

Микробиология, 2020, T. 89, № 5, стр. 535-546
Микробные биопленки в карстовой пещере Прощальная (Дальний Восток, Россия)
Л. М. Кондратьева a, О. С. Шадрина a, З. Н. Литвиненко a, *, Е. М. Голубева b, Н. С. Коновалова b
a Институт водных и экологических проблем ДВО РАН
680000 Хабаровск, Россия
b Институт тектоники и геофизики им. Ю.А. Косыгина ДВО РАН
680000 Хабаровск, Россия
* E-mail: zoyalitvinenko@gmail.com
Поступила в редакцию 24.07.2019
После доработки 31.10.2019
Принята к публикации 24.05.2020
Аннотация
В статье приводятся результаты исследования абиотических (состав капельной и трещинной вод) и биотических (состав и активность микробных комплексов) факторов, влияющих на формирование биопленок в карстовой пещере Прощальная (Дальний Восток, Россия). Рассмотрено влияние природных инфильтрационных вод на структуру культивируемых микробных комплексов и элементный состав натечного образования “лунное молоко”, формирующегося на стенах пещеры. Максимальное содержание таких элементов, как Fe и Mn было обнаружено в капельных водах в весенний период. Показана важная роль органических соединений, определяющих интенсивность микробиологических процессов на биогеохимическом барьере вода–порода и влияющих на элементный состав “лунного молока” и структуру сообществ, участвующих в осаждении карбонатов. Для исследования структуры микробных комплексов, входящих в состав “лунного молока” использовали методы культивирования и количественный ПЦР-анализ. В составе “лунного молока” были идентифицированы железобактерии родов Rhodoferax и Geothrix и представители рода Bacillus. Микроструктура и элементный состав “лунного молока” из пещеры Прощальная были исследованы с помощью сканирующего электронного микроскопа (VEGA 3 LMH, “Tescan”, Чехия) и кремний-дрейфового рентгеновского детектора X-MAX 80 мм2 (“Oxford Instruments”, Великобритания). На электронных изображениях масса “лунного молока” представлена различными микроструктурами и характеризовалась высокой гетерогенностью. Трубчатые микроструктуры имели богатый элементный состав (C, O, Ca, Fe, Mn, Si, Al и S) по сравнению с булавовидными образованиями, состоящими из карбоната кальция (СаСО3). Связывающий матрикс в составе “лунного молока” был представлен тонкими нановолокнами.
Согласно современной научной концепции большинство микроорганизмов в естественных экосистемах существуют в виде структурированных, прикрепленных к поверхности сообществ – биопленок (БП) (Николаев, Плакунов, 2007; Costerton et al., 2007). Бактерии в биопленках способны в определенных пределах быстро эволюционировать в ответ на изменения окружающей среды. В природных условиях БП чаще всего представлены сложными консорциумами. Для своего выживания микроорганизмы используют механизм коллективной координации – эффект “quorum sensing” (QS). QS играет важную роль в метаболизме хемолитотрофных бактерий, контролирует адгезию клеток к минеральным частицам, образование полисахаридов и формирование биопленок (Farah et al., 2005). Благодаря сложной структуре, внутри биопленки создаются симбиотические отношения между микроорганизмами разных видов, что позволяет им выжить в экстремальных местах обитания (Fleming, Wingender, 2010). К ним относятся подземные водоносные горизонты и поровое пространство горных пород, где микроорганизмы участвуют в важных биогеохимических процессах, происходящих в контактной зоне вода–порода (Perry et al., 2004).
Растворение карбонатных минералов и морфогенез карстовых полостей часто связывают с бактериальной активностью (Заварзин, Колотилова, 2001; Hill, Forti, 2007). Различные органические соединения и микроорганизмы, способные колонизировать поверхность горных пород, попадают в карстовые пещеры из наземных биотопов с инфильтрационными водами, паводковыми водами и атмосферным воздухом. В результате бактериального метаболизма карбонатные породы могут быть полностью разрушены или трансформированы в другие вторичные минералы (Максимович, Хмурчик, 2012).
В результате анализа микробных сообществ, отобранных со стен пещер, расположенных в Испании, Чешской Республике и Словении (Porca et al., 2012), была выдвинута гипотеза о том, что заселение пещер микроорганизмами происходило путем проникновения воды сквозь вышележащие породы и почвы. Роль микроорганизмов в пещерах может быть связана с несколькими функциями, включая синтез нового органического вещества (хемолитоавтотрофия), трансформацию горных пород и разложение органических веществ (органотрофия). Было показано, что образование многих специфических натечных структур (спелеотем) в пещерах происходит в результате активности хемолитоавтотрофных организмов (Мазина, Семиколенных, 2010).
Микроорганизмы способны изменять минеральный состав и растворимость карбонатов, а также размер и морфологию кристаллов, о чем свидетельствуют крупные плохо растворимые кристаллы СаСО3, образующиеся в присутствии Bacillus pasteurii (Mitchell, Ferris, 2006). Также было показано, что микробные клетки могут выступать центрами осаждения и кристаллизации многих элементов (Barton, Northup, 2007). Следует отметить, что так называемые кальцитовые нановолокна, которые повсеместно распространены при взаимодействии воды с карбонатными породами, считают вторичными карбонатами. Однако их генезис до сих остается спорным вопросом, предполагаются разные механизмы их образования: в результате физико-химических процессов, минерализации палочковидных бактерий или осаждения кристаллов кальцита на органических матрицах; кальцинирования мицелия грибов или актинобактерий (Bindschedler et al., 2014; Maciejewska et al., 2017). Типичным вторичным кальцитом также считают игольчатые волокна кальцита (needle fibre calcite) (Cailleau et al., 2009). Однако проведенный изотопный анализ показал, что это различные кальциты, и это вносит еще больше путаницы в их происхождение. Поэтому четкое признание природы разных ультраструктурных кальцитов имеет большое значение для понимания процессов, с которым они могут быть связаны (Dupraz et al., 2009). Некоторые специалисты считают, что присутствие нановолокон или игольчатых волокон кальцита во вторичных отложениях CaCO3 можно использовать для характеристики палеоклимата и оценки экологической обстановки. Их соотношение может свидетельствовать о чередовании засушливых и полузасушливых климатических условий, хотя обе эти формы кальцитов могут встречаться и при влажном климате (Shankar, Achyuthan, 2007).
