине деревьев, изменяясь от 0.8 до 0.4, что указывает на относительно хороший прирост большинства особей сосны данного поколения. Показатель эксцесса изменяется с 1.7 до 0.4.
Смещение показателей асимметрии по диаметру в сторону нулевого значения с 1.3 до 0.9 говорит об увеличении числа крупных деревьев в древостое сосняка черничного (ППП 10). Эксцесс по толщине снижается, приобретая черты плосковершинной кривой. В третьем поколении деревьев сосны коэффициент асимметрии стремится к нулевому значению с –1.2 до –0.7, тогда как в младшем поколении асимметрия толщины остается на прежнем уровне (0.3). По высоте показатели меры косости в поколениях смещаются также в сторону нулевых значений. Коэффициент эксцесса высоты как в древостое, так и в поколениях в процессе его развития варьирует. Следует отметить, что за наблюдаемый период (восемь лет) все деревья сосны перешли в первый ярус сосняка. Такой переход определяется отпадом отставших в росте деревьев младших поколений и хорошим приростом их по высоте.
Согласно распределению деревьев по возрасту на ППП 4 (рис. 3а) видно, что ель в возрасте от 105 до 174 лет параллельно с сосной активно занимала пространство после пожара 1841 г. С поступлением в еловую часть древостоя тонкомерных деревьев отмечается снижение ее средних таксационных показателей. Увеличиваются значения варьирования по всем наблюдаемым признакам. Асимметрия диаметра и высоты смещаются в сторону преобладания мелких деревьев ели. Эксцесс диаметра переходит в положительную сторону, тогда как по высоте кривая становится более плосковершинной, смещаясь в отрицательную сторону.
Рис. 3.
Распределение деревьев по возрасту хвойных видов в сосняках: а – черничном (ППП 4); б – брусничном (ППП 12); 1 – сосна обыкновенная, 2 – ель сибирская; 3 – сосна сибирская (кедр). Условные обозначения см. рис. 1.
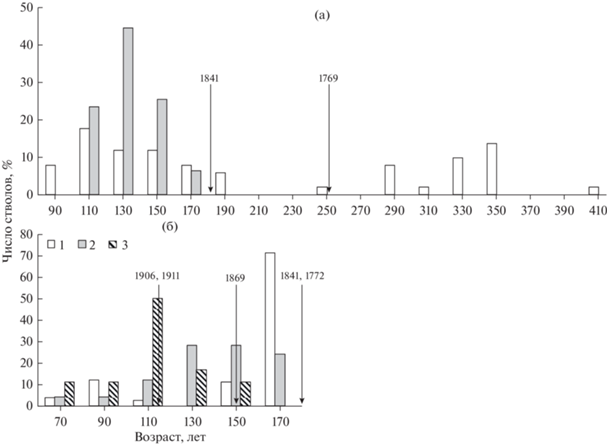
В северотаежных сосняках Архангельской области и Республики Коми при длительном отсутствии или слабой пожарной активности ель способна занимать значительные пространства в лишайниковых сообществах, вытесняя из них сосну (Листов, 1986; Семенов и др., 1998). При высокой интенсивности пожара в сосняках темнохвойные виды гибнут неминуемо (Непомилуева, 1974). На участках сосняка брусничного и черничного (ППП 4, 12) при длительном отсутствии пожаров или их слабой активности наблюдается тенденция смены светлохвойных сосновых сообществ на темнохвойные. Так, по характеру горения последние три пожара в сосняке брусничном на ППП 12 (1869, 1906 и 1911 гг.), по-видимому, носили характер слабой пирогенной активности, о чем свидетельствует присутствие в составе древостоя сосняка большого количества особей ели и кедра. В сосняке входящие в состав ель и кедр представлены одним поколением деревьев, имеющим приблизительно схожий максимальный и минимальный возраст (табл. 2). Прошедшие беглые низовые пожары в сосняке не вызывали полную гибель темнохвойных видов, а, напротив, способствовали их увеличению в составе древостоя (рис. 3б). Пожар 1869 г. вызвал “вспышку” возобновления ели и кедра. Однако после этого пожара возобновление сосны практически отсутствовало, что, видимо, связано с загущенностью древостоя особями сосны первого поколения, препятствующего появлению нового поколения соснового подроста. Последние два пожара, прошедшие с коротким интервалом в пять лет (1906 и 1911 гг.), уничтожили часть деревьев подроста светлохвойных и темнохвойных видов, способствуя появлению второго поколения сосны, а также вспышке возобновления кедра. Как показали наблюдения в данном древостое, пожары низкой интенсивности иногда могут содействовать смене более пожароустойчивых светлохвойных сообществ на темнохвойные. Теневыносливые ель и кедр, развивающиеся на автоморфных подзолистых почвах, хотя и отличаются замедленным ростом по диаметру и высоте, но способны сдерживать возобновление сосны и даже вытеснять ее со временем при длительном отсутствии или слабой активности пожаров. Соответственно, появление в составе древостоев сосняков темнохвойных видов приведет к межвидовой конкуренции, что, в свою очередь, отразится на размерной и возрастной структуре древостоя (Lilja, Kuuluvainen, 2005).
