

Лесоведение, 2022, № 6, стр. 643-657
Ценотическое разнообразие и долговременная динамика массива “Вепсский лес”
Е. В. Шорохова a, b, *, А. А. Корепин a, Е. А. Капица a, Г. В. Березин a, А. А. Шорохов c, М. А. Шорохова a
a Санкт-Петербургский государственный лесотехнический университет имени С.М. Кирова
194021 Санкт-Петербург, Институтский пер., д. 5, Россия
b Институт леса КарНЦ РАН,
185910 Петрозаводск, ул. Пушкинская, д. 11, Республика Карелия, Россия
c ООО “ИКЕА Индастри Тихвин”
187556 Ленинградская обл., Тихвин, Шведский проезд, д. 15, Россия
* E-mail: shorohova@es13334.spb.edu
Поступила в редакцию 26.12.2021
После доработки 19.02.2022
Принята к публикации 07.06.2022
- EDN: BNGSZW
- DOI: 10.31857/S0024114822060109
Аннотация
В свете проблемы сохранения устойчивости наземных экосистем в условиях изменения климата особенно важно понимание механизмов поддержания биоразнообразия в массивах эталонных лесов, не затронутых хозяйственной деятельностью. Целью работы являлся синтез комплексных многолетних исследований структурно-функционального разнообразия и динамики массива коренных среднетаежных лесов “Вепсский лес”. В задачи входил анализ: 1) режима естественных нарушений; 2) представленности биогеоценозов различных форм динамики, сопряженных с разными лесорастительными условиями, а также ветровальных окон; 3) возрастной структуры лесов массива; 4) динамики породного состава, запаса древесины, числа деревьев, крупных древесных остатков, естественного возобновления и отпада древостоя в массиве в целом и по типам биогеоценозов; 5) факторов, определяющих специфику пространственно-временной организации изучаемого лесного массива. Решение поставленных задач возможно лишь в рамках комплексного подхода. Использовали данные аэрофотосъемки, описаний на трансектах, маршрутных обследований, картирования состава и возрастной структуры лесов, типов местообитаний и почв, а также описаний на регулярно заложенных постоянных круговых пробных площадей фиксированного радиуса. Повторяемость и интенсивность ветров обусловливает структурное разнообразие и динамику изучаемого лесного массива. В свою очередь, ценотическое разнообразие массива определяет интенсивность ветровальных нарушений, характер и скорость восстановления сообществ после них. Преобладающие по площади ельники на дренированных местообитаниях подвержены ветровалам средней и сильной интенсивности – 19% их общей площади составляют ветровальные окна. В большинстве своем (43%) они имеют относительно разновозрастную структуру с преобладанием возрастного поколения 161–200 лет. Древостои, имеющие абсолютно разновозрастную структуру (15%), приурочены к недостаточно дренированным местообитаниям. Средний запас древостоев массива с 1991 по 2019 гг. уменьшился на 15% (с 321 до 274 м3 га–1) в результате ветровалов. Запас крупных древесных остатков, представленных, в основном, валежом и зависшими деревьями, составил в среднем 71% от запаса живых деревьев. Его вариация отражала фазы динамики древостоя в связи с ветровальными нарушениями. Число биогеоценозов, находящихся в фазе стабилизации, уменьшилось, в том время как доля биогеоценозов, находящихся в дигрессионных и демутационных фазах динамики, увеличилась. Преобладает “верховой” отпад, т.е. отмирают, в основном, более крупные деревья. Среднегодовой отпад на момент последнего учета составил 3.5% от общего запаса древостоя. Доля ели в составе древостоев массива остается достаточно стабильной. Результаты многолетних исследований в массиве “Вепсский лес” могут быть использованы для решения множества задач, в том числе для разработки оптимальных сценариев ведения лесного хозяйства, а также для сохранения и/или восстановления биоразнообразия в управляемых лесах.
Коренные леса, развивающиеся без существенного воздействия человека в течение времени, сопоставимого с предельным биологическим возрастом преобладающей древесной породы, являются эталонами устойчивых лесных сообществ (Сукачев, Дылис, 1964; Дыренков, 1984; Пугачевский, 1992; Frelich, Reich, 2003; Potapov et al., 2017; Watson et al., 2018). Вне зависимости от географического положения и флористического состава такие леса представляют собой множество несинхронно развивающихся элементов подвижной мозаично-ярусной структуры (patch mosaic) (Коротков, 1991; Бекмансуров и др., 2004; Manabe et al., 2009; Chambers et al., 2013; Král et al., 2014; Martin et al., 2018). Мозаики разных уровней качественно различаются по составляющим их компонентам, структуре, ведущим факторам и формам динамики, степени экологической однородности и характерному времени протекания ведущих процессов (Мазинг, 1988; Юрцев, 1992; Маслов, 1990; Spies, Turner, 1999; Trotsiuk et al., 2014; Meigs et al., 2017). Понимание естественных процессов, действующих внутри лесных сообществ разных уровней с оценкой их устойчивости к внешним воздействиям, – необходимое условие при планировании хозяйственной деятельности, нацеленной на сохранение всех экосистемных функций леса (Пугачевский, 2013; Лукина и др., 2020; Gauthier et al., 2021).
Структура и динамика относительно простого по видовому составу древесного яруса и сложных по структуре коренных таежных лесов хорошо исследована на уровне отдельных биогеоценозов (БГЦ) (Дыренков, 1984; Пугачевский, 1992; Shorohova et al., 2009, 2011; Martin et al. 2018, 2021). Лесной массив – пространственно единый комплекс лесных биогеоценозов на территории, однородной в ландшафтном и хозяйственно-историческом отношениях (Федорчук и др., 2012), – остается наименее изученным объектом в иерархическом ряду лесных экосистем (Федорчук и др., 2012; Venier et al., 2018; Martin et al., 2021). Как в классических, так и в современных работах подчеркивается, что понятия “девственный”, “коренной”, “старовозрастный лес” корректнее относить не к отдельным БГЦ, а к лесным массивам (Морозов, 1930; Leibungut, 1982; Громцев, 2007; Martin et al., 2020, 2021). Выявление режимов естественных нарушений – событий, разрушающих или изменяющих структуру экосистемы, сообщества или популяции, изменяющих доступность ресурсов или субстрата или трансформирующих физическую среду (Pickett, White, 1985), – возможно только на уровне лесного массива (Shorohova et al., 2022). Последними достижениями лесной науки и в некоторой степени практики являются системы лесного хозяйства, направленные на сохранение устойчивого функционирования социально-экологических систем в условиях глобального изменения климата – ecosystem forest management с употребляемыми реже синонимами resilient forest management и adaptive forest management. Эти системы предполагают адаптацию к режимам нарушений и ведение лесного хозяйства на уровне лесного массива (Messier et al., 2013; Gauthier et al., 2021; Shorohova et al., 2022).
