

Лесоведение, 2022, № 6, стр. 631-642
Биологическое разнообразие коренных типов леса в заповедных лесных участках Московского региона
А. А. Маслов *
Институт лесоведения РАН
143030 Московская обл., п/о Успенское, ул. Советская, д. 21, Россия
* E-mail: amaslov@ilan.ras.ru
Поступила в редакцию 14.03.2022
После доработки 11.05.2022
Принята к публикации 07.06.2022
- EDN: KSYPOF
- DOI: 10.31857/S0024114822060067
Аннотация
В результате маршрутных обследований заповедных лесных участков (лесных резерватов) Московского региона (зона хвойно-широколиственных лесов) получен массив геоботанических описаний в коренных типах леса, включая ельники, сосняки и широколиственные леса. Данные описания (применительно к региону) можно рассматривать в качестве “эталонных” при сравнении с молодыми, вторичными и антропогенно сильно нарушенными лесами. Цель работы – определить показатели видового разнообразия для ненарушенных (рубками, рекреацией) коренных лесных сообществ в основных типах леса на территории Московского региона. Разделение описаний на группы (типы леса) проведено в соответствии с принципами школы В.Н. Сукачева (на основе признаков местообитания и состава диагностических групп видов) с уточнением результатов путем ординации описаний в многомерных осях DCA. Впервые для коренных типов леса Московского региона по единой методике рассчитаны показатели биоразнообразия (для площади описаний 300 м2): видовая насыщенность, индекс Шеннона, построены графики доминирования–разнообразия. Максимальная видовая насыщенность древесного яруса (A) наблюдается в сложных типах леса – сосняке (4.9) и ельнике (4.4), яруса кустарников (B2) – в ельниках сложном (6.1), кисличном (5.3), травяно-кустарничкового яруса (C) – в неморальных ельниках (более 29), яруса D (мохово-лишайникового) – в сосняке брусничном (7.6). В целом для типа леса (все ярусы) максимальная видовая насыщенность обнаружена в неморальных типах ельников – кисличном (43.0) и сложном (42.7). В этих же типах леса наблюдается максимальное видовое разнообразие по индексу Шеннона (2.80 и 2.82 соответственно), а форма кривых доминирования–разнообразия видов нижних ярусов (C + D) имеет минимальный наклон и близка к S-образной. Такая форма кривых характерна для зрелых природных сообществ.
Расположенный в зоне хвойно-широколиственных лесов Московский регион11 интересен для изучения биологического разнообразия тем, что на сравнительно небольшой равнинной территории в экотоне между подзоной южной тайги и зоной широколиственных лесов (Курнаев, 1982; Hytteborn et al., 2005) представлены самые разные типы ландшафтов и типы леса – от бореальных ельников до типичных дубрав.
Изучению лесов региона посвящено достаточно много исследований. На видовом (флористическом) уровне вопрос был рассмотрен в сводке Л.П. Рысина “Конспект лесной флоры …” (2009). На ценотическом уровне первой работой стала статья Н.А. Коновалова (1929) о типах леса подмосковных лесничеств. Разнообразие типов леса было рассмотрено в монографии С.Ф. Курнаева (1968) и в серии монографий “Леса Восточного Подмосковья” (1979), “Леса Западного Подмосковья” (1982), “Леса Южного Подмосковья” (1985), “Леса Северного Подмосковья” (1993) и в книге “Леса Москвы …” (2001). Все эти (и другие) публикации были обобщены в сводке Л.П. Рысина “Леса Подмосковья” (2012).
На первый взгляд кажется, что вопросы биоразнообразия лесов региона изучены очень подробно. Однако внимательное знакомство с “классическими” работами позволяет выявить ряд проблем. Почти все описания типов леса относятся к “ближнему” Подмосковью (территории бывшего лесопаркового пояса столицы), леса которого сильно нарушены рекреацией, рубками, загрязнением; размер площадок геоботанических описаний, как правило, не указан, или он может быть очень велик (до 0.5–1 га), а отсутствие полноценных фитоценотических таблиц делает невозможным расчет показателей видового богатства. Таким образом, для, казалось бы, хорошо изученных лесов Московского региона сохраняется объективная потребность в количественной оценке биоразнообразия типов леса на площадках стандартизированного размера и по единой (или хотя бы сопоставимой) методике – аналогично тому, как это сделано для лесных сообществ Великобритании (Rodwell, 1991).
Первые попытки поставить анализ биологического разнообразия лесов Московского региона на количественную основу были предприняты в сборнике “Оценка и сохранение …” (2000), где приводятся данные по видовому богатству и видовой насыщенности для леспаркхоза “Горки” и Приокско-Террасного заповедника. Дальнейшее развитие эти подходы получили в серии статей, где параметры описания лесных сообществ соответствуют стандартам проекта “European vegetation survey” (Rodwell et al., 1997) и базе данных “European vegetation archive” (Сhytrý et al., 2016). Эти работы посвящены как отдельным районам Московского региона (Черненькова и др., 2018) или отдельным типам сообществ (Морозова и др., 2021), так и лесам Московской области в целом (Черненькова и др., 2020). В последней работе оценки средней видовой насыщенности (числа видов на единицу площади) даны для каждого синтаксона.
Переход к количественной оценке видового богатства делает актуальным вопрос о сравнении получаемых результатов между собой и с “эталонами” коренных заповедных сообществ (Рысин, Савельева, 1980), поскольку в малонарушенных заповедных сообществах биоразнообразие не является максимальным (Guo, 2005). О такой особенности коренных лесов упоминалось еще в классической монографии Э. Мэгарран (1992) на примере десяти лесных массивов в Северной Ирландии.