В притундровых сосняках брусничных и черничных типов по мере увеличения числа поколений деревьев в древостоях коэффициенты варьирования возраста, диаметра и высоты увеличиваются, иногда превышая 70% (Семенов и др., 1998). Построенная нами ранее (Кутявин, 2018) модель связи коэффициентов изменчивости возраста с диаметром стволов в сосняках Северного Приуралья и модель, предложенная ранее для ельников Архангельской области И.И. Гусевым (1962), позволила достаточно точно прогнозировать изменения возрастной структуры древостоев в динамике (рис. 2). Представленные области: I – условно-разновозрастный, II – относительно-разновозрастный с демутационными фазами динамики и III – ступенчато-разновозрастный тип возрастной структуры – были построены для исследованных постпирогенных сосняков ранее (Кутявин, Манов, 2020). Из приведенной модели видно смещение вариации диаметра и высоты на ППП из одного типа возрастной структуры в другой при первом и последующем исследованиях (на рис. 2 показаны стрелками). Так, в сосняках брусничных с появлением нового поколения (ППП 1) или пополнением молодого поколения новыми деревьями сосны (ППП 12) вариация смещается или стремится в область ступенчато-разновозрастных древостоев. В сосняках черничных на ППП 4 и 10 прослеживается тенденция снижения коэффициентов варьирования, что связано с распадом старых материнских поколений и, как следствие, с их переходом в область условно-разновозрастных (ППП 10) или относительно-разновозрастных с демутационными фазами динамики (ППП 4) типов возрастной структуры.
Чистые по составу древостои сосняков Сибири находятся в состоянии постоянного динамического развития, в котором отпад деревьев маловероятен (Верхунов, 1976). Сосняки брусничные в Карелии при длительном отсутствии пожаров носят цикличный (волнообразный) тип распределения деревьев по возрасту, и этот тип сохраняется при пожарах слабой интенсивности. Возобновительный процесс в них непрерывен, поэтому разрывы между поколениями деревьев в древостоях не образуются (Зябченко, 1984). Проведенные нами ранее исследования возрастной структуры в древостоях коренных среднетаежных сосняков Республики Коми показывают, что цикличная структура распределения деревьев не встречается. Наиболее часто сосняки Приуралья представлены ступенчато-разновозрастной структурой с разорванными поколениями деревьев сосны и формируются даже при длительном отсутствии огня или его полном отсутствии, например, сосняки сфагновые (Кутявин, 2018; Кутявин, Манов, 2020).
Исследования, проведенные в центральной и северной Швеции, показали, что немаловажным фактором волнообразного возобновления сосны является климат (Agren, Zackrisson, 1990). Согласно нашим данным, в чистом по составу сосняке брусничном (ППП 1) и сосняках черничных (ППП 4, 10) с примесью в составе ели древостои характеризуются ступенчатой возрастной структурой с разрывами между поколениями. При этом в двух случаях разрывы между поколениями вызваны не пожарной активностью, а перезагущенностью пространства деревьями верхнего яруса и как следствие полным отсутствием возобновления. В сосняке брусничном (ППП 1) до момента образования второго поколения возобновление отсутствовало на протяжении 117 лет (рис. 1а). В сосняке черничном (ППП 10), несмотря на активное усыхание деревьев первых поколений, сопровождаемое освобождением пространства для возобновления, количество соснового подроста остается незначительным (табл. 1). Длительное отсутствие возобновления сосны также может ограничиваться беглыми низовыми пожарами слабой интенсивности (Листов, 1986). Такие пожары достаточно сложно установить при датировке пирогенной активности на участках (Кутявин, 2018). Ранее показано (Галенко, 1983), что в черничных сосняках молодые особи сосны под пологом древостоя не испытывают недостатка светового довольствия. Основной причиной отсутствия возобновления сосны под пологом древостоев, развивающейся на автоморфных почвах, является нехватка элементов минерального питания в почве, в частности, таких как азот, калий, фосфор, а также влияние старших поколений деревьев сосны, перехватывающих основные питательные элементы (Листов, 1986). Такие условия ведут к жестким конкурентным отношениям между особями деревьев, причем, чем старше древостой, тем сильнее влияние микроценотических эффектов (Демаков, 2018). Известно также, что после пожаров в сосняках на автоморфных почвах происходит улучшение эдафических условий, приводящее к “взрыву” возобновления сосны (Engelmark, 1993; Angelstam, 1998).