Режим нарушений, биоразнообразие, устойчивость и динамику естественно развивающихся лесных массивов характеризуют по ряду показателей. Для описания режима нарушений лесного массива используют следующие параметры: а) вид нарушения (пожар, ветровал, усыхание древостоев в результате воздействия насекомых и др.); б) пространственные характеристики (площадь нарушения, произошедшего за одно событие или за определенный период; пространственное распределение и сопряженность нарушенных структурных элементов, в связи с, например, экотопическими условиями); в) временны̀е характеристики (частота нарушений, выражаемая как число событий за единицу времени; интервал между нарушениями); г) специфичность нарушения (по видовому составу, по сукцессионному состоянию экосистем, по элементам рельефа); д) величина нарушения (severity, степень нарушенности экосистемы) и др. (Pickett, White, 1985; Громцев, 2007; Shorohova et al., 2021).
Группа показателей, характеризующих структурно-динамическое разнообразие массива, включает площадь и пространственное расположение различных БГЦ и структурных элементов, например, “окон” – прорывов в пологе леса (M-cCarthy, 2001; Shorohova et al., 2011; Федорчук и др., 2012). Выделяют БГЦ, различающиеся по видовому составу растительных сообществ, пространственной структуре, сукцессионному состоянию и др. Распределение общей площади окон по размерам и времени образования может использоваться для реконструкции параметров прошлых нарушений и описания динамических процессов, выраженных на уровне комплекса лесных биогеоценозов. Возрастная структура основных лесообразующих пород характеризует динамическое состояние и устойчивость лесного массива, степень равномерности или дискретности возобновительного процесса в целом по массиву (Дыренков, 1984; Пугачевский, 1992). Общее количество, породный состав и состояние подроста отражают специфику возобновительного процесса и реакции экосистем различного динамического состояния на нарушения.
Динамика лесного массива описывается изменением общего запаса древесины, средних величин запаса на гектар и среднего возраста и их пространственной изменчивости, породного состава древостоев, структуры популяций, а также изменением соотношения разного рода структурных единиц (Федорчук и др., 1998, 2012). Запас и структурное разнообразие крупных древесных остатков (КДО) отражают историю развития массива и слагающих его БГЦ (Федорчук и др., 2011; Шорохова и др., 2021). Их характеристики в совокупности с характеристиками сообществ живых организмов, связанных с ними на всех этапах их разложения, рассматриваются как один из показателей устойчивости лесных экосистем (Устойчивость лесов …, 2018).
Целью работы является синтез комплексных многолетних исследований структурно-функционального разнообразия и динамики массива коренных среднетаежных лесов “Вепсский лес”. В задачи входил анализ: 1) режима естественных нарушений; 2) представленности БГЦ различных форм динамики, сопряженных с разными лесорастительными условиями, а также ветровальных окон; 3) возрастной структуры лесов массива; 4) динамики породного состава, запаса древесины, числа деревьев, крупных древесных остатков (КДО), естественного возобновления и отпада древостоя в массиве в целом и по типам БГЦ; 5) факторов, определяющих специфику пространственно-временной организации изучаемого лесного массива. На основании литературных данных и ранее полученных результатов исследования массива “Вепсский лес” была сформулирована основная рабочая гипотеза. Предполагали, что ценотическое разнообразие массива, выраженное как площадное соотношение структурных единиц – древостоев различного динамического состояния (вариантов возрастной структуры и фаз возрастной циклической динамики), сопряженных с различными лесорастительными условиями, – определяет характер, направление и скорость сукцессионных процессов. Работа продолжает серию публикаций по результатам долговременных исследований в резервате “Вепсский лес”. Данные, представленные в статье, дополняют наблюдения ученых по ряду показателей (Федорчук и др., 1998, 2002, 2011, 2012, 2014; Березин и др., 2020; Корепин и др., 2020) и включают новые показатели, связанные с характеристикой КДО (Шорохова и др., 2021).
ОБЪЕКТЫ И МЕТОДИКА
В качестве объекта исследования выбран лесной массив, находящийся в резервате “Вепсский лес” одноименного природного парка, расположенного в Балтийско-Белозерском таежном районе (60°12′19.8″ N 35°08′04.7″ E, площадь 889 га, рис. 1). Массив находится на высоте 220–260 м над уровнем моря, на приподнятом участке древнего карбонового плато (Федорчук и др., 1998). Среднегодовая температура составляет +2.8°C (1950–2021), среднегодовое количество осадков – 750 мм. На территории исследуемой части резервата, на площади около 600 га сохранились незатронутые хозяйственной деятельностью естественно развивающиеся преимущественно еловые леса; около 300 га занимают болотные экосистемы (Федорчук и др., 2012). На XII-ом международном ботаническом конгрессе в Ленинграде в 1975 году лесной массив резервата “Вепсский лес” признан эталоном природы средней тайги. В лесном массиве преобладают относительно разновозрастные еловые леса на дренированных и недостаточно дренированных почвах на моренных суглинках и двучленных наносах (серии типов леса – черничная, долгомошно-черничная), заболоченные торфянистые и болотно-торфяные почвы (серии типов леса – сфагново-черничная, сфагново-майниковая). Возраст основного поколения еловых древостоев достигает 200–300, а отдельных деревьев – более 400 лет (Федорчук и др., 1998).
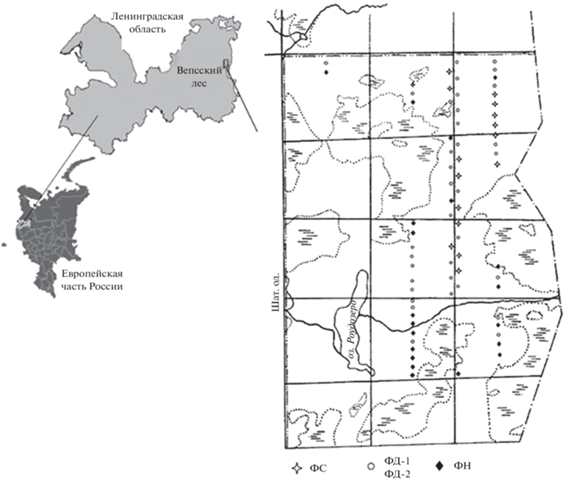
Рис. 1.
Расположение объекта исследований и регулярно заложенных постоянных круговых пробных площадей (КПП) в рамках квартальной сети. На схеме показано пространственное расположение биогеоценозов, находящихся в разных фазах сукцессионной динамики древостоя по данным инвентаризации 2018–2019 гг. ФС – фаза стабилизации, ФД – фаза дигрессии с двумя подфазами – дигрессии запаса (ФД1) и активного формирования нового поколения древостоя (ФД2), ФН – фаза нарастания запаса.
