

Лесоведение, 2022, № 6, стр. 674-686
Флористическое разнообразие ксеро-мезофитных широколиственных лесов Юго-Запада России
Ю. А. Семенищенков *
Брянский государственный университет имени академика И.Г. Петровского
241036 Брянск, ул. Бежицкая, д. 14, Россия
* E-mail: yuricek@yandex.ru
Поступила в редакцию 22.03.2022
После доработки 08.04.2022
Принята к публикации 07.06.2022
- EDN: SFSMAR
- DOI: 10.31857/S0024114822060092
Аннотация
Ксеро-мезофитные широколиственные леса – важная составляющая общего разнообразия лесной растительности Юго-Запада России. Эти сообщества относятся к двум эколого-географическим группам: дубовым разнотравным флористически богатым лесам Сарматского региона и дубовым разнотравным остепненным лесам Понтического региона. Цель настоящей статьи – дать анализ флоры ксеро-мезофитных лесов для обоснования их природоохранной значимости в регионе. В ценофлоре ксеро-мезофитных широколиственных лесов отмечены 508 видов сосудистых растений и 30 видов мохообразных. Сообщества характеризуются наибольшей флористической насыщенностью среди всех групп широколиственных лесов в изучаемом регионе, что было установлено на основе сравнения 1296 геоботанических описаний широколиственных лесов Юго-Запада России. Проявление высокого флористического разнообразия ксеро-мезофитных сообществ – значительная ботанико-географическая и экологическая гетерогенность ценофлоры, что демонстрируют спектры геоэлементов/полизональных групп и экобиоморф. В соответствии с результатами DCA-ординации, наибольшие значения флористической насыщенности и выровненности обилия видов наблюдаются у субконтинентальных сообществ ксеро-мезофитных лесов в северной части их ареала в местообитаниях с наиболее бедными минеральным азотом умеренно увлажненными почвами. В ценофлоре ксеро-мезофитных широколиственных лесов Юго-Запада России выявлено 79 видов сосудистых растений, занесенных в Красные книги России (2008) или субъектов РФ на Юго-Западе страны. Среди охраняемых наиболее многочисленны виды трех групп: тепло- и светолюбивые южные по происхождению субпонтические (лесостепные), мезофильные неморальные виды широколиственных лесов и ксерофильные понтические (степные) виды. Можно считать сообщества изучаемого типа своеобразными проводниками лесостепных и степных видов к северу, в зону широколиственных лесов; к юго-востоку, напротив, такие сообщества становятся рефугиумами некоторых северных, в том числе бореальных и суббореальных видов.
Флористическое разнообразие лесных сообществ – важнейший показатель их экологического состояния, индикатор антропогенных нарушений и способности растительного покрова к восстановлению в условиях глобального преобразования экосистем человеком. Анализу зависимости основных показателей разнообразия флоры (состава, структуры, динамики) посвящены многочисленные исследования фитоценологов и экологов (Margalef, 1958; Whittaker, 1972; Magurran, 1988; Василевич, 1992; Юрцев, 1992; Zobel, 1992; Palmer, 1994; Hooper, Vitousek, 1997; Zobel et al., 1998; Cardinale et al., 2000; Duffy, 2002; Лебедева и др., 2004; Hooper et al., 2005; Морозова, 2008; Grime, 2009; Миркин, Наумова, 2012; Онипченко, 2013; и др.).
Особую эколого-географическую группу лесной растительности Юго-Запада России составляют так называемые ксеро-мезофитные леса, неоднократно привлекавшие внимание исследователей в этом регионе (Булохов, 1991; Морозова, 1999; Булохов, Соломещ, 2003; Булохов, Харин, 2008; Семенищенков, 2009, 2012, 2016; Зеленая книга …, 2012; Булохов, Семенищенков, 2013; Семенищенков, Полуянов, 2014; и др.). В соответствии с лесной типологией, эти леса можно назвать дубовыми (и производными) или дубовыми с участием сосны разнотравными; их местообитания соответствуют типам лесорастительных условий Д2 и С2 (они иногда формируются в условиях суборей с участием сосны). С позиций флористической классификации растительность ксеромезофитных широколиственных лесов на Юго-Западе России представлена сообществами двух союзов. Союз Betonico officinalis–Quercion roboris Goncharenko et Semenishchenkov in Goncharenko et al. 2020 объединяет ксеро-мезофитные флористически богатые дубовые леса Сарматского региона. Другой союз – Scutellario altissimae–Quercion roboris Goncharenko in Goncharenko et al. 2020 – представляет ксеро-мезофитные остепненные разнотравные дубовые леса Понтического региона (Goncharenko et al., 2020). Эти единицы входят в состав порядка Quercetalia pubescenti-petraeae Klika 1933 и класса Quercetea pubescentis Doing-Kraft ex Scamoni et Passarge 1959, объединяющие в Восточной Европе дубовые леса теплых и умеренно прохладных регионов неморальной зоны. В пределах всего своего ареала сообщества этих союзов соседствуют с широколиственными лесами других эколого-географических групп. Мезофитные леса на северо-западе региона относятся к союзу хвойно-широколиственных и широколиственно-хвойных лесов Русской равнины и Прибалтики Querco roboris–Tilion cordatae Solomeshch et Laiviņš ex Bulokhov et Solomeshch in Bulokhov et Semenishchenkov 2015; в юго-восточной части, за пределами ареала ели европейской (Picea abies L.) на плакорах, – к союзу среднерусско-приволжских широколиственных лесов Aceri campestris–Quercion roboris Bulokhov et Solomeshch in Bulokhov et Semenishchenkov 2015 (класс Carpino–Fagetea sylvaticae Jakucs ex Passarge 1968). Ацидофитные дубовые и сосново-дубовые леса на бедных почвах в центральной части региона относятся к союзу Vaccinio myrtilli–Quercion roboris Bulokhov et Solomeshch 2003 класса Quercetea robori-petraeae Br.-Bl. et Tx. ex Oberd. 1957. Пойменные преимущественно дубовые леса объединяет союз Fraxino–Quercion roboris Passarge 1968 в составе класса древесной пойменной растительности Alno glutinosae–Populetea albae P. Fukarek et Fabijanić 1968.
