

Лесоведение, 2022, № 5, стр. 451-460
Модели фитомассы деревьев березы и осины для дистанционного зондирования в климатических градиентах Евразии
В. А. Усольцев a, b, *, И. С. Цепордей a, И. М. Данилин c
a Ботанический сад УрО РАН
620144 Екатеринбург, ул. 8 Марта, д. 202а, Россия
b Уральский государственный лесотехнический университет
620100 Екатеринбург, ул. Сибирский тракт, д. 37, Россия
c Институт леса им. В.Н. Сукачева Сибирского отделения Российской академии наук
660036 Красноярск, Академгородок, 50, Россия
* E-mail: Usoltsev50@mail.ru
Поступила в редакцию 04.10.2021
После доработки 30.12.2021
Принята к публикации 06.04.2022
- EDN: DCZNPG
- DOI: 10.31857/S0024114822050072
Аннотация
Лесные экосистемы играют большую роль в стабилизации климата, и непрерывный мониторинг их фитомассы имеет первостепенное значение. Технология бортового лазерного сканирования получила широкое распространение при оценке фитомассы деревьев путем дистанционной регистрации таких таксационных показателей, как диаметр и площадь проекции кроны и высота дерева. В работе использована авторская база данных 755 и 296 модельных деревьев соответственно березы (Betula spp.) и осины (Populus spp.) Евразии с замеренными показателями высоты дерева и диаметра кроны, а также фитомассы ствола, листвы, ветвей и корней. Для всех компонентов фитомассы выявлена положительная связь с диаметром кроны и высотой дерева. На статистически достоверном уровне установлено, что фитомасса листвы, ветвей, стволов и надземная у деревьев березы соответственно на 4.7, 1.3, 9.6 и 8.0% больше, чем у осины, при равных значениях как диаметра кроны, так и высоты дерева. Это вызвано тем, что при одной и той же высоте дерева диаметр кроны у осины на 12% больше, чем у березы. Фитомасса всех компонентов равновеликих деревьев описывается пропеллеро-образной 3D-зависимостью от зимних температур и осадков. В теплых регионах при повышении осадков фитомасса увеличивается, но по мере перехода к холодным регионам она характеризуется противоположным трендом. При повышении температуры во влажных регионах фитомасса возрастает, но по мере перехода в сухие условия снижается. Вклад таксационных показателей деревьев, их видовой принадлежности и климатических переменных в объяснение изменчивости компонентов фитомассы составляет соответственно 79.0, 4.6 и 16.4%. Климатические переменные в наибольшей степени объясняют изменчивость фитомассы стволов, ветвей и надземной (от 18 до 22%) и в наименьшей степени – листвы (около 7%). Полученные результаты могут быть полезны при лазерном мониторинге фитомассы лесов и при прогнозировании возможных изменений в структуре фитомассы деревьев при климатических отклонениях.
Антропогенные изменения окружающей среды влияют на экосистемы на всех уровнях организации (Díaz et al., 2019). В функциональной биогеографии предполагается, что свойства растений отражают адаптацию растительности к изменяющимся условиям окружающей среды, в том числе климата (Reichstein et al., 2014). Современные глобальные модели растительности нацелены на реализацию адаптивных реакций растений в контексте теории оптимальности (Корзухин, Семевский, 1992). Поскольку реакция растений на стрессы проявляется в изменении их фитомассы, представляет интерес знание того, как изменения климата повлияют на фитомассу и способность лесов депонировать углерод (Franklin et al., 2020; Tautenhahn et al., 2020).
В течение последних двух десятилетий для оценки наземной фитомассы интенсивно используется технология бортового лазерного зондирования, обеспечивающая получение высокоточной информации о пространственных и темпоральных характеристиках лесов. Благодаря способности проникновения сквозь толщу полога, данные лазерного зондирования, представляющие трехмерные облака точек, дают подробную 3D-картину структуры леса (Данилин и др., 2005; Ковязин и др., 2020; Camarretta et al., 2021).