Режим ветровальных нарушений изучали на основании данных аэрофотосъемки, описаний на трансектах и маршрутных обследований по просекам и визирным линиям. Число и давность пожаров реконструировали с помощью дендрохронологических методов: 1) анализа радиального прироста деревьев и 2) реконструкции пожаров по пожарным подсушинам.
Представленность различных БГЦ изучали на основании результатов картирования состава и возрастной структуры лесов, типов местообитаний и почв, а также по материалам наблюдений на 74-х регулярно заложенных постоянных круговых пробных площадях (КПП) фиксированного радиуса (17.85 м). Представленность ветровальных окон определяли на основании анализа спектрозональных аэрофотоснимков разных лет и учетов на трансектах.
Естественную динамику породного состава и запаса древостоя, естественного возобновления, КДО и древесного отпада в массиве в целом и по типам БГЦ, а также породную и возрастную структуры популяций лесообразующих древесных пород изучали по данным учетов на КПП. 74 КПП (каждая площадью 0.1 га) были заложены в 1991 году в еловых (ель европейская (Picea abies (L.) Karst), ель финская (P. fennica (P. abies × P. obovata Ledeb.)) и смешанных древостоях, имеющих значительное участие ели (более 30% по запасу) (рис. 1). Для КПП определяли тип леса, вариант возрастной структуры и фазу динамики древостоя. Выделяли условно одновозрастные (УО), относительно разновозрастные (ОР) и абсолютно разновозрастные (АР) древостои (Дыренков, 1984). Вариант возрастной структуры древостоя определяли на характерных участках вблизи пробных площадей глазомерно с уточняющим бурением отдельных деревьев. Отнесение БГЦ к одному из вариантов возрастной структуры имело предварительный характер и требовало дальнейшего уточнения. Фаза зрелости (ФЗ) характеризовалась относительно стабильной величиной запаса древостоя, а также равной вероятностью основных видов отпада (сухостоя или ветровала-бурелома). В фазе дигрессии (ФД1) наблюдалось уменьшение общего запаса древостоя за счет увеличения величины отпада (более 2–3% запаса древостоя в год), преобладание “верхового” отпада (более крупномерных деревьев). В конце фазы (ФД2) характерно преобладание в отпаде ветровальных и буреломных стволов, неоднородность полога крон деревьев (обилие “окон”), интенсивное появление и активное развитие подроста. Для фазы нарастания запаса характерно увеличение прироста и запаса древостоя, “низовой” отпад (преобладают тонкомерные деревья) преимущественно в виде сухостоя с небольшим объемом (1.0–1.5% от запаса древостоя в год), высокая сомкнутость древостоя, незначительная численность и медленное развитие подроста (Федорчук и др., 2011).
Характеристики древостоя получены с использованием стандартных методов таксации на основании перечетов деревьев по породам и элементам леса (ярусам) и (или) возрастным группам через каждые пять лет. Возрастное поколение деревьев определяли глазомерно, с уточняющим бурением 2–3 деревьев у шейки корня. Высоту древостоя элемента леса (возрастного поколения каждой древесной породы) определяли по графику высот, построенному по результатам измерения высоты и диаметра 25–55 деревьев. Запас древесины рассчитывали по объемным таблицам по ступеням толщины и разрядам высот. Естественное возобновление и КДО учитывали на двух перпендикулярных трансектах шириной два (четыре для КДО) метра, проходящих через центры пробных площадей. Анализировали все экземпляры древесных растений основных лесообразующих пород, попадающие в границы трансекты по следующим категориям размерности по высоте: до 0.05 м, 0.06–0.25 м, 0.26–0.5 м, 0.51–1.5 м, более 1.5 м. По внешним признакам особи подразделяли на две категории: жизнеспособные и нежизнеспособные. КДО учитывали по породам, положению (сухостой, валеж, зависшие стволы и пни) и классам разложения.
Более подробно методики описаны в публикациях Федорчука с соавторами (1998, 2012) и Шороховой с соавторами (2021).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Режим естественных нарушений. На территории лесного массива преобладающим видом нарушений являются ветровалы, приводящие к варьирующей во времени и пространстве оконной динамике лесов. На аэрофотоснимках разных лет не отмечено заметных прорывов в древесном пологе до 1980-х годов. В то же время возрастная структура массива, а именно значительная общая доля условно одновозрастных древостоев одного возрастного поколения предположительно отражает влияние сильных ветровалов, произошедших в период с 1800 по 1830 гг. (Федорчук и др., 2002). Начало очередного периода активизации ветровальных нарушений отмечено в 1982 г. Образовавшиеся в 1980–1990-х годах ветровальные окна увеличились в последующие десятилетия, отмечено увеличение размера существующих и появление новых прорывов в древесном пологе. В целом в массиве преобладают ветровалы средней интенсивности. Пространственное расположение окон отражает направление наиболее сильных ветров – большинство окон вытянуты с северо-запада на юго-восток. Наибольшая площадь окон отмечена в еловых древостоях, однако их относительная площадь максимальна в осинниках, где она достигает 55% площади, тогда как для ельников эта величина составляет 14%. Наиболее уязвимыми к ветровалам оказались ельники черничные: 19% их площади составляют окна. Более половины крупных (более 0.25 га) окон также приурочено к ельникам черничным. Наиболее подвержены ветровалам относительно разновозрастные (16% их площади) и условно одновозрастные (12%) ельники; в абсолютно разновозрастных древостоях окна занимают около 10% площади (Федорчук и др., 2012). Пожары 1869 и 1952 гг., зарегистрированные вблизи озер, носят локальный характер (Федорчук и др., 2012). В последнее годы наблюдаются куртины усыхания елей.
Ветровальная динамика, наблюдаемая в массиве “Вепсский лес”, характерна для таежных лесных массивов с преобладанием ельников (Скворцова и др., 1983; Георгиевский, 1992; Пугачевский, 1992; Сукцессионные процессы …, 1999). Отрывочные сведения об интервале между ветровальными нарушениями в таежных лесах свидетельствуют о его высокой вариабельности (Смолоногов, 1995; Waldron et al., 2013; De Grandpré et al., 2018). Интенсивность ветровалов также отличается вариабельностью в зависимости от взаимодействия между скоростью ветра, топографией и почвенными условиями, историей нарушений и характеристиками древостоев и лесных ландшафтов (Скворцова и др., 1983; Everham, Brokaw, 1996; Ruel, 2000; Ulanova 2000; Saad et al., 2017). Во многих работах отмечается, что ветровалы средней интенсивности являются характерной чертой сообществ коренных таежных лесов (Сукцессионные процессы …, 1999; Girard et al., 2014).