Высокое флористическое разнообразие ксеромезофитных лесов региона ранее отмечалась в литературе (Булохов, Семенищенков, 2013). Они обнаруживают значительное флористическое сходство с аналогичными сообществами из Центральной и Южной Европы, где подробно описаны в литературе (Семенищенков, 2016; Goncharenko et al., 2020). Однако распространение этих лесов за северо-восточными пределами ареалов некоторых важнейших древесных эдификаторов (в первую очередь граба обыкновенного (Carpinus betulus L.), бука лесного (Fagus sylvaticus L.), дуба скального (Quercus petraea (Matt.) Liebl.), дуба пушистого (Q. pubescens Willd.)) и утрата на градиенте Запад–Восток многочисленных западных и южных представителей флоры определяют ботанико-географическое своеобразие их сообществ в Восточной Европе (Булохов, Семенищенков, 2013; Семенищенков, 2016; Goncharenko et al., 2020). Уникальность данных лесов на Юго-Западе России связана с тем, что в их состав входят многочисленные различные по географическому происхождению виды, охраняемые в России или в ее субъектах. Однако до сих пор не был проведен специальный анализ флоры этих сообществ в регионе для обоснования их природоохранной значимости, что и стало целью настоящей работы.
Природные условия района исследования. Район исследования охватывает Юго-Запад России, лежащий практически полностью в пределах Среднерусской возвышенности Русской равнины. Ксеро-мезофитные леса встречаются здесь на эрозионных лессовых плато, в долинах наиболее крупных рек (Десна, Ипуть, Ока), в ландшафтах ополий, сформировавшихся по возвышенным речным правобережьям, на древних речных террасах.
Данная территория располагается в области умеренного климата с четко выраженными холодными и теплыми периодами года. В соответствии с классификацией Кеппена–Гейгера (Kottek et al., 2006), регион характеризуется Dfb-типом климата (континентальный влажный с теплым летом). Среднегодовое количество осадков – от 450 мм – на северо-западе до 620 мм – на юго-востоке. Среднегодовая температура воздуха изменяется на данном географическом градиенте от 4.7 до 6.9°С.
В северо-западной части региона преобладают серые лесные суглинистые почвы, сформировавшиеся под широколиственными лесами. Серые и темно-серые почвы приурочены преимущественно к водораздельным пространствам и пологим склонам, в то время как светло-серые занимают более крутые склоны (Природа …, 2012). На склоновых местностях представлен комплекс в разной степени смытых балочных почв. Для юго-востока региона характерно сочетание в разной степени оподзоленных и выщелоченных, типичных, среднегумусных и тучных мощных черноземов с серыми лесными почвами (Атлас …, 2000). Леса изучаемого типа занимают и специфические местообитания – склоны речных долин, балок и прибалочные плато с близким залеганием и выходом на поверхность карбонатных пород (мела, мергеля, известняка).
Данная территория представляет собой староосвоенный регион со значительно трансформированной природой. Лесистость территории – от 45% – на северо-западе до 4–5% – на юго-востоке. В связи с интенсивным уничтожением человеком широколиственные леса сохранились мелкими фрагментами на фоне сельскохозяйственных земель. Лесные сообщества охраняются в многочисленных ООПТ разного уровня, в том числе заповедниках “Белогорье”, “Брянский лес”, “Воронежский”, “Галичья Гора”, “Куликово поле”, “Центрально-Черноземный им. проф. В.В. Алехина”.
По ботанико-географическому районированию северо-восточная часть района исследования лежит в пределах Среднерусской подпровинции Восточноевропейской широколиственнолесной провинции Европейской широколиственнолесной области (зона широколиственных лесов); юго-восточная – Среднерусской (Верхнедонской) подпровинции Восточноевропейской лесостепной провинции Евразиатской степной области (лесостепная зона). Широколиственные леса являются зональным типом растительности на всех перечисленных территориях (Исаченко, Лавренко, 1980).
ОБЪЕКТЫ И МЕТОДИКА
Анализ флористического разнообразия ксеромезофитных широколиственных лесов Юго-Запада России проведен на основе базы данных, включающей 234 геоботанических описания из Белгородской, Брянской, Воронежской, Калужской, Курской, Липецкой, Орловской, Тульской областей России. В нее вошли 170 ранее опубликованных (Морозова, 1999; Булохов, Соломещ, 2003; Семенищенков, 2009; Семенищенков, Телеганова, 2013; Семенищенков, Полуянов, 2014; Семенищенков, 2016; Семенищенков, Шапурко, 2018) и неопубликованные авторские описания (64).
Оценка экологических режимов местообитаний сообществ проведена с использованием шкал Х. Элленберга (Ellenberg et al., 1992) на основе невзвешенных средних значений в программе JUICE (Tichý, 2002). DCA-ординация реализована средствами пакета R (https://www.r-project. org), интегрированного с программой JUICE. Корреляции осей с экологическими факторами определена с помощью коэффициента корреляции Кендалла в программе PC-ORD 5.0 (McCune, Mefford, 2006).
Для редких видов сосудистых растений верность выборке геоботанических описаний ксеромезофитных лесов установлена с использованием статистического φ-коэффициента верности (C-hytrý et al., 2002); к верным отнесены виды со значениями коэффициента более 20 (p < 0.05).
В качестве показателей флористического разнообразия определены флористическая насыщенность (видовое богатство на площадке в 400 м2), значения индекса Шеннона-Уивера (Margalef, 1958) на основе данных процентного обилия/покрытия видов в сообществах и относительное альфа-разнообразие, отражающее гетерогенность ценофлоры (Миркин, Наумова, 2012). Сравнительный анализ данных показателей проведен с использованием базы данных по лесной растительности, включающей 1296 авторских и ранее опубликованных разными авторами геоботанических описаний (Семенищенков, 2016; с добавлением). Типы геоэлементов сосудистых растений даны по H. Walter (1977), полизональных групп и экобиоморф – по А.Д. Булохову (2004). Статистическая обработка результатов выполнена в среде MS Excel.