В ходе исследований по Международной биологической программе для трех климатических зон Сибири были предложены зависимости фитомассы крон сосны обыкновенной (Pinus sylvestris L.) от ширины кроны (Поздняков et al., 1969). За последние годы достигнуты существенные успехи в области обнаружения отдельных деревьев и регистрации их формы и структуры (диаметр кроны, высота дерева, площадь проекции и объем кроны) на основе новых высокопроизводительных алгоритмов и применения беспилотных летательных аппаратов (БПЛА, или дронов) (Ковязин и др., 2016; Усольцев, 2016). При этом диаметр кроны может обладать большей способностью объяснять изменчивость надземной фитомассы (10.5%) по сравнению с высотой дерева (6.0%) (Goodman et al., 2014).
Для нескольких видов лиственницы Китая (лиственницы сибирской (Larix sibirica Ledeb.), лиственницы Гмелина (L. gmelinii (Rupr.) Rupr.), лиственницы ольгинской (L. olgensis A. Henry.), лиственицы тонкочешуйной (L. kaempferi (Lamb.) Carr.), лиственицы принца Рупрехта (L. principis-rupprechtii Mayr.), лиственницы Мастерса (L. mastersiana Rehder & E.H. Wilson), лиственницы Потанина (L. himalaica W.C. Cheng et L.K. Fu) и лиственницы Гриффитца (L. speciosa W.C.Cheng & Y.W.Law)) были проанализированы аллометрические модели связи надземной и подземной фитомассы деревьев с диаметром ствола, а также с диаметром ствола и высотой дерева. Включение в аллометрические модели климатических показателей в качестве дополнительных независимых переменных существенно улучшило точность оценок и дало возможность прогнозирования фитомассы при изменении климата (Zeng et al., 2017; Fu et al., 2017). Однако такие модели разработаны без деления надземной фитомассы на компоненты и применимы лишь для территории Китая.
Березовые и осиновые леса повсеместно распространены в бореальной зоне и часто произрастают совместно с преобладанием того или другого вида. Береза и осина предпочитают разные экологические ниши, которые часто перекрываются. Работ, посвященных изучению растительности этих формаций, сравнительно немного (Дегтева и др., 2001; Попов, 2017). Лесостепная зона Западной Сибири представлена смешанными березово-осиновыми древостоями, в 90% площади которых преобладает береза и в 10% площади преобладает осина (Грибанов и др., 1970). Эти леса имеют характер небольших островков (колков) площадью от 0.2 до 30 га, которые занимают плоские блюдцеобразные впадины, сформировавшиеся вследствие оседания грунта под влиянием выщелачивания. По опушкам таких “блюдец” растет обычно береза повислая (Betula pendula Roth), которая ближе к центру сменяется березой пушистой (Betula pubescens Ehrh.), а центр “блюдца”, часто подверженный весеннему застойному увлажнению, бывает занят осиной (Усольцев, 1985). В последние годы происходит гибель колков на территории Западной Сибири вследствие переувлажнения (Володченкова, Гуц, 2011). Это явление циклически повторяется каждые 30–40 лет, но масштабы нынешней гибели колков беспрецедентны (Перепечина, 2009). Являются ли причины нынешней гибели колков антропогенными или климатическими и какое влияние это окажет на соотношение березы и осины в регионе, пока неизвестно.
В субарктическом регионе Швеции в течение последних десятилетий отмечена интенсивная экспансия осины обыкновенной (Populus tremula L.) в экотоне, в котором в течение последних 4 тыс. лет доминировала береза Черепанова (Betula pubescens Ehrh. ssp. czerepanovii (Orlova) Hämet-Ahti). В экспансии осины решающее значение имело поражение березы в результате вспышек травяной моли, вредоносному влиянию которой осина не подвержена. Предполагается, что при дальнейшем потеплении экспансия осины в субарктический экотон продолжится (Van Bogaert et al., 2009; 2010). При прогнозировании устойчивости и продуктивности популяций березы и осины в связи с изменением климата необходимо знать реакцию этих видов на стрессы, выраженную в том числе изменением структуры их фитомассы.