О том, какие именно типы леса можно считать “коренными”, в российской литературе имеются разные точки зрения. В данной работе мы соблюдаем традиции московской школы геоботаников, согласно которым к коренным типам леса принято относить сообщества, где (по эмпирическим представлениям авторов) древостой и нижние ярусы соответствуют типу условий произрастания при отсутствии явной сукцессионной динамики. При этом сообщества коренного типа могут иметь одновозрастный древостой и не являются “климаксом”22. В число коренных типов леса Московского региона традиционно включают не только ельники, липняки и дубравы, но и различные типы сосновых лесов на относительно бедных почвах (Алехин, 1947; Курнаев, 1968; Рысин, 1979; Ильинская и др., 1982; Рекомендации …, 1982; Огуреева и др., 1996).
Цель настоящей работы – определить показатели видового разнообразия для ненарушенных (рубками, рекреацией) коренных лесных сообществ в основных типах леса на территории заповедных лесных участков Московского региона. В задачи работы входили: выполнение лесоводственно-геоботанических описаний, классификация (ординация) описаний для уточнения принадлежности к типу леса, отбор эталонных описаний, определение видовой насыщенности, индекса Шеннона и построение кривых доминирования–разнообразия.
ОБЪЕКТЫ И МЕТОДИКА
Исследования проводили на территории заповедных лесных участков (лесных резерватов) Московского региона (Рысин, Савельева, 1985; Маслов, 2009). Все резерваты находятся в “дальнем Подмосковье” – за пределами бывшего лесопаркового пояса, при этом часть резерватов теперь расположена в Новой Москве. Хорошо представлены все физико-географические районы, за исключением лесостепного Заокского (рис. 1). Лесоводственно-геоботанические описания были выполнены в ходе маршрутных исследований 1980–1992 гг. Таким образом, в полевых материалах зафиксировано состояние лесов Московского региона на конец XX в., что представляет (сейчас) и исторический интерес, имеется в виду, что бóльшая часть старовозрастных ельников существенно пострадала или совсем исчезла в результате катастрофической вспышки короеда типографа в 2011–2013 гг.
Рис. 1.
Объекты исследований – сеть резерватов (заповедных лесных участков) Московской обл. (по состоянию на 2000 г.). Точками обозначены номера резерватов. Красная линия – границы области, желтые линии – границы физико-географических районов по Е.Д. Смирновой (Московская область, 1979): I. Верхневолжская низменность; II. Смоленско-Московская возвышенность; III. Москворецко-Окская равнина; IV. Мещерская низменность; V. Заокское плато.
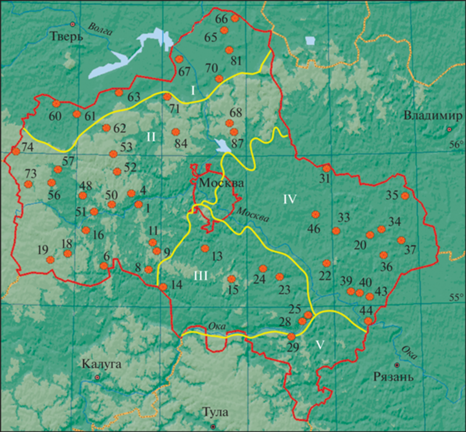
Планирование маршрутов по территории резерватов для выполнения описаний осуществляли по картам и материалам лесоустройства с данными о составе и возрасте насаждений. Первоочередное внимание уделяли участкам естественных ненарушенных (рекреацией, рубками, выпасом) старовозрастных (старше 100 лет) насаждений из коренных пород деревьев (ель, сосна, липа, дуб). При отсутствии таких насаждений описания делали в средневозрастных коренных лесах. Еще более низкий приоритет имели старые насаждения из вторичных пород (осина, береза); в таких сообществах описания проводили лишь при отсутствии в резервате коренных насаждений. Дополнительными критериями при выборе места для описания были полнота насаждения и отсутствие свежих естественных нарушений, таких как ветровальные комплексы, усыхания или порои. Однако в местах находок редких и краснокнижных видов растений описания делали независимо от характера древостоя и/или нарушенности.
Геоботанические описания выполняли путем закладки круговых площадок размером 300–400 м2, что соответствует практике описаний в лесах Европы (Chytrý, Otýpková, 2003). Учет видового состава проводили отдельно по ярусам: A – древостой, B1 – подрост (деревья с диаметром <6 см), B2 – подлесок, C – травы и кустарнички, D – мхи и лишайники (только напочвенные). Для каждого вида указывали обилие по шкале Браун-Бланке. Образцы трудных для определения видов брали в гербарий33. Дополнительно фиксировали параметры местообитания, а также (в ряде случаев) закладывали прикопки для описания почвы. Месторасположение описаний привязывали к карте масштаба 1 : 10 000 и наносили на нее. Всего в ходе маршрутных исследований было сделано 205 описаний в ельниках, сосняках, хвойно-широколиственных и широколиственных лесах.
Каждое описание с учетом комплекса признаков местообитания и растительности предварительно относили к определенному типу леса. Классификационная схема базировалась на принципах школы В.Н. Сукачева (Рысин, 1982), а также “Рекомендациях по выделению коренных и производных групп типов леса…” (1982). Тип леса в последней работе понимается относительно широко и включает восемь синтаксонов: сосняк лишайниковый, сосняк брусничный, сосняк черничный, сосняк сложный, ельник черничный, ельник кисличный, ельник сложный, дубняк липовый44. Материалы по болотным лесам не включены в настоящую публикацию в связи со значительной их спецификой.