Названия сосудистых растений приведены в соответствии с базой The Euro+Med PlantBase (http://www.emplantbase.org/).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Общая характеристика флористического разнообразия и его экологические тренды. В ценофлоре ксеро-мезофитных широколиственных лесов на Юго-Западе России отмечено 508 видов сосудистых растений и 30 видов мохообразных. Эти сообщества характеризуются наибольшей флористической насыщенностью среди всех групп широколиственных лесов в изучаемом регионе (табл. 1).
Таблица 1.
Показатели флористического разнообразия основных эколого-географических групп широколиственных лесов Юго-Западной России
| Союзы | Количество описаний | Количество видов в ценофлоре | Флористическая насыщенность | Индекс Шеннона-Уивера | Относительное альфа-разнообразие | ||
|---|---|---|---|---|---|---|---|
| мини-мальная | макси-мальная | средняя | |||||
| Ксеромезофитные леса | |||||||
| Ксеро-мезофитные дубовые разнотравные леса Сарматского региона | 210 | 500 | 18 | 96 | 44.2 ± 0.98 | 2.77 ± 0.03 | 1.76 |
| Ксеро-мезофитные остепненные дубовые разнотравные леса Понтического региона | 24 | 160 | 12 | 63 | 35.3 ± 1.94 | 2.13 ± 0.07 | 1.44 |
| Мезофитные леса | |||||||
| Среднерусско-приволжские широколиственные леса неморального состава без участия Picea abies | 500 | 280 | 7 | 38 | 22.4 ± 0.24 | 2.04 ± 0.01 | 1.38 |
| Елово-широколиственные леса неморального состава Русской равнины и Прибалтики | 322 | 244 | 18 | 48 | 28.1 ± 0.48 | 2.20 ± 0.09 | 1.06 |
| Гигро-мезофитные леса | |||||||
| Пойменные разнотравные дубовые леса Русской равнины | 143 | 322 | 13 | 68 | 28.7 ± 0.90 | 2.18 ± 0.04 | 1.91 |
| Ацидофитные леса | |||||||
| Дубовые и сосново-дубовые разнотравные леса на бедных почвах Юго-Запада России | 97 | 201 | 24 | 52 | 33.02 ± 0.74 | 2.24 ± 0.06 | 0.85 |
В пределах ксеро-мезофитных лесов отмечены сообщества с максимальными значениями видового богатства (96 видов – леса Сарматского региона). Данный показатель высок, хотя заметно ниже для группы сообществ Понтического региона (63). Эти факты подтверждают высокое флористическое разнообразие сообществ данной эколого-географической группы. Высокое видовое богатство характерно для светлых разреженных сообществ обычно с редким или угнетенным подлеском, что ведет к “олуговению” с насыщением свето- и теплолюбивыми опушечными и луговыми видами. Способствует этому и характерная “открытая” структура фитоценозов, сформированных преимущественно так называемой “поздней” формой дуба с высокими очищенными от ветвей стволами.
С возрастанием флористической насыщенности закономерно на статистически значимом уровне возрастает выровненность обилия видов (рис. 1). Это объясняется уменьшением их вклада в общее обилие при увеличении количества видов. В многовидовых сообществах возрастает количество малообильных видов, нередко представленных единичными экземплярами.
Рис. 1.
Зависимость выровненности обилия видов (индекс Шеннона-Уивера) от флористической насыщенности в сообществах ксеро-мезофитных широколиственных лесов Юго-Запада России.
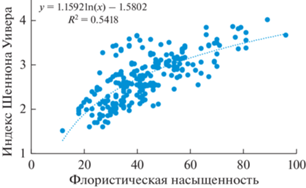
Следует отметить, что высокого значения видового богатства достигают и сообщества пойменных дубрав (68 видов), что объясняется некоторыми особенностями их состава, и в том числе экотонным эффектом с повышением биоразнообразия у границ контрастных сообществ поймы. В сухих экотопах, например, на песчаных гривах в речных поймах, дубравы иногда содержат многие характерные для ксеро-мезофитных лесов травянистые виды (Семенищенков, 2020).
Информативно характеризуют ценофлоры лесов сравниваемых типов значения средней флористической насыщенности, которые, несмотря на наличие отдельных сообществ с низким флористическим разнообразием, достигают максимальных значений для группы ксеро-мезофитных лесов. Сообщества с низким видовым богатством образуются в нескольких случаях. Это леса на крутых склонах балок или речных долин в условиях интенсивной почвенной эрозии, постпожарные и нарушенные выпасом или прогоном скота сообщества. Перечисленные факторы нередко ведут к значительному снижению видового богатства, хотя многие характерные виды лесных ассоциаций и союзов сохраняются в ценофлоре. В целом же ксеро-мезофитные леса характеризуются более высокой флористической насыщенностью в сравнении со средним значением (35 видов на 400 м2), установленным ранее для широколиственных лесов разного состава на примере днепровского бассейна (Юго-Запад России) (Семенищенков, 2016). Необходимо обратить внимание на то, что мохообразные (в описаниях отмечены только эпигейные виды) имеют низкое разнообразие (не более 3 видов в сообществе); моховой ярус в фитоценозах отсутствует.
Одно из проявлений высокого флористического разнообразия ксеро-мезофитных лесов – значительная ботанико-географическая и экологическая гетерогенность ценофлоры сосудистых растений, что наглядно демонстрируют спектры геоэлементов/полизональных групп и экобиоморф соответственно.
