

Микробиология, 2019, T. 88, № 6, стр. 631-644
Термокарстовые озера – экосистемы с интенсивными микробными процессами цикла метана
А. Ю. Каллистова a, *, А. С. Саввичев a, И. И. Русанов a, Н. В. Пименов a
a Институт микробиологии им. С.Н. Виноградского, ФИЦ Биотехнологии РАН
119071 Москва, Россия
* E-mail: kallistoanna@mail.ru
Поступила в редакцию 20.05.2019
После доработки 02.07.2019
Принята к публикации 10.07.2019
Аннотация
Термокарстовые озера образуются в результате вытаивания высокольдистых многолетнемерзлых пород, приводящего к появлению просадочных форм рельефа, которые при равнинном характере территории и наличии положительного водного баланса заполняются водой. Активация процесса термокарста является одним из возможных проявлений деградации вечной мерзлоты в условиях глобального потепления. Термокарстовые озера распространены в зонах сплошного, прерывистого и островного залегания вечной мерзлоты, т.е. на территориях Сибири, Аляски, Канады и северной Скандинавии. В этих озерах развиваются комплексные микробные сообщества, адаптированные к длительному периоду воздействия низких температур и различающиеся интенсивностью аэробного и анаэробного метаболизма в зависимости от минерального состава донных отложений, количества доступного органического вещества, лимнологических и гидрохимических особенностей водоемов. Для многих термокарстовых озер характерны высокие скорости эмиссии метана. Исследования последних лет показали, что в термокарстовых озерах происходит интенсивное образование метана по разным путям метаногенеза, а также аэробное и анаэробное метанокисление с участием разнообразных метаногенных и метанотрофных бактерий и архей. При этом остается открытым вопрос о механизмах и микроорганизмах, вовлеченных в процесс анаэробного окисления метана, за счет которого в термокарстовых озерах может потребляться до 80% метана. Кроме того, недостаточно изучены микроорганизмы, активно функционирующие подо льдом в течение длительного зимнего периода, крайне важные для водных экосистем северных регионов. Приходится также признать практически полное отсутствие серьезных микробиологических исследований термокарстовых озер России, несмотря на то, что в нашей стране вечная мерзлота занимает до 65% территории, процесс термокарста распространен и термокарстовые озера многочисленны.
ВВЕДЕНИЕ
Вечная мерзлота (многолетнемерзлые породы, permafrost) − это “такой находящийся на некоторой глубине от дневной поверхности слой почвы, который имеет отрицательную температуру, длящуюся непрерывно минимум два года, максимум тысячелетия и десятки тысячелетий” (цит. стр. 20, Сумгин, 1927). Вечная мерзлота Северного полушария занимает ~15% земной поверхности (Saunois et al., 2016 и ссылки там же) и является одним из крупнейших наземных пулов органического углерода (Ciais et al., 2013 и ссылки там же). Многолетнемерзлые породы подстилают 65% территории России, поэтому неудивительно, что в течение последних 100 лет криолитозона активно изучается российскими учеными различных специальностей (Сумгин, 1927; Ефимов, 1950; Общее мерзлотоведение, 1978; Gilichinsky et al., 1995; Леин и соавт., 2003, 2005; Rivkina et al., 2004; Shcherbakova et al., 2004, 2016; Leibman et al., 2005; Ривкина и соавт., 2006; Drozdov et al., 2015; Romanovsky et al., 2017 и др.). За рубежом интерес к исследованию вечной мерзлоты возник сравнительно недавно – в начале 1980-х гг. (Jorgenson et al., 2001 и ссылки там же), и растет год от года в связи с многочисленными драматичными предсказаниями повсеместного ее таяния, которое ожидается к концу XXI века (Lawrence, Slater, 2005; Romanovsky et al., 2017; Hjort et al., 2018). Согласно предполагаемому сценарию, глобальное потепление климата будет способствовать оттаиванию многолетнемерзлых пород, высвобождая захороненный в них углерод, результатом чего станет резкое увеличение эмиссии парниковых газов (СН4, СО2) из высокоширотных экосистем (Zimov et al., 2006), а также повреждение инфраструктуры арктических регионов и серьезный ущерб окружающей среде (Romanovsky et al., 2017; Hjort et al., 2018). В настоящее время в год публикуется в среднем 1000 статей, так или иначе связанных с проблемой таяния вечной мерзлоты (Oliva, Fritz, 2018).
Нет сомнений в том, что с потеплением климата сократится протяженность многолетнемерзлых отложений, результатом чего станет оттаивание некоторой части захороненного в них органического углерода. При этом величина возможных потерь оттаявшего углерода в виде эмиссии в атмосферу CO2 и CH4 достоверно не известна. Последствия высвобождения углерода из вечной мерзлоты сложно оценить, поскольку не известны масштабы и скорости процесса оттаивания, доля углерода, которая при этом высвободится, возможный смягчающий эффект различных факторов, а также роль локальных процессов (пространственной изменчивости деградации вечной мерзлоты). Пока нет четких доказательств того, что оттаивание вечной мерзлоты вносит значимый вклад в современный глобальный бюджет CO2 и CH4. Моделирование поведения многолетнемерзлых пород и эмиссии из них парниковых газов указывает на относительно медленную положительную динамику, исчисляемую сотнями лет. При этом потенциальное количество углерода, которое может быть высвобождено в качестве CO2 и CH4 из вечной мерзлоты, близко к таковому для других экосистем, например, к потенциальному количеству CO2, которое может быть выделено наземными почвами в ответ на глобальное потепление. Специалисты, тем не менее, утверждают, что с потеплением Арктики эмиссия парниковых газов увеличится, что приведет к положительной обратной реакции со стороны климата (Ciais et al., 2013 и ссылки там же).
В результате оттаивания вечной мерзлоты в пресноводные системы поступают захороненные в ней биогенные элементы в составе органических и минеральных веществ, где они концентрируются и становятся доступными для ремобилизации (Vonk et al., 2015). Арктические пресноводные системы занимают около 16% зоны залегания многолетнемерзлых пород (Vonk et al., 2015) и включают в себя, в том числе, озера различного генезиса – тектонические, ледниковые, пойменные и термокарстовые (Кравцова, 2009). Термокарстовые озера составляют значимую часть арктической пресноводной системы и обусловливают специфику тундровых и лесотундровых ландшафтов (Кравцова, 2009). Они широко распространены в Сибири, на Аляске, в Канаде и северной Скандинавии (Кравцова, 2009, Grosse et al., 2013; Vonk et al., 2015, Wik et al., 2016).
Термокарстовые озера образуются в результате процесса вытаивания высокольдистых многолетнемерзлых пород, приводящего к появлению просадочных форм рельефа, которые при равнинном характере территории и наличии положительного водного баланса заполняются водой, т.е. процесса термокарста (термин введен д.г.-м.н., проф. М.М. Ермолаевым в 1932 г.) (Общее мерзлотоведение, 1978). Активация термокарстовых процессов является одним из наиболее вероятных проявлений деградации вечной мерзлоты (Кравцова, 2009). Кроме этого, многие термокарстовые озера характеризуются высокими скоростями эмиссии метана, и в них предполагается усиление процесса метаногенеза при дальнейшем оттаивании вечной мерзлоты (Walter et al., 2007; Laurion et al., 2010; Покровский и соавт., 2012; Heslop et al., 2015; Martinez-Cruz et al., 2015; Vonk et al., 2015; Wik et al., 2016; Townsend-Small et al., 2017; Serikova et al., 2019). И, напротив, отдельные исследования указывают на то, что некоторые термокарстовые экосистемы, бывшие в начале своего образования источником углерода, превратились за последние тысячелетия в его сток (Walter Anthony et al., 2014; Bouchard et al., 2017).
В обзоре кратко изложены довольно хорошо изученные к настоящему времени принципы образования термокарстовых озер, данные об их распространении на территории России и лимнологические характеристики озер. Основное внимание уделено обобщению современных данных о микробных процессах и микроорганизмах цикла метана в термокарстовых озерах.
ОБРАЗОВАНИЕ ТЕРМОКАРСТОВЫХ ОЗЕР
Настоящий раздел составлен с использованием материалов диссертации на соискание степени к.г.н. Т.В. Родионовой (2013). Развитие процесса термокарста происходит при совокупном соблюдении нескольких обязательных условий: наличии высокольдистых многолетнемерзлых пород; увеличении глубины сезонного или многолетнего протаивания, при котором глубина протаивания должна превышать глубину залегания подземного льда или сильнольдистой породы; равнинном характере территории, затрудняющем сток поверхностных вод; наличии положительного водного баланса (Общее мерзлотоведение, 1978; Романовский, 1993).
Вода, образовавшаяся при оттаивании льда, отфильтровывается вверх, а талая кровля проседает, заполняя образующуюся полость и создавая на поверхности понижение. Образование термокарстовых озер может инициироваться скоплением атмосферных осадков в естественных или антропогенно-обусловленных западинах на участках развития льдистых отложений. При дренаже процессы термокарста не приводят к формированию озер, поскольку в массиве пород на месте вытаявших тел остаются пустоты. Это способствует развитию термоэрозии, образованию долинообразных понижений (аласных долин) и систем стока из соединившихся термокарстовых котловин (Общее мерзлотоведение, 1978; Романовский, 1993; Grosse et al., 2013).