Идея подражания природе и ведения лесного хозяйства в соответствии с режимами естественных нарушений широко обсуждается (Attiwill, 1994; Kuuluvainen, 2002; Gauthier et al., 2009). В вовлеченных в хозяйственный оборот лесных массивах с преобладанием ветровальных нарушений средней интенсивности наиболее перспективным представляется сочетание сплошнолесосечного хозяйства с применением выборочных, в том числе оконных (группово-выборочных) рубок, а также лесное хозяйство, нацеленное на сохранение лесной среды – variable retention felling (Shorohova et al., 2008; Koivula et al., 2014; Крышень и др., 2020). Поддержание ценотического разнообразия и устойчивости лесов при этом достигается определенным сочетанием на уровне лесного массива участков с различными режимами, интенсивностью и сроками хозяйственного вмешательства. Так, для наименее дренированных участков, в частности, чернично-сфагновых, майниково-сфагновых и долгомошно-черничных серий типов леса, следует выбирать самый жесткий режим ограничений по рубкам, ориентируясь в первую очередь на формирование в таких лесорастительных условиях наибольшей разновозрастности и пространственной неоднородности древостоев. Напротив, на дренированных участках, прежде всего, на вершинах холмов, перспективен вариант мелкоконтурных, сплошных (котловинных) рубок. При этом сроки их проведения должны быть максимально разнесены по десятилетним периодам для получения в долгосрочной перспективе равномерного распределения древостоев по группам возраста. Между двумя описанными крайними вариантами возможны выборочные формы хозяйства, представленность которых будет определяться степенью ландшафтной неоднородности лесного массива, а также целевым уровнем его устойчивости и структурного разнообразия.
Структура и разнообразие БГЦ. В исследуемом массиве преобладают ельники (82% – по данным лесоустройства и 99% – по данным учетов на КПП). При лесоустройстве выделены также сосняки (16%) и березняки (1%). Осинники составляют 1%.
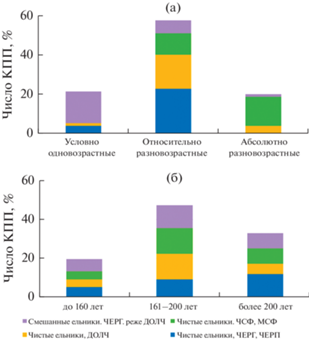
Преобладающие в исследуемом массиве ельники на дренированных местообитаниях подвержены ветровалам средней интенсивности и в большинстве имеют относительно разновозрастную структуру (43%) с преобладанием поколения 161–200 лет (рис. 2). Древостои, имеющие абсолютно разновозрастную структуру (15%), приурочены к недостаточно дренированным местообитаниям. Результаты, характеризующие представленность БГЦ разной возрастной структуры древостоя (рис. 2), полученные по материалам КПП, мало отличаются от материалов картирования (Федорчук и др., 2012). Вопрос о вероятности смены варианта возрастной структуры в процессе возрастной динамики БГЦ в исследуемом лесном массиве остается открытым до следующего учетного периода.
Рис. 2.
Ценотическое разнообразие лесного массива “Вепсский лес”. (а) Распределение биогеоценозов по вариантам возрастной структуры древостоя и типам леса, (б) Распределение биогеоценозов по возрасту основного поколения ели и типам леса.
Породный состав древостоев массива и преобладание ели обусловливает породный состав КДО, представленный в основном елью: на долю березы, осины и сосны приходится в совокупности не более 12% от общего объема КДО. Интересно, что запас КДО, составляющий в среднем 195 м3 га–1 (71% от запаса живых деревьев) и варьирующий от 30 до 532 м3 га–1, отражает фазы динамики древостоя в связи с ветровальными нарушениями. Ветровальная динамика древостоев массива резервата “Вепсский лес” обусловливает также структуру пула КДО, который представлен в основном валежом и зависшими деревьями. Более высокие объемы КДО в древостоях черничного типа леса, по сравнению с древостоями долгомошно-черничного и чернично-сфагнового типов леса, и в древостоях, находящихся в фазе дигрессии запаса, по сравнению с древостоями фаз стабилизации и нарастания запаса, отражают особенности возрастной динамики древостоев массива в связи с нарушениями (Шорохова и др., 2021).
Динамика. Анализ многолетней динамики состояния массива показывает уменьшение среднего запаса древостоев массива на 15% (табл. 1, рис. 3). Произошло также изменение соотношения площадей древостоев, находящихся в разных фазах возрастной циклической динамики (табл. 2).
Таблица 1.
Количество КПП (шт.) по изменению запаса древостоя
| Изменение запаса, % | Годы учета | ||||
|---|---|---|---|---|---|
| 1991–1997 | 1997–2001 | 2001–2008 | 2008–2013 | 2013–2019 | |
| ≤–10 | 7 | 12 | 14 | 16 | 22 |
| ≥+10 | 4 | 6 | 9 | 16 | 14 |
| от –10 до +10 | 63 | 56 | 51 | 42 | 38 |
Рис. 3.
Динамика (а) среднего запаса древостоя по породам, среднего запаса (б, г) и отпада (в, д) древостоя в зависимости от лесорастительных условий (б, в) и вариантов возрастной структуры древостоя (г, д) в массиве “Вепсский лес” с 1991 по 2019 год, м3 га–1. Е – Ель (Picea abies (L.) H. Karst), Б – Береза (Betula pendula Roth, B. pubescens Ehrh.), Ос – Осина (Populus tremula L.), C – Сосна (Pinus sylvestris L.). ДОЛЧ – долгомошно-черничные; ЧЕРГ – черничные типы леса на дренированных суглинках и двучленных наносах, ЧЕРП – то же на дренированных песках и супесях; ЧСФ – чернично-сфагновые; МСФ – майниково-сфагновые типы леса. АР – абсолютно разновозрастные; ОР (СП) – относительно разновозрастные, смена пород; Уо – условно одновозрастные; Уо (СП) – условно одновозрастные, смена пород; ОР – относительно разновозрастные.