В ботанико-географическом спектре (рис. 2) выявлены 7 типов зональных геоэлементов и 6 полизональных групп. Характерно преобладание неморальных видов мезофитных широколиственных лесов со значительным участием свето- и теплолюбивых субпонтических лесостепных, южносибирских лесо-опушечных и даже понтических степных видов. Участие ряда суббореальных и бореальных представителей флоры характерно преимущественно для северной части ареала лесов данной группы. Среди полизональных видов наиболее представительны виды с широкими ареалами, распространенные от средиземноморья до зон тундры (Pzb) или тайги (Pzt). Это многочисленные растения, участвующие в составе растительности азонально-зональных типов на широком ботанико-географическом градиенте, например, дудник лесной (Angelica sylvestris L.), колокольчик рапунцелевидный (Campanula rapunculoides L.), осока сближенная (Carex contigua Huds.), ежа сборная (Dactylis glomerata L.), подмаренник бореальный (Galium boreale L.), борщевик сибирский (Heracleum sibiricum L.), бородавник обыкновенный (Lapsana communis L.), вербейник обыкновенный (Lysimachia vulgaris L.), лапчатка прямостоячая (Potentilla erecta (L.) Raeusch.), крапива двудомная (Urtica dioica L.), горошек заборный (Vicia sepium L.), короставник полевой (Knautia arvensis (L.) J.M. Coult.) и др. Большинство из них проявляют свойства гелиофитов, широко встречаются на лугах и опушках широколиственных лесов в достаточно большом диапазоне эдафических условий. Интересным является распространение в ксеро-мезофитных сообществах некоторых центральноевропейских видов у восточной границы ареала. Например, неморальные виды: крестообразник голый (Cruciata glabra (L.) Ehrend.), наперстянка крупноцветковая (Digitalis grandiflora Mill.), зверобой горный (Hypericum montanum L.) – в России известны только из юго-западных регионов, где встречаются в основном в ксеро-мезофитных лесах.
Рис. 2.
Спектр геоэлементов и полизональных групп ксеро-мезофитных широколиственных лесов Юго-Запада России. Геоэлементы (по: Walter, 1977): B – бореальный, Bs – суббореальный, N – неморальный, Sp – cубпонтический, Po – понтический, Js – южносибирский, P – плюрирегиональный; полизональные группы (по: Булохов, 2004): Pca – субсредиземноморско-арктическая, Pcb – субсредиземноморско-бореальная, Pct – субсредиземноморско-умеренная, Pza – средиземноморско-арктическая, Pzb – средиземноморско-бореальная, Pzt – средиземноморско-умеренная.
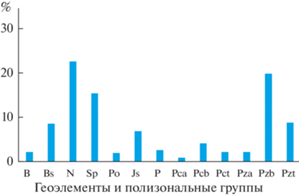
В целом описанное выше ботанико-географическое разнообразие ценофлоры, вероятно, связано со значительным распространением данного более юго-западного по происхождению типа сообществ к северу на Русской равнине, где они создают благоприятное фитоценотическое окружение для многочисленных видов разной зональной приуроченности.
Спектр экобиоморф также отличается значительным разнообразием: в нем отмечены 11 типов (рис. 3). Он характеризует данные леса как ксеромезофитные, так как в ценофлоре явно преобладают виды мезоморфной экобиоморфы (среди них большинство неморальных) с наиболее заметным участием ксероморфных и “переходных” к ним ксеро-мезо- и мезо-ксероморфных растений.
Рис. 3.
Спектр экобиоморф ксеро-мезофитных широколиственных лесов Юго-Запада России.
Экобиоморфы (по: Булохов, 2004): he – геломорфные, he/m – гело-мезоморфные, hg – гигроморфные, hg/m – гигро-мезоморфные, k – ксероморфные, k/he – ксеро-геломорфные, k/m – ксеро-мезоморфные, m – мезоморфные, m/he – мезо-геломорфные, m/hg – мезо-гигроморфные, m/k – мезо-ксероморфные.
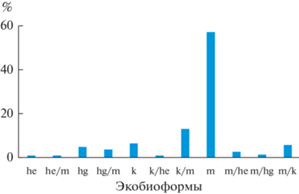
В целом на значительную гетерогенность ценофлоры ксеро-мезофитных лесов указывают высокие значения показателя относительного альфа-разнообразия (табл. 1). Фактически это связано с большим варьированием флористического состава сообществ при сохранении комплекса характерных видов ассоциаций и союзов; данное обстоятельство, следуя Б.М. Миркину (Миркин, Наумова, 2012), существенно затрудняет классификацию растительности.
Какие же факторы в наибольшей степени определяют флористическое разнообразие ксеромезофитных лесов? DCA-ординация (рис. 4, табл. 2 и 3) продемонстрировала роль ведущих факторов среды в дифференциации геоботанических описаний, которые характеризуются, в свою очередь, индивидуальными показателями флористического разнообразия: флористической насыщенностью и выровненностью обилия видов, оцененной с помощью индекса Шеннона-Уивера. Наибольший вклад в эколого-флористическую дифференциацию сообществ вносят факторы влажности, кислотности почвы, а также температуры, градиенты которых в наибольшей степени коррелируют с осью DCA1 ординации с наибольшей нагрузкой. С ней на статистически значимом уровне коррелирует и значение выровненности обилия видов. Однако флористическая насыщенность обнаруживает корреляцию в большей степени с осью DCA2. Она отчасти соответствует градиенту почвенного богатства минеральным азотом и континентальности. Таким образом, наибольшие значения флористической насыщенности и выровненности наблюдаются у субконтинентальных сообществ ксеро-мезофитных лесов в северной части их ареала в местообитаниях с небогатыми минеральным азотом, умеренно увлажненными почвами. На изучаемой территории такие местообитания приурочены преимущественно к древним речным террасам; именно здесь описаны сообщества с максимальным видовым богатством.
Рис. 4.
Диаграмма DCA-ординации (оси 1 и 2) ксеро-мезофитных широколиственных лесов Юго-Запада России. Эколого-географические группы: 1 – дубовые разнотравные леса Сарматского региона, 2 – остепненные дубовые разнотравные леса Понтического региона.
Векторы экологических факторов: CONT – континентальность, LIGHT – освещенность, MOIST – влажность почвы, NUTR – богатство почвы минеральным азотом, REACT – реакция почвы, TEMP – температурное число (определены по шкалам H. Ellenberg et al., 1992). N – флористическая насыщенность, H – индекс Шеннона-Уивера.
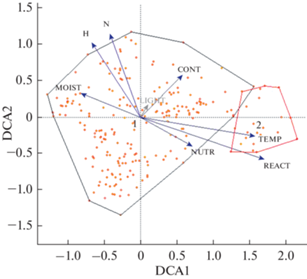
Таблица 2.
Численные параметры осей ординации
| Оси ординации | DCA1 | DCA2 |
|---|---|---|
| Нагрузка на ось | 0.365 | 0.204 |
| Длина оси | 3.427 | 2.792 |
Таблица 3.