Особый интерес представляет биомасса натечного образования “лунное молоко” (moonmilk), которое формируется на стенах и сводах карстовых пещер и относится к наиболее распространенному типу карбонатных отложений, образующихся в пещерах. Описаны различные формы отложения “лунного молока”, в том числе инкрустации, пленки, толстые слои отложений вторичных карбонатов и каналы в глине (Summers et al., 2013; Cacchio et al., 2014).
В формировании микроструктур натечных образований значимую роль играет качественный состав поступающих поверхностных и подземных вод, который зависит от климатических и сезонных факторов. Например, в пещерах Италии в спелеотеме “лунное молоко” кроме кальция были определены органические углерод и азот, а также алюминий, кремний и железо (Summers et al., 2013). Однако их генезис не обсуждается, хотя авторы считают, что элементный состав в значительной степени определяется поступающей водой. В пещерах Красноярского края (Большая Орешная, Геологов-2) в составе натечного образования “лунное молоко” были определены карбонаты. В качестве обоснования был использован факт, что сезонный цикл влажности воздуха в пещере “увлажнение–просыхание” приводит к растворению поверхности коренной породы с немедленной рекристаллизацией карбоната (Мазина, Семиколенных, 2010).
Долгое время ученые учитывали только геологические процессы, влияющие на образование кальцитов. В последнее время стали уделять больше внимания микробиологическому фактору с использованием физиолого-геохимического подхода (Reitschuler et al., 2016). Присутствие микроорганизмов в формациях “лунного молока” наблюдается в пещерах по всему миру, от тропиков до высоких широт. В этом биологически обусловленном процессе верхние поверхностные слои активно формируются, в то время как более глубокие и старые слои постепенно обезвоживаются, инкрустируются и становятся неактивными. Поэтому метаболическая активность сложных микробных сообществ может играть существенную роль в образовании “лунного молока” (Portillo, Gonzales, 2011).
Во многих работах приведены убедительные данные о присутствии в “лунном молоке” различных групп микроорганизмов, показана их важная роль в процессе растворения карбонатов. Доказано экспериментально, что некоторые бактериальные культуры, выделенные из “лунного молока” на селективных средах, содержащих растворимые соли кальция, были способны к кристаллизации кальцита (Baskar et al., 2006). На основании исследований, проведенных в пещере Снежная (Абхазия) (Кондратьева и соавт., 2016), было выдвинуто предположение, что элементный состав подземных вод и структура микробных сообществ природных вод различного генезиса определяют элементный состав “лунного молока”.
Целью настоящей работы было определение активности микроорганизмов, выделенных из проб воды разного генезиса и “лунного молока” из пещеры Прощальная (Дальний Восток, Россия), с использованием методов культивирования на агаризованных питательных средах, выделения отдельных штаммов бактерий, определения их физиологической и биохимической активности и анализа наноструктур в массе “лунного молока” при помощи сканирующей электронной микроскопии с определением их элементного состава.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Место исследования. Пещера Прощальная относится к ландшафтному памятнику природы краевого значения и находится на правом берегу реки Сагды-Селанка (водораздел реки Амур) в Хабаровском крае (Россия). Ее правый склон крутой и сложен мраморированными известняками, которые подвержены карстовым процессам (Махинов и соавт., 2016). Подземная полость пещеры Прощальная представляет собой трехмерный лабиринт, состоящий из крупных (длиной до 120 м) залов, ходов, галерей и колодцев. В целом пещера вытянута в северо-восточном направлении и имеет три явно выраженных яруса, которые в свою очередь состоят из 6–8 этажей. Верхний ярус находится в обвальной стадии развития и характеризуется скоплением обрушившихся со свода и стен глыб, плит и блоков коренной породы. Средний ярус – место широкого распространения натечных образований. Нижний ярус частично обводнен. В пещере есть внутренний водоток, множество источников капельной и трещинной воды; стены и потолки покрыты различными натечными образованиями, включая “лунное молоко”. Карстовые воды Дальнего Востока России по химическому составу относят, чаще всего, к гидрокарбонатно-кальциевым, реже – к хлоридно-гидрокарбонатно-кальциевым со средней степенью минерализации. Значительную роль в пещере играют конденсационные и инфильтрационные воды (Берсенёв, 1989).
Исследование проб воды различного генезиса из пещеры Прощальная в 2015–2017 гг. показали, что за время циркуляции воды в пещере внутренний водоток обогащается ионами кальция, и его сток оказывает влияние на химический состав поверхностных вод р. Сагды-Селанка, протекающей рядом с пещерой. За весь период наблюдений максимальное содержание Ca, Fe и Mn было обнаружено в капельной воде в весенний период. Содержание индикаторного элемента кальция в капельной и трещинной водах в пещере Прощальная зависело от поступления талой воды и количества осадков. В мае 2015 года в исследуемом районе среднее количество осадков составляло 105 мм, в 2016 году – 136 мм, а в 2017 году – 48 мм. В ноябре 2015 года их количество составило 9.9 мм. Обогащение вод железом происходит как вследствие выщелачивания и растворения железистых минералов и пород, так и в результате обменных реакций, происходящих в поровых растворах горных пород (Шадрина, Кондратьева, 2018).