В данном исследовании мы намерены:
1) установить, имеются ли статистически значимые различия фитомассы деревьев березы и осины при оценке ее с помощью бортовой лидарной техники;
2) выяснить, как компоненты фитомассы соотносятся не только с таксационными показателями деревьев, но и с колебаниями температур и осадков на территории Евразии;
3) какой вклад в объяснение изменчивости компонентов фитомассы вносят таксационные показатели деревьев, видовая принадлежность и климатические переменные.
ОБЪЕКТЫ И МЕТОДИКА
Для решения поставленных задач мы использовали авторскую базу эмпирических данных о фитомассе лесообразующих пород Евразии (Усольцев, 2016). Из нее отобрано 755 и 296 модельных деревьев соответственно березы и осины с замеренными таксационными и биопродукционными показателями (табл. 1). Род Береза представлен в основном видами березы пушистой и березы повислой и в меньшей мере видами березы даурской (B. dahurica Pall.), березы ребристой (B. costata Trautv.), березы плосколистной (B. platyphylla Suk.). Род Осина представлен в основном видом осины обыкновенной и в меньшей мере видом тополя Давида (P. davidiana Dode). Обработка экспериментального материала выполнена по программе многофакторного регрессионного анализа Statgraphics-19 (http://www.statgraphics.com/).
Таблица 1.
Статистики модельных деревьев березы и осины
| Обозначение статистик* | Анализируемые показатели** | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| h | lcr | dcr | Ps | Pb | Pf | Pa | Pr | Pr/Pa | |
| Betula | |||||||||
| Mean | 15.6 | 7.2 | 2.9 | 98.6 | 16.2 | 2.5 | 117.3 | 47.6 | 0.35 |
| Min | 1.50 | 0.70 | 0.30 | 0.02 | 0.003 | 0.004 | 0.03 | 0.06 | 0.24 |
| Max | 27.8 | 18.1 | 13.4 | 984.8 | 267.2 | 29.5 | 1279.1 | 458.8 | 0.5 |
| SD | 6.4 | 3.2 | 1.9 | 130.6 | 29.8 | 3.5 | 159.7 | 105.0 | 0.066 |
| CV, % | 40.7 | 44.8 | 64.7 | 132.4 | 183.7 | 142.5 | 136.2 | 220.5 | 18.9 |
| n | 747.0 | 720.0 | 755.0 | 755.0 | 755.0 | 755.0 | 755.0 | 20.0 | 20.0 |
| Populus | |||||||||
| Mean | 15.4 | 6.3 | 3.1 | 102.3 | 22.4 | 3.5 | 128.2 | 18.3 | 0.19 |
| Min | 2.10 | 0.70 | 0.40 | 0.10 | 0.001 | 0.001 | 0.12 | 4.60 | 0.17 |
| Max | 28.8 | 15.0 | 11.0 | 955.0 | 354.4 | 38.6 | 1091.1 | 31.9 | 0.22 |
| SD | 6.6 | 2.9 | 1.9 | 161.9 | 48.9 | 6.0 | 210.6 | 19.3 | 0.037 |
| CV, % | 43.1 | 47.0 | 59.9 | 158.2 | 218.2 | 170.9 | 164.2 | 105.8 | 19.5 |
| n | 296.0 | 295.0 | 296.0 | 296.0 | 296.0 | 296.0 | 296.0 | 2.0 | 2.0 |
* Mean, Min и Max – соответственно среднее, минимальное и максимальное значения; SD – стандартное отклонение; CV – коэффициент вариации; n – число наблюдений. ** h – высота дерева, м; lcr – длина кроны, м; dcr – диаметр кроны, м; Ps, Pb, Pf, Pa, Pr – соответственно фитомасса ствола в коре, ветвей, листвы, надземной и корней в абсолютно сухом состоянии, кг.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Известно, что наиболее информативными независимыми переменными при оценке фитомассы деревьев путем дистанционного зондирования являются диаметр кроны и высота дерева. Принята структура двухфакторной аллометрической модели (Вейисов, Каплин, 1976; Усольцев, 2016):
(1)
$\ln {{P}_{i}} = {{a}_{0}} + {{a}_{1}}\left( {\ln {{d}_{{cr}}}} \right) + {{a}_{2}}\left( {\ln h} \right) + {{a}_{3}}X.$С целью установить различие березы и осины по фитомассе деревьев (или его отсутствие), в модели (1) в качестве дополнительной независимой переменной введена бинарная переменная Х (Freese, 1964), которой кодируются данные по березе (Х = 0) и осине (Х = 1). Результаты расчета моделей (1) показаны в табл. 2.