Накопленная под пологом сосняка черничного (ППП 4) ель, несмотря на то, что характеризуется относительно низкими темпами роста, способна препятствовать возобновлению сосны. Обследования в 2009 и 2019 гг. показали наличие в его составе подроста единичных особей сосны, находящихся на стадии самосева или подроста мелкой категории крупности (0.25–0.5 м), располагающихся преимущественно в окнах и на микроповышениях. За наблюдаемые десять лет под пологом сосняка более чем в два раза увеличивается количество подроста с преобладанием темнохвойных видов. В составе подроста значительно (до 60%) увеличивается доля кедра. Береза и ель участвуют в равных долях (20%), тогда как сосна теряет свои позиции, снижая участие в возобновлении с 10 до 4% (табл. 1). За десятилетний период наблюдений отмечается более чем в два раза увеличение количества в составе древостоя березы, пополняющейся также из тонкомера и поросли старых поваленных деревьев. Крупные особи березы имеют схожий возраст с сосной (82–170 лет) пятого поколения и елью. Максимальное значение возраста березы свидетельствует о ее одновременном послепожарном возобновлении с сопутствующими в составе породами. Максимальный возраст кедра составляет 123 года, что говорит о его закреплении на данном участке сосняка только через 50 лет после пожара (табл. 2).
Большая роль в продуктивности лесов в последнее время связана с изменением климатических условий. Согласно исследованиям в управляемых лесах Финляндии за период с 1971 по 2010 гг., средний прирост объема древесины составил порядка 37%, где наиболее высокая продуктивность отмечается в северных районах страны, достигая 45% от общих объемов (Henttonen et al., 2017). Проведенные недавние исследования на ППП в смешанных горных лесах Словакии (Sumichrast et al., 2020) показали довольно высокий прирост древесины и увеличение густоты в хвойно-лиственных древостоях. По мнению авторов, одной из причин увеличения продукционной способности этих древостоев является потепление климата. По нашим данным, полученным на пробных площадях, продуктивность стволовой древесины в коренных сосняках остается на довольно стабильном уровне. Увеличение запаса древесины отмечается в сосняках брусничных на 1.8 м3 га–1 (ППП 1), что связано с появлением в древостое новых поколений деревьев сосны, а также относительно хорошим приростом старших поколений в возрасте 160 лет на 7 м3 га–1 (ППП 12). Черничные сосняки, наоборот, теряют запасы древесины на 7.5–18.6 м3 га–1, что связано с распадом старых материнских поколений и длительным отсутствием возобновления сосны под их пологом. Постепенное снижение продукционной способности сосны в сосняках черничных (ППП 4, 10) обусловлено длительным отсутствием пожарной активности и сильной загущенностью медленно развивающейся ели, препятствующей возобновлению сосны. С.С. Зябченко (1984) указывал, что пирогенная активность во многом способствует образованию, росту и развитию новых поколений сосны в сосняках и увеличивает устойчивость и продукционную способность коренных сосновых экосистем Севера.
ЗАКЛЮЧЕНИЕ
Десятилетние наблюдения за динамикой развития древостоев в коренных среднетаежных сосняках с давностью пирогенного воздействия более ста лет назад показали значительную динамичность изменения в их строении. Вариационный анализ размерной и возрастной структуры указывает на относительно большие изменения статистических показателей морфоструктуры древостоев. С отпадом угнетенных деревьев сосны в старших поколениях отмечается увеличение средней высоты, что, вероятно, связано с увеличением конкуренции со стороны младших поколений и сопутствующих в составе древесных пород и приводит к усыханию угнетенных деревьев старших поколений. При этом показатели асимметрии диаметра в данных поколениях находятся на относительно стабильном уровне. Приведенная модель связи варьирования возраста с изменением диаметра стволов для среднетаежных сосняков наглядно показывает переход древостоев из одного типа возрастной структуры в другой. При длительном отсутствии или слабой пожарной активности, а также при участии в составе древостоя ели наблюдается тенденция сукцессионной смены сосновых сообществ на еловые. Важными факторами в развитии древостоев в сосновых фитоценозах являются климатические и почвенно-экологические факторы. Дальнейшее накопление знаний о динамике развития коренных сосняков Северного Приуралья позволит глубже познать их современную природу в пространственно-временном интервале развития, а также разработать охранно-хозяйственные мероприятия.