Образование термокарстовых озер в зонах сплошного распространения вечной мерзлоты проходит через несколько последовательных стадий (рис. 1): понижение поверхности почвы в начальной фазе таяния льда; заполнение просадок водой и образование озерка; расширение и углубление озерной просадочной котловины при полном оттаивании льда на данном участке; образование довольно полноводного озера; постепенное высыхание или дренирование термокарстового озера (Ефимов, 1950). В связи с более суровыми климатическими условиями на приморских низменностях севера Якутии, в северных областях Западно-Сибирской низменности и на арктических островах возможно устойчивое существование мелких (глубиной до 1–1.2 м) термокарстовых озер, донные отложения которых сезонно оттаивают, но под сезонноталым слоем залегают высокольдистые породы. Таким образом, под термокарстовыми озерами может устойчиво существовать как сезонноталый слой с температурой поверхности пород дна водоема ниже 0°С, так и талики с температурой поверхности пород дна водоема выше 0°С. Талики и таликовые (талые) зоны – это толщи талых пород, развитые с поверхности земли или под водоемами и водотоками и непрерывно существующие более одного года (Романовский, 1983). Талики, ограниченные мерзлыми породами только по боковым поверхностям, являются сквозными. По ним осуществляется взаимосвязь поверхностных вод и вод глубоких подмерзлотных горизонтов. Талики, подстилаемые на некоторой глубине мерзлыми породами, называются несквозными или надмерзлотными. По несквозным водоносным таликам происходит приповерхностный сток (Бойцов, 2011). Мощность подозерного талика и его размеры в плане зависят от глубины и площади озера, времени его существования, распределения температур донных отложений по площади. Под озерами, размер которых по короткой оси превышает удвоенную мощность многолетнемерзлой толщи, образуются сквозные талики (Общее мерзлотоведение, 1978; Бойцов, 2011).
Рис. 1.
Образование и развитие термокарстового озера в зоне сплошного распространения вечной мерзлоты (упрощенная схема развития аласного рельефа по Соловьеву (1962)): 1 – рельеф в зоне сплошного распространения вечной мерзлоты; 2 – понижение поверхности почвы в начальной фазе таяния льда и заполнение просадок водой (зачаточный былар); 3 – расширение и углубление озерной просадочной котловины при полном оттаивании льда на данном участке, образование полноводного озера (тымпа, впоследствии элементарный алас); 4 – постепенное высыхание или дренирование термокарстового озера, повторное льдообразование (зрелый алас).
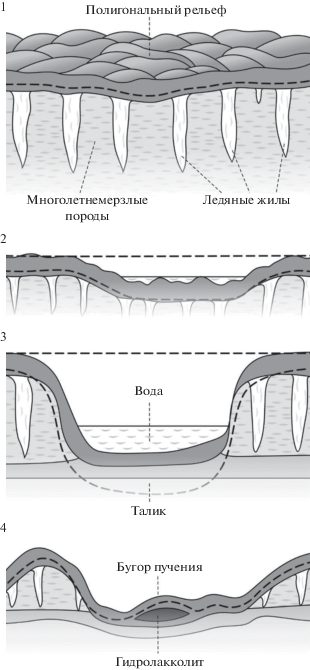
Как правило, термокарст прекращается при достижении равенства между глубиной сезонного оттаивания и мощностью слоя талых отложений, а чем меньше льдистость пород, тем быстрее растет доля талых отложений (Общее мерзлотоведение, 1978). Если вода удаляется из термокарстового понижения и прекращается ее “отепляющее воздействие”, происходит промерзание протаявших пород, сопровождающееся повторным льдообразованием и возникновением ледяных ядер – гидролакколитов, часто называемых буграми пучения (синонимы: пинго, булгуннях, туфур, тебелер). Впоследствии возможно повторное протаивание новообразовавшихся ледяных ядер, приводящее к формированию крупных округлых и овальных озер или заполненных водой впадин (Общее мерзлотоведение, 1974; Попов и соавт., 1985). Следует отметить, что бугры пучения могут и не иметь в основании льды и льдистые породы. В их основании может скапливаться, например, метан, выброс которого в атмосферу приводит к формированию глубоких кратеров – воронок газового выброса (ВГВ), которые постепенно заполняются водой и превращаются в озера. Генезис озер, образовавшихся на месте ВГВ, не связан с процессом термокарста (Leibman et al., 2014; Savvichev et al., 2018).
В зонах прерывистого распространения вечной мерзлоты процесс термокарста представляет собой различные стадии эволюции плоскобугристых болот (пальсы) от малых термокарстовых просадок и депрессий до зрелых озер и остатков – хасыреев (см. определение ниже). Пальса (palsa) образуется в заболоченных регионах на сезонноталом слое мерзлоты и может рассматриваться как разновидность бугра пучения, ядром которого является высокольдистый замороженный торф и минеральные отложения. В многолетнемерзлых минеральных почвах образуются аналогичные возвышенности, называемые литальсами (lithalsas) (Payette et al., 2004; Calmels et al., 2008; Покровский и соавт., 2012; Bouchard et al., 2017).
Широкое развитие термокарста и массовое образование термокарстовых озер произошло при оттаивании плейстоценовых отложений в эпоху голоценового климатического оптимума, когда температура воздуха повысилась, по сравнению с современной, на 2–2.5°С, и наблюдалось увеличение количества осадков (Качурин, 1961; Романовский, 1993; Родионова, 2013).
РАСПРОСТРАНЕНИЕ И ДИНАМИКА ПЛОЩАДЕЙ ТЕРМОКАРСТОВЫХ ОЗЕР НА ТЕРРИТОРИИ РОССИИ
Первая карта распространения термокарстовых озер на территории России, составленная на основании анализа космических снимков с интернет-портала Google Earth, представлена в статье Кравцовой (2009). Охарактеризованы размеры озер и густота их сети во всех районах их распространения. Автор подчеркивает, что в районах развития вечной мерзлоты в России встречаются озера разного генезиса, но считает, что преобладают именно термокарстовые, составляющие абсолютное большинство (Кравцова, 2009). Геологически обоснованные доказательства такого предположения в статье не приводятся.
Выше уже упоминалось, что многолетнемерзлые породы занимают до 65% территории России. При этом на большую часть территории севера и северо-востока страны приходится зона сплошной мерзлоты мощностью от 100–500 до более 1000 м. Согласно исследованию Кравцовой (2009), сопоставление границ районов массового распространения термокарстовых озер с границами развития многолетнемерзлых пород показало, что термокарстовые озера характерны для зон сплошного, прерывистого и островного распространения вечной мерзлоты в России. Наибольшую площадь занимают районы распространения термокарстовых озер в зонах тундр и лесотундр субарктического пояса: восточная часть Кольского полуострова, север Восточно-Европейской равнины, южная часть Ямала, Гыданский и Тазовский полуострова, север Западно-Сибирской низменности, южная часть Таймыра, депрессия в районе оз. Ессей, южная часть Колымской низменности, Анадырская низменность. Вся дельта Лены представляет собой район развития термокарстовых озер. Равнины Центральной Якутии являются зоной развития аласов (Кравцова, 2009; Кравцова, Родионова, 2016). Аласы (хасыреи) – это котловины бывших тер-мокарстовых озер, опустевших из-за тер-моэрозионных процессов. Аласы часто характ-еризуются обширными площадями просевшей земной поверхности с остаточными озерами (подробнее см. в обзоре Crate et al., 2017). Районы распространения термокарстовых озер заходят также в зону северной тайги – хвойных лесов умеренного пояса на севере Восточно-Европейской равнины, значительной части Западно-Сибирской низменности, на Центрально-Якутской равнине, Камчатке, Тихоокеанском побережье Дальнего Востока. В межгорных впадинах Забайкалья термокарстовые озера распространены на участках южной тайги и островных южно-сибирских степей. В ряде районов России распространение термокарстовых озер выходит за пределы современных границ многолетнемерзлых пород (Кравцова, 2009; Кравцова, Родионова, 2016).
В последние годы публикуется довольно много русскоязычных статей, посвященных исследованию распространения, динамики и количества термокарстовых озер на территории России по данным космических снимков (например, Брыксина и соавт., 2007, 2011; Кравцова, 2009; Кравцова, Тарасенко, 2011; Кравцова, Родионова, 2016; Полищук и соавт., 2016, 2017 и др.). Метод дистанционного зондирования также широко используется для мониторинга изменений площадей термокарстовых озер на Аляске, в Канаде и Швеции (Romanovsky et al., 2017 и ссылки там же). Динамика изменения площадей и количества термокарстовых озер привлекает внимание ученых как возможный показатель состояния криолитозоны при потеплении климата. Однако, помимо повышения температуры воздуха (потепления климата), которое может активизировать процесс термокарста, на изменчивость термокарстовых озер влияет целый комплекс взаимосвязанных гидрологических, метеорологических, тектонических и антропогенных факторов. За 30-летний период мониторинга термо-карстовых озер на территории России выявлено преобладание сокращения площадей и количества озер над их увеличением. Массовое сокращение площадей озер связано с гидрологическими факторами, зарастанием водоемов и, в отдельных случаях, с антропогенным влиянием. Однако в отдельных регионах зафиксировано увеличение площадей озер, связанное как с активацией термокарстового процесса (на Яно-Индигирской низменности, побережье Северного Ледовитого океана, п-овах Ямал и Гыданский), так и с гидрологическими факторами (в речных долинах на территории Колымской низменности, дельтах рек Печора и Яна, котловинах Забайкалья). Цикличные изменения количества атмосферных осадков (например, на территории Центральной Якутии) приводят как к увеличению площади озер, так и к уменьшению в зависимости от наступления многоводного или маловодного периода. Антропогенная деятельность, особенно в районах активной нефтедобычи (например, на территории Среднеобской низменности на юге Западной Сибири) также приводит к увеличению площадей озер. В сложном комплексе факторов, влияющих на изменения размеров озер, выделить влияние современного потепления климата пока не представляется возможным (Кравцова, Родионова, 2016).
Исследования последних лет с использованием дистанционного зондирования документируют снижение уровня вод и усиление дренажа в озерах, расположенных в термокарстовых ландшафтах, с заметными различиями между зонами сплошного и прерывистого залегания вечной мерзлоты. Неясно, связаны ли такие серьезные гидрологические сдвиги исключительно с климатом или с термокарстовой активностью, или с их сочетанием. Обнаружение процессов, ответственных за изменения уровня воды в термокарстовых ландшафтах, важно для более точного прогнозирования экологических последствий, которые могут повлиять как на местную флору и фауну, так и на традиционный образ жизни населения северных регионов (Bouchard et al., 2017 и ссылки там же).