Корреляция осей DCA-ординации со значениями экологических факторов и параметрами флористического разнообразия синтаксонов
| Оси ординации | DCA1 | DCA2 |
|---|---|---|
| Освещенность | 0.001 | 0.000 |
| Температура | 0.320 | 0.000 |
| Континентальность | 0.051 | 0.382 |
| Влажность почвы | 0.156 | 0.001 |
| Реакция почвы | 0.453 | 0.004 |
| Богатство почвы минеральным азотом | 0.074 | 0.221 |
| Видовое богатство | –0.069 | 0.304 |
| Индекс Шеннона-Уивера | –0.220 | 0.173 |
Следует отметить, что фактически неучтенными при ординации остаются некоторые факторы, которые, на наш взгляд, могут оказывать влияние на флористическое разнообразие: склоновые процессы в долинных и балочных местообитаниях и антропогенные нарушения. В полной мере статистически учесть эти факторы не представляется возможным. Однако, как и другие типы широколиственных сообществ, ксеро-мезофитные леса являются чувствительными к антропогенным нарушениям. Среди важнейших воздействий человека на них следует отметить палы травы и низовые пожары, приводящие к упрощению структуры и снижению видового разнообразия травяного яруса и подлеска, повреждению и выпадению из травостоя лесных травянистых видов, распространению устойчивых к пирогенному воздействию длиннокорневищных многолетников (например, вейника наземного (Calamagrostis epigeios (L.) Roth)). Изреживание или угнетение древостоя и подлеска приводит к осветлению, сопровождающемуся “олуговением” ценофлоры; ранее это явление было отмечено для городских нарушенных лесов (Булохов, Харин, 2008) и разреженных пойменных дубрав Нечерноземья (Семенищенков, 2020). Следует отметить влияние выпаса и прогона скота, в основном в краевых частях лесных массивов, ведущих к уплотнению почвы, внедрению в сообщества синантропных или даже чужеземных видов (например, райграса высокого (Arrhenatherum elatius (L.) J. Presl & C. Presl), мелколепестника однолетнего (Erigeron annuus (L.) Desf.), золотарника канадского (Solidago canadensis L.), иногда борщевика Сосновского (Heracleum sosnowskyi Manden.)). Благодаря открытости для инвазий и ксеро-мезофитности местообитаний сообщества могут стать местами распространения перечисленных чужеземных видов.
Раритетные компоненты флоры ксеро-мезофитных лесов и вопросы ее охраны. В ценофлоре ксеро-мезофитных широколиственных лесов Юго-Запада России выявлено 79 видов сосудистых растений, занесенных в Красные книги России (2008) или ее субъектов на Юго-Западе страны (табл. 4).
Таблица 4.
Охраняемые в субъектах РФ виды сосудистых растений и их константность в анализируемой выборке описаний ксеро-мезофитных широколиственных лесов Юго-Запада России
| Субъекты РФ (области) | Белгородская | Брянская | Воронежская | Калужская | Курская | Липецкая | Орловская | Тульская | Констант-ность в выборке описаний, % |
|---|---|---|---|---|---|---|---|---|---|
| Борец шерстистоустый (Aconitum lasiostomum) | . | + | + | . | + | + | . | . | 3 |
| Воронец колосистый (Actaea spicata L.) | . | . | + | . | . | . | . | . | 9 |
| Бубенчик лилиелистный (Adenophora liliifoliaL.) | + | + | + | + | + | + | + | + | 3 |
| Адонис весенний (Adonis vernalis L.) | + | + | + | . | + | + | + | + | 1 |
| Ветреница лесная (Anemone sylvestris L.) | + | + | . | + | + | + | . | . | 3 |
| Венечник ветвистый (Anthericum ramosum L.) | . | . | . | + | . | . | . | + | 20 |
| Подмаренник трехтычинковый (Asperula tinctoria L. [= Galium triandrum Hyl.]) | . | . | . | + | . | . | . | . | 7 |
| Барбарис обыкновенный (Berberis vulgaris L.) | . | + | . | . | . | . | . | . | 1 |
| Колокольчик жестковолосистый (Campanula cervicaria L.) | . | . | + | . | . | . | . | . | 1 |
| Колокольчик широколистный (C. latifolia L.) | + | . | . | . | + | + | . | . | 2 |
| Осока Микели (Carex michelii Host ) | . | . | . | . | . | . | + | + | 5 |
| Бодяк венгерский (Cirsium pannonicum L.) | . | + | . | . | . | . | . | + | 1 |
| Ломонос цельнолистный (Clematis integrifolia L.) | + | . | . | . | + | + | + | . | 2 |
| Ломонос прямой (C. recta L.) | . | + | . | + | . | . | . | + | 9 |
| Боярышник украинский (Crataegus ucrainica Pojark.) | . | + | . | . | . | . | . | . | 1 |
| Скерда сибирская (Crepis sibirica L.) | . | . | . | + | . | . | . | . | 3 |
| Венерин башмачок настоящий(Cypripedium calceolus L.) | + | + | + | + | + | + | + | + | 1 |
| Пузырник ломкий (Cystopteris fragilis L.) | . | + | . | . | . | . | . | . | 1 |
| Живокость клиновидная (Delphinium cuneatum Steven ex DC.) | . | . | + | . | . | + | + | + | 1 |
| Гвоздика Фишера (Dianthus fischeri Spreng.) | . | + | . | . | . | . | + | . | 1 |
| Гвоздика пышная (D. superbus L.) | + | + | + | + | + | + | + | + | 1 |
| Наперстянка крупноцветковая (Digitalis grandiflora Mill.) | . | + | . | . | + | . | + | . | 35 |
| Змееголовник Рюйша (Dracocephalum ruyschiana L.) | + | + | + | + | + | . | + | + | 1 |
| Дремлик чемерицевидный (Epipactis helleborine (L.) Crantz) | + | . | + | . | + | + | . | . | 18 |
| Бересклет европейский (Euonymus europaea L.) | . | . | . | + | . | . | . | . | 11 |
| Молочай полумохнатый (Euphorbia semivillosa Prokh.) | . | . | . | + | . | . | . | . | 4 |