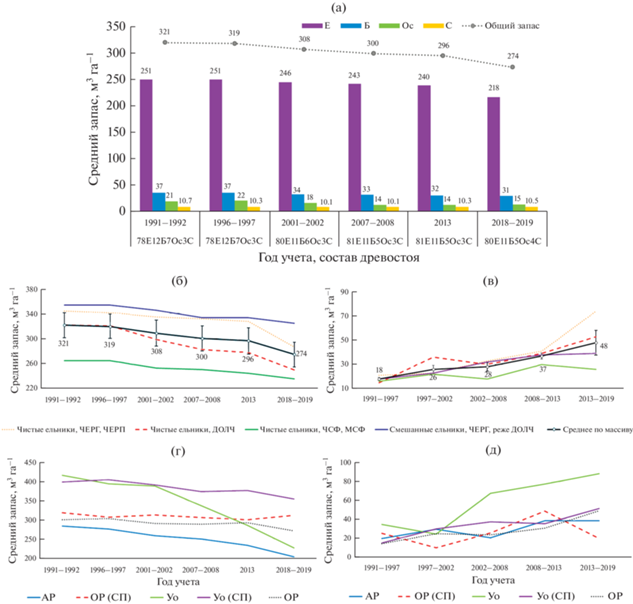
Таблица 2.
Распределение КПП по фазам динамики БГЦ в резервате “Вепсский лес” за период с 1991 по 2019 годы учета
| Количество КПП по фазам динамики | |||
|---|---|---|---|
| Годы учета | 1991–1992 | 2007–2008 | 2018–2019 |
| Фаза стабилизации | 46 | 32 | 17 |
| Фаза дигрессии | 14 | 14 | 14 |
| Фаза дигрессии-2 | 0 | 13 | 25 |
| Фаза нарастания | 14 | 15 | 18 |
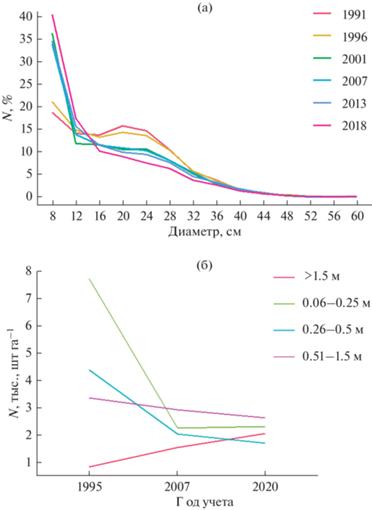
Общее уменьшение среднего запаса древостоя в первую очередь связано с устойчивым снижением запаса ели, особенно усилившимся в последние годы (рис. 3). Сравнение среднего диаметра живых и перешедших в категорию “отпад” деревьев позволяет сделать вывод о преобладании в массиве “верхового” отпада (табл. 3). Процесс перехода деревьев из категории “живые” в категорию “отпад” ускорялся в последние годы, о чем свидетельствует рост величины среднегодового отпада, который на момент последнего учета составил 3.5% от общего запаса древостоя (табл. 4), а также изменение формы распределения деревьев по диаметру – сглаживания пиков и приближение формы кривой к экспоненциальной (рис. 4). Вместе с тем доля ели в составе древостоев массива остается достаточно стабильной (рис. 3). Устойчивость положения ели подтверждается фактом значительного увеличения числа новых деревьев, пополняющих перечетную часть древостоя после учета 1996 г. (рис. 4) за счет интенсивного процесса естественного возобновления после периода ветровалов в 1980-х – начале 1990-х годов (Федорчук и др., 2012).
Таблица 3.
Средний диаметр (см) деревьев в массиве “Вепсский лес” в разные учетные периоды
| Порода | Категория | Год учета | |||||
|---|---|---|---|---|---|---|---|
| 1991–1992 | 1996–1997 | 2001–2002 | 2007–2008 | 2013 | 2018–2019 | ||
| средний диаметр, см | |||||||
| Ель | Живые | 19.4 | 19.1 | 17.0 | 17.2 | 16.9 | 15.6 |
| Отпад | – | 19.1 | 19.4 | 20.0 | 20.1 | 21.6 | |
| Береза | Живые | 25.0 | 25.5 | 25.1 | 24.3 | 23.6 | 22.0 |
| Отпад | – | 20.9 | 26.4 | 23.9 | 25.1 | 26.1 | |
| Осина | Живые | 43.5 | 44.4 | 42.7 | 35.2 | 36.7 | 39.1 |
| Отпад | – | 46.7 | 51.6 | 58.0 | 61.3 | – | |
| Сосна | Живые | 39.0 | 38.4 | 39.9 | 40.3 | 41.6 | 42.3 |
| Отпад | – | 38.7 | 40.0 | – | 34.0 | 32.0 | |
Таблица 4.
Распределение объема древесного отпада (м3 га–1) по породам в массиве “Вепсский лес” в разные учетные периоды
| Порода | Год учета | ||||
|---|---|---|---|---|---|
| 1991–1997 | 1997–2002 | 2002–2008 | 2008–2013 | 2013–2019 | |
| древесный отпад, м3 га–1 | |||||
| Ель | 13 | 18 | 20 | 31 | 44 |
| Береза | 3 | 4 | 3 | 4 | 3 |
| Осина | 1 | 4 | 5 | 2 | 0 |
| Сосна | 1 | 0 | 0 | 0 | 0 |
| Итого | 18 | 26 | 28 | 37 | 48 |
Рис. 4.
Динамика (а) количества деревьев ели (Picea abies (L.) H. Karst) по ступеням толщины (%) и (б) естественного возобновления по группам высот в резервате “Вепсский лес” с 1991 по 2019 гг. N – число особей.
Отмечено влияние сильных ветров в конце 20‑го века на динамику других малонарушенных лесных массивов. В этот период средний запас ельников и всех древостоев массивов Центрально-Лесного и Висимского заповедников и части национального парка “Водлозерский” уменьшался, а площадь еловых лесов первых двух классов возраста увеличивалась (Федорчук и др., 2012).
Наблюдается снижение численности мелкого подроста и увеличение численности крупного подроста ели (рис. 4; Березин и др., 2020). Таким образом, динамика естественного возобновления ели обеспечивает непрерывное пополнение материнского древостоя новыми деревьями.
Вышеперечисленные особенности характерны для лесных сообществ, находящихся на заключительных стадиях фазы дигрессии (Федорчук и др., 2011). По всей видимости, фаза дигрессии запаса в настоящее время выражена не только на уровне отдельных БГЦ, но и на уровне лесного массива в целом. Тем не менее, наличие лесных сообществ, находящихся в демутационных фазах, позволяет судить о разнонаправленности динамических процессов на уровне массива (Корепин и др., 2020). Дальнейшее снижение запаса древостоев массива, вероятно, продолжится в ближайшие годы; на это указывает достаточно высокая доля древостоев с возрастом основного поколения старше 160–180 лет (рис. 2). Тенденция к увеличению количества пробных площадей с древостоями, постепенно переходящими в стадию нарастания запаса (табл. 2, рис. 1), может сохраниться за счет активизации процессов возобновления, формирования новых поколений на месте последних нарушений. Далее, по мере увеличения доли древостоев демутационных фаз в структуре массива, наблюдаемое в последние годы снижение среднего по массиву запаса, возможно, замедлится. Со временем переход этих древостоев в фазу активного роста, предположительно, выразится в увеличении среднего по массиву запаса.