Количество культивируемых гетеротрофных бактерий в природных водах определяли на агаризованных питательных средах: крахмал-аммиачный агар (КАА), рыбо-пептонный агар (РПА) и РПА, разбавленный в десять раз. Для определения количества железомарганцевых бактерий использовали агаризованную среду Бромфильда. Посев на питательные среды осуществляли методом предельных разведений. Пищевые потребности бактериальных изолятов определяли в жидкой питательной среде с использованием различных источников углерода (моносахаридов, дисахаридов, аминокислот, спиртов и др.).
Пробы натечного образования “лунное молоко” разной консистенции были отобраны в пещере Прощальная (рис. 1) в мае 2014–2016 гг., ноябре 2015 г., феврале 2017 г. Микробиологические исследования проб “лунного молока” проведены с применением методов культивирования на агаризованных питательных средах и методом количественной ПЦР. Анализ структуры микробного сообщества натечного образования “лунное молоко” и определение общего количества культивируемых гетеротрофных бактерий проводили с использованием агаризованных сред (РПА, РПА:10, КАА). Культивирование проводили при 20–23°С в течение 7 сут. Для анализа активности роста выделенных штаммов на жидких питательных средах использовали спектрофотометр Unico 2800 (“United Products & Instruments”, США), длина волны 600 нм. Экспериментальные исследования проводили в трех повторностях.
Рис. 1.
Пещера Прощальная: (а) галерея Альбатроса; (б) натечное образование “лунное молоко” на стене.

Подсчет общего количества копий ДНК эубактерий и количества копий ДНК бактерий рода Rhodoferax и Geothrix проводили с использованием специализированных праймеров, разработанных в Берлинском техническом университете под руководством профессора Ульриха Шевчика. Показатели для калибровочных кривых: R2 > 0.99, эффективность от 92–98%. Количественную ПЦР проводили согласно общепринятым методикам (Kubista et al., 2006). Выделение ДНК проводили при помощи набора GeneMATRIX soil DNA purification kit в соответствии с инструкцией производителя (“EURx Ltd.”, Германия). Пробы исследуемой культуральной жидкости фильтровали через мембранные фильтры Sartorius stedium (Германия) из ацетата целлюлозы с размером пор 0.2 мкм.
Исследование микроструктуры и элементного состава “лунного молока” проводили в аналитическом центре Института тектоники и геофизики ДВО РАН на сканирующем электронном микроскопе VEGA 3 LMH (“Tescan”, Чехия) в режиме вторичных электронов с напылением платины, в сочетании с кремний-дрейфовым рентгеновским детектором X-MAX 80 мм2 (“Oxford Instruments”, Великобритания).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Растворенные минеральные вещества, поступающие с капельной и трещинной водой, образуют натечные образования (спелеотемы) и массивные корки в пещерах. Различные органические соединения и микроорганизмы, способные колонизировать поверхность горных пород, попадают в карстовые пещеры из наземных биотопов с инфильтрационными водами.
Исследования воды различного генезиса в пещере Прощальная. Исследования элементного состава природных вод показали, что содержание кальция в капельной и трещинной водах в пещере Прощальная зависит от поступления талой воды и количества осадков. Его максимальное содержание отметили в мае 2016 г., когда количество выпавших осадков было фактически в 3 раза больше, чем в 2017 г. Высокое содержание кальция связано с процессами растворения карбонатных пород при взаимодействии с природными водами, особенно при увеличении их кислотности в присутствии органических веществ. Содержание ионов кальция во внутреннем водотоке было значительно выше, чем в реке Сагды-Селанка (Шадрина, Кондратьева, 2018).
На примере пещеры Снежная (Абхазия) было показано, что при избытке карбоната кальция и в присутствии примесей азотсодержащих органических веществ in vitrо процесс растворения кристаллов СаСО3 в водной среде ускорялся. Определяющую роль играли присутствующие в воде микроорганизмы, способные синтезировать полимерный матрикс. В результате обильного бактериального слизеобразования формировались биопленки, синтезирующие органические кислоты, которые способствовали растворению карбонатов кальция (Кондратьева и соавт., 2016).
Органические вещества играют важную роль в определении интенсивности микробиологических процессов на биогеохимическом барьере вода–порода и влияют на содержание растворенных форм железа и марганца. В пробах воды из внутреннего водотока пещеры Прощальная, независимо от времени года, среди представителей микробного сообщества преобладали бактерии, предпочитающие низкие концентрации органических веществ, а также нитрифицирующие и железомарганцевые бактерии. Максимальное увеличение количества гетеротрофных микроорганизмов (6.28 × 103 КОЕ/мл) было зафиксировано во внутреннем водотоке в весенний период. Это может быть связано с таянием снега и поступлением органических веществ из почвы с поверхностными водами. В речной воде в это время наблюдали значительное уменьшение количества микроорганизмов из-за увеличения скорости потока. В осенний период в обоих водотоках отмечали снижение численности всех физиологических групп культивируемых микроорганизмов.
Большинство штаммов, выделенных из природных вод пещеры Прощальная, активно использовали сахара (β-галактозу, глюкозу, маннозу, арабинозу) и аминокислоты (аргинин, орнитин) в качестве источника углерода (табл. 1). Утилизация лактозы и сахарозы происходит в присутствии фермента карбогидразы, которая участвует одновременно в двух циклах – углерода и кальция (Müller et al., 2014).
Таблица 1.