Бинарная переменная Х в моделях (1) значима на уровне p < 0.001, за исключением массы листвы, для которой Х значима на уровне p < 0.01. Она имеет знак “минус”, и это означает, что при одних и тех же значениях высоты дерева и диаметра кроны фитомасса всех надземных компонентов у осины существенно меньше, чем у березы, хотя по средним значениям экспериментальных данных фитомассы всех компонентов мы видим прямо противоположное соотношение (табл. 1).
По-видимому, в совокупности всех видов в пределах рода береза по сравнению с осиной имеет более узкую крону, а поскольку мы сравниваем их при равенстве как высоты дерева, так и диаметра кроны, то у деревьев березы концентрация фитомассы оказывается большей. Это предположение подтвердилось после расчета уравнения связи диаметра кроны с высотой дерева для берез и осин:
(2)
$\begin{gathered} \ln {{d}_{{cr}}} = - 1.4970 + 0.9359\left( {\ln h} \right) + 0.1136X; \\ adj{{R}^{2}} = 0.522;\,\,\,\,SE = 0.45. \\ \end{gathered} $Оказалось, что при одинаковой высоте дерева диаметр кроны у осины на 12% больше, чем у березы, и это различие статистически значимо (t = = 3.7 > t999 = 3.29).
Для ответа на второй вопрос нашего исследования имеющиеся данные географических координат модельных деревьев нанесены на карты средней январской температуры (https://store.mapsofworld.com/ image/cache/data/map_2014/currents-and-temperature- jan-enlarge-900x700.jpg) и среднегодовых осадков (http://www.mapmost.com/world-precipitation-map/ free-world-precipitation-map/) (World Weather Maps, 2007) (рис. 1 и 2) и одновременно совмещены с таксационными и биопродукционными показателями деревьев в одной общей матрице, которая включена затем в процедуру регрессионного анализа (http://www.statgraphics.com/). Построение моделей фитомассы деревьев на уровне родов, а не видов, а также отказ от использования средней годовой температуры в пользу средней температуры января обоснованы ранее (Усольцев, Цепордей, 2021а).
Рис. 1.
Распределение данных о фитомассе 755 модельных деревьев березы (красные кружки) и 296 деревьев осины (желтые кружки) на карте изотерм января, °C (World Weather Maps, 2007).
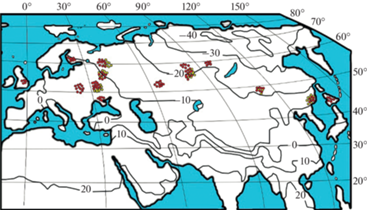
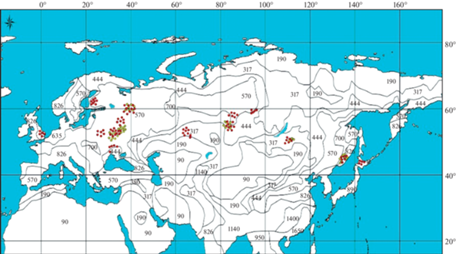
Рис. 2.
Распределение данных о фитомассе 755 модельных деревьев березы (красные кружки) и 296 деревьев осины (желтые кружки) на карте среднегодовых осадков, мм (World Weather Maps, 2007).