Ценотическое разнообразие массива определяет характер его долговременной динамики (рис. 3). Так, динамика запаса древостоев и интенсивность отпада деревьев наиболее выражены в ельниках черничного и долгомошно-черничного типов. Наиболее стабильными оказываются низко продуктивные заболоченные чернично-сфагновые и майниково-сфагновые ельники и продуктивные БГЦ со смешанным составом древостоя (рис. 3б, 3в).
Структурное разнообразие ценозов является одним из механизмов поддержания их устойчивости и адаптации к существующему режиму естественных нарушений. Наиболее стабильными по динамике древесного запаса оказались относительно разновозрастные и смешанные по составу лесные сообщества. Возможно, это временное явление, связанное с процессом смены древесных пород в лесных сообществах. Достаточно устойчивой формой организации лесных сообществ также являлись относительно чистые абсолютно разновозрастные ельники, формирующиеся в данных ландшафтных условиях преимущественно в недостаточно дренированных местообитаниях. Часть таких биогеоценозов в настоящее время находится в дигрессивных фазах, но они менее выражены, по сравнению с более простыми по организации сообществами на дренированных местообитаниях. По величине древесного отпада особенно выделяются просто организованные БГЦ условно одновозрастных ельников (рис. 3г, 3д).
Таким образом, устойчивость лесного массива может определяться как относительной стабильностью всех составляющих его компонентов, так и уравновешенной разнонаправленностью динамических процессов, происходящих внутри отдельных БГЦ, составляющих массив (Дыренков, 1984; Федорчук и др., 2014; Корепин и др., 2020). Встречающиеся в лесных экосистемах различные по интенсивности и частоте естественные нарушения поддерживают их структурное разнообразие (Synek и др., 2020.; Shorohova et al., 2022).
ЗАКЛЮЧЕНИЕ
Полученные данные позволяют утверждать, что ветровальные нарушения в различные исторические периоды затрагивали большую часть древостоев исследуемого лесного массива. Ценотическое разнообразие массива, выраженное как площадное соотношение структурных единиц – древостоев различного динамического состояния (вариантов возрастной структуры и фаз возрастной циклической динамики), сопряженных с различными лесорастительными условиями, – определяет интенсивность ветровалов, характер и скорость восстановления сообществ после них.
Анализ динамики различных по лесорастительным условиям и структурной организации лесных сообществ показывает наибольшую выраженность динамических процессов в просто организованных условно одновозрастных еловых древостоях черничного типа леса, где последние 30 лет наблюдались значительное снижение запаса древостоя и рост древесного отпада. Наиболее стабильными по динамике древесного запаса оказались относительно разновозрастные смешанные древостои и чистые абсолютно разновозрастные ельники.
Динамические процессы на уровне лесного массива в целом зависят от степени участия в его структуре древостоев сходных форм динамики. Преобладание в лесном массиве древостоев старших возрастов позволяет предположить, что процессы дигрессии, выраженные в настоящее время не только на уровне отдельных биогеоценозов, но и на уровне лесного массива, будут продолжаться еще некоторое время. Однако успешность естественного возобновления в сочетании с увеличением количества пробных площадей, где древостои находятся в демутационных фазах динамики, свидетельствует об устойчивом восстановительном процессе.
Результаты работы могут быть использованы для разработки систем ведения лесного хозяйства в Балтийско-Белозерском и сходных по ландшафтным характеристикам таежных районах. Сохранение структурно-функционального разнообразия БГЦ на уровне массива и сохранение элементов БГЦ, специфичных для коренных лесов, в управляемых лесах позволит минимизировать различия между вовлеченными и не вовлеченными в хозяйственную деятельность лесами и сохранить их экосистемные функции (Burton et al., 1999; Bauhus et al., 2009; Puettmann et al., 2009; Kuuluvainen et al., 2011, 2012; Halme et al., 2013).
***
Авторы выражают глубокую признательность одному из родоначальников исследований на территории “Вепсского леса” Виктору Николаевичу Федорчуку за идейное руководство и вдохновение, а также всем коллегам, принимавшим участие в полевых и организационных работах, в особенности А.А. Шварц, М.Л. Кузнецовой, Г.В. Филиппову, А.А. Гладышеву, И.П. Коготько, Д.М. Докучаеву, А.М. Иванову, Л.А. Пегову.
Список литературы
Бекмансуров М.В., Бобровский М.В., Браславская Т.Ю., Евстигнеев О.И., Заугольнова Л.Б., Коротков В.Н., Калуцкова Н.Н., Леонова Н.А., Лихацкий Ю.П., Макарова В.А., Морозова А.С. Восточноевропейские леса: история в голоцене и современность. М.: Наука, 2004. 575 с.
Березин Г.В., Шорохова Е.В., Шорохов А.А., Капица Е.А., Корепин А.А. Естественное возобновление ели европейской в коренных лесах Балтийско-Белозерского таежного района. СПб.: Политех-пресс, 2020. 36–38 с.
Георгиевский А.Б. Фаза окон в коренных еловых лесах южной тайги // Ботанический журнал. 1992. Т. 77. № 6. С. 52–62.
Громцев А.Н. Динамика коренных таежных лесов в Европейской части России при естественных нарушениях // Актуальные проблемы геоботаники. III Всероссийская школа-конференция. Лекции. Петрозаводск: КарНЦ РАН, 2007. С. 283–301.
Дыренков С.А. Структура и динамика таежных ельников. Л.: “Наука”, 1984. 174 с.
Корепин А.А., Шорохов А.А., Шорохова А.А. Анализ многолетней динамики массива коренных лесов эталонного значения Балтийско-Белозерского таежного района. СПб.: Политех-пресс, 2020. С. 147–150 с.
Коротков В.Н. Новая парадигма в лесной экологии // Биологические науки. № 8. 1991. Вып. 8. С. 7–20.
Крышень А.М., Синькевич С.М., Шорохова Е.В. Variable retention forestry — лесоводство, ориентированное на непрерывное в пространстве и во времени сохранение лесной среды // Растительные ресурсы. 2020. Т. 56. № 3. С. 1–7.
Лукина Н.В., Гераськина А.П., Горнов А.В., Шевченко Н.Е., Куприн А.В., Чернов Т.И., Чумаченко С.И., Шанин В.Н., Кузнецова А.И., Тебенькова Д.Н., Горнова М.В. Биоразнообразие и климаторегулирующие функции лесов: актуальные вопросы и перспективы исследований // Вопросы лесной науки. 2020. Т.3. № 4. 90 с.