ХАРАКТЕРИСТИКА ТЕРМОКАРСТОВЫХ ОЗЕР
Термокарстовые озера подробно охарактеризованы в обзорах Вонк и соавт. (Vonk et al., 2015) и Бушара и соавт. (Bouchard et al., 2017). В исчерпывающем обзоре Вонк и соавт. (2015) обстоятельно изложены лимнологические и биогеохимические особенности термокарстовых озер. Работа Бушара с соавт. (2017) является обобщением последних палеолимнологических реконструкций, основанных на многопараметрическом анализе осадочных отложений водоемов, расположенных в зоне вечной мерзлоты, включая север Канады, Аляску и дельту Лены (северо-восток Сибири), которые позволяют понять временные закономерности образования озера, эрозии водосборного бассейна, продуктивности воды, гидрологической эволюции и связанных с этим ландшафтных изменений. Мы не будем подробно анализировать данные упомянутых работ, ограничившись лишь краткой характеристикой термокарстовых озер; для более детального ознакомления рекомендуем обратиться к первоисточникам.
Размеры и площади термокарстовых озер варьируют в широком диапазоне от нескольких метров (пруды, ponds) до километров. Обычно термокарстовые озера мелководны, однако встречаются и глубокие озера (≥10 м). Лимнологические и биогеохимические особенности озера зависят от того, сформировалось ли оно при таянии вечной мерзлоты едомного или неедомного типа (табл. 1) (Sepulveda-Jauregui et al., 2015; Vonk et al., 2015). Едома – это тип вечной мерзлоты, обогащенной органическим углеродом (2–5% по массе) с преобладанием позднеплейстоценового лeсса со льдом. Едома представляет собой положительную форму рельефа (возвышенность) и широко распространена в Восточной Сибири, Канаде и на Аляске (Zimov et al., 2006; Heslope et al., 2015; Vonk et al., 2015). Термокарстовые озера, образованные в мерзлоте едомного типа, обычно глубокие, гумусовые, подстилаемые таликом и зачастую стратифицированные. Слабо окисленные или восстановленные условия и наличие доступного органического вещества благоприятны для прохождения процесса метаногенеза в этих озерах (Walter et al., 2006; Sepulveda-Jauregui et al., 2015; Vonk et al., 2015).
Таблица 1.
Сравнительная физико-химическая характеристика термокарстовых озер Аляски, расположенных в зонах сплошного, прерывистого и островного залегания вечной мерзлоты едомного и неедомного типа (Sepulveda-Jauregui et al., 2015)
| Характеристика озера* | Тип вечной мерзлоты | |
|---|---|---|
| едома | неедома | |
| Трофический статус | Миксотрофные | Ультраолиготрофные и олиготрофные |
| Температура воды, зима/лето, °С | 1.1/11.3 | 1.6/14.9 |
| рН воды, зима/лето | 7.1/8.2 | 6.7/7.7 |
| Окислительно-восстановительный потенциал воды, зима/лето, мВ | 84/187 | 222/295 |
| Хлорофилл a, мкг л–1 | 34.5 | 14.5 |
| Ортофосфаты, мкг Р л–1 | 27.9 | 5.3 |
| Нитраты, мг ${\text{NO}}_{3}^{ - }$−N л–1 | 0.09 | 0.02 |
| Сульфаты, мг ${\text{SO}}_{4}^{{2 - }}$−S л–1 | 0.44 | 5.39 |
| Общий органический углерод, вода, мг л–1 | 26.6 | 10.1 |
| Общий органический углерод, поверхностные осадки, % | 7.6 | 10.0 |
| Общий азот, вода, мг л–1 | 2.0 | 0.6 |
| Общий азот, поверхностные осадки, % | 1.0 | 1.0 |
Термокарстовые озера, образованные в мерзлоте неедомного типа, обычно нестратифицированы и имеют насыщенные кислородом придонные воды. Однако в некоторых мелководных (<3 м глубиной) озерах и прудах наблюдается температурная стратификация в летний сезон и, как следствие, метаногенез. При этом выделяется гораздо меньше метана, чем в озерах, образованных в мерзлоте едомного типа. Зимой глубокие термокарстовые озера имеют выраженную перевернутую стратификацию подо льдом, а мелководные озера замерзают полностью. Многие термокарстовые озера, например, тундровые, являются полимиктическими. Слабая стратификация или ее отсутствие летом в таких озерах может быть связана с незащищенностью озерных вод от перемешивания ветром в отсутствие густой растительности, со сниженной способностью холодных вод к стратификации, повышенным конвективным перемешиванием, меньшим поверхностным поглощением света и нагреванием. Стратификация термокарстовых озер также зависит от наличия градиентов солености и концентрации газов (Vonk et al., 2015 и ссылки там же).
Когда вечная мерзлота оттаивает, баланс между выбросом органического вещества в виде парниковых газов, включением углерода в микробные пищевые цепи, его захоронением в осадках или вымыванием зависит от того, какое органическое вещество – растворимое или взвешенное – поступает в водоем при оттаивании, а также от состава органического вещества, морфологических и стратификационных характеристик озера. Любые изменения в стратификации, окислительно-восстановительном режиме, доступе кислорода и др. влияют на этот баланс (Vonk et al., 2015). Исследования термокарстовых просадок, малых, больших озер и хасыреев в зоне островной и прерывистой мерзлоты (зона плоскобугристых болот – пальсы), выявили систематическое увеличение концентраций растворенного органического углерода, CO2, CH4, основных ионов, микроэлементов и тяжелых металлов с уменьшением размера водных объектов. Основным механизмом, ответственным за формирование состава воды озер от начала таяния мерзлоты в пальсе до образования больших термокарстовых озер, вероятно, является прогрессивная минерализация растворенного органического вещества торфа аэробными микроорганизмами. Это предполагает, что в начале процесса таяния пальсы концентрация растворенного органического вещества и, как следствие, парниковых газов в водных объектах будет увеличиваться. Более того, таяние мерзлоты в Западной Сибири может привести к замещению больших (зрелых) термокарстовых озер, которые дренируются в речную сеть, на меньшие по площади, но многочисленные просадки и озерки. Эти малые водные объекты площадью 1–100 м2 могут оказаться крайне важными и пока не учитываемыми источниками эмиссии парниковых газов в атмосферу и потока растворенного органического и других веществ в реки (Покровский и соавт., 2012; Shirokova et al., 2013).
Считается, что следствием образования термокарстовых озер является быстрое разложение древнего (≥11 500 лет до н.э.) оттаявшего углерода и его эмиссия в атмосферу в виде парниковых газов. Однако измерение эмиссии CH4 и CO2 из озер Аляски различного генезиса, включая термокарстовые, показало, что основным (~90% от общего объема эмиссии) источником парниковых газов являлся не плейстоценовый углерод вечной мерзлоты, а современное и “молодое” органическое вещество (<3300 лет до н.э.). При этом годовая эмиссия СО2 значительно превышала годовую эмиссию СН4 (Elder et al., 2018), что может быть связано как с преобладанием аэробного процесса разложения органического вещества и/или со слабым метаногенезом, так и с высокой интенсивностью метанокисления в этих озерах.
МИКРОБНЫЕ ПРОЦЕССЫ ЦИКЛА МЕТАНА В ТЕРМОКАРСТОВЫХ ОЗЕРАХ
В термокарстовых озерах развиваются сложные микробные сообщества, адаптированные к длительному периоду воздействия низких температур и различающиеся по интенсивностям аэробного и анаэробного метаболизма в зависимости от состава донных отложений, качественного состава и количества растворенного vs взвешенного органического вещества, минеральных веществ, ионов, лимнологических и гидрохимических особенностей водоема (Vonk et al., 2015). Термокарстовые озера являются одним из значимых источников метана, выделяя около 4.1 ± 2.2 Тг CH4 год–1, что составляет ~25% от ежегодной эмиссии метана из северных пресноводных экосистем (озер и прудов) (Wik et al., 2016). Неудивительно поэтому, что в современных работах по исследованию термокарстовых озер важное место занимает изучение микробных процессов цикла метана – метаногенеза и метанокисления, а также филогенетического разнообразия микроорганизмов, участвующих в этих процессах.
Метаногенез
Метаногенез является основной терминальной стадией разложения органического вещества в пресноводных стратифицированных водоемах с низким содержанием сульфата и, в частности, в термокарстовых озерах. Высокая потенциальная метаногенная активность (5.95 мкг C–CH4 г сух. вещества–1 сут–1; 125.9 мкг C–CH4 г ${\text{C}}_{{{\text{орг}}}}^{{--1}}$ сут–1) выявлена в плотных, обогащенных органическим веществом поверхностных осадках термокарстового озера Волт (Vault, Аляска), а также в молодом (недавно оттаявшем) слое отложений на дне подозерного талика (1.18 мкг C-CH4 г сух. вещества–1 сут–1; 59.6 мкг C-CH4 г ${\text{C}}_{{{\text{орг}}}}^{{--1}}$ сут–1). Более низкие скорости образования метана наблюдались в отложениях, которые оттаивали в талике в течение более длительного периода времени. Для сравнения, в почвенных образцах вечной мерзлоты образования метана не происходило. Потенциальные скорости метаногенеза в осадках озера Волт, которое образовалось при таянии мерзлоты едомного типа, на несколько порядков превышали скорости метаногенеза в осадках нетермокарстовых арктических озер (Heslop et al., 2015).