По результатам маршрутных обследований лесных заповедных участков в нескольких типах леса были заложены постоянные пробные площади (ППП). Программа работ на ППП опубликована ранее (Маслов, 1990). На каждой ППП учеты видов нижних ярусов проводили на постоянных трансектах из нескольких тысяч площадок размером 20 × 20 см, что позволяет детально оценивать значимость видов и строить кривые доминирования-разнообразия.
Для обработки и хранения данных по растительности и параметрам местообитания в качестве оболочки применяли специализированную базу данных TurboVeg for Windows (Hennekens, Schaminee, 2001). Данная база уже давно фактически стала стандартом для работ с геоботаническими массивами данных, в частности, в рамках проекта “European Vegetation Survey” (Schaminée et al., 2009).
Количественная обработка описаний выполнена в пакетах PC-ORD (McCune, Mefford, 2016) и в TurboVeg for Windows. Важнейшей задачей было формирование выборки “эталонных” описаний коренных типов леса; при этом отбор “эталонов” включал два этапа: 1) проверку (корректировку) принадлежности каждого описания к типу леса;
2) улучшение качества выборки описаний в рамках отдельного типа леса.
На первом этапе методом DCA (детрендового анализа соответствий) было осуществлено упорядочивание описаний вдоль многомерных осей максимального варьирования (Jongman et. al., 1995). При проведении DCA, в соответствии с рекомендациями разработчиков, из общей сводной таблицы были исключены самые редкие виды (встреченные меньше, чем в 10 описаниях), для остальных редких видов применяли опцию взвешивания. Ординацию выполняли с учетом баллов обилия видов, конвертированных в % покрытия по стандартной шкале TurboVeg.
Корректировку признака “тип леса” для конкретного описания проводили по результатам ординации, при этом за координаты описания в пространстве ординации принимались его нагрузки на первые две оси. Положение описаний относительно осей зависит от сходства их видового состава. Чем более сходен видовой состав, тем ближе расположены в пространстве ординации, соответствующие описаниям точки. Выделив группы близких (в пространстве осей) описаний, можно уточнить тип леса, перейдя, таким образом, от процедуры ординации сообществ к процедуре классификации. Часть описаний, далеко отстоящих в пространстве ординации от коренных типов леса, была отбракована на этом этапе. Дополнительно при определении принадлежности описания к конкретному типу леса использовали признаки местообитания и состав диагностических фитоцено-экологических групп видов (Маслов, 1998; Федорчук и др., 2002). Таким образом, включение описания, например, в тип леса “сосняк брусничный” не обязательно означает доминирование брусники в данном конкретном сообществе – в отличие от доминантного подхода, примененного, в частности, в монографии “Сосновые леса России” (Рысин, Савельева, 2008).
На втором этапе формирование выборки для каждого типа леса осуществляли путем ручной отбраковки описаний, в меньшей степени соответствующих критериям “эталонов” коренных ненарушенных насаждений. Отбракованы описания с потенциально неполным списком видов (сделанные ранней весной или поздней осенью) и описания с “выбросами” по числу видов. Описания из последней группы анализировали вручную на предмет флористического списка и исключали из обработки при “излишнем” количестве сорных и/или лугово-лесных видов (свидетельство давнего выпаса). На последнем шаге селекции для каждого типа леса были отобраны описания с бóльшим возрастом древостоя и полнотой не менее 0.6. В ельниках и дубравах после селекции преобладающий возраст древостоев в описаниях составил 90–100 лет, а в сосняках – 70–80 лет. Всего по итогам селекции было исключено 25% описаний, и окончательный размер выборки для расчетов видового разнообразия составил 151 описание.
Показатели видового разнообразия определяли для каждого описания и каждого типа леса. Для описания выявляли видовую насыщенность (число видов) по ярусам и в целом для описания; видовую насыщенность в типе леса определяли как среднее из всех описаний типа. В качестве меры разнообразия был выбран индекс Шеннона, наиболее широко используемый в литературе (Мэгарран, 1992; Ханина и др., 2016). При расчете индекса ярусы A и B1 объединяли в один во избежание двойного учета одних и тех же древесных видов.
Ранговые кривые доминирования–разнообразия построены для отдельных ППП по данным учета встречаемости видов нижних ярусов (C и D) на площадках размером 0.2 × 0.2 м. Количество площадок на каждой из ППП составляет от 2000 до 2800.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
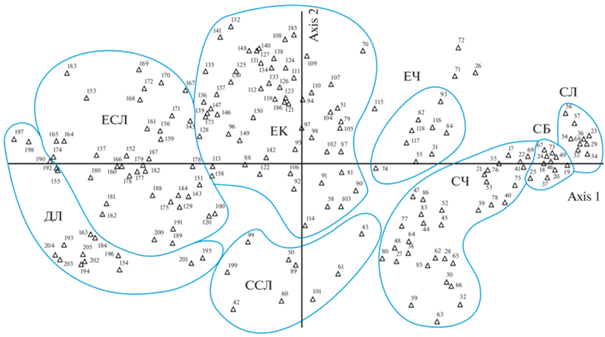
На рис. 2 представлены результаты ординации описаний незаболоченных заповедных лесов методом детрендового анализа соответствий (DCA). Группы точек, соответствующие предварительно выделенным типам леса, на диаграмме обведены, при этом диалектическое единство дискретности и континуальности растительного покрова наглядно видно из графика. Так, плавные переходы в пространстве ординации наблюдаются между ельниками кисличными и ельниками сложными, а также между бореальными сосняками – лишайниковыми, брусничными, черничными. В то же время собственно бореальные сосняки и бореальные ельники достаточно изолированы в пространстве ординации от неморальных типов леса. По результатам ординации предварительные названия типов леса для ряда описаний на границах между типами были изменены, а часть описаний, далеко “улетающих” за пределы групп, исключена из дальнейшей обработки.