Предложена структура модели, включающая в качестве независимых переменных как морфометрические характеристики деревьев, так и климатические показатели:
(3)
$\begin{gathered} \ln {{P}_{i}} = {{a}_{0}} + {{a}_{1}}\left( {\ln {{d}_{{cr}}}} \right) + {{a}_{2}}\left( {\ln h} \right) + {{a}_{3}}X + \\ + \,\,{{a}_{4}}\left[ {\ln \left( {T + 30} \right)} \right] + {{a}_{5}}\left( {\ln {\kern 1pt} PR} \right) + \\ + \,\,{{a}_{6}}\left[ {\ln \left( {T + 30} \right)} \right]\left( {\ln PR} \right), \\ \end{gathered} $Как следует из табл. 1, количество фактических данных о фитомассе корней березы и осины соответственно в 38 и 150 раз меньше по сравнению с данными о надземной фитомассе, и это объясняется несопоставимо большей трудоемкостью получения фактических данных о фитомассе корней. По причине недостаточной представленности данных о фитомассе корней, мы рассчитали модели (3) для относительного показателя, а именно – для Pr/Pa, и связали оба вида в одной общей модели путем кодирования их бинарной переменной Х. Результаты расчета моделей (3) показаны в табл. 3.
Таблица 3.
Результаты расчета моделей (3)
| ln(Y) | а0 | ln dcr | ln h | X | ln(T + 30) | lnPR | [ln(T + 30)] × × (lnPR) | adjR2 | SE |
|---|---|---|---|---|---|---|---|---|---|
| ln(Ps) | 32.8198 | 0.7613 | 2.8258 | –0.1869 | –13.1784 | –6.1190 | 2.1487 | 0.954 | 0.40 |
| ln(Pf) | 9.0958 | 1.3389 | 1.3102 | –0.1055 | –2.5647(*) | –2.3434(*) | 0.4775(*) | 0.776 | 0.75 |
| ln(Pb) | 45.9217 | 1.6241 | 2.0017 | –0.1972 | –17.6957 | –8.4270 | 2.9153 | 0.865 | 0.75 |
| ln(Pa) | 33.5231 | 0.8848 | 2.6453 | –0.1843 | –13.1757 | –6.1667 | 2.1562 | 0.947 | 0.42 |
| ln(Pr/Pa) | 31.5608 | – | – | – | –13.0107 | –5.1040 | 2.0173 | 0.336 | 0.20 |
Все регрессионные коэффициенты моделей, представленных в табл. 3, достоверны на уровне p < 0.001, за исключением двух, помеченных как (*) для фитомассы листвы. В модели для Pr/Pa оказались значимыми лишь климатические переменные.
Геометрическая интерпретация моделей (3) (рис. 3) для фитомассы березы получена путем подстановки в (3) среднего значения h для березы, равного 15.6 м (табл. 1) и расчетных значений dcr, равных для березы и осины соответственно 2.93 и 3.28 м, рассчитанных по модели (2).
Рис. 3.
Связь компонентов фитомассы деревьев березы с средней температурой января (T) и среднегодовыми осадками (PR). Обозначения см. в табл. 1.
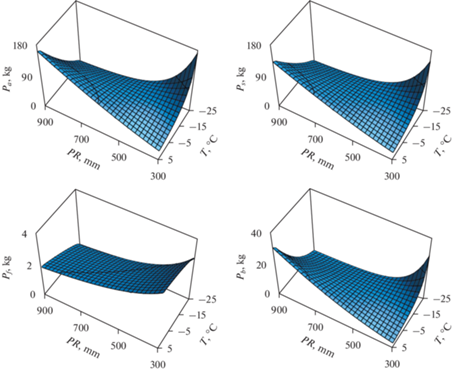
Согласно рис. 3, зависимость всех компонентов фитомассы, за исключением листвы, от температур и осадков описывается пропеллеро-образной 3D-поверхностью. В холодных регионах при повышении осадков фитомасса снижается, но по мере перехода к теплым регионам она характеризуется противоположным трендом. При повышении температуры во влажных регионах фитомасса увеличивается, но по мере перехода в сухие условия –снижается. Масса листвы возрастает по мере перехода от влажных к влагодефицитным регионам, независимо от температурного режима, и несколько снижается в направлении холодных регионов, но только при достаточном влагообеспечении. Для осины рис. 3 повторяется, но 3D-поверхности для фитомассы листвы, ветвей, стволов и надземной сдвинуты вниз по оси ординат согласно моделям (3) соответственно на 4.7, 1.3, 9.6 и 8.0%, и эти сдвиги статистически значимы.