Известно четыре катаболических пути анаэробного образования метана метаногенными археями – гидрогенотрофный, ацетокластический, метилотрофный и метил-редуцирующий (подробнее о путях метаногенеза см. в обзоре Каллистовой и соавт., 2017). В одних термокарстовых озерах выявлен исключительно ацетокластический путь метаногенеза, в других – гидрогенотрофный, а в некоторых озерах оба пути вносят примерно одинаковый вклад в общий процесс образования метана (Negandhi et al. 2013; Vonk et al., 2015; Matheus Carnevali et al., 2015, 2018; Townsend-Small et al., 2017; de Jong et al., 2018). В термокарстовых озерах выявлен также метилотрофный путь метаногенеза (de Jong et al., 2018).
Влияние температуры и субстратов метаногенеза на скорость образования метана исследовали в термокарстовых озерах Аляски. При этом наблюдалась взаимосвязь между количеством образованного метана и количеством лабильного органического вещества в осадках (Matheus Carnevali et al., 2015). Добавление таких субстратов метаногенеза, как ацетат, Н2/СО2 и триметиламин приводило к увеличению потенциальной метаногенной активности осадков, по сравнению с контролем (без субстратов) при разных температурах (4 и 10°C). Максимальная скорость образования метана наблюдалась при более высокой температуре (10°С) и обогащении осадков триметиламином (метилотрофный путь); в сообществе при этом доминировали метаногены семейства Methanosarcinaceae. Ацетокластические представители семейства Methanosaetaceae преобладали в инкубационных вариантах с добавлением ацетата при обеих температурах. При обогащении осадков смесью метилированных соединений с водородом (метил-редуцирующий путь метаногенеза) образования метана не происходило, несмотря на увеличение в процессе инкубации относительного вклада некультивируемых представителей класса Thermoplasmata. Способность к метаногенезу доказана далеко не для всех архей этого класса (de Jong et al., 2018).
Представителей семейств Methanosarcinaceae и Methanosaetaceae часто детектируют в различных термокарстовых озерах (Negandhi et al., 2013; de Jong et al., 2018; Matheus Carnevali et al., 2018). Также выявляют гидрогенотрофных представителей порядков Methanomicrobiales, Methanobacteriales и Methanocellales (Negandhi et al., 2013; Matheus Carnevali et al., 2018). В термокарстовых озерах Аляски и Канады помимо известных метаногенов обнаружены некультивируемые метаногены кластера Rice Cluster II (Negandhi et al., 2013; de Jong et al., 2018). У доминирующего в тающей мерзлоте представителя этого кластера Candidatus “Methanoflorens stordalenmirensis” с помощью геномного анализа удалось обнаружить гены, участвующие в гидрогенотрофном метаногенезе (Mondav et al., 2014).
Интересной для понимания метаногенеза, хотя и не относящейся конкретно к термокарстовым озерам, является работа Вея и соавт. (Wei et al., 2018), в которой на примере Тибетского нагорья (Qinghai-Tibetan Plateau, Китай), самой большой высокогорной области вечной мерзлоты в мире, показано, как процесс таяния мерзлоты влияет на активность и состав микробного сообщества. Оказалось, что таяние мерзлоты приводит к смене доминирующей метаногенной составляющей сообщества и переключению пути метаногенеза с гидрогенотрофного на ацетокластический. В ответ на таяние мерзлоты резко возрастает численность активных метаногенов, и увеличение эмиссии метана является прямым следствием этого (Wei et al., 2018).
Следует отметить, что помимо метаногенов в термокарстовых озерах Аляски выявлены разнообразные некультивируемые археи, относящиеся к филумам Lokiarchaeota (суперфилум Asgard), Woesearchaeota и Pacearchaeota (суперфилум DPANN), Bathyarchaeota и Thaumarchaeota (суперфилум TACK), Soil Crenarchaeota Group (SCG), а также анаэробные метанотрофы кластеров ANME (de Jong et al., 2018; Matheus Carnevali et al., 2018), о которых речь пойдет ниже.
Окисление метана
Метанотрофы, наряду с метаногенами, играют ключевую роль в процессе круговорота метана, окисляя этот парниковый газ и снижая тем самым его эмиссию в атмосферу. В ассоциированных с вечной мерзлотой местообитаниях метанотрофы отвечают за потребление 20–60% образованного метана (Singleton et al., 2018 и ссылки там же). В термокарстовых озерах происходит как аэробное, так и анаэробное окисление метана.
Аэробные метанотрофные бактерии окисляют метан с использованием кислорода в качестве акцептора электронов (Hanson, Hanson, 1996). В пресных озерах высокие скорости окисления метана обычно приурочены к границе перехода окисленных условий в восстановленные (oxic-anoxic interface), т.е. наблюдаются в поверхностном слое осадков нестратифицированных водоемов или в оксиклине стратифицированных (Lidström, Somers 1984; Borell et al., 2011). При измерении аэробного окисления метана в 30 озерах Аляски выявлены факторы, регулирующие этот процесс. Показано, что зимой окисление метана главным образом зависит от содержания растворенного кислорода, тогда, как летом – от концентрации метана. Другой ключевой фактор, регулирующий уровень аэробного метанокисления, это тип вечной мерзлоты: озера, образовавшиеся при таянии мерзлоты едомного типа, характеризуются более высокими потенциальными скоростями метанокисления (средние значения: зима/лето – 0.18/0.71 мг л–1 сут–1) по сравнению с термокарстовыми и нетермокарстовыми озерами, сформированными в мерзлоте неедомного типа (средние значения: зима/лето – 0.08/0.29 мг л–1 сут–1) (Martinez-Cruz et al., 2015).
Аэробные метанотрофные представители классов Alpha- (тип II) и Gammaproteobacteria (тип I) являются типичными обитателями пресноводных озер, при этом в различных озерах, включая бореальные, наиболее часто выявляют гаммапротеобактерии рода Methylobacter (сем. Methylococcaceae) (Borrel et al., 2011; He et al., 2012; Crevecoeur et al., 2017; Martinez-Cruz et al., 2017; Rissanen et al., 2018; Kallistova et al., 2019). Преобладание представителей семейства Methylococcaceae также выявлено в отдельных термокарстовых озерах Аляски (de Jong et al., 2018; Matheus Carnevali et al., 2018) и Канады (Crevecoeur et al., 2015, 2017). Нетипичные метанотрофы обнаружены в мелководных термокарстовых водоемах Канады: в полигональных прудах (polygonal ponds) доминировали Crenothrix sp. – аэробные метанотрофы I типа, в канавных прудах (runnel ponds) преобладали представители Verrucomicrobia subdiv. 3, впервые обнаруженные в кислом горячем источнике Йеллоустонского Национального парка (Negandhi et al., 2014). Последовательности 16S рРНК представителей порядка Methylacidiphilales (Verrucomicrobia) найдены и в других Канадских термокарстовых прудах (Crevecoeur et al., 2015), а рода Crenothrix – в термокарстовых озерах Аляски (Matheus Carnevali et al., 2018). Метанотрофы II типа также вносят вклад в метанокисляющие сообщества термокарстовых прудов (thaw ponds) (Negandhi et al., 2014; Crevecoeur et al., 2017).
Влияние процесса оттаивания вечной мерзлоты на автохтонное метанотрофное сообщество было изучено на примере Стордален Майер (Stordalen Mire, Швеция). Выявлены последовательные изменения в структуре сообщества, активности и метанокисляющем потенциале. Состав и активность метанотрофного сообщества значительно изменялись по мере того, как прогрессировало оттаивание, начиная с пальсы до частично оттаявшего и полностью оттаявшего болота. Метанотрофная популяция, развивающаяся в градиенте оттаивания, состояла как из классических метанотрофов, так и из новых некультивируемых представителей кластера Upland Soil Cluster Alpha (USCα), а также потенциально метанотрофных Hyphomicrobiaceae. Вероятно, пальса, представляющая собой местообитание с низким содержанием метана, исходно была заселена факультативно метанотрофными представителями кластера USCα, окисляющими атмосферный метан, благодаря высокому сродству к субстрату. При постепенном оттаивании увеличивался вклад представителей семейства Methylocystaceae (тип II), которые справлялись с нестабильными условиями по метану, благодаря наличию pMMO-изоферментов и способности к использованию других альтернативных субстратов. В полностью оттаявших болотах, характеризующихся высокой концентрацией метана, преобладали и были активны облигатные метанотрофы семейства Methylococcaceae (Singleton et al., 2018).