Мазинг В.В. Структура, состав и динамика бореальных растительных сообществ. Ученые записки Тарт. ун-та. Тарту, 1988. № 812. С. 122–141.
Маслов А.А. Количественный анализ горизонтальной структуры лесных сообществ. М.: Наука, 1990. 160 с.
Морозов Г.Ф. Учение о лесе. М.-Л., 1930. 440 с.
Пугачевский А.В. Ценопопуляции ели: структура, динамика, факторы регуляции. Минск: Наука и техника, 1992. 204 с.
Пугачевский А.В. Оценка экологических рисков в системе информационного обеспечения лесного хозяйства // Ботаника (исследования): Сборник научных трудов. Минск: Ин-т эксперимент. бот. НАН Беларуси, 2013. № 42. С. 243–258.
Скворцова Е.Б., Уланова Н.Г., Басевич В.Ф. Экологическая роль ветровалов. М.: Лесная промышленность, 1983. 192 с.
Смолоногов Е.П. Лесообразовательный процесс и ветровалы // Последствия катастрофического ветровала для лесных экосистем. Екатеринбург: УрО РАН, 2000. С. 12–18.
Сукачев В.Н., Дылис Н.В. Основы лесной биогеоценологии. М.: Наука, 1964. 566 с.
Сукцессионные процессы в заповедниках России и проблемы сохранения биологического разнообразия / Под ред. О.В. Смирновой, Е.С. Шапошникова. СПб.: РБО, 1999. 548 с.
Устойчивость лесов. Теория и практика биогеоценотических исследований. М.: Товарищество научных изданий КМК, 2018. 171 с.
Федорчук В.Н., Кузнецова М.Л., Андреева А.А., Моисеев Д.В. Резерват “Вепсский лес”. Лесоводственные исследования. СПб: СПбНИИЛХ, 1998. 208 с.
Федорчук В.Н., Шорохов А.А., Григорьева С.О. Охрана и изучение коренных еловых лесов Ленинградской области // Лесоведение. 2002. № 6. С. 23–28.
Федорчук В.Н., Шорохов А.А., Шорохова Е.В., Кузнецова М.Л. Динамика коренных еловых лесов европейской России // Лесоведение. 2014. № 2. С. 11–19.
Федорчук В.Н., Шорохов А.А., Шорохова Е.В., Кузнецова М.Л., Тетюхин С.В. Массивы коренных еловых лесов: структура, динамика, устойчивость // СПб.: Изд-во Политехнического ун-та, 2012. 140 с.
Федорчук В.Н., Шорохова Е.В., Шорохов А.А., Кузнецова М.Л. Возрастная динамика еловых древостоев северо-западной части Русской равнины // Лесоведение. 2011. № 3. С. 3–13.
Шорохова М.А., Березин Г.В., Капица Е.А., Шорохова Е.В. Характеристики крупных древесных остатков в лесном массиве “Вепсский лес” – эталоне природы средней тайги // Известия Санкт-Петербургской лесотехнической академии, 2021. № 236. С. 198–211.
Юрцев Б.А. Эколого-географическая структура биологического разнообразия и стратегия его учета и охраны // Биологическое разнообразие: подходы к изучению и сохранению. СПб., 1992. С. 7–21.
Attiwill P.M. The disturbance of forest ecosystems: the ecological basis for conservative management // Forest Ecology and Management. 1994. V. 63. № 2–3. P. 247–300.
Bauhus J., Puettmann K., Messier C. Silviculture for old-growth attributes // Forest Ecology and Management. 2009. V. 258. № 4. P. 525–537.
Burton P.J., Kneeshaw D.D., Coates K.D. Managing forest harvesting to maintain old growth in boreal and sub-boreal forests // For Chron. 1999. V. 75. № 4. P. 623–631.
Chambers J.Q., Negron-Juarez R.I., Marra D.M., Di Vittorio A., Tews J., Roberts D., Ribeiro G.H., Trumbore S.E., Higuchi N. The steady-state mosaic of disturbance and succession across an old-growth Central Amazon forest landscape // Proceedings of the National Academy of Sciences. 2013. V. 110. № 10. P. 3949–3954.
De Grandpré L., Waldron K., Bouchard M., Gauthier S., Beaudet M., Ruel J.C., Hébert C., Kneeshaw D.D. Incorporating insect and wind disturbances in a natural disturbance-based management framework for the boreal forest // Forests. 2018. V. 9. № 8. P. 471.
Everham E.M., Brokaw N.V.L. Forest damage and recovery from catastrophic wind // The Botanical Review. 1996. V. 62. № 2. P. 113–185.
Frelich L.E., Reich P.B. Perspectives on development of definitions and values related to old-growth forests // Environmental Reviews. 2003. V. 11. № 1. P. S9–S22.
Gauthier S., Leduc A., Bergeron Y., Kuuluvainen T., Vaillancourt M.A., Montoro Girona M., Macdonald E., Bélilse A.C., Shorohova E., Shvidenko A. The boreal forests in the face of global change: Is ecosystem management still part of the solution? In: Sustainable forest management of the boreal forests in the face of climate change. Springer, 2021 (in press).
Gauthier S., Vaillancourt M.A., Kneeshaw D.D., Drapeau P., Grandpré L De, Claveau Y., Paré D. Forest ecosystem management: origins and foundations // Ecosystem management in the boreal forest. Presses de l’Université du Québec. Quebec. 2009. P. 13–18.
Girard F., De Grandpré L., Ruel J.C. Partial windthrow as a driving process of forest dynamics in old-growth boreal forests // Canadian Journal of Forest Research. 2014. V. 44. № 10. P. 1165–1176.
Halme P., Allen K.A., Auniņš A., Bradshaw R.H.W., Brumelis G., Čada V., Clear J.L., Eriksson A.M., Hannon G., Hyvärinen E., et al. Challenges of ecological restoration: Lessons from forests in northern Europe // Biological Conservation. 2013. V. 167. P. 248–256.
Koivula M., Kuuluvainen T., Hallman E., Kouki J., Siitonen J., Valkonen S. Forest management inspired by natural disturbance dynamics (DISTDYN) – a long-term research and development project in Finland // Scandinavian Journal of Forest Research. 2014. V. 29. P. 579–592.
Král K., McMahon S.M., Janík D., Adam D., Vrška T. Patch mosaic of developmental stages in central European natural forests along vegetation gradient // Forest Ecology and Management. 2014. V. 330. P. 17–28.
Kuuluvainen T. Natural variability of forests as a reference for restoring and managing biological diversity in boreal Fennoscandia // Silva Fennica. 2002. V. 36. № 1. P. 97–125.