Список литературы
Березовская Ф.С., Карев Г.П. Моделирование динамики древостоев // Сибирский лесной журн. 2015. № 3. С. 7–19. https://doi.org/10.15372/SJFS20150302
Верхунов П.М. Закономерности строения разновозрастных сосняков. Новосибирск: Наука, 1976. 255 с.
Войнов Г.С., Чупров Н.П., Ярославцев С.В. (ред.) Лесотаксационный справочник по северо-востоку европейской части Российской Федерации (нормативные материалы для Ненецкого автономного округа, Архангельской, Вологодской областей, Республики Коми). Архангельск: Правда Севера, 2012. 672 с.
Гавриков В.Л. О концептуальности модели смены поколений хвойных // Структурно-функциональные взаимосвязи и продуктивность фитоценозов. Красноярск, 1983. С. 20–31.
Галенко Э.П. Фитоклимат и энергетические факторы продуктивности хвойного леса Европейского Севера. Л.: Наука, 1983. 129 с.
Гусев И.И. К вопросу о возрастной структуре ельников Архангельской области // Лесной журн. 1962. № 2. С. 20–27.
Гусев И.И. Моделирование экосистем. Архангельск: АГТУ, 2002. 112 с.
Демаков Ю.П. Структура и закономерности развития лесов Республики Марий Эл. Йошкар-Ола: Поволжский гос. технолог. ун-т, 2018. 432 с.
Дыренков С.А. Структура и динамика таежных ельников. Л.: Наука, 1984. 174 с.
Жила С.В., Иванова Г.А., Иванов В.А, Цветков П.А. Лесовозобновление после пожаров разной интенсивности в сосняках Средней Сибири // Сибирский лесной журн. 2019. № 6. С. 53–62. https://doi.org/10.15372/SJFS20190606
Зябченко С.С. Сосновые леса Европейского Севера. Л.: Наука, 1984. 244 с.
Комин Г.Е., Семечкин И.В. Возрастная структура древостоев и принципы ее типизации // Лесоведение. 1970. № 2. С. 24–33.
Кузьмичев В.В. Закономерности динамики древостоев: принципы и модели. Новосибирск: Наука, 2013. 208 с.
Кутявин И.Н. Сосновые леса Северного Приуралья: строение, рост, продуктивность. Сыктывкар: ИБ Коми НЦ УрО РАН, 2018. 176 с. DOI
Кутявин И.Н., Манов А.В. Возрастная структура древостоев под воздействием пожаров в сосняках Северного Приуралья (республика Коми) // Вестник Северо-Восточного центра ДВО РАН. Биологические науки. 2020. № 3. С. 47–59. https://doi.org/10.34078/1814-0998-2020-3-47-59
Листов А.А. Боры-беломошники. М.: Агропромиздат, 1986. 181 с.
Манов А.В., Кутявин И.Н. Размерная, возрастная и пространственная структура древостоев постпирогенных среднетаежных сосняков на автоморфных почвах (на примере Республики Коми) // Сибирский лесной журн. 2019. № 6. С. 100–110. https://doi.org/10.15372/SJFS20190611
Манов А.В. Динамика горизонтальной структуры древостоя коренного разнотравно-черничного ельника подзоны средней тайги Республики Коми // Известия РАН. Серия биологическая. 2017. № 5. С. 573–580. https://doi.org/10.7868/S0002332917050137
Мельников Е.С. Конкуренция и компромиссное распределение ресурсов в древостое: Результаты изучения автоколебания радиального прироста // Известия СПбЛТА. 2002. Т. 168. Вып. 10. С. 25–31.
Мишко А.Е., Ставрова Н.И., Горшков В.В. Онтогенентическая структура популяций Picea obovata (Pinacea) на разных стадиях послепожарных сукцессий в северотаежных сосново-еловых лесах // Ботанический журн. 2018. Т. 103. № 9. С. 1124–1152. https://doi.org/10.7868/S0006813618090041
Нагимов З.Я., Нагимов В.З., Артемьева И.Н. Ход роста сосновых древостоев в лишайниковом типе леса Ханты-Мансийского автономного округа // Известия высших учебных заведений “Лесной журн.”. 2010. № 5. С. 7–11.
Непомилуева Н.И. Кедр сибирский (Pinus sibirica Du Tour) на северо-востоке европейской части СССР. Л.: Наука, 1974. 184 с.