Аэробное окисление метана было изучено и в термокарстовых озерах, расположенных на территории нашей страны. Зоной активных исследований в России служит дельта Лены, которая представляет собой район массового развития термокарстовых озер. Большинство исследований в дельте Лены выполнено немецкими учеными. Следует отметить работы таких авторов, как С. Лиебнер (C. Liebner), И. Буссманн (I. Bussmann) и Д. Вагнер (D. Wagner), посвященные аэробному окислению метана. Например, Осудар и соавт. (Osudar et al., 2016) определяли скорости метанокисления, вклад в процесс и разнообразие аэробных метанотрофов в водной толще озерно-речных комплексов (lake–river transects), расположенных на о. Самойловский (дельта Лены). Комплексы включали несколько мелководных термокарстовых озер (до 6 м глубиной), их стоки, участки реки неподалеку от стока и на середине реки; исследования проводили в летний сезон. В результате показано, что концентрация метана сильно варьировала в озерах и стоках, при этом в зоне стоков была зачастую выше, чем в озерах. Скорости метанокисления и обилие аэробных метанотрофов в целом коррелировали с содержанием метана. Скорости метанокисления сильно варьировали (от 0.2 до 480 нмоль л–1 ч–1), попадая, тем не менее, в диапазон значений, полученных ранее для других северных озер и рек. Максимальные концентрации метана и скорости метанокисления выявлены в зоне стоков и пойменной части озерных комплексов. Однако ни концентрация метана, ни обилие метанотрофов не могли полностью объяснить наблюдаемые различия в скоростях метанокисления. Например, в двух озерах концентрация метана была близка к таковой в реках, однако количество метанотрофов в озерах было ниже, а скорость метанокисления выше, чем в реках. Авторы предположили, что в этих озерах развивается специфическое метанотрофное сообщество, более эффективно окисляющее метан. Действительно, молекулярный анализ выявил различия в составе метанотрофных сообществ озер, стоков и рек. В реках преобладали представители рода Methylocystis (тип IIa) и кластера FW (freshwater lakes, пресноводные озера) (тип Ib). Метанотрофная популяция в озерах и стоках была более гетерогенной по составу: помимо универсальных метанотрофов, обитающих во всех исследованных сайтах, каждое озеро и сток содержали своих собственных, уникальных метанотрофов. Среди них можно выделить представителей типа Ib (кластеры RPC, JRC (rice field soil, почва рисовников)) и OSC (organic soil, почва с высоким содержанием органического вещества), типа Ia (род Methylobacter), типа IIa (род Methylocystis) и типа IIb (род Methylocapsa); классификация метанотрофов по Dumont et al., 2014. Стоки характеризовались наибольшим разнообразием метанотрофов. Метанотрофы, доминирующие в реках, являлись типичными обитателями водных экосистем, а в озерах и стоках – почвенных. Это указывает на заселение озер и стоков почвенной микрофлорой в результате эрозии и разлива. При этом в озерах преобладали метанотрофы I типа, а в реках и стоках – II типа. В целом, большинство исследователей отмечали преобладание в пресноводных местообитаниях метанотрофов I типа над метанотрофами II типа. Авторы цитируемой работы предполагают, что доминирование метанотрофов II типа может быть связано либо с достаточно стабильными условиями окружающей среды, либо с повышением температуры поверхностных вод в летний сезон (Osudar et al., 2016).
В термокарстовых озерах также обнаруживаются метилотрофные бактерии (Martinez-Cruz et al., 2017; de Jong et al., 2018), которые являются типичными спутниками аэробных метанотрофов в различных местообитаниях (Chistoserdova, 2015).
Процесс анаэробного окисления метана (AOM) с использованием различных акцепторов электронов (например, ${\text{SO}}_{4}^{{2 - }},$ ${\text{NO}}_{3}^{ - },$ ${\text{NO}}_{2}^{ - },$ Fe3+, Mn4+ и др.) в последние годы вызывает интерес у многих исследователей. Сульфат- и нитратзависимое AOM осуществляют ANME-археи по пути обратного метаногенеза, а нитритзависимое АОМ − бактерии группы NC10 (Ca. “Methylomirabilis”) по внутриаэробному пути (Welte et al., 2016; Mc-Glynn, 2017; Timmers et al., 2017). Механизмы металлозависимого АОМ и таксономическая принадлежность микроорганизмов, вовлеченных в этот процесс, до сих пор остаются неясными. Предполагалось участие в этом процессе различных бактерий и архей (Egger et al., 2015; Fu et al., 2016; Timmers et al., 2017; He et al., 2018), и лишь для архей кластера ANME-2d (Ca. “Methanoperedens nitroreducens”) получены доказательства их способности к металлозависимому АОМ наряду с нитратзависимым (Ettwig et al., 2016).
AOM-микроорганизмы обнаружены в термокарстовых озерах Аляски. Например, нитратзависимые археи Ca. “Methanoperedens” (ANME-2d) присутствовали в нативных образцах осадков оз. Эмейксаун (Emaiksoun) и оз. Безымянное (Unnamed), указывая на возможность протекания процесса AOM в этих термокарстовых озерах. Однако в инкубационных экспериментах с использованием ${\text{NO}}_{3}^{ - },$ ${\text{NO}}_{2}^{ - }$ в качестве акцепторов электронов не удалось показать АОМ-активность. В совокупности с данными об очень низкой концентрации ${\text{NO}}_{3}^{ - }$ in situ это могло свидетельствовать о том, что нитратзависимое АОМ не играет важной роли в изученных озерах (de Jong et al., 2018). Археи ANME-2d (нитратзависимое АОМ) и ANME-2 a/b (сульфатзависимое АОМ) были также обнаружены в озере Сакок (Sukok) (Matheus Carnevali et al., 2018).
Наиболее интригующие результаты были получены для оз. Волт (Vault), в котором выявлено участие в процессе АОМ аэробных метанотрофов рода Methylobacter. В составе микробного сообщества они окисляли в анаэробных условиях до 32% метана, образующегося в верхнем слое осадков этого мелководного железосодержащего термокарстового озера (Martinez-Cruz et al., 2017). Похожие результаты были получены ранее для Канадского пруда, в анаэробных наддонных водах которого обнаружен высокий относительный вклад последовательностей 16S рРНК аэробных метанотрофов (до 23%) (Crevecoeur et al., 2015). Такие результаты можно объяснить, например, периодическим внесением кислорода в наддонные воды при перемешивании или тем, что данные метанотрофы, попав из аэробной зоны в анаэробную, сохраняют жизнеспособность в течение некоторого, возможно, довольно длительного периода времени (Vonk et al., 2015). В случае с оз. Волт дальнейшие исследования выявили относительно высокий вклад в микробное сообщество осадков pmoA-генов аэробных метанотрофов и низкий – бактерий, родственных нитритзависимым Ca. “Methylomirabilis” (группа NC10) (Martinez-Cruz et al., 2018). В недавно опубликованной работе было показано, что, действительно, в оз. Волт от 41 до 83% растворенного метана потреблялось анаэробно. Агентами АОМ-процесса при этом выступали не аэробные метанотрофные бактерии, а анаэробные нитратзависимые археи ANME-2d (Ca. “Methanoperedens”) (Winkel et al., 2019). Аналогичная ситуация сложилась вокруг меромиктического (но не термокарстового) оз. Цуг (Швейцария), в анаэробном гиполимнионе которого детектированы физиологически активные метанотрофные гаммапротеобактерии. Для них предположено сопряжение процесса АОМ с нитрат/нитритредукцией или с использованием оксидов железа и марганца в качестве альтернативных кислороду акцепторов электронов (Oswald, 2016, 2017). Позднее, однако, в этом озере были обнаружены нитритзависимые АОМ-бактерии группы NC10 (Ca. “Methylomirabilis”), вклад которых в общую бактериальную популяцию достигал 27% (Graf et al., 2018). Таким образом, остается открытым вопрос: действительно ли аэробные метанотрофные гаммапротеобактерии участвуют в процессе АОМ в этих озерах, или анаэробные микроорганизмы (NC10 и ANME-2d), ранее не выявленные в силу методологических ограничений, отвечают за этот процесс? А может быть, в процессе АОМ участвуют обе группы, или какие-то новые организмы, которых мы пока не умеем детектировать?
Другие микроорганизмы термокарстовых озер
В термокарстовых озерах обнаружено высокое разнообразие бактерий, не участвующих напрямую в цикле метана. Среди них отмечено преобладание органотрофных Bacteroidetes, Betaproteobacteria и Actinobacteria (Negandhi et al., 2014, 2016; de Jong et al., 2018; Matheus Carnevali et al., 2018). Betaproteobacteria и Bacteriodetes ассоциированы с водоемами, обогащенными органическим веществом, и предпочитают лабильный углерод (Negandhi et al., 2014 и ссылки там же). Acidobacteria, Firmicutes, Verrumicrobia, Gemmatimonadetes и Chlroflexi также детектируются, например, в Канадских термокарстовых прудах (Negandhi et al., 2016). Высокая относительная численность аммоний-окисляющих бактерий порядка Nitrosomonadales и представителей порядка Anaerolineales выявлена в термокарстовых озерах Аляски (de Jong et al., 2018). Различие в составе микробных сообществ в зависимости от происхождения озерного метана (термогенное vs биогенное) показано на примере двух озер Аляски. В этих озерах потенциально активная (rRNA-доминирующая) микробная фракция состояла из таксонов, типичных для других мерзлотных экосистем – Bacteroidetes, Actinobacteria, Nitrospirae и Chloroflexi. Микроорганизмы, напрямую вовлеченные в круговорот метана (метаногены и метанотрофы) и азота (Nitrospirae), развивались в озере, в котором метан имел биогенное происхождение. В озере, обогащенном термогенным метаном, преобладали таксоны, участвующие в биогеохимической трансформации серы и фотосинтезе (Matheus Carnevali et al., 2018). Фототрофы широко распространены в термокарстовых озерах и представлены как оксигенными, так и аноксигенными организмами, включая эукариотические водоросли, цианобактерии, пикоцианобактерии, фотосинтезирующие серные и несерные бактерии (Vonk et al., 2015 и ссылки там же). Было предположено, что в начале своего образования озеро инокулируется микроорганизмами из окружающей почвы, поэтому исходное биоразнообразие сообщества идентично микробному составу инокулята (почвы). Затем микробное сообщество озера подвергается процессу видовой сукцессии. Таяние вечной мерзлоты приводит к более массовому переносу почвенных микроорганизмов в водные экосистемы (Crump et al., 2012; Vonk et al., 2015 и ссылки там же; Osudar et al., 2016).