Утилизация различных источников углерода штаммами, выделенными из воды различного генезиса в мае 2017 г. (пещера Прощальная)
| Источник углерода | Речные воды | Водоток в пещере | Капельная вода | Трещинная вода | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Бактериальные штаммы | ||||||||||
| В 44 | В 45 | В 46 | В 19 | В 21 | В 25 | В 26 | В 32 | В 38 | В 42 | |
| Глюкоза | + | + | + | + | + | + | – | + | + | + |
| Манноза | + | + | + | + | + | + | + | + | + | + |
| Арабиноза | + | + | + | + | + | + | + | + | + | + |
| Лактоза | + | – | + | + | – | + | + | + | + | – |
| Сахароза | + | – | – | + | – | + | – | – | + | – |
| Аргинин | + | + | + | + | + | + | + | + | + | + |
| Лизин | – | – | – | – | – | + | – | – | + | + |
| Орнитин | + | + | + | + | + | + | + | + | + | + |
| Инозит | + | – | + | + | + | + | – | + | + | + |
| Маннит | + | – | + | + | + | – | + | + | + | + |
| Сорбитол | – | – | + | + | + | – | + | + | + | + |
| β-Галактоза | + | + | – | – | – | – | – | – | – | + |
| Малонат натрия | + | – | + | + | + | + | + | + | + | + |
| Цитрат натрия | + | – | + | + | + | + | + | + | + | + |
| Мочевина | – | – | + | – | + | + | – | + | – | + |
Штаммы, выделенные из проб внутреннего водотока и трещинных вод, активно использовали спирты (инозит, сорбит и маннит). Самыми активными оказались штаммы, выделенные из трещинной воды, которые использовали более широкий спектр источников углерода. Возможно, среди них присутствовали аллохтонные почвенные микроорганизмы, поступившие с поверхностными водами.
Микробиологические исследования “лунного молока”. Проведенные исследования показали, что численность культивируемых бактерий, входящих в состав биопленки “лунное молоко”, значительно варьировала в зависимости от его консистенции (табл. 2). Во всех образцах “лунного молока” преобладали гетеротрофные микроорганизмы, предпочитающие высокие концентрации азотсодержащих органических веществ и незначительно отличающиеся по доминирующему морфотипу колоний. Более того, в тонком слое творожистой массы численность разных групп была выше, чем в более толстом слое творожистой массы “лунного молока”. При культивировании на КАА, содержащем крахмал в качестве источника углерода, численность бактерий была низкой в трех образцах. Предпочтения в использовании азотсодержащих ОВ представителями, входящими в состав “лунного молока”, можно объяснить особенностями их метаболизма. Экспериментально была показана сопряженность двух важных метаболических механизмов аммонификации пептидов/аминокислот и карбонатного осаждения кальция (Maciejewska et al., 2017).
Таблица 2.
Структура и численность микробных сообществ “лунного молока” из разных экологических ниш (май 2016 г.)
| Среда | Морфотипколонии | Пробы “лунного молока” | ||
|---|---|---|---|---|
| 1 (тонкий слой творожистой массы) | 2 (плотный слой слизистой массы) | 3 (толстый слой творожистой массы) | ||
| численность микроорганизмов, тыс. КОЕ/мл | ||||
| РПА | ПС | 111 ± 10.5 | 15 ± 3.9 | 142 ± 16.8 |
| ПМ | 41 ± 6.4 | 3 ± 1.7 | 5 ± 2.2 | |
| Ж | 74 ± 8.6 | – | 3 ± 1.7 | |
| Всего | 226 ± 25.5 | 18 ± 4.6 | 150 ± 12.7 | |
| РПА:10 | СП | 74 ± 8.6 | – | – |
| МЗ | 32 ± 5.6 | 5 ± 2.2 | – | |
| Б | 2 ± 1.4 | – | – | |
| СПал | – | 22 ± 4.7 | 6 ± 2.4 | |
| ММ | – | – | 86 ± 9.3 | |
| Всего | 108 ± 14.6 | 27 ± 6.9 | 92 ± 10.7 | |
| КАА | МС | 70 ± 8.4 | – | – |
| МТ | 22 ± 4.7 | более 500 | 52 ± 5.2 | |
| Всего | 92 ± 13.1 | – | 52 ± 5.2 | |
Следует отметить, что в тонком слое “лунного молока” среди первичных колонизаторов присутствовали бактерии, образующие слизистые полупрозрачные колонии (рис. 2). Согласно литературным данным, слизистые полимерные соединения способствует более интенсивной адгезии биопленок к твердой поверхности (Perry et al., 2004). В дальнейшем микробные комплексы, синтезирующие полимерные субстанции, оказывают влияние на растворение карбонатов кальция. Периодически из “лунного молока” на среде Бромфильда, содержащей Fe(OH)3, выделили колонии фиолетового цвета, характерные для железомарганцевых бактерий. Это может быть связано с поступлением железосодержащих подземных вод, влияющих на структуру формируемых биопленок “лунного молока”.
Рис. 2.
Колонии из суспензии “лунного молока” из пещеры Прощальная: (а) рост на РПА; (б) рост на РПА:10.
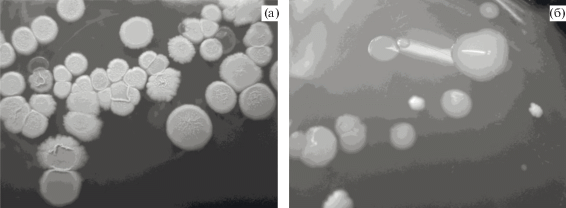
Минимальное разнообразие морфотипов колоний и низкая численность отмечены в образце “лунного молока” в виде плотной слизистой массы. Известно, что высокие концентрации карбоната кальция способны осаждаться в слизистой матрице и препятствовать развитию бактерий. Возможно, физиологическая адаптация бактерий к токсичным ионам Ca2+ происходит путем кальцификации поверхности бактериальных клеток. Такая активность создает условия для начала формирования биогенных кристаллов, которые способствуют образованию вторичных отложений CaCO3 в пещерах (Banks et al., 2010). Во многих случаях микроорганизмы и их внеклеточные полимерные вещества выступают в качестве эффективных центров формирования новых структур, которые могут привести к пассивной инкрустации биопленок (Flemming, Wingender, 2010).