Осипов А.Ф., Бобкова К.С. Биологическая продуктивность и фиксация углерода среднетаежными сосняками при переходе из средневозрастных в спелые // Лесоведение. 2016. № 5. С. 346–354.
Проскуряков М.А. Проблема хронобиологической цикличности движения свойств лесных экосистем. Сообщение 1 // Сибирский лесной журн. 2015. № 2. С. 71–84. https://doi.org/10.15372/SJFS20150607
Рысин Л.П., Вакуров А.Д., Павлова В.Ф. Значение постоянных пробных площадей в лесоводственных исследованиях // Лесоведение. 1981. № 1. С. 60–66.
Семенов Б.А., Цветков В.Ф., Чибисов Г.А., Елизаров Ф.П. Притундровые леса европейской части России (природа и ведение хозяйства). Архангельск: Пресс А, 1998. 334 с.
Цветков В.Ф. Сосняки Кольской лесорастительной области и ведение хозяйства в них. Архангельск: АГТУ, 2002. 380 с.
Швиденко А.З., Щепащенко Д.Г. Кракснер Ф., Онучин А.А. Переход к устойчивому управлению лесами России: теоретико-методические предпосылки // Сибирский лесной журн. 2017. № 6. С. 3–25. https://doi.org/10.15372/SJFS20170601
Agren J., Zackrisson O. Age and size structure of Pinus sylvestris population on mires in central and northern Sweden // J. Ecology. 1990. V. 78. № 4. P. 1049–1062.
Angelstam P.K. Maintaining and restoring biodiversity in European boreal forests by developing natural disturbance regimes // J. Vegetation Science. 1998. V. 9. № 4. P. 593–602.
Bastos A., Gouvenia C.V., Trigo R.M. Analyzing the spatio-temporal impacts of the 2003 and 2010 extreme heatwaves on plant productivity in Europe // Biogeosciences. 2014. V. 11. № 13. P. 3421–3435. https://doi.org/10.5194/bg-11-3421-2014
Engelmark O. Early post-fire tree regeneration in a Picea-Vaccinium forests in northern Sweden // J. Vegetation Science. 1993. V. 4. № 6. P. 791–794.
Henttonen H.M., Pekka N. Makinen H. Environment-induced growth changes in the Finnish forests during 1971–2010 – An analysis based on National Forest Inventory // Forest Ecology and Management. 2017. V. 386. P. 22–36. https://doi.org/10.1016/j.foreco.2016.11.044
Holmes R.L. Computer-assisted quality control in tree-ring dating and measurement // Tree-Ring Bulletin. 1983. V. 43. P. 69–78.
Kulha N., Pasanen L., Holmstrom L., Grandpre L.D., Gauthier S., Kuuluvainen T., Aakala T. The structure of boreal old-growth forests changes at multiple spatial scales over decades // Landscape Ecology. 2020. V. 35(4). P. 843–858. https://doi.org/10.1007/s10980-020-00979-w
Kullman L. Pine (Pinus sylvestris) treeline dynamics during the past millennium – a population study in west-central Sweden // Ann. Bot. Fennici. 2005. V. 42. P. 95–106.
Kurz W.A., Dymond C.C., White T.M., Stinson G., Shaw C.H., Rampley G.J., Smyth C., Simpson B.N., Neilson E.T., Trofymow J.A., Metsaranta J., Apps M.J. CBM-CFS3: a model of carbon-dynamics in forestry and land-use change implementing IPCC standards // Ecological Modeling. 2009. V. 220. P. 480–504.
Lilja S., Kuuluvainen T. Structure of old Pinus sylvestris dominated Forest Stands along a geographic and human impact gradient in mid-boreal Fennoscandia // Silva Fennica. 2005. V. 39. № 3. P. 407–428.
Madany M.N., Swetnam T.W., West N.E. Comparison of two approaches for determining fire dates from tree scars // Forest Science. 1982. V. 28. № 4. P. 856–861.
Muller J., Boch S., Blaser S., Fisher M., Prati D. Effects of forest management on bryophyte communities on deadwood // Nova Hedwigia. 2015. V. 100. P. 423–438.
Rinn F. Tsap version 3.5. Reference Manual. Computer program for tree-ring analysis and presentation. Germany, Helenberg, Frank Rinn, 1996. 264 p.
Sumichrast L., Vencurik J., Pittner J., Kucbel S. The long-term dynamics of the old-growth structure in the National Nature Reserve Badinsky prales // J. Forest Science. 2020. V. 66. № 12. P. 501–510. https://doi.org/10.17221/139/2020-JFS
Дополнительные материалы отсутствуют.