Рис. 2.
Ординация описаний в незаболоченных коренных типах леса Московского региона методом детрендового анализа соответствий (DCA). Обозначения типов леса: СЛ – сосняк лишайниковый, СБ – сосняк брусничный, СЧ – сосняк черничный, ССЛ – сосняк сложный, ЕЧ – ельник черничный, ЕК – ельник кисличный, ЕСЛ – ельник сложный, ДЛ – дубняк липовый. Цифрами обозначены номера описаний; часть полностью налегающих друг на друга описаний на диаграмме опущена.
Показатели видового разнообразия коренных типов леса даны в табл. 1. Число описаний по типам существенно отличается. Максимальным числом описаний (46) в таблице представлены ельники кисличные, минимальным (8) – сосняки лишайниковые. Доля каждого типа леса в общем количестве описаний отражает примерно встречаемость коренных насаждений данного типа в пределах Московского региона. В колонке “число видов” дано количество видов во флористическом списке (ненормированное по числу описаний).
Таблица 1.
Видовое разнообразие коренных типов леса Московского региона
| Тип леса | N | Число видов | Видовая насыщенность по ярусам и общая | H | |||||
|---|---|---|---|---|---|---|---|---|---|
| A | B1 | B2 | C | D | общая | ||||
| СЛ | 8 | 69 | 1.0 ± 0.0 | 1.8 ± 0.3 | 4.1 ± 0.4 | 19.4 ± 1.7 | 6.5 ± 0.7 | 32.1 ± 1.9 | 2.26 |
| СБ | 11 | 88 | 2.5 ± 0.2 | 2.8 ± 0.3 | 2.4 ± 0.5 | 21.2 ± 2.5 | 7.6 ± 0.8 | 34.6 ± 2.5 | 2.42 |
| СЧ | 21 | 99 | 2.8 ± 0.2 | 2.5 ± 0.3 | 4.1 ± 0.2 | 20.2 ± 1.3 | 4.8 ± 0.4 | 32.8 ± 1.3 | 2.54 |
| ССЛ | 11 | 97 | 4.9 ± 0.2 | 4.1 ± 0.3 | 4.5 ± 0.6 | 24.7 ± 1.3 | 2.7 ± 0.6 | 37.9 ± 1.9 | 2.72 |
| ЕЧ | 9 | 73 | 3.2 ± 0.2 | 2.6 ± 0.5 | 3.1 ± 0.3 | 19.3 ± 2.2 | 6.0 ± 0.5 | 32.7 ± 2.1 | 2.61 |
| ЕК | 46 | 172 | 3.3 ± 0.1 | 2.4 ± 0.2 | 5.3 ± 0.3 | 29.7 ± 0.9 | 3.8 ± 0.3 | 43.0 ± 1.0 | 2.80 |
| ЕСЛ | 35 | 143 | 4.4 ± 0.3 | 3.4 ± 0.2 | 6.1 ± 0.3 | 29.1 ± 1.1 | 2.0 ± 0.2 | 42.7 ± 1.4 | 2.82 |
| ДЛ | 10 | 87 | 3.8 ± 0.3 | 3.0 ± 0.4 | 4.3 ± 0.6 | 22.5 ± 0.8 | 0.3 ± 0.2 | 32.1 ± 1.1 | 2.53 |
Обозначения: N – число описаний размером 300(400) м2, A – древостой, B1 – подрост, B2 – кустарниковый ярус, C – травяно-кустарничковый ярус, D – мохово-лишайниковый ярус, H – индекс Шеннона (при расчете объединялись ярусы A, B1). Обозначения типов леса см. на рис. 2. Для видовой насыщенности приводится среднее значение и его стандартная ошибка.
Видовая насыщенность
Максимальная видовая насыщенность древесного яруса (A) наблюдается в сложных типах леса – сосняке (4.9) и ельнике (4.4), что вполне ожидаемо, исходя из определения данных типов. Минимальная видовая насыщенность (1.0) присутствует в типе леса сосняк лишайниковый, где, кроме сосны, в древостое нет других видов. Почти в том же порядке располагаются типы леса и по богатству видов подроста (B1), при этом в сложных типах леса видовая насыщенность подроста несколько меньше, чем насыщенность древостоя, а в сосняке лишайниковом – больше, так как, кроме сосны, в подросте встречаются дуб и береза. По богатству видов кустарников (B2) порядок расположения типов леса не совпадает с порядком расположения по богатству древесного яруса. Самый разнообразный подлесок мы наблюдаем в ельнике сложном (6.1), но на втором месте – ельник кисличный (5.3). Минимальная видовая насыщенность видов кустарников – в сосняке брусничном (2.4). Большее число видов кустарников в сосняке лишайниковом (4.1) связано с тем, что сообщества данного типа чаще встречаются в южных районах области и имеют в своем составе элементы флоры лесостепей (ракитник русский (Chamaecytisus ruthenicus (Fisch. ex Woloszcz.) Klásk.) и дрок красильный (Genista tinctoria L.)).