| Дрок германский (Genista germanica L.) | . | + | . | . | . | . | + | . | 1 |
| Горечавка крестовидная (Gentiana cruciata L.) | + | + | + | + | + | . | . | . | 1 |
| Шпажник черепитчатый (Gladiolus imbricatus L.) /шпажник тонкий (Gladiolus tenuis M. Bieb.) | + | + | + | + | + | + | + | + | 1 |
| Зверобой волосистый (Hypericum hirsutum L.) | . | + | . | + | . | . | . | . | 6 |
| Зверобой горный (H. montanum L.) | . | + | . | . | . | . | . | . | 1 |
| Ирис безлистный (Iris aphylla L.) | + | + | + | + | + | + | + | + | 6 |
| Тонконог крупноцветковый (Koeleria macrantha (Ledeb.) Schult. [= Koeleria cristata (L.) Pers.]) | . | . | . | + | . | . | . | . | 2 |
| Гладыш широколистный (Laserpitium latifolium L.) | + | . | . | + | + | + | . | + | 37 |
| Гладыш прусский (L. prutenicum L.) | . | . | . | . | . | . | . | . | 7 |
| Чина молочно-белая (Lathyrus lacteus) | . | . | . | . | . | . | + | + | 1 |
| Чина гороховидная (L. pisiformis L.) | . | + | . | + | . | . | . | . | 14 |
| Лилия царские кудри (Lilium martagon L.) | + | + | + | + | + | + | + | + | 19 |
| Тайник яйцевидный (Listera ovata L.) | + | + | + | . | + | + | + | . | 1 |
| Воробейник лекарственный (Lithospermum officinale L.) | . | . | . | + | . | . | . | . | 2 |
| Дрёма двудомная (Silene dioica L.) | . | + | . | . | . | . | + | . | 1 |
| Перловник трансильванский (Melica transsilvanica Schur ) | . | . | . | . | . | . | . | + | 1 |
| Гнездовка настоящая (Neottia nidus-avis (L.) Rich.) | + | . | + | . | + | + | . | . | 5 |
| Вороний глаз четырехлистный (Paris quadrifolia L.) | . | . | + | . | . | . | . | . | 12 |
| Оленник Ривиниуса (Peucedanum cervaria (L.) Lapeyr. [= Cervaria rivinii Gaertn.]) | . | . | . | . | + | + | . | . | 6 |
| Любка двулистная (Platanthera bifolia L.) | + | . | + | . | + | + | . | . | 7 |
| Любка зеленоцветковая (Platanthera chlorantha (Custer) Rchb.) | + | + | + | + | + | + | + | . | 3 |
| Лапчатка белая (Potentilla alba L.) | + | . | . | + | . | + | . | . | 47 |
| Лапчатка семилисточковая (P. heptaphylla L.) | . | . | . | + | . | . | . | . | 1 |
| Черноголовка крупноцветковая (Prunella grandiflora L.) | . | + | . | . | . | . | + | . | 1 |
| Вишня кустарниковая (Prunus fruticosa Pall.) | . | + | . | . | . | . | . | . | 7 |
| Слива колючая (P. spinosa L.) | . | + | . | + | . | . | . | . | 15 |
| Поповник щитковый (Tanacetum corymbosum (L.) Sch. Bip. [= Pyrethrum corymbosum (L.) Scop.]) | . | . | . | + | . | . | . | . | 35 |
| Жостер слабительный (Rhamnus cathartica L.) | . | . | . | + | . | . | . | . | 19 |
| Подлесник европейский (Sanicula europaea L.) | . | + | . | . | . | . | + | + | 1 |
| Пролеска сибирская (Scilla siberica Haw.) | . | . | . | . | + | . | + | + | 1 |
| Козелец пурпурный (Scorzonera purpurea L.) | + | + | . | + | . | . | . | . | 1 |
| Шлемник высочайший (Scutellaria altissima L.) | . | . | . | . | . | + | . | + | 3 |
| Шлемник копьелистный (S. hastifolia L.) | . | . | . | . | . | . | + | + | 1 |
| Кадения сомнительная (Selinum dubium (Schkuhr) Leute [= Kadenia dubia (Schkuhr) Lavrova & V.N. Tikhom.]) | . | . | . | + | . | . | . | . | 1 |
| Серпуха венценосная (Serratula coronata L.) | . | . | . | + | . | . | . | . | 1 |
| Серпуха красильная (S. tinctoria L.) | . | . | . | + | . | . | . | . | 44 |
| Жабрица однолетняя (Seseli annuum L.) | . | . | . | + | . | . | . | . | 1 |
| Гулявник прямой (Sisymbrium strictissimum L.) | . | . | . | . | . | . | + | . | 1 |
| Спирея городчатая (Spiraea crenata L.) | . | . | . | . | + | . | + | + | 1 |
| Чистец прямой (Stachys recta L.) | . | . | . | + | . | . | . | . | 1 |
| Купальница европейская (Trollius europaeus L.) | + | . | + | . | + | + | + | + | 4 |
| Черника обыкновенная (Vaccinium myrtillus L.) | . | . | . | . | + | + | . | + | 6 |
| Брусника обыкновенная (V. vitis-idaea L.) | . | . | . | . | + | + | . | . | 6 |
| Чемерица черная (Veratrum nigrum L.) | . | + | + | + | . | . | . | . | 5 |
| Вероника седая (Veronica incana L.) | . | . | . | + | . | . | . | . | 1 |
| Вероника ненастоящая (V. spuria L.) | . | + | . | + | . | . | . | . | 1 |
| Горошек кашубский (Vicia cassubica L.) | . | . | . | + | . | . | . | + | 2 |
| Горошек гороховидный (V. pisiformis L.) | . | + | . | + | . | . | + | + | 9 |
| Горошек тонколистный (V. tenuifolia) | . | . | . | + | . | . | . | . | 5 |
| Барвинок травянистый (Vinca herbacea Waldst. & Kit.) | + | . | . | . | . | . | . | . | 1 |
| Ластовень ласточкин (Vincetoxicum hirundinaria Medik.) | . | . | . | + | . | . | . | . | 26 |
Среди перечисленных редких наиболее многочисленны виды трех групп: тепло- и светолюбивые южные по происхождению субпонтические (лесостепные) (39%), мезофильные неморальные виды широколиственных лесов (24%) и ксерофильные понтические (степные) виды (10%). Можно считать сообщества изучаемого типа своеобразными проводниками лесостепных и степных видов к северу, в зону широколиственных лесов. К юго-востоку, напротив, такие сообщества становятся рефугиумами некоторых северных, в том числе бореальных и суббореальных видов.