У других древесных видов, в частности, ели (Усольцев, Цепордей, 2021б), закономерности несколько отличаются по некоторым компонентам от представленных на рис. 3. По-видимому, это связано с биологическими свойствами древесных видов и с особенностями распределения ассимилятов дерева в различные его компоненты (Palumets, 1990).
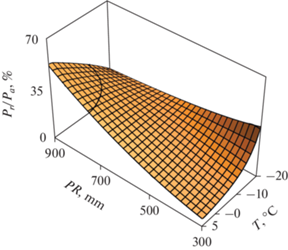
Изменчивость отношения (Pr/Pa) объясняется независимыми переменными в наименьшей степени (34%) по сравнению с компонентами надземной фитомассы (78–95%), и различие (Pr/Pa) для березы и осины недостоверно. Графическая интерпретация этой зависимости показана на рис. 4, в котором мы можем видеть, что отношение Pr/Pa для деревьев березы и осины изменяется в градиентах температур и осадков аналогично изменениям фитомассы стволов, ветвей и надземной.
Рис. 4.
Связь относительного показателя фитомассы корней березы и осины (Pr/Pa) с средней температурой января (T) и среднегодовыми осадками (PR).
Результаты многофакторного регрессионного анализа дали ответ на третий вопрос нашего исследования (табл. 4). Мы видим в табл. 4, что средние значения вкладов таксационных показателей деревьев, видовой принадлежности и климатических переменных в объяснение изменчивости компонентов фитомассы составляют соответственно 79.0, 4.6 и 16.4%. Климатические переменные в наибольшей степени объясняют изменчивость фитомассы стволов, ветвей и надземной (от 18 до 22%) и в наименьшей степени – листвы (около 7%).
Таблица 4.
Вклад независимых переменных уравнений (3) в объяснение изменчивости зависимых переменных, %
| ln(Y) | Независимые переменные | |||||||
|---|---|---|---|---|---|---|---|---|
| ln dcr (I) |
ln h (II) |
I + II | Х | ln(T + 30) (III) | lnPR (IV) |
[ln(T + 30)] × × (lnPR) (V) | III + IV + V | |
| ln(Ps) | 19.7 | 57.2 | 76.9 | 4.9 | 6.0 | 6.1 | 6.1 | 18.2 |
| ln(Pf) | 50.4 | 38.5 | 88.9 | 4.0 | 1.7 | 3.4 | 2.0 | 7.1 |
| ln(Pb) | 37.4 | 36.0 | 73.4 | 4.6 | 7.2 | 7.5 | 7.3 | 22.0 |
| ln(Pa) | 19.7 | 57.2 | 76.9 | 4.9 | 6.0 | 6.1 | 6.1 | 18.2 |
| М ± σ(а) | 31.8 ± 14.9 | 47.2 ± 11.6 | 79.0 ± 6.8 | 4.6 ± 0.4 | 5.2 ± 2.4 | 5.8 ± 1.7 | 5.4 ± 2.3 | 16.4 ± 6.4 |
Поскольку деревья разных древесных пород обладают свойственной им конфигурацией вертикального профиля, эта видовая специфика сегодня успешно распознается с помощью бортовых лазерных сенсоров (Puttonen et al., 2011; Åkerblom et al., 2017).
ВЫВОДЫ
1. На основе авторской базы данных о фитомассе березы и осины Евразии установлено, что масса листвы, ветвей, стволов и надземная у деревьев березы равного диаметра кроны и равной высоты дерева соответственно на 4.7, 1.3, 9.6 и 8.0% больше, чем у осины. Это вызвано тем, что при одной и той же высоте дерева диаметр кроны у осины на 12% больше, чем у березы.