Согласно молекулярным исследованиям в массе “лунного молока”, отобранной из пещеры Прощальная, присутствовали железобактерии родов Rhodoferax и Geothrix (табл. 3), которые обычно встречаются в железосодержащих подземных водах. Представители рода Rhodoferax являются психротолерантными факультативными анаэробами, которые используют Fe(OH)3 в качестве акцептора электронов. Geothrix fermentans часто находят в подземных водах в зоне восстановления Fe(OH)3. Было установлено, что железобактерии прикрепляются к поверхности минеральных частиц за счет продуцирования адгезивных биополимеров. Можно предположить, что Rhodoferax и Geothrix инициируют первую стадию формирования биопленки и создают условия, благоприятные для роста других гетеротрофных бактерий. Железо- и марганецокисляющие бактерии неоднократно обнаруживали в натечных образованиях некоторых пещер (Ghosh et al., 2017).
Таблица 3.
Результаты ПЦР-анализа проб “лунного молока” из пещеры Прощальная
| Образец | Общее количество копий генов ДНК эубактерий/мл | Количество копий генов ДНК рода Rhodoferax/мл | Количество копий генов ДНК рода Geothrix/мл |
|---|---|---|---|
| М 1 | 1.17 × 109 | 5.64 × 106 | 3.25 × 105 |
| М 2 | 1.08 × 109 | 1.44 × 106 | 3.76 × 105 |
Из массы “лунного молока” разной консистенции были выделены около 100 штаммов бактерий, которые отличались по спектру утилизируемых источников углерода. Штаммы бактерий, выделенных из плотных слоев творожистой массы, фактически не использовали для своей жизнедеятельности многие углеводы. Это может быть связано с замедлением биохимической активности при формировании биопленок на этапе продуцирования защитного слизистого полимерного матрикса и накопления в нем различных химических элементов, в том числе Al, Si, Fe и Mn. Из тонкого слоя слизистой массы “лунного молока” выделены штаммы, активно утилизирующие глюкозу, маннозу, арабинозу, лактозу и сахарозу. Большинство штаммов, выделенных из творожистой и плотной массы “лунного молока”, предпочитали в качестве источника углерода лактозу, глюкозу и маннозу, но не использовали для своего метаболизма аминокислоты. Такие потребности обусловлены отбором тех форм, которые могли активно развиваться за счет продуцирования другими членами консорциума полимерного матрикса (экзополимеров).
Согласно идентификационной таблице с биохимическими тестами (Васильев, 2013) в биомассе “лунного молока” были идентифицированы бактерии рода Bacillus. Можно предположить, что поверхностные воды, дренирующие почву и карстовые породы, являются источником проникновения микроорганизмов рода Bacillus в состав биопленок “лунного молока”. Два штамма были отнесены к бактериям Bacillus mesentericus и B. megaterium (табл. 4). Они относятся к типичным почвенным хемоорганотрофным микроорганизмам, которые встречаются в пресных водах, участвуют в процессах круговорота азота и могут восстанавливать железо. Ранее сообщалось, что бактерии рода Bacillus активно продуцируют полимерную слизь и выступают в качестве катализаторов выветривания горных пород и биогенной минерализации (Ercole et al., 2007). На стадии прорастания спор бактерий рода Bacillus происходит выделение в среду кальциевой соли дипиколиновой кислоты (пиридин-2,6-дикарбоксиловая кислота), которая способствует прикреплению бактериальных клеток к поверхности горных пород. В присутствии Bacillus pasteurii наблюдали образование крупных кристаллов СаСО3 (Mitchel, Ferris 2006).
Таблица 4.
Биохимические свойства штаммов бактерий, выделенных в мае 2015 г. из “лунного молока” пещеры Прощальная
| Тесты | Бактерии рода Bacillus | |||
|---|---|---|---|---|
| штамм П 1 | B. mesentericus | штамм П 2 | B. megaterium | |
| Анаэробный рост | – | – | – | – |
| Каталаза | + | + | + | + |
| Тест Фогес−Проскауера | – | – | – | – |
| Подвижность | + | + | – | +/– |
| Рост при 45°С | + | + | + | + |
| Цитрат | + | + | + | + |
| Мочевина | – | – | – | – |
| H2S | + | + | – | – |
| Арабиноза | + | + | + | + |
| Маннит | + | + | +/– | + |
| Раффиноза | – | – | + | + |
| Ксилоза | + | + | – | +/– |
| Салицин | – | – | – | – |
Молодые клетки, продуцирующие полимерный матрикс, формируют биопленки, в которых аккумулируются различные элементы, и происходит их кристаллизация. При электронном микроскопировании одного из штаммов бактерий, выделенного из “лунного молока”, был зафиксирован начальный этап кристаллообразования (рис. 3).
Рис. 3.
СЭМ изображение начального этапа кристаллообразования бактериальными клетками. Масштабная метка – 5 мкм.

Таким образом, выделенные штаммы спорообразующих бактерий рода Bacillus могут принимать участие в формировании и накоплении биомассы “лунного молока” в пещере Прощальная, способствуя в дальнейшем процессам кристаллизации и образования вторичных биоминералов. Источниками питания для формирующихся биопленок могут выступать органические вещества, поступающие с инфильтрационными водами, и метаболиты хемолитотрофных первопоселенцев.