Видовая насыщенность травяно-кустарникового яруса (C) наибольших значений (более 29) достигает в неморальных типах ельников – кисличном и сложном. Наименьшее число видов (в интервале от 19 до 21) наблюдается в четырех бореальных типах леса – сосняке лишайниковом, черничном, брусничном и в ельнике черничном. Что касается мохово-лишайникового яруса (D), то для него – картина обратная. Наибольшая видовая насыщенность обнаружена в сосняке брусничном (7.6), сосняке лишайниковом (6.6), ельнике черничном (6.0), наименьшая – в дубо-липняке (0.3) и ельнике сложном (2.0).
Показатели видовой насыщенности в целом для типа леса даны в предпоследней колонке табл. 1. При этом общая видовая насыщенность не является в точности суммой оценок по отдельным ярусам, так как ярусы древостоя (A) и подроста (B1) частично включают одни и те же древесные виды. Как оказалось, максимальная видовая насыщенность на сообщество наблюдается в типах леса ельник кисличный (43.0) и ельник сложный (42.7). На третьем месте (с заметным отрывом) – сложный сосняк (37.9). Все остальные: бореальные сосняки, ельник черничный и дубо-липняк – попадают в группу близких значений разнообразия в интервале 32.1–34.6. Можно отметить, что максимальная видовая насыщенность в кисличном и сложном ельниках положительно коррелирует с максимальным для данных типов числом видов во флористическом списке (даже с учетом бóльших размеров выборки в данных типах).
Результаты для относительно старовозрастных коренных лесов интересно сравнить с литературными данными по аналогичным синтаксонам, где описания не проходили столь строгий отбор на возраст и ненарушенность. Наиболее полно такие данные недавно представлены в работе Т.В. Черненьковой с коллегами (Черненькова и др., 2020). В этой работе нет данных по соснякам лишайниковым, брусничным, а в характеристиках синтаксонов представлены данные только по общей видовой насыщенности (без разбивки по ярусам) и не дается ошибка средней. Несколько отличаются также размер площадки (400 м2), подход к выделению и объем синтаксонов. Тем не менее, шесть из восьми типов леса в табл. 1 имеют аналоги в виде групп ассоциаций (Черненькова и др., 2020). Несмотря на различия в методах, результаты исследований принципиально сходны, что подтверждает их объективный характер. Так, совпадает расположение синтаксонов в порядке снижения общей видовой насыщенности. На первом месте – аналог кисличного ельника (ельник мелкотравно-широкотравный). Второе и третье место занимают аналоги ельника сложного и сложного сосняка (широкотравный ельник, елово-широколиственный широкотравный лес и широкотравный сосняк). На четвертом месте – аналог черничного ельника (ельник кустарничковый мелкотравно-зеленомошный).
По абсолютным значениям общей насыщенности сравнение результатов с данными Т.В. Черненьковой и коллег (2020) привело к несколько неожиданным результатам. Сами значения – в целом близки. Но в четырех из шести синтаксонах общая видовая насыщенность в коренных типах леса несколько выше, чем в “широкой” выборке у коллег. Так, в типе леса “ельник-кисличник” она составила 43.0 (табл. 1) против 39 в синтаксоне “ельник мелкотравно-широкотравный”; в ельнике сложном – 42.7 (табл. 1) против 36 в синтаксоне “широкотравный ельник”. С учетом большего возраста, меньшей нарушенности и незначительно меньшей площади описаний мы ожидали, что в нашей выборке коренных сообществ видовая насыщенность будет меньше, чем в “широкой” выборке у коллег. Можно предположить, что более низкая видовая насыщенность в “широкой выборке” (Черненькова и др., 2020) является следствием присутствия в описаниях большого числа лесных культур с обедненным видовым составом. Действительно, в описаниях синтаксонов (особенно ельников) авторы часто упоминают культуры. Интересно, что в коренном типе леса “дубняк липовый” и синтаксоне широкой выборки “дубовые с липой леса” числа общей насыщенности совпали полностью (32 вида на описание).
Индекс Шеннона
Интегральная оценка видового разнообразия в изученных типах леса проводилась путем вычисления информационного индекса Шеннона. Во всех восьми типах леса значения индекса располагаются в диапазоне от 2.26 до 2.82. Не очень большие различия частично связаны с полевой методикой, так как балльные шкалы покрытия нелинейно зависят от покрытия (фитомассы) и потому (по мнению Э. Мэгарран, 1992) “дают не слишком точные результаты” при расчете индексов. Вторая причина небольшого разброса значений – присутствие во всех типах леса видов–доминантов с высоким баллом обилия. Как показали В.К. Шитиков и Г.С. Розенберг (2005), при сильном доминировании кривая зависимости индекса Шеннона Н от числа видов S стремится к значению 2.0. Интересно, что разброс значений индекса мало зависит даже от группы изучаемых организмов: в монодоминантных еловых культурах Ирландии для видов птиц было получено значение индекса 2.056, а для естественных дубовых лесов – 2.404 (Batten, 1976; цит. по: Мэгарран, 1992).
Максимальное видовое разнообразие (равномерность распределения) по индексу Шеннона характерно для ельника сложного (2.82) и для ельника кисличного (2.80). Минимальное видовое разнообразие по Шеннону наблюдается в сосняке лишайниковом (2.26) и сосняке брусничном (2.42).