2. Введение в аллометрическую модель температуры и осадков в качестве дополнительных независимых переменных показало, что фитомасса равновеликих деревьев описывается пропеллеро-образной 3D-зависимостью. В холодных регионах при повышении осадков фитомасса снижается, но по мере перехода к теплым регионам характеризуется противоположным или нейтральным трендом. При повышении температуры во влажных регионах фитомасса увеличивается, но по мере перехода в сухие условия постепенно снижается.
3. Средние значения вклада таксационных показателей деревьев, их видовой принадлежности и климатических переменных в объяснение изменчивости компонентов фитомассы составляют соответственно 79.0, 4.6 и 16.4%.
Полученные результаты могут быть полезны при мониторинге фитомассы лесов на основе лазерного зондирования.
Список литературы
Вейисов С., Каплин В.Г. К методике определения надземной фитомассы белого саксаула в Восточных Каракумах // Проблемы освоения пустынь. 1976. № 1. С. 60–64.
Володченкова Л.А., Гуц А.К. Математическое моделирование стадий вымокания березовых лесов с помощью теории катастроф // Математические структуры и моделирование. 2011. № 24. С. 19–33.
Грибанов Л.Н., Лагов И.А., Чабан П.С. Леса Казахстана // Леса СССР. Т. 5. М.: Наука, 1970. С. 5–77.
Данилин И.М., Медведев Е.М., Мельников С.Р. Лазерная локация земли и леса: учебное пособие. Красноярск: Ин-т леса СО РАН, 2005. 182 с.
Дегтева С.В., Железнова Г.В., Пыстина Т.Н., Шубина Т.П. Ценотическая и флористическая структура лиственных лесов европейского Севера. Спб.: Наука, 2001. 269 с.
Ковязин В.Ф., Богданов В.Л., Гарманов В.В., Осипов А.Г. Мониторинг зеленых насаждений с применением беспилотных летательных аппаратов // Аграрный научный журн. Естественные, технические и экономические науки. 2016. № 4. С. 14–19.
Ковязин В.Ф., Виноградов К.П., Киценко А.А., Васильева Е.А. Воздушное лазерное сканирование для уточнения таксационных характеристик древостоев // Известия вузов. Лесной журн. 2020. № 6. С. 42–54.
Перепечина Ю.И. Влияние периодического переувлажнения на состояние лесов Курганской области // Актуальные проблемы лесного комплекса. 2009. Т. 23. С. 54–56. https://www.elibrary.ru/contents.asp?issueid= 1357980
Поздняков Л.К., Протопопов В.В., Горбатенко В.М. Биологическая продуктивность лесов Средней Сибири и Якутии. Красноярск: Книжное изд-во, 1969. 120 с.
Попов С.Ю. Растительность березовых и осиновых лесов Пинежского заповедника // Заповедная наука. 2017. Т. 2. № 2. С. 66–83.
Усольцев В.А. Моделирование структуры и динамики фитомассы древостоев. Красноярск: Изд-во Красноярского ун-та, 1985. 191 с. http://elar.usfeu.ru/handle/ 123456789/3353
Усольцев В.А. Фитомасса модельных деревьев лесообразующих пород Евразии: база данных, климатически обусловленная география, таксационные нормативы. Екатеринбург: Уральский государственный лесотехнический университет, 2016. 336 с. http://elar.usfeu. ru/handle/123456789/5696
Усольцев В.А., Цепордей И.С. Прогнозирование биомассы стволов сосновых деревьев естественных древостоев и лесных культур в связи с изменением климата // Сибирский лесной журн. 2021 а. № 2. С. 72–81.
Усольцев В.А., Цепордей И.С. Принцип пространственно-временнóго замещения в экологии и прогнозирование биомассы Picea spp. при изменении климата // Хвойные бореальной зоны. 2021 б. Т. 39. № 4. С. 269–275.