Структура и элементный состав “лунного молока” из пещеры Прощальная. Ранее в результате сканирующей электронной микроскопии “лунного молока” из пещеры Снежная (Абхазия) нами было установлено, что его элементный состав определяется микроструктурной организацией. Так, в нем были обнаружены микроструктуры в виде пластинок с высоким содержанием Mg (около 22 вес. %), кубические кристаллы с высоким содержанием Fe. Доля Ca в этих микроструктурах была невысокой (Кондратьева и соавт., 2016).
Визуально гомогенная слизистая масса “лунного молока” из пещеры Прощальная при электронном микроскопировании была представлена различными наноструктурами. При увеличении более тысячи раз на электронных изображениях были обнаружены различные по форме образования в виде тонких нитей (волокон), трубок (полутрубок) и булавовидных структур (рис. 4). Нитчатые наноструктуры были похожи на поликристаллические кальцитовые волокна, обнаруженные в “лунном молоке” из Бельгийской пещеры (Collembola) (Maciejewska et al., 2017). Отмечено сходство трубчатых структур “лунного молока” из пещеры Прощальная с кальцитовыми трубками, обнаруженными в карбонатных породах Швейцарии (Bindschedler et al., 2014).
Рис. 4.
СЭМ изображения массы “лунного молока” из пещеры Прощальная при разном увеличении: а, б, в – общий вид массы “лунного молока”, увеличение в 50, 500, 1000 раз; г – булавовидные, трубчатые и нитчатые микроструктуры, увеличение в 10 тыс. раз; д – трубчатые и нитчатые структуры, увеличение в 15 тыс. раз; е – трубчатые микроструктуры (из публикации Bindschedler et al., 2014).

В элементном составе наноструктур, присутствующих в “лунном молоке” пещеры Прощальная, доминировали C, O, Ca, что подтверждает их карбонатное происхождение. Самый богатый элементный состав был характерен для трубчатых наноструктур, в них присутствовали такие элементы как Fe, Al, Si и реже Mn. В отдельных локусах редко встречались микропримеси магния и серы (табл. 5). Максимальное содержание Ca зафиксировано в трубчатых микроструктурах “лунного молока”. Булавовидные образования характеризовались более бедным элементным составом, характерным для карбонатов кальция, хотя в них присутствовали примеси натрия. В отличие от “лунного молока” из пещеры Снежная, в аналогичном по визуальным признакам натечном образовании из пещеры Прощальная магний отсутствовал. Ранее было высказано предположение, что в период формирования массы “лунного молока” в пещере Прощальная на фоне снижения количества осадков происходило в основном осаждение кальцитов, не содержащих магний, несмотря на его присутствие в грунтовых водах (Шадрина, Кондратьева, 2018).
Таблица 5.
Элементный состав наноструктур, входящих в состав “лунного молока” в пещере Прощальная (атомный вес, %) (Шадрина, Кондратьева, 2018)
| Элементы | Атомный вес, % | ||
|---|---|---|---|
| трубчатые наноструктуры | булововидные наноструктуры | мицелиальные формы | |
| С | 18–20 | 20–24 | 20–23 |
| О | 58–60 | 62–68 | 62–66 |
| Ca | 15–20 | 6–13 | 10–15 |
| Na | – | 0.15–0.5 | 0.5–1.32 |
| Fe | 0.18–0.26 | – | – |
| Mn | 0.10–0.11 | – | – |
| Si | 0.7–1.04 | – | 0.08–0.19 |
| Al | 0.53–0.73 | – | – |
| Mg | – | – | – |
| S | 0.05–0.07 | – | 0.26–0.75 |
Стоит подчеркнуть, что локальная аккумуляция нерастворимых форм железа в природных системах тесно связана с микробиологической активностью. Гидроксиды Fe часто называют биогенными, они создают высокореакционную поверхность, ответственную за вторичное отложение широкого спектра других микроэлементов (Hill, Forti, 2007).
Не менее важную роль в аккумуляции элементов на поверхности бактериальных клеток играют экзогенные полимерные субстанции (ЭПС), которые выполняют множество различных эколого-физиологических функций. Способность продуцировать экзополимеры позволяет микроорганизмам контролировать условия в окружающей среде (Ras et al., 2011). При высоких концентрациях кальция в щелочных условиях (рН > 8.4) осаждение карбонатных минералов может происходить внутри матрицы ЭПС. Сродство к кальцию снижается в присутствии конкурирующих двухвалентных катионов, таких как магний, железо, барий и стронций. Кислотные функциональные группы экзополимеров способствуют так называемой органоминерализации карбонатов (Braissant et al., 2007). При использовании X-ray фотоэлектронной спектроскопии была показана важная роль серосодержащих функциональных групп – тиолов, сульфиновых и сульфоновых кислот в осаждении карбонатов кальция (Beech et al., 1999). В состав полимерного матрикса могут входить сахара (рамноза, ксилоза, манноза, галактоза, глюкоза), органические кислоты (пируват, сукцинат), а также функциональные группы – сульфаты и/или фосфаты (Sutherland, 2001). В результате деградации ЭПС при участии гетеротрофных бактерий вновь происходит высвобождение ионов кальция и формирование вторичных биоминералов.
Присутствие в массе “лунного молока” разнообразных по элементному составу наноструктур можно связать с различными механизмами трансформации веществ, осуществляемых разными представителями микробного консорциума. Фактически натечное образование “лунное молоко” является биопленкой, в которой происходят синтез полимерного матрикса, аккумуляция на поверхности клеток разных сочетаний элементов и вторичное образование биоминералов со своеобразной наноструктурой.