В заключение раздела следует отметить, что проведенный нами обзор различных литературных источников выявил крайне мало микробиологических и биогеохимических исследований термокарстовых озер России, выполненных российскими учеными. В качестве единичных примеров можно привести работы Покровского и соавт. (2012), Широковой и соавт. (Shirokova et al., 2013) и Забелиной и соавт. (2017). В двух первых публикациях описывается одна и та же работа по исследованию просадок и термокарстовых озер, образованных при таянии пальсы центральной и северной части Западной Сибири (районы Нового Уренгоя и Ноябрьска, Ямало-Ненецкий автономный округ) в лесных, лесотундровых и тундровых ландшафтах, развитых на сплошной, прерывистой и островной зонах мерзлоты. Была определена численность гетеротрофного бактериопланктона (эвтрофных и олиготрофных бактерий) высевом на плотные среды. При этом отмечена тенденция снижения доли эвтрофной составляющей в процессе развития экосистемы озера с момента образования термокарстовой просадки до формирования зрелой экосистемы озера. В озерах глубиной >5 м отмечено уменьшение содержания гетеротрофного бактериопланктона с глубиной, с превалированием эвтрофной составляющей. Слабая корреляционная связь между численностью эвтрофного бактериопланктона и растворенным органическим углеродом указывала на то, что растворенный углерод находится в трудноусвояемой форме в “молодых” термокарстовых озерах, поступает в водную толщу в основном из донного торфа и при береговой абразии мерзлых торфяников. Однако в целом наблюдается тенденция к увеличению численности гетеротрофов и интенсивности аэробной деструкции органического вещества с увеличением содержания растворенного органического углерода. В исследованных водных объектах преобладал процесс аэробного разложения органического вещества бактериопланктоном; первичная продукция не была зафиксирована. Возможной причиной этого является то, что объектами изучения были в основном несформировавшиеся озерные экосистемы, где фитопланктонные сообщества отсутствуют в силу достаточно низких показателей кислотности среды (Покровский и соавт., 2012; Shirokova et al., 2013).
Забелина и соавт. (2017) исследовали термокарстовые озера Большеземельской тундры, расположенные в пределах тундровой зоны с преобладанием пальсы (Ненецкий автономный округ). В работе определяли общую численность и биомассу бактериопланктона, а также численность аэробных гетеротрофов высевом на плотные питательные среды. Отмечено увеличение численности бактерий в процессе развития озер от просадок и депрессий до больших сформировавшихся озер (Забелина и соавт., 2017).
ЗАКЛЮЧЕНИЕ
Термокарстовые озера образуются в результате процесса вытаивания высокольдистых многолетнемерзлых пород, приводящего к появлению просадочных форм рельефа. Активация процесса термокарста является одним из проявлений деградации вечной мерзлоты в условиях глобального потепления климата. Когда вечная мерзлота оттаивает, захороненные в ней биогенные элементы поступают в водоемы, где они становятся доступными для ремобилизации. В таких водоемах развиваются микробные сообщества, адаптированные к длительному периоду воздействия низких температур и различающиеся по интенсивностям аэробного и анаэробного метаболизма в зависимости от состава донных отложений, количества растворенного vs взвешенного органического вещества, минеральных веществ, ионов, лимнологических и гидрохимических особенностей водоема. Для многих термокарстовых озер характерны высокие скорости эмиссии метана, которая, в основном, происходит за счет дисбаланса между микробными процессами метаногенеза и метанокисления. Поэтому для оценки и прогнозирования величины эмиссии парниковых газов из термокарстовых озер в условиях таяния вечной мерзлоты приоритетное значение имеет исследование активности микроорганизмов цикла метана. Работы последних лет показали, что в термокарстовых озерах происходят как интенсивное образование метана по разным путям метаногенеза, так и аэробное и анаэробное метанокисление с участием разнообразных метаногенных и метанотрофных бактерий и архей. При этом остается открытым вопрос о механизмах и микроорганизмах, вовлеченных в процесс анаэробного окисления метана, за счет которого в термокарстовых озерах может потребляться до 80% метана. Кроме того, недостаточно хорошо изучены микробные сообщества, активно функционирующие подо льдом в течение длительного зимнего периода, крайне важные для водных экосистем северных регионов. К сожалению, приходится признать практически полное отсутствие серьезных микробиологических исследований термокарстовых озер России. Этот факт открывает широкие перспективы на ближайшие годы для детального изучения таксономического и функционального разнообразия микробных сообществ, обеспечивающих протекание основных процессов биогеохимического круговорота углерода в термокарстовых озерах.
Список литературы
Бойцов А.В. Геокриология и подземные воды криолитозоны. Учебное пособие. Тюмень: ТюмГНГУ, 2011. 178 с.
Брыксина Н.А., Евтюшкин А.В., Полищук Ю.М. Изучение динамики изменений термокарстовых форм рельефа с использованием космических снимков // Современные проблемы дистанционного зондирования Земли из космоса. 2007. Т. 4. № 2. С. 123–128.
Брыксина Н.А., Полищук В.Ю., Полищук Ю.М. База данных по термокарстовым озерам Западной Сибири на основе космических снимков и возможности ее практического использования // Современные проблемы дистанционного зондирования Земли из космоса. 2011. Т. 8. № 3. С. 175–181.
Ефимов А.И. К вопросу о развитии термокарстовых озер в Центральной Якутии // Исследование вечной мерзлоты в Якутской республике. Вып. 2. М.: Изд-во АН СССР, 1950. С. 98–114.
Забелина С.А., Широкова Л.С., Ковалев О.Д., Чупаков А.В., Чупакова А.А. Структура бактериопланктона термокарстовых озер большеземельской тундры // Труды ИБВВ РАН. 2017. Вып. 79 (82). С. 58–62.
Каллистова А.Ю., Меркель А.Ю., Тарновецкий И.Ю., Пименов Н.В. Образование и окисление метана прокариотами // Микробиология. 2017. Т. 86. № 6. С. 661–683.
Kallistova A.Yu., Merkel A.Yu., Tarnovetskii I.Yu., Pimenov N.V. Methane formation and oxidation by prokaryotes // Microbiology (Moscow). 2017. V. 86. P. 671–691.
Качурин С.П. Термокарст на территории СССР. Изд-во АН СССР, 1961. 289 с.
Кравцова В.И. Распространение термокарстовых озер в России // Вестн. Моск. ун-та. Сер. география. 2009. № 3. С. 33–42.
Кравцова В.И., Родионова Т.В. Исследование динамики площади и количества термокарстовых озер в различных районах криолитозоны России по космическим снимкам // Криосфера Земли. 2016. Т. 20. № 1. С. 81–89.
Kravtsova V.I., Rodionova T.V. Variations in size and number of thermokarst lakes in different permafrost regions: spaceborne evidence // Earth’s Cryosphere. 2016. V. 20. P. 75–81.
Кравцова В.И., Тарасенко Т.В. Динамика термокарстовых озер Центральной Якутии при изменениях климата с 1950 года // Криосфера Земли. 2011. Т. 15. № 3. С. 31–42.
Леин А.Ю., Лейбман М.О., Саввичев А.С., Миллер Ю.М., Пименов Н.В. Изотопно-биогеохимические особенности подземного пластового льда полуостровов Югорского и Ямал // Геохимия. 2003. Т. 41. № 10. С. 1084–1104.
Lein A.Yu., Leibman M.O., Savvichev A.S., Miller Yu.M., Pimenov N.V. Isotopic and biogeochemical characteristics of tabular ground ice on the Yugorskii and Yamal peninsulas // Geochem. Int. 2003. V. 41. P. 993–1012.
Леин А.Ю., Саввичев А.С., Лейбман М.О., Передня Д.Д. Ледовая летопись: пример расшифровки с помощью изотопных трассеров // Природа. 2005. № 7. С. 25–34.
Попов А.И., Розенбаум Г.Э., Тумель Н.В Криолитология: учебное пособие. М.: Изд-во МГУ, 1985. 239 с.
Общее мерзлотоведение / Под ред. Кудрявцева В.А. М.: Изд-во Моск. ун-та, 1978. 463 с.
Покровский О.С., Широкова Л.С., Кирпотин С.Н. Микробиологические факторы, контролирующие цикл углерода в термокарстовых водных объектах Западной Сибири // Вестн. Томского гос. ун-та. Биология. 2012. № 3 (19). С. 199–217.
Полищук Ю.М., Брыксина Н.А., Куприянов М.А. Сравнительный анализ изменений размеров озер в зоне мерзлоты Западной и восточной Сибири по космическим снимкам // Геоинформатика. 2016. № 1. С. 64–67.
Полищук Ю.М., Куприянов М.А., Брыксина Н.А. Дистанционное исследование динамики и площади озер в сплошной криолитозоне Сибири // География и природные ресурсы. 2017. № 3. С. 164–170.
Ривкина Е.М., Краев Г.Н., Кривушин К.В., Лауринавичюс К.С., Федоров-Давыдов Д.Г., Холодов А.Л., Щербакова В.А., Гиличинский Д.А. Метан в вечномерзлых отложениях Северо-Восточного сектора Арктики // Криосфера Земли. 2006. Т. 10. № 3. С. 23–41.
Родионова Т.В. Исследование динамики термокарстовых озер в различных районах криолитозоны России по космическим снимкам. Дисс. … канд. геогр. н. Москва, 2013. 196 с.
Романовский Н.Н. Подземные воды криолитозоны / Под ред. Всеволожского В.А. М.: МГУ, 1983. 232 с.
Романовский Н.Н. Основы криогенеза литосферы. М.: Изд-во Моск. ун-та, 1993. 336 с.
Соловьев П.А. Аласный рельеф Центральной Якутии и его происхождение // Многолетнемерзлые породы и сопутствующие им явления на территории Якутской АССР. М.: Изд-во АН СССР, 1962. С. 38–53.
Сумгин М.И. Вечная мерзлота почвы в пределах СССР. Изд-во НКЗ, Дальневосточная геофизическая обсерватория, Владивосток, 1927. 372 с. http://books.e-heritage.ru/book/10080861.
Borrel G., Jézéquel D. Biderre-Petit C., Morel-Desrosiers N., Morel J.P., Peyret P., Fonty G., Lehours A.C. Production and consumption of methane in freshwater lake ecosystems // Res. Microbiol. 2011. V. 162. P. 832–847.
Bouchard F., MacDonald L.A., Turner K.W., Thienpont J.R., Medeiros A.S., Biskaborn B.K., Korosi J., Hall R.I., Pienitz R., Wolfe B.B. Paleolimnology of thermokarst lakes: a window into permafrost landscape evolution // Arctic Sci. 2017. V. 3. P. 91–117.