К сожалению, данные по расчетам индекса Шеннона для лесов Московского региона в литературе отсутствуют, а по другим регионам методически отличаются и фрагментарны, что затрудняет сравнение результатов. В одной из первых отечественных работ по расчету индекса (Дыренков и др., 1981) приводятся данные для трех типов коренных ельников южной тайги (Ленинградская обл.). Хотя сравнивать данные по всем ярусам (табл. 1) и по ярусу C не совсем корректно, значения индекса в аналогичных типах довольно близки. Так, в ельнике сложном (табл. 1) индекс Шеннона равняется 2.82; в аналогичном травяно-дубравном (Дыренков и др., 1981) – 2.91. В этой связи не очень понятно, как были получены значения индекса более 4.0 для ряда лесных сообществ в “Калужских засеках” (Ханина и др., 2016).
Кривые доминирования-разнообразия
Важную информацию о параметрах видового разнообразия в сообществах разных типов дает построение кривых значимости видов (Уиттекер, 1980). Их называют также кривыми рангового распределения обилий (Мэгарран, 1992) или кривыми доминирования–разнообразия (Шитиков, Розенберг, 2005). В отечественной литературе такие кривые нередко используют для анализа видового разнообразия групп животных (Мухаметова, 2010; Трошкова и др., 2015), однако для видов лесных растений нам удалось обнаружить лишь график для сосняков лесотундры (Черненькова, 2002).
На рис. 3 представлены кривые доминирования–разнообразия видов нижних ярусов на постоянных пробных площадях в четырех типах леса. Порядок видов вдоль оси X соответствует рангу вида в ряду от видов с наибольшей значимостью до видов с наименьшей значимостью (Уиттекер, 1980).
Рис. 3.
Кривые доминирования–разнообразия нижних ярусов на постоянных пробных площадях в четырех типах леса. По оси X – ранг вида, по оси Y – встречаемость вида на площадках 0.2 × 0.2 м, %.
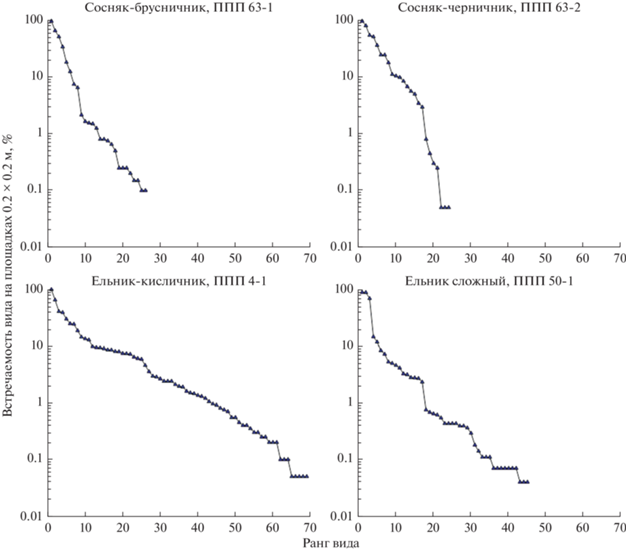
Характер кривых в двух бореальных сосняках и двух неморальных ельниках существенно отличается, хотя сравнение каждого из распределений с теоретическими моделями (геометрический, логнормальный ряд) не позволило статистически достоверно отнести наблюдаемые распределения к одной из этих моделей.
Для сосняка брусничного и сосняка черничного характерен резкий наклон кривой с большими различиями значимости между соседними по рангу видами. Данная форма графиков более соответствует геометрическому ряду и гипотезе перехвата ниш сильными доминантами. Как правило, такое распределение наблюдается в бедных видами местообитаниях с явными лимитирующими факторами или на ранних стадиях сукцессии (Уиттекер, 1980). Оба критерия справедливы в отношении сосняка брусничного (Маслов, Логофет, 2020). Видовая насыщенность и индекс Шеннона в сосняках бореального типа находятся в группе самых низких значений.
В ельнике сложном и особенно в ельнике кисличном наклон кривой существенно меньше (рис. 3). В последнем сообществе вообще отсутствуют большие различия значимости между соседними по рангу видами, а форма пологой кривой близка к S-образной, характерной для логнормального распределения. Это означает, что встречаемость видов определяется в данном сообществе большим числом независимых переменных, которые неодинаково влияют на разные виды (Уиттекер, 1980). Такая форма кривой наблюдается в зрелых, разнообразных природных сообществах (Мэгарран, 1992). Видовая насыщенность и индекс Шеннона в ельниках неморального типа находятся в группе самых высоких значений. Интересно, что первый по рангу вид, кислица (Oxalis acetosella L.), имеет в кисличном ельнике встречаемость около 100%, то есть формально является доминантом. Однако общая форма кривой доминирования–разнообразия свидетельствует о незначительных эдификаторных свойствах кислицы, что подтверждает работа Т.В. Черненьковой (1982).
ЗАКЛЮЧЕНИЕ
Массив геоботанических описаний в заповедных, наиболее сохранившихся коренных лесах Московского региона конца XX века представляет большую ценность в качестве “эталона” природных сообществ при сравнении с молодыми, вторичными и антропогенно нарушенными лесами. В настоящей работе представлены данные по оценке биоразнообразия на уровне видов растений. В дальнейшем предполагается провести анализ разнообразия на уровне синтаксонов, включая и заболоченные леса. С учетом достаточно точной географической привязки данные о встречаемости отдельных видов можно использовать для уточнения ареалов, как это было сделано на примере ветреницы дубравной (Anemone nemorosa L.) (Маслов, 2010).