Некоторые из перечисленных выше редких видов растений проявляют высокую верность выборке ксеро-мезофитных лесов на статистически значимом уровне: венечник ветвистый (φ = 37.9), оленник Ривиниуса (20.2), наперстянка крупноцветковая (31.5), дремлик чемерицевидный (29.4), гладыш широколистный (52.9), чина гороховидная (30.5), лилия царские кудри (34.7), любка двулистная (20.5), лапчатка белая (60.4), вишня кустарниковая (21.0), слива колючая (30.5), поповник щитковый (49.3), жостер слабительный (23.6), серпуха красильная (59.0), ластовень ласточкин (29.7). Наиболее высокую константность в изучаемой выборке описаний имеют венечник ветвистый (20%), наперстянка крупноцветковая (35%), дремлик чемерицевидный (18%), гладыш широколистный (37%), чина гороховидная (14%), лилия царские кудри (19%), лапчатка белая (47%), слива колючая (15%), поповник щитковый (35%), жостер слабительный (19%), серпуха красильная (44%), ластовень ласточкин (26%). Учитывая низкую значимость административных границ при расселении видов, ксеро-мезофитные леса на Юго-Западе России можно считать единым рефугиумом многочисленных регионально-редких видов сосудистых растений.
ЗАКЛЮЧЕНИЕ
В ценофлоре ксеро-мезофитных широколиственных лесов Юго-Запада России отмечено 508 видов сосудистых растений и 30 видов мохообразных. Сообщества характеризуются наибольшей флористической насыщенностью среди всех групп широколиственных лесов в изучаемом регионе, что было установлено на основе сравнения 1296 геоботанических описаний широколиственных лесов Юго-Запада России. Высокое флористическое разнообразие ксеро-мезофитных сообществ является отражением экологической и ботанико-географической гетерогенности ценофлоры, что демонстрируют спектры геоэлементов/полизональных групп и экобиоморф. Наибольшие значения параметров флористического разнообразия – флористической насыщенности и его выровненности – наблюдаются у субконтинентальных сообществ ксеро-мезофитных лесов в северной части их ареала в местообитаниях с наиболее бедными минеральным азотом умеренно увлажненными почвами.
В ценофлоре ксеро-мезофитных широколиственных лесов Юго-Запада России выявлено 79 видов сосудистых растений, занесенных в Красные книги России (2008) или субъектов РФ на Юго-Западе страны. Среди охраняемых наиболее многочисленны виды трех групп: тепло- и светолюбивые южные по происхождению субпонтические (лесостепные), мезофильные неморальные виды широколиственных лесов и ксерофильные понтические (степные) виды. Учитывая низкую значимость административных границ при расселении видов, ксеро-мезофитные леса на Юго-Западе России можно считать своеобразными проводниками лесостепных и степных видов к северу, в зону широколиственных лесов; к юго-востоку, напротив, такие сообщества становятся рефугиумами некоторых северных, в том числе бореальных и суббореальных видов.
Проведенное исследование имеет важно значение в связи с подготовкой к реализации общероссийского проекта по созданию фундаментальной сводки по растительности России и готовящимся переизданием Зеленой книги Брянской области, расположенной на Юго-Западе страны.
Список литературы
Атлас Курской области. М.: Федер. служба геодезии и картографии России, 2000. 48 с.
Булохов А.Д. Синтаксономия лесной растительности Южного Нечерноземья. 1. Порядок Quercetalia pubescentis Br.-Bl. 1931 // Биологические науки. М. 1991. 48 с.
Булохов А.Д. Фитоиндикация и ее практическое применение. Брянск: БГУ, 2004. 245 с.
Булохов А.Д., Семенищенков Ю.А. Ботанико-географические особенности ксеро-мезофитных широколиственных лесов союза Quercion petraeae Zólyomi et Jakucs ex Jakucs 1960 Южного Нечерноземья России // Бюллетень Брянского отделения РБО. 2013. № 1 (1). С. 10–24.
Булохов А.Д., Соломещ А.И. Эколого-флористическая классификация лесов Южного Нечерноземья России. Брянск: БГУ, 2003. 359 с.
Булохов А.Д., Харин А.В. Растительный покров Брянска и его пригородной зоны. Брянск: БГУ, 2008. 310 с.
Василевич В.И. Разнообразие растительности в пределах ландшафта // Биологическое разнообразие: подходы к изучению и сохранению. СПб., 1992. С. 34–41.
Зеленая книга Брянской области (растительные сообщества, нуждающиеся в охране) / Булохов А.Д., Семенищенков Ю.А., Панасенко Н.Н., Анищенко Л.Н., Федотов Ю.П., Аверинова Е.А., Харин А.В., Кузьменко А.А., Шапурко А.В. Брянск: БГУ, 2012. 144 с.
Красная книга Российской Федерации (растения и грибы) / Под ред. Трутнева Ю.П. М.: Товарищество научных изданий КМК, 2008. 855 с.
Исаченко Т.И., Лавренко Е.М. Ботанико-географическое районирование // Растительность европейской части СССР. Л., 1980. С. 10–20.
Лебедева Н.В., Дроздов Н.Н., Криволуцкий Д.А. Биологическое разнообразие: уч. пособие для студентов высш. уч. заведений. М.: Гуманитар. изд. центр Владос, 2004. 432 с.
Миркин Б.М., Наумова Л.Г. Современное состояние основных концепций науки о растительности. Уфа: Гилем, 2012. 488 с.
Морозова О.В. Леса заповедника “Брянский лес” и Неруссо-Деснянского Полесья (синтаксономическая характеристика). Брянск, 1999. 98 с.
Морозова О.В. Таксономическое богатство флоры Восточной Европы: факторы пространственной дифференциации. М.: Наука, 2008. 328 с.
Онипченко В.Г. Функциональная фитоценология: синэкология растений. М: URSS, 2013. 576 с.