Calmels F., Allard M., Delisle G. Development and decay of a lithalsa in northern Quebec: a geomorphological history // Geomorphology. 2008. V. 97. P. 287–299.
Chistoserdova L. Methylotrophs in natural habitats: current insights through metagenomics // Appl. Microbiol. Biotechnol. 2015. V. 99. P. 5763–5779.
Ciais P., Sabine C., Bala G., Bopp L., Brovkin V., Canadell J., Chhabra A., DeFries R., Galloway J., Heimann M., Jones C., Le Quéré C., Myneni R.B., Piao S., Thornton P. Carbon and other biogeochemical cycles // Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change / Eds. Stocker T.F., Qin D., Plattner G.-K., Tignor M., Allen S.K., Boschung J., Nauels A., Xia Y., Bex V., Midgley P.M. Cambridge: Cambridge University Press, UK, 2013. P. 465–570. Crate S., Ulrich M., Habeck J.O., Desyatkin A.R., Desyatkin R.V., Fedorov A.N., Hiyama T., Iijima Y., Ksenofontov S., Mészáros C., Takakura H. Permafrost livelihoods: A transdisciplinary review and analysis of thermokarst-based systems of indigenous land use // Anthropocene. 2017. V. 18. P. 89–104.
Crevecoeur S., Vincent W.F., Comte J., Lovejoy C. Bacterial community structure across environmental gradients in permafrost thaw ponds: Methanotroph-rich ecosystems // Front. Microbiol. 2015. V. 6. 192. https://doi.org/10.3389/fmicb.2015.00192
Crevecoeur S., Vincent W.F., Comte J., Matveev A., Lovejoy C. Diversity and potential activity of methanotrophs in high methane-emitting permafrost thaw ponds // PLoS One. 2017. V. 12. e0188223.
Crump B.C., Amaral-Zettler L.A., Kling G.W. Microbial diversity in arctic freshwaters is structured by inoculation of microbes from soils // ISME J. 2012. V. 6. P. 1629–1639.
de Jong A.E.E., In 't Zandt M.H., Meisel O.H., Jetten M.S.M., Dean J.F., Rasigraf O., Welte C.U. Increases in temperature and nutrient availability positively affect methane-cycling microorganisms in Arctic thermokarst lake sediments // Environ. Microbiol. 2018. V. 20. P. 4314–4327.
Drozdov D.S., Malkova G.V., Romanovsky V.E., Rumyantseva Ya.V., Abramov A.A., Konstantinov P.Ya., Sergeev D.O., Shiklomanov N.I., Kholodov A.L., Ponomareva O.E., Streletskiy D.A. Monitoring of permafrost in Russia. Russian database and the international GTN-P project // Proc. 68th Canadian Geotechnical Conf., Quebec, Canada, September 2015.
Dumont M.G, Lüke C., Deng Y., Frenzel P. Classification of pmoA amplicon pyrosequences using BLAST and the lowest common ancestor method in MEGAN // Front Microbiol. 2014. V. 5. 34. https://doi.org/10.3389/fmicb.2014.00034
Elder C.D., Xu X., Walker J., Schnell J.L., Hinkel K.M., Townsend-Small A., Arp C.D., Pohlman J.W., Gaglioti B.V., Czimczik C.I. Greenhouse gas emissions from diverse Arctic Alaskan lakes are dominated by young carbon // Nature Climate Change. 2018. V. 8. P. 166–171.
Egger M., Rasigraf O., Sapart C.J., Jilbert T., Jetten M.S., Röckmann T., van der Veen C., Bândă N., Kartal B., Ettwig K.F., Slomp C.P. Iron-mediated anaerobic oxidation of methane in brackish coastal sediments // Environ. Sci. Technol. 2015. V. 49. P. 277–283.
Ettwig K.F., Zhu B., Speth D., Keltjens J.T., Jetten M.S.M., Kartal B. Archaea catalyze iron-dependent anaerobic oxidation of methane // Proc. Natl. Acad. Sci. USA. 2016. V. 113. P. 12792–12796.
Fu L., Li S.W., Ding Z.W., Ding J., Lu Y.Z., Zeng R.J. Iron reduction in the DAMO/Shewanella oneidensis MR-1 coculture system and the fate of Fe(II) // Water Res. 2016. V. 88. P. 808–815.
Gilichinsky D.A., Wagener S., Vishnevetskaya T.A. Permafrost microbiology // Permafrost and Periglacial Processes. 1995. V. 6. P. 281–291.
Graf J.S., Mayr M.J., Marchant H.K., Tienken D., Hach P.F., Brand A., Schubert C.J., Kuypers M.M.M., Milucka J. Bloom of a denitrifying methanotroph, “Candidatus Methylomirabilis limnetica”, in a deep stratified lake // Environ. Microbiol. 2018. V. 20. P. 2598–2614.
Grosse G., Jones B., Arp C. Thermokarst lakes, drainage, and drained basins // Treatise on Geomorphology / Ed. Shroder J.F. San Diego. USA: Academic Press, 2013. P. 325–353.
Hanson R.S, Hanson T.E. Methanotrophic bacteria // Microbiol. Rev. 1996. V. 60. P. 439–471.
He R., Wooller M.J., Pohlman J.W., Quensen J., Tiedje J.M., Leigh M.B. Shifts in identity and activity of methanotrophs in arctic lake sediments in response to temperature changes // Appl. Environ. Microbiol. 2012. V. 78. P. 4715–4723.
He Z., Zhang Q., Feng Y., Luo H., Pan X., Gadd G.M. Microbiological and environmental significance of metal-dependent anaerobic oxidation of methane // Sci. Total Environ. 2018. V. 610–611. P. 759–768.
Heslop J.K., Walter Anthony K.M., Sepulveda-Jauregui A., Martinez-Cruz K., Bondurant A., Grosse G., Jones M.C. Thermokarst lake methanogenesis along a complete talik profile // Biogeosci. 2015. V. 12. P. 4317–4331.
Hjort J., Karjalainen O., Aalto J., Westermann S., Romanovsky V.E., Nelson F.E., Etzelmüller B., Luoto M. Degrading permafrost puts Arctic infrastructure at risk by mid-century // Nat. Commun. 2018. V. 9. 5147. https://doi.org/10.1038/s41467-018-07557-4
Jorgenson M.T., Racine C.H., Walters J.C., Osterkamp T.E. Permafrost degradation and ecological changes associated with a warming climate in Central Alaska // Climatic Change. 2001. V. 48. P. 551–579.
Kallistova A., Kadnikov V., Rusanov I., Kokryatskaya N., Beletsky A., Mardanov A., Savvichev A., Ravin N., Pimenov N. Microbial communities involved in aerobic and anaerobic methane cycling in a meromictic ferruginous subarctic lake // Aquat. Microb. Ecol. 2019. V. 82. P. 1–18.
Lawrence D.M., Slater A.G. A projection of severe near-surface permafrost degradation during the 21st century // Geophys. Res. Let. 2005. V. 32. L24401. https://doi.org/10.1029/2005GL025080
Laurion I., Vincent W.F., MacIntyre S., Retamal L., Dupont C., Francus P., Pienitz R. Variability in greenhouse gas emissions from permafrost thaw ponds // Limnol. Oceanogr. 2010. V. 55. P. 115–133.
Leibman M.O., Arkhipov S.M., Perednya D.D., Savvichev A.S., Vanshtein B.G., Hubberten H.-W. Geochemical properties of the water–snow–ice complexes in the area of Shokalsky glacier, Novaya Zemlya, in relation to tabular ground-ice formation // Annals of Glaciology. 2005. V. 42. P. 249–254.
Leibman M.O., Kizyakov A.I., Plekhanov A.V., Streletskaya I.D. New permafrost feature – deep crater in Central Yamal (West Siberia, Russia) as a response to local climate fluctuations // Geography, Environment, Sustainability. 2014. V. 7. P. 68–79.
Lidström M.E., Somers L. Seasonal study of methane oxidation in Lake Washington // Appl. Environ. Microbiol. 1984. V. 47. P. 1255–1260.
Martinez-Cruz K., Sepulveda-Jauregui A., Walter Anthony K., Thalasso F. Geographic and seasonal variation of dissolved methane and aerobic methane oxidation in Alaskan lakes // Biogeosciences. 2015. V. 12. P. 4595–4606.
Martinez-Cruz K., Leewis M.C., Herriott I.C., Sepulveda-Jauregui A., Walter Anthony K., Thalasso F., Leigh M.B. Anaerobic oxidation of methane by aerobic methanotrophs in sub-Arctic lake sediments // Sci. Total Environ. 2017. V. 607–608. P. 23–31.
Martinez-Cruz K., Sepulveda-Jauregui A., Casper P., Walter Anthony K., Smemo K.A., Thalasso F. Ubiquitous and significant anaerobic oxidation of methane in freshwater lake sediments // Water Res. 2018. V. 144. P. 332–340.
Matheus Carnevali P.B., Rohrssen M., Williams M.R., Michaud A.B., Adams H., Berisford D., Love G.D., Priscu J.C., Rassuchine O., Hand K.P., Murray A.E. Methane sources in arctic thermokarst lake sediments on the North Slope of Alaska // Geobiology. 2015. V. 13. P. 181–197.
Matheus Carnevali P.B., Herbold C.W., Hand K.P., Priscu J.C., Murray A.E. Distinct microbial assemblage structure and archaeal diversity in sediments of Arctic thermokarst lakes differing in methane sources // Front. Microbiol. 2018. V. 9. 1192. https://doi.org/10.3389/fmicb.2018.01192
McGlynn S.E. Energy metabolism during anaerobic methane oxidation in ANME archaea // Microbes Environ. 2017. V. 32. P. 5–13.