Впервые для коренных типов леса Московского региона по единой методике рассчитаны показатели биоразнообразия: видовая насыщенность, индекс Шеннона, построены графики доминирования–разнообразия.
Наибольшая видовая насыщенность древесного яруса (A) наблюдается в сложных типах леса – сосняке (4.9) и ельнике (4.4). Видовая насыщенность травяно-кустарничкового яруса (C) достигает наибольших значений (более 29) в неморальных ельниках – кисличном и сложном. Для яруса D (мохово-лишайникового) наибольшая видовая насыщенность обнаружена в типе леса сосняк брусничный (7.6).
В целом для типа леса (все ярусы) наибольшая видовая насыщенность обнаружена в неморальных типах ельников – кисличном и сложном (около 43). В этих же типах леса показано максимальное видовое разнообразие по индексу Шеннона (около 2.8), а форма кривых доминирования–разнообразия видов нижних ярусов имеет минимальный наклон. Такая форма кривой характерна для зрелых природных сообществ.
Наименьшая видовая насыщенность в целом для типа леса обнаружена в бореальных типах сосняков и ельников. Наименьших значений здесь достигает и индекс Шеннона, а форма кривых доминирования–разнообразия видов нижних ярусов характеризуется резким наклоном, что свойственно для бедных сообществ с явными лимитирующими факторами и/или сильными доминантами.
* * *
Автор благодарен коллегам – участникам группы по изучению заповедных лесных участков – Ю.В. Петерсону, Л.И. Савельевой и Е.С. Комиссарову, а также Ю.Е. Алексееву и Е.А. Игнатовой – за помощь в определении трудных видов растений и S.M. Hennekens – за предоставленную возможность работы с лицензионной версией TurboVeg.
Список литературы
Алехин В.В. Растительность и геоботанические районы Московской и сопредельных областей. М.: МОИП, 1947. 71 с.
Дыренков С.А., Федорчук В.Н., Григорьева С.О. Видовое разнообразие растительных сообществ коренных таежных ельников // Экология. 1981. № 2. С. 26–33.
Ильинская С.А., Матвеева А.А., Речан С.П., Орлова М.А., Казанцева Т.Н. Типы леса // Леса Западного Подмосковья. М.: Наука, 1982. С. 20–150.
Коновалов Н.А. Типы леса подмосковных опытных лесничеств Ц.Л.О.С. М.; Л.: Сельхозгиз, 1929. 159 с.
Курнаев С.Ф. Основные типы леса средней части Русской равнины. М.: Наука, 1968. 355 с.
Курнаев С.Ф. Дробное лесорастительное районирование Нечерноземного центра. М.: Наука, 1982. 118 с.
Леса Восточного Подмосковья / Под ред. С.Ф. Курнаева. М.: Наука, 1979. 184 с.
Леса Западного Подмосковья / Под ред. Л.П. Рысина. М.: Наука, 1982. 236 с.
Леса Москвы: Опыт организации мониторинга / Под ред. Л.П. Рысина. М.: Грааль, 2001. 148 с.
Леса Северного Подмосковья / Под ред. Л.П. Рысина. М.: РАН, 1993. 316 с.
Леса Южного Подмосковья / Под ред. Л.П. Рысина. М.: Наука, 1985. 280 с.
Лесная энциклопедия. Т. 1. М.: Сов. энциклопедия, 1985. 563 с.
Маслов А.А. Количественный анализ горизонтальной структуры лесных сообществ. М.: Наука, 1990. 160 с.
Маслов А.А. Динамика фитоцено-экологических групп видов и типов леса в ходе природных сукцессий заповедных лесов центра Русской равнины // Бюл. МОИП. Отд. биол. 1998. Т. 103. № 2. С. 34–43.
Маслов А.А. Мониторинг биоразнообразия и процессов природной динамики в заповедных лесных участках: программа и итоги работ за 25 лет // Структура и функции лесов Европейской России. М.: Товарищество научных изданий КМК, 2009. С. 172–190.
Маслов А.А. Ветреница дубравная Anemone nemorosa L. в заповедных лесных участках Московской области // Восстановление и мониторинг природной флоры. М.: Товарищество научных изданий КМК, 2010. С. 85–91.
Маслов А.А. Разграничение видов березы по форме листьев: сравнение дискриминантных методов // Лесоведение. 2021. № 5. С. 523–530.
Маслов А.А., Логофет Д.О. Совместная динамика популяций черники и брусники в заповедном послепожарном сосняке-зеленомошнике. Модель с осредненными вероятностями перехода // Журнал общей биологии. 2020. Т. 81. № 4. С. 243–256.
Морозова О.В., Беляева Н.Г., Гнеденко А.Е., Суслова Е.Г., Черненькова Т.В. Синтаксономия и экология черноольшаников Московской области // Растительность России. 2021. № 42. С. 42–62.
Московская область: Атлас / Под ред. Э.Г. Галиуллиной, В.И. Юрченко. М.: ГУГК, 1979. 40 с.
Мухаметова О.Н. Сравнительная характеристика ихтиопланктона лагунных озер юго-восточной части Сахалина // Вопросы ихтиологии. 2010. Т. 50. № 5. С. 685–695.
Мэгарран Э. Экологическое разнообразие и его измерение. М.: Мир, 1992. 184 с.