На скорость и стадии образования массы “лунного молока” может влиять формирование биопленок в водоносном горизонте. Изменение физико-химических условий и поступление органических веществ в составе инфильтрующихся вод могут регулировать переотложение ионов кальция в поровом пространстве. Как следствие может происходить цементация порового пространства, прекращение инфильтрации природных вод (трещинной и капельной) и прекращение нарастания массы “лунного молока”.
Таким образом, полученные данные свидетельствуют о том, что биопленка “лунное молоко” из пещеры Прощальная сформирована при участии разнообразных микроорганизмов, которые отличаются своим метаболизмом и способностью синтезировать слизистые полимеры. Присутствие органических веществ в водной среде оказывает существенное влияние на структуру биопленок “лунного молока”. Под влиянием факторов окружающей среды, включая количество осадков, изменяется структура микробного сообщества и характер метаболизма, в частности, это может привести либо к растворению карбонатов кальция, либо к осаждению отдельных элементов (кальций, железо, марганец) с образованием биогенных минералов.
Согласно полученным результатам, можно предположить, что взаимодействие различных физиологических групп бактерий в составе биопленки “лунного молока” приводит к накоплению его биомассы и изменению соотношения между отдельными элементами за счет продуцирования или деградации экзополимеров. Наличие в биомассе натечных образований серосодержащих функциональных групп может быть признаком активной аккумуляции ионов кальция в полимерной матрице и последующей литификации этих новообразований.
Данные исследования приобретают актуальность в условиях глобального изменения климата. Существуют риски ускорения процессов карстообразования за счет активизации микробиологической активности на биогеохимическом барьере вода–порода, которая тесно связана с элементным составом подземных вод и антропогенным преобразованием почвенно-растительного покрова.
БЛАГОДАРНОСТИАвторы выражают благодарность В.О. Шадрину, руководителю экспедиции в пещеру Прощальная в 2015–2017 гг., Н.С. Коноваловой за проведение сканирующей электронной микроскопии и коллегам за помощь в отборе проб.СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВНастоящая статья не содержит результатов каких-либо исследований с использованием животных в качестве объектов.КОНФЛИКТ ИНТЕРЕСОВАвторы заявляют об отсутствии конфликта интересов.
Список литературы
Берсенёв Ю.И. Карст Дальнего Востока. М.: Наука, 1989. 172 с.
Васильев Д.А., Калдыркаев А.И., Феоктистова Н.А., Алёшкин А.В. Идентификация бактерий Bacillus cereus на основе их фенотипической характеристики. Ульяновск: НИИЦМиБ; УГСХА им. П.А. Столыпина, 2013. 98 с.
Заварзин Г.А., Колотилова Н.Н. Введение в природоведческую микробиологию. М.: Издательский Дом “Университет”, 2001. 256 с.
Николаев Ю.А., Плакунов В.К. Биоплёнка – “город микробов” или аналог многоклеточного организма? // Микробиология. 2007. Т. 76. С. 149–163.
Nikolaev Yu.A., Plakunov V.K. Biofilm – “city of microbes” or an analogue of multicellular organisms? // Microbiology (Moscow). 2007. V. 76. P. 125–138.
Кондратьева Л.М., Полевская О.С., Литвиненко З.Н., Голубева Е.М., Коновалова Н.С. Роль микробного сообщества в формировании натечного образования “лунное молоко” в карстовой пещере Снежная (Абхазия) // Микробиология. 2016. Т. 85. С. 598–608.
Kondratyeva L.M., Polevskaya O.S., Litvinenko Z.N., Golubeva E.M., Konovalova N.S. Role of the microbial community in formation of speleothem (moonmilk) in the Snezhnaya carst cave (Abkhazia) // Microbiology (Moscow). 2016. V. 85. P. 629–637.
Мазина С.Е., Семиколенных А.А. Различные формы лунного молока пещер России в свете проблемы генезиса // Пещеры. 2010. № 33. С. 34–44.
Максимович Н.Г., Хмурчик В.Т. Влияние микроорганизмов на минеральный состав и свойства грунтов // Вестник Пермского ун-та. 2012. Т. 3. № 16. С. 47–54.
Махинов А.Н., Крюкова М.В., Махинова А.Ф. Роль карстовых явлений в формировании долинного ландшафта реки Сагды-Селанка (центральный Сихотэ-Алинь) // Вестник ДВО РАН. 2016. № 5. С. 86–93.
Шадрина О.С., Кондратьева Л.М. Особенности формирования спелеотемы “лунное молоко” в пещере Прощальная (Хабаровский край) // Вопросы географии. 2018. № 147. С. 322–341.
Banks E.D., Taylor N.M., Gulley J., Lubbers B.R., Giarrizzo J.G., Bullen H.A., Hoehler T.M., Barton H.A. Bacterial calcium carbonate precipitation in cave environments: a function of calcium homeostasis // Geomicrobiol. J. 2010. V. 27. P. 444–454.
Barton H.A., Northup D.E. Geomicrobiology in cave environments: past, current and future perspectives // J. Cave. Karst Stud. 2007. V. 69. P. 163–178.
Baskar S., Baskar R., Mauclaire L., McKenzie J.A. Microbially induced calcite precipitation in culture experiments: Possible origin for stalactites in Sahastradhara caves, Dehradun, India // Curr. Sci. 2006. V. 90. P. 58–64.
Beech I.B., Zinkevich V., Tapper R., Gubner R., Avci R. Study of the interaction of sulphate-reducing bacteria exopolymers with iron using X-ray photoelectron spectroscopy and time-of-flight secondary ionization mass spectrometry // J. Microbiol. Methods. 1999. V. 36. P. 3–10.