Mondav R., Woodcroft B.J., Kim E.-H., McCalley C.K., Hodgkins S.B., Crill P.M., Chanton J., Hurst G.B., VerBerkmoes N.C., Saleska S.R., Hugenholtz P., Rich V.I., Tyson G.W. Discovery of a novel methanogen prevalent in thawing permafrost // Nat. Commun. 2014. V. 5. 3212. https://doi.org/10.1038/ncomms4212
Negandhi K., Laurion I., Whiticar M.J., Galand P.E., Xu X., Lovejoy C. Small thaw ponds: An unaccounted source of methane in the Canadian High Arctic // PLoS One. 2013. V. 8. e78204.
Negandhi K., Laurion I., Lovejoy C. Bacterial communities and greenhouse gas emissions of shallow ponds in the High Arctic // Polar Biol. 2014. V. 37. P. 1669–1683.
Negandhi K., Laurion I., Lovejoy C. Temperature effects on net greenhouse gas production and bacterial communities in arctic thaw ponds // FEMS Microbiol. Ecol. 2016. V. 92. fiw117.
Oliva M., Fritz M. Permafrost degradation on a warmer Earth: Challenges and perspectives // Curr. Opin. Environ. Sci. Health. 2018. V. 5. P. 14–18.
Osudar R., Liebner S., Alawi M., Yang S., Bussmann I., Wagner D. Methane turnover and methanotrophic communities in arctic aquatic ecosystems of the Lena Delta, Northeast Siberia // FEMS Microbiol Ecol. 2016. V. 92. fiw116.
Oswald K., Milucka J., Brand A., Hach P., Littmann S., Wehrli B., Kuypers M.M., Schubert C.J. Aerobic gammaproteobacterial methanotrophs mitigate methane emissions from oxic and anoxic lake waters // Limnol. Oceanogr. 2016. V. 61. P. S101–S118.
Oswald K., Graf J.S., Littmann S., Tienken D., Brand A., Wehrli B., Albertsen M., Daims H., Wagner M., Kuypers M.M., Schubert C.J., Milucka J. Crenothrix are major methane consumers in stratified lakes // ISME J. 2017. V. 11. P. 2124−2140.
Payette S., Delwaide A., Caccianiga M., Beauchemin M. Accelerated thawing of subarctic peatland permafrost over the last 50 years // Geophys. Res. Lett. 2004. V. 31. L18208.
Rissanen A.J., Saarenheimo J., Tiirola M., Peura S., Aalto S.L., Karvinen A., Nykänen H. Gammaproteobacterial methanotrophs dominate methanotrophy in aerobic and anaerobic layers of boreal lake waters // Aquat. Microb. Ecol. 2018. V. 81. P. 257–276.
Rivkina E., Laurinavichius K., McGrath J., Tiedje J., Shcherbakova V., Gilichinsky D. Microbial life in permafrost // Adv. Space Res. 2004. V. 33. P. 1215–1221.
Romanovsky V., Isaksen K., Drozdov D., Anisimov O., Instanes A., Leibman M., McGuire A.D., Shiklomanov N., Smith S., Walker D. Changing permafrost and its impacts // Snow, Water, Ice and Permafrost in the Arctic (SWIPA) / AMAP, Oslo, 2017. P. 65–102.
Savvichev A., Leibman M., Kadnikov V., Kallistova A., Pimenov N., Ravin N., Dvornikov Y., Khomutov A. Microbiological study of Yamal lakes: a key to understanding the evolution of gas emission craters // Geosciences. 2018. V. 8. 478. https://doi.org/10.3390/geosciences8120478
Saunois M., Bousquet P., Poulter B., Peregon A., Ciais P., Canadell J.G., Dlugokencky E.J., Etiope G., Bastviken D., Houweling S., Janssens-Maenhout G., Tubiello F.N., Castaldi S., Jackson R.B., Alexe M., Arora V.K., Beerling D.J., Bergamaschi P., Blake D.R., Brailsford G., Brovkin V., Bruhwiler L., Crevoisier C., Crill P., Covey K., Curry C., Frankenberg C., Gedney N., Höglund-Isaksson L., Ishizawa M., Ito A., Joos F., Kim H.-S., Kleinen T., Krummel P., Lamarque J.-F., Langenfelds R., Locatelli R., Machida T., Maksyutov S., McDonald K.C., Marshall J., Melton J.R., Morino I., Naik V., O’Doherty S., Parmentier F.-J.W., Patra P.K., Peng C., Peng S., Peters G.P., Pison I., Prigent C., Prinn R., Ramonet M., Riley W.J., Saito M., Santini M., Schroeder R., Simpson I.J., Spahni R., Steele P., Takizawa A., Thornton B.F., Tian H., Tohjima Y., Viovy N., Voulgarakis A., van Weele M., van der Werf G.R., Weiss R., Wiedinmyer C., Wilton D.J., Wiltshire A., Worthy D., Wunch D., Xu X., Yoshida Y., Zhang B., Zhang Z., Zhu Q. The global methane budget 2000–2012 // Earth. Syst. Sci. Data. 2016. V. 8. P. 697–751.
Sepulveda-Jauregui A., Walter Anthony K.M., Martinez-Cruz K., Greene S., Thalasso F. Methane and carbon dioxide emissions from 40 lakes along a north–south latitudinal transect in Alaska // Biogeosciences. 2015. V. 12. P. 3197–3223.
Serikova S., Pokrovsky O.S., Laudon H., Krickov I.V., Lim A.G., Manasypov R.M., Karlsson J. High carbon emissions from thermokarst lakes of Western Siberia // Nat. Commun. 2019. V. 10. 1552. https://doi.org/10.1038/s41467-019-09592-1
Shcherbakova V., Rivkina E., Laurinavichuis K., Pecheritsina S., Gilichinsky D. Physiological characteristics of bacteria isolated from water brines within permafrost // Int. J. Astrobiol. 2004. V. 3. P. 37–43.
Shcherbakova V., Yoshimura Y., Ryzhmanova Y., Taguchi Y., Segawa T., Oshurkova V., Rivkina E. Archaeal communities of Arctic methane-containing permafrost // FEMS Microbiol. Ecol. 2016. V. 92. fiw135.
Shirokova L.S., Pokrovsky O.S., Kirpotin S.N., Desmukh C., Pokrovsky B.G., Audry S., Viers J. Biogeochemistry of organic carbon, CO2, CH4, and trace elements in thermokarst water bodies in discontinuous permafrost zones of Western Siberia // Biogeochemistry. 2013. V. 113. P. 573–593.
Singleton C.M., McCalley C.K., Woodcroft B.J., Boyd J.A., Evans P.N., Hodgkins S.B., Chanton J.P., Frolking S., Crill P.M., Saleska S.R., Rich V.I., Tyson G.W. Methanotrophy across a natural permafrost thaw environment // ISME J. 2018. V. 12. P. 2544–2558.
Timmers P.H., Welte C.U., Koehorst J.J., Plugge C.M., Jetten M.S., Stams A.J. Reverse methanogenesis and respiration in methanotrophic archaea // Archaea. 2017. 1654237.
Townsend-Small A., Åkerström F., Arp C.D., Hinkel K.M. Spatial and temporal variation in methane concentrations, fluxes, and sources in lakes in Arctic Alaska // J. Geophys. Res: Biogeosci. 2017. V. 122. P. 2966–2981.
Vonk J.E., Tank S.E., Bowden W.B., Laurion I., Vincent W.F., Alekseychik P., Amyot M., Billet M.F., Canário J., Cory R.M., Deshpande B.N., Helbig M., Jammet M., Karlsson J., Larouche J., MacMillan G., Rautio M., Walter Anthony K.M., Wickland K.P. Reviews and syntheses: Effects of permafrost thaw on Arctic aquatic ecosystems // Biogeosciences. 2015. V. 12. P. 7129–7167.
Walter K.M., Zimov S.A., Chanton J.P., Verbyla D., Chapin F.S. Methane bubbling from Siberian thaw lakes as a positive feedback to climate warming // Nature. 2006. V. 443. P. 71–75.
Walter K.M., Smith L.C., Chapin F.S. Methane bubbling from northern lakes: present and future contributions to the global methane budge // Philos. Trans. A Math. Phys. Eng. Sci. 2007. V. 365. P. 1657–1676.
Walter Anthony K.M., Zimov S.A., Grosse G., Jones M.C., Anthony P.M., Chapin F.S., Finlay J.C., Mack M.C., Davydov S., Frenzel P., Frolking S. A shift of thermokarst lakes from carbon sources to sinks during the Holocene epoch // Nature. 2014. V. 511. P. 452–456.
Wei S., Cui H., Zhu Y., Lu Z., Pang S., Zhang S., Dong H., Su X. Shifts of methanogenic communities in response to permafrost thaw results in rising methane emissions and soil property changes // Extremophiles. 2018. V. 22. P. 447–459.
Welte C.U., Rasigraf O., Vaksmaa A., Versantvoort W., Arshad A., Op den Camp H.J., Jetten M.S., Lüke C., Reimann J. Nitrate- and nitrite-dependent anaerobic oxidation of methane // Environ. Microbiol. Rep. 2016. V. 8. P. 941–955.
Wik M., Varner R.K., Walter Anthony K., MacIntyre S., Bastviken D. Climate-sensitive northern lakes and ponds are critical components of methane release // Nat. Geosci. 2016. V. 9. P. 99–105.
Winkel M., Sepulveda-Jauregui A., Martinez-Cruz K., Heslop J.K., Rijkers R., Horn F., Liebner S., Walter Anthony K.M. First evidence for cold-adapted anaerobic oxidation of methane in deep sediments of thermokarst lakes // Environ. Res. Commun. 2019. V. 1. 021002.
Zimov S.A., Schuur E.A.G., Chapin F.S. Permafrost and the global carbon budget // Science. 2006. V. 312. P. 1612–1613.
Дополнительные материалы отсутствуют.
Инструменты
Микробиология