Огуреева Г.Н., Микляева И.М., Суслова Е.Г., Швергунова Л.В. Растительность Московской области. Пояснительный текст к карте (м-б 1 : 200 000). М.: Экор, 1996. 45 с.
Оценка и сохранение биоразнообразия лесного покрова в заповедниках европейской России / Под ред. Л.Б. Заугольновой. М.: Научный мир, 2000. 196 с.
Рекомендации по выделению коренных и производных групп типов леса лесной зоны европейской части РСФСР / Побединский А.В., Лазарев Ю.А., Ханбеков Р.И., Орлов А.Я., Абатуров Ю.Д. М.: ВНИИЛМ, 1982. 40 с.
Рысин Л.П. Типы леса Восточного Подмосковья // Леса Восточного Подмосковья. М.: Наука, 1979. С. 39–125.
Рысин Л.П. Лесная типология в СССР. М.: Наука, 1982. 217 с.
Рысин Л.П. Конспект лесной флоры средней полосы Русской равнины (сосудистые растения). М.: Товарищество научных изданий КМК, 2009. 177 с.
Рысин Л.П. Леса Подмосковья. М.: Товарищество научных изданий КМК, 2012. 256 с.
Рысин Л.П., Савельева Л.И. Эталонные леса, их значение и критерии выбора // Ботанический журнал. 1980. Т. 65. № 1. С. 133–140.
Рысин Л.П., Савельева Л.И. Лесные заповедные участки. М.: Агропромиздат, 1985. 168 с.
Рысин Л.П., Савельева Л.И. Сосновые леса России. М.: Товарищество научных изданий КМК, 2008. 290 с.
Трошкова И.Ю., Трошков Н.Ю., Никитский Н.Б. Жужелицы (Coleoptera, Carabidae) лесов приокской террасы в Серпуховском районе Московской области // Бюллетень МОИП. Отд. биол. 2015. Т. 120. № 1. С. 26–34.
Уиттекер Р. Сообщества и экосистемы. М.: Прогресс, 1980. 327 с.
Федорчук В.Н., Егоров А.А., Гаубервиль К., Чернов И.М. Краткий определитель типов леса Ленинградской области. СПб: Арт Юнион, 2002. 36 с.
Ханина Л.Г., Смирнов В.Э., Бобровский М.В. Оценка видового богатства растительных сообществ заповедника “Калужские засеки” на основе индексов Шеннона и Симпсона и чисел Хилла // Лесоведение. 2016. № 1. С. 65–73.
Черненькова Т.В. Влияние эколого-фитоценотических факторов на кислицу обыкновенную (Oxalis acetosella L.) // Бюллютень МОИП. Отд. биол. 1982. Т. 87. № 5. С. 59–66.
Черненькова Т.В. Реакция лесной растительности на промышленное загрязнение. М.: Наука, 2002. 190 с.
Черненькова Т.В., Морозова О.В., Беляева Н.Г., Пузаченко М.Ю. Современная организация лесных сообществ с участием широколиственных пород в зоне широколиственно-хвойных лесов (на примере Московской области) // Растительность России. 2018. № 33. С. 107–130.
Черненькова Т.В., Суслова Е.Г., Морозова О.В., Беляева Н.Г., Котлов И.П. Биоразнообразие лесов Московского региона // Экосистемы: экология и динамика. 2020. Т. 4. № 3. С. 61–144.
Шитиков В.К., Розенберг Г.С. Оценка биоразнообразия: попытка формального обобщения // Количественные методы экологии и гидробиологии. Тольятти: СамНЦ РАН, 2005. С. 91–129.
Chytrý M., Otýpková Z. Plot sizes used for phytosociological sampling of European vegetation // Journal of Vegetation Science. 2003. V. 14. № 4. P. 563–570.
Chytrý M. et al. European Vegetation Archive (EVA): an integrated database of European vegetation plots // Applied vegetation science. 2016. V. 19. № 1. P. 173–180.
Guo Q. Ecosystem maturity and performance // Nature. 2005. V. 435. P. E6.
Hennekens S.M., Schaminee J.H.J. TURBOVEG, a comprehensive data base management system for vegetation data // Journal of Vegetation Science. 2001. V. 12. № 4. P. 589–591.
Hytteborn H., Maslov A.A., Nazimova D.I., Rysin L.P. Boreal forests of Eurasia // Coniferous forests. Amsterdam, etc.: Elsevier, 2005. P. 23–99. (Ecosystems of the World, V. 6).
Jongman R.H.G., Ter Braak C.J.F., van Tongeren O.F.R. Data analysis in community and landscape ecology. Cambridge: Cambridge University Press, 1995. 299 p.
McCune B., Mefford M.J. PC-ORD. Multivariate analysis of ecological data. Version 7. Gleneden Beach, Oregon: MjM Software Design, 2016. 34 p.
Rodwell J.S. (ed.). British plant communities. V. 1. Woodlands and scrub. Cambridge: Cambridge Univ. Press, 1991. 395 p.
Rodwell J.S., Mucina L., Pignatti S., Schaminée J.H.J., Chytr-ý M. European vegetation survey: The context of the case studies // Folia Geobot. et Phytotaxon. 1997. V. 32. № 2. P. 113–115.
Schaminée J., Hennekens S., Chytry M., Rodwell J. Vegetation-plot data and databases in Europe: An overview // Preslia. 2009. V. 81. № 3. P. 173–185.
Дополнительные материалы отсутствуют.