Bindschedler S., Cailleau G., Braissant O., Millière L., Job D., Verrecchia E.P. Unravelling the enigmatic origin of calcitic nanofibres in soils and caves: purely physicochemical or biogenic processes? // Biogeosciences. 2014. V. 11. P. 2809–2825.
Braissant O., Decho A.W., Duprazi C., Glunk C., Przekop K.M., Visscher P.T. Exopolymeric substances of sulfate-reducing bacteria: interactions with calcium at alkaline pH and implication for formation of carbonate minerals // Geobiology. 2007. V. 5. P. 401–411.
Cacchio P., Ferrini G., Ercole C., Del Gallo M., Lepidi A. Biogenicity and characterization of moonmilk in the Grotta Nera (Majella National Park, Abruzzi, central Italy) // J. Cave Karst Studies. 2014. V. 76. P. 88–103.
Cailleau G., Verrecchia E.P., Braissant O., Emmanuel L. The biogenic origin of needle fibre calcite // Sedimentology. 2009. V. 56. P. 1858–1875.
Costerton J.W. The biofilm primer. Berlin: Springer Hiedelberg, 2007. 199 p.
Dupraz C., Reid R.P., Braissant O., Decho A.W., Norman R.S., Visscher P.T. Processes of carbonate precipitation in modern microbial mats // Earth-Science Rev. 2009. V. 96. P. 141–162.
Ercole C., Cacchio P., Botta A.L., Centi V., Lepidi A. Bacterially induced mineralization of calcium carbonate: the role of exopolysaccharides and capsular polysaccharides // Microsc. Microanal. 2007. V. 13. P. 42–50.
Farah C., Vera M., Morin D., Haras D., Jerez C.A., Guiliani N. Evidence for a functional quorum-sensing type AI-1 system in the extremophilic bacterium Acidithiobacillus ferrooxidans // Appl. Environ. Microbiol. 2005. V. 71. P. 7033–7040.
Flemming H.C., Wingender J. The biofilm matrix // Nature Rev. Microbiol. 2010. V. 8. P. 623–633.
Ghosh S., Paine E., Wall R., Kam G., Lauriente T., Sangarmangkang P.C., Derrick Horne D., Cheeptham N. In situ cultured bacterial diversity from iron Curtain Cave, Chilliwack, British Columbia, Canada // Diversity. 2017. V. 9 (3). P. 1–15. https://doi.org/10.3390/d9030036
Hill C.A., Forti P. Cave mineralogy and the NSS: past, present, future // J. Cave Karst Stud. 2007. V. 69. P. 35–45.
Kubista M., Andrade J.M., Bengtsson M., Forootan A., Jonake J., Lind K., Sindelka R., Sjoback R., Sjogreen B., Strombom L., Stahlberg A., Zoric N. The real-time polymerase chain reaction // Mol. Aspects Med. 2006. V. 27. P. 95–125.
Maciejewska M., Adam D., Naômé A., Martinet L., Tenconi E., Całusińska M., Delfosse P., Hanikenne M., Baurain D., Compère P., Carnol M., Barton H.A., Rigali S. Assessment of the potential role of Streptomyces in cave moonmilk formation // Front. Microbiol 2017. V. 8. P. 1181–1199.
Mitchell A.C., Ferris F.G. The influence of Bacillus pasteurii on the nucleation and growth of calcium carbonate // Geomicrobiol. J. 2006. V. 23. P. 213–226.
Müller W.E., Schlossmacher U., Schröder H.C., Lieberwirth I., Glasser G., Korzhev M., Neufurth M., Wang X. Enzyme-accelerated and structure-guided crystallization of calcium carbonate: role of the carbonic anhydrase in the homologous system // Acta Biomaterialia. 2017. V. 10. P. 450–462.
Perry T.D., Duckworth O.W., McNamara C.J., Martin S.T., Mitchell R. Effects of the biologically produced polymer alginic acid on macroscopic and microscopic calcite dissolution rates // Environ. Sci. Technol. 2004. V. 38. P. 3040–3046.
Porca E., Jurado V., Zgur-Bertok D., Saiz-Jimenez C., Pasic L. Comparative analysis of yellow microbial communities growing on the walls of geographically distinct caves indicates a common core of microorganisms involved in their formation // FEMS Microbiol. Ecol. 2012. V. 81. P. 255–266.
Portillo M.C., Gonzales J.M. Moonmilk deposits originate from specific bacterial communities in Altamira Cave (Spain) // Microbial Ecol. 2011. V. 61. P. 182–189.
Ras M., Lefebvre D., Derlon N., Paul E., Girbal-Neuhauser E. Extracellular polymeric substances diversity of biofilms grown under contrasted environmental conditions // Water Res. 2011. V. 45. P. 1529–1538.
Reitschuler C., Spötl C., Hofmann K., Wagner A.O., Illmer P. Archaeal distribution in moonmilk deposits from Alpine Caves and their ecophysiological potential // Microbial Ecol. 2016. V. 71. P. 686–699.
Shankar N., Achyuthan H. Genesis of calcic and petrocalcic horizons from Coimbatore, Tamil Nadu: Micromorphology and geochemical studies // Quatern. Internat. 2007. V. 175. P. 140–154.
Summers E.A., Paoletti M.G., Beggio M., Dorigo L., Pamio A., Gomiero T., Furlan C., Brilli M., Dreon A.L., Bertoni R., Squartini A. Comparative microbial community composition from secondary carbonate (moonmilk) deposits: implications for the Cansiliella servadeii cave hygropetric food web // Int. J. Speleol. 2013. V. 42. P. 181–192.
Sutherland I.A. The biofilm matrix – an immobilized but dynamic microbial environment // Trends Microbiol. 2001. V. 9. P. 222–227.
Дополнительные материалы отсутствуют.