Kuuluvainen T., Aakala T. Natural forest dynamics in boreal Fennoscandia: a review and classification // Silva Fennica. 2011. V. 45. № 5. P. 823–841.
Kuuluvainen T., Tahvonen O., Aakala T. Even-Aged and Uneven-Aged Forest Management in Boreal Fennoskandia: A Review // Ambio. 2012. V. 41. № 7. P. 720–737.
Leibundgut H. Europäische Urwälder der Bergstufe. Bern, Stuttgart: Verlag Paul Haupt, 1982. 308 p.
Manabe T., Shimatani K., Kawarasaki S., Aikawa S.I., Yamamoto, S.I. The patch mosaic of an old-growth warm-temperate forest: patch-level descriptions of 40-year gap-forming processes and community structures // Ecological research. 2009. V. 24 № 3. P. 575–586.
Martin M., Boucher Y., Fenton N.J., Marchand P., Morin H. Forest management has reduced the structural diversity of residual boreal old-growth forest landscapes in Eastern Canada // Forest Ecology and Management. 2020. V. 458. P. 1–10.
Martin M., Fenton N.J., Morin H. Structural diversity and dynamics of boreal old-growth forests case study in Eastern Canada // Forest Ecology and Management. 2018. V. 422. P. 125–136.
Martin M., Girona M.M., Morin H. Driving factors of conifer regeneration dynamics in eastern Canadian boreal old-growth forests // PLoS ONE. 2020. V. 15. № 7. P. e0230221.
Martin M., Shorohova E., Fenton N.J. Boreal old-growth forests are ecosystems of exceptional ecological and social value. In: Sustainable forest management of the boreal forests in the face of climate change. Springer. 2021 (in press).
McCarthy J. Gap dynamics of forest trees: a review with particular attention to boreal forests // Environmental Reviews. 2001. V. 9. № 1. P. 1–59.
Meigs G.W., Morrissey R.C., Bače R., Chaskovskyy O., Čada V., Després T., Donato D.C., Janda P., Lábusová J., Seedre M. et al. More ways than one: Mixed-severity disturbance regimes foster structural complexity via multiple developmental pathways // Forest Ecology and Management. 2017. V. 406. P. 410–426.
Messier C., Puettmann K., Coates D.J. Managing forests as complex adaptive systems: building resilience to the challenge of global change // London, 2013. 368 p.
Pickett S.T.A., White P. The ecology of natural disturbance and patch dynamics // Orlando: Academic Press, 1985. 632 p.
Potapov P., Hansen M.C., Laestadius L., Turubanova S., Yaroshenko A., Thies C., Smith W., Zhuravleva I., Komarova A., Minnemeyer S. et al. The last frontiers of wilderness: Tracking loss of intact forest landscapes from 2000 to 2013 // Science Advances. 2017. V. 3. № 1. P. 1–14.
Puettmann K.J., Coates K.D., Messier C. A Critique of Silviculture: Managing for Complexity. Washington D.C.: Island Press, 2009. 190p.
Ruel J.C. Factors influencing windthrow in balsam fir forests: from landscape studies to individual tree studies // Forest Ecology and Management. 2000. V. 135. P. 169–178.
Saad C., Boulanger Y., Beaudet M., Gachon P., Ruel J.C., Gauthier S. Potential impact of climate change on the risk of windthrow in eastern Canada’s forests // Climatic Change. 2017. V. 143. P. 487–501.
Shorohova E, Kuuluvainen T, Kangur A, Jõgiste K. Natural stand structures, disturbance regimes and successional dynamics in the Eurasian boreal forests: a review with special reference to Russian studies // Annals of Forest Science. 2009. V. 66. № 2. P. 1–20.
Shorohova E., Aakala T., Gauthier S., Kneeshaw D., Koivula M., Ruel J.C., Ulanova N. Natural Disturbances from the Perspective of Forest Ecosystem-based Management. In: Sustainable forest management of the boreal forests in the face of climate change. Springer. 2022 (in press).
Shorohova E., Kneeshaw D., Kuuluvainen T., Gauthier S. Variability and dynamics of old-growth forests in the circumboreal zone: implications for conservation, restoration and management // Silva Fennica. 2011. V. 45. № 5. P. 785–806.
Shorohova E., Fedorchuk V.N., Kuznetsova M.L., Shvedova O. Wind-induced successional changes in pristine boreal Picea abies forest stands: evidence from long-term permanent plot records // Forestry. 2008. V. 81. P. 335–359.
Spies T.A., Turner M.A. Dynamic forest mosaics // In the book “Maintaining biodiversity in forest ecosystem”. Ed by L. Malcolm & J. Hunter: Cambridge Univ. Press., 1999. P. 95–160.
Synek M., Janda P., Mikolas M., Nagel T.A., Schurman J.S., Pettit J.L., Trotsiuk V., Morrissey R.C., Bace R., Cada V., Brang P., Bugmann H., Begovic K., Chaskovskyy O., Dusatko M., Frankovic M., Kameniar O., Knir T., Kozak D., Langbehn T., Malek J., Rodrigo R., Saulnier M., Teodosiu M., Vostarek O., Svoboda M. Contrasting patterns of natural mortality in primary Picea forests of the Carpathian Mountains // Forest Ecology and Management. 2020. V. 457. P. 117 734.
Trotsiuk V., Svoboda M., Janda P., Mikolas M., Bace R., Rejzek J., Samonil P., Chaskovskyy O., Korol M., Myklush S. A mixed severity disturbance regime in the primary Picea abies (L.) Karst. forests of the Ukrainian Carpathians // Forest Ecology and Management. 2014. V. 334. P. 144–153.
Ulanova N.G. The effects of windthrow on forests at different spatial scales: a review // Forest Ecology and Management. 2000. V. 135. P. 155–168.
Venier L.A., Walton R., Thompson I.D., Arsenault A., Titus B.D. A review of the intact forest landscape concept in the Canadian boreal forest: Its history, value, and measurement // Environmental Review. 2018. V. 26. № 4. P. 369–377.
Waldron K., Ruel J.C., Gauthier S. The effects of site characteristics on the landscape-level windthrow regime in the North Shore region of Quebec, Canada // Forestry. 2013. V. 86. № 2. P. 159–171.
Watson J.E.M., Evans T., Venter O., Williams B., Tulloch A., Stewart C., Thompson I., Ray J.C., Murray K., Salazar A., et al. The exceptional value of intact forest ecosystems // Nature Ecology & Evolution. 2018. V. 2. № 4. P. 599–610.
Weldon J., Grandin U. Major disturbances test resilience at a long-term boreal forest monitoring site // Ecology and Evolution. 2019. V. 9. № 7. P. 4275–4288.
Дополнительные материалы отсутствуют.