Åkerblom M., Raumonen P., Mäkipää R., Kaasalainen M. Automatic tree species recognition with quantitative structure models // Remote Sensing of Environment. 2017. V. 191. P. 1–12. https://doi.org/10.1016/j.rse.2016.12.002
Camarretta N., Harrison P.A., Lucieer A., Potts B.M., Davidson N., Hunt M. Handheld laser scanning detects spatiotemporal differences in the development of structural traits among species in restoration plantings // Remote Sensing. 2021. V. 13. № 9. Article 1706. https://doi.org/10.3390/rs13091706
Díaz S., Settele J., Brondızio E.S., Ngo H.T., Agard J., Arneth A., Balvanera P., Brauman K.A, Butchart S.H.M., Chan K.M.A., Lucas A.G., Ichii K., Liu J., Subramanian S.M., Midgley G.F., Miloslavich P., Molnár Z., Obura D., Pfaff A., Polasky S., Purvis A., Razzaque J., Reyers B., Chowdhury R.R., Shin Y.J., Visseren-Hamakers I., Willis K.J., Zayas C.N. Pervasive human-driven decline of life on Earth points to the need for transformative change // Science. 2019. V. 366. Article 1327.
Franklin O., Harrison S.P., Dewar R., Farrior C.E., Brännström Å., Dieckmann U., Pietsch S., Falster D., Cramer W., Loreau M., Wang H., Mäkelä A., Rebel K.T., Meron E., Schymanski S.J., Rovenskaya E., Stocker B.D., Zaehle S., Manzoni S., van Oijen M., Wright I.J., Ciais P., van Bodegom P.M., Peñuelas J., Hofhansl F., Terrer C., Soudzilovskaia N.A., Midgley G., Prentice I.C. Organizing principles for vegetation dynamics // Nature Plants. 2020. V. 6. P. 444–453.
Freese F. Linear regression methods for forest research. USDA Forest Service. Res. Paper FPL 17. Madison, 1964. 136 p.
Fu L., Sun W., Wang G. A climate-sensitive aboveground biomass model for three larch species in northeastern and northern China // Trees. 2017. V. 31. P. 557–573.
Goodman R.C., Phillips O.L., Baker T.R. The importance of crown dimensions to improve tropical tree biomass estimates // Ecological Applications. 2014. V. 24. № 4. P. 680–698. https://doi.org/10.1890/13-0070.1
Madgwick H.A.I. Above-ground weight of forest plots – comparison of seven methods of estimation // New Zealand J. Forestry Science. 1983. V. 13. P. 100–107.
Palumets Ya.K. Modeling of the distribution of spruce phytomass // Soviet Forest Sciences. 1990. № 3. P. 36–40 (Allerton Press, Inc.).
Puttonen E., Jaakkola A., Litkey P., Hyyppä J. Tree classification with fused mobile laser scanning and hyperspectral data // Sensors. 2011. V. 11. № 5. P. 5158–5182. https://doi.org/10.3390/s110505158
Reichstein M., Bahn M., Mahecha M.D., Kattge J., Baldocchi D.D. Linking plant and ecosystem functional biogeography // Proceedings of the National Academy of Sciences, USA. 2014. V. 111. Article 201216065.
Tautenhahn S., Migliavacca M., Kattge J. News on intra-specific trait variation, species sorting, and optimality theory for functional biogeography and beyond // New Phytologist. 2020. V. 228. P. 6–10.
Van Bogaert R., Jonasson C., De Dapper M., Callaghan T.V. Range expansion of thermophilic aspen (Populus tremula L.) in the Swedish Subarctic // Arctic, Antarctic, and Alpine Research. 2010. V. 42. P. 362–375. https://doi.org/10.1657/1938-4246-42.3.362
Van Bogaert R., Jonasson C., De Dapper M., Callaghan T.V. Competitive interaction between aspen and birch moderated by invertebrate and vertebrate herbivores and climate warming // Plant Ecology & Diversity. 2009. V. 2. № 3. P. 221–232. https://doi.org/10.1080/17550870903487456
World Weather Maps, 2007. Available at: https://www.mapsofworld.com/referrals/weather
Zeng W.S., Duo H.R., Lei X.D., Chen X.Y., Wang X.J., Pu Y., Zou W.T. Individual tree biomass equations and growth models sensitive to climate variables for Larix spp. in China // European J. Forest Research. 2017. V. 136. P. 233–249.
Дополнительные материалы отсутствуют.
Инструменты
Лесоведение