Природа и природные ресурсы Брянской области / Под ред. Л.М. Ахромеева. Брянск: Курсив, 2012. 320 с.
Семенищенков Ю.А. Фитоценотическое разнообразие Судость-Деснянского междуречья. Брянск: БГУ, 2009. 400 с.
Семенищенков Ю.А. Сообщества союза Aceri tatarici–Quercion roboris Zólyomi et Jakucs ex Jakucs 1960 в бассейне реки Ворсклы (Белгородская область) // Вестник Тверского гос. ун-та. Сер.: Биология и экология. 2012. Вып. 28. № 25. С. 54–62.
Семенищенков Ю.А. Эколого-флористическая классификация как основа ботанико-географического районирования и охраны лесной растительности бассейна Верхнего Днепра (в пределах Российской Федерации): дис. … докт. биол. наук: 03.02.01. Уфа, 2016. 558 с.
Семенищенков Ю.А. Экологические эффекты в формировании флористического состава и их отражение в синтаксономии пойменных дубрав бассейна Верхнего Днепра // Растительность России. 2020. № 39. С. 24–36. https://doi.org/10.31111/vegrus/2020.39.26
Семенищенков Ю.А., Полуянов А.В. Остепненные широколиственные леса союза Aceri tatarici–Quercion Zólyomi 1957 на Cреднерусской возвышенности // Растительность России. 2014. № 24. С. 101–123.
Семенищенков Ю.А., Телеганова В.В. Синтаксономия и экология ксеромезофитных дубрав долины Оки в пределах Калужской области // Уч. зап. Орловского гос. ун-та. 2013. № 3 (53). С. 132–138.
Семенищенков Ю.А., Шапурко А.В. Распространение и особенности экологии гибрида Pulmonaria × notha A. Kern (Boraginaceae) в Брянской области // Вестник Воронежского гос. ун-та. Сер.: Химия. Биология. Фармация. 2018. № 2. С. 246–254.
Юрцев Б.А. Эколого-географическая структура биологического разнообразия и стратегия его учета и охраны // Биологическое разнообразие: подходы к изучению и сохранению. СПб., 1992. С. 7–21.
Cardinale B.J., Nelson K., Palmer M.A. Linking species diversity to the functioning of ecosystems: on the importance of environmental context // Oikos. 2000. V. 91. № 1. P. 175–183. https://doi.org/10.1034/j.1600-0706.2000.910117.x
Chytrý M., Tichý L., Holt J., Botta-Dukat Z. Determination of diagnostic species with statistical fidelity measures // Journal of Vegetation Science. 2002. V. 13. № 1. P. 79–90. https://doi.org/10.1658/1100-9233(2002)013[0079:DODSWS] 2.0.CO;2
Duffy J.E. Biodiversity and ecosystem function: the consumer connection // Oikos. 2002. V. 99. № 2. P. 201–219. https://doi.org/10.1034/j.1600-0706.2002.990201.x
Ellenberg H., Weber H. E., Düll R., Wirth W., Paulißen D. Zeigerwerte von Pflanzen in Mitteleuropa. 2 Aufl. Göttingen, 1992. 258 p.
Goncharenko I.V., Semenishchenkov Y.A., Tsakalos J.L., Mucina L. Thermophilous oak forests of the steppe and forest-steppe zones of Ukraine and Western Russia // Biologia. 2020. V. 75. № 1. https://doi.org/10.2478/s11756-019-00413-w
Grime J.P. Declining plant diversity: empty niches or functional shifts? // Journal of Vegetation Science. 2009. V. 13. № 4. P. 457–460. https://doi.org/10.1111/j.1654-1103.2002.tb02072.x
Hooper D., Vitousek P. The effect of plant composition and diversity on ecosystem processes // Science. 1997. V. 277. № 5330. P. 1302–1305. https://doi.org/10.1126/science.277.5330.1302
Hooper D.U., Chapin F.S., Ewel J.J., Hector A., Inchausti P., Lavorel S., Lawton J.H., Lodge D.M., Loreau M., Naeem S., Schmid B., Setälä H., Symstad A.J., Vandermeer J., Wardle D.A. Effects of biodiversity on ecosystem functioning: a consensus of current knowledge // Ecological Monographs. 2005. V. 75. № 1. P. 3–35. https://doi.org/10.1890/04-0922
Kottek M., Grieser J., Beck Ch., Rudolf B., Rubel F. World Map of the Köppen-Geiger Climate Classification Updated // Meteorologische Zeitschrift. 2006. V. 15. № 3. P. 259–263. https://doi.org/10.1127/0941-2948/2006/0130
Magurran A.E. Ecological diversity and its measurement. New-York, 1988. 179 p. https://doi.org/10.1007/978-94-015-7358-0
Margalef D.R. Information theory in ecology // General Systems. 1958. V. 3. P. 36–71.
McCune B., Mefford M.J. PC-ORD. Multivariate analysis of Ecological Data, Version 5. Oregon, 2006. 28 p.
Palmer M.W. Variation in species richness: towards a unification of hypothesis // Folia Geobotanica et Phytotaxonomica. 1994. V. 29. № 4. P. 511–530.
The Euro + Med PlantBase – the information resource for Euro-Mediterranean plant diversity. URL: http://www. emplantbase.org/home.html. Дата обращения: 08.04.2021.
Tichý L. JUICE, software for vegetation classification // Journal of Vegetation Science. 2002. V. 13. P. 451–453. https://doi.org/10.1658/1100-9233(2002)013[045:JSFVC] 2.0.CO;2
Walter H. Vegetationzonen und Klima: der okologisher Gliederung der Biogeosphare. Studgart; Ulmar, 1977. 309 p.
Whittaker R.H. Evolution and measurement of species diversity // Taxon. 1972. V. 21. P. 213–251.
Zobel M. Plant species coexistence – the role of historical, evolutionary and ecological factors // Oikos. 1992. V. 65. № 2. P. 312–320. https://doi.org/10.2307/3545024
Zobel M., van der Maarel E., Dupre C. Species pool: the concept, its determination and significance for community restoration // Applied Vegetation Science. 1998. V. 1. P. 55–66. https://doi.org/10.1111/jvs.12333
Дополнительные материалы отсутствуют.