

Микробиология, 2022, T. 91, № 6, стр. 647-665
Редактирование геномов метанотрофных бактерий: возможные мишени и доступный инструментарий
В. Н. Хмеленина a, С. Ю. Бут a, b, О. Н. Розова a, b, И. Ю. Ошкин b, Н. В. Пименов b, С. Н. Дедыш b, *
a Институт биохимии и физиологии микроорганизмов им. Г.К. Скрябина, Пущинский научный центр
биологических исследований Российской академии наук
142290 Пущино, Московская обл., Россия
b Институт микробиологии им. С.Н. Виноградского, ФИЦ Биотехнологии РАН
119071 Москва, Россия
* E-mail: dedysh@mail.ru
Поступила в редакцию 17.07.2022
После доработки 26.07.2022
Принята к публикации 26.07.2022
- EDN: NXTCDH
- DOI: 10.31857/S0026365622600584
Аннотация
Аэробные метанотрофные бактерии – это прокариотические микроорганизмы, обладающие уникальными ферментами, метанмонооксигеназами, позволяющими им использовать метан (СН4) в качестве ростового субстрата. Эта метаболическая особенность метанотрофов делает их привлекательными объектами биотехнологий, основанных на использовании метана для производства микробного белка и ряда целевых метаболитов. Растущий интерес к этим технологиям обусловлен высокой доступностью СН4, являющегося основным компонентом природного газа и биогаза, образуемого в результате анаэробной переработки органических отходов. Аэробные метанотрофы окисляют метан при температуре и давлении окружающей среды, поэтому перспективны в качестве биокатализаторов преобразования СН4 в продукты с добавленной стоимостью. Развитие биотехнологий конверсии метана с применением метанотрофов предполагает привлечение методов геномного редактирования для улучшения характеристик штаммов этих бактерий, используемых для производства. Специфика метаболизма СН4-использующих бактерий, а также сложности работы с этими объектами долгое время сдерживали развитие метаболической инженерии метанотрофов. В настоящем обзоре рассмотрены успехи последних десятилетий в области метаболической инженерии аэробных метанотрофов, описаны вероятные мишени и доступный инструментарий редактирования геномов этих микроорганизмов. Использование этого инструментария открывает возможности получения штаммов с биотехнологически ценными характеристиками, а также более глубокого изучения метаболических особенностей аэробных метанотрофов.
Аэробные метанотрофные бактерии – это группа прокариотических микроорганизмов, специализированных на использовании метана (СН4) в качестве единственного источника углерода и энергии (Hanson, Hanson, 1996; Гальченко, 2001; Trotsenko, Murrell, 2008; Chistoserdova, Lidstrom, 2013; Khmelenina et al., 2018). Способность к окислению метана обусловлена наличием у этих бактерий уникального фермента ‒ метанмонооксигеназы (ММО), которая существует в двух структурно и биохимически различных формах (Murrell et al., 2000; Murrell, Smith, 2010). Одна из форм, мембранная MMO (particulate MMO, pMMO), имеется у подавляющего большинства известных метанотрофов и локализована во внутрицитоплазматических мембранах (ВЦМ), выявляемых на ультратонких срезах клеток метанотрофов. Другая форма этого фермента, растворимая ММО (soluble MMO, sMMO), присутствует лишь у некоторых метанотрофов и локализована в цитоплазме. Структурные и каталитические характеристики этих форм ММО, а также история попыток гетерологичной экспрессии этих ферментов в неметанотрофных микроорганизмах освещены в недавнем обзоре (Khider et al., 2021).
Метанотрофные бактерии населяют широкий спектр экосистем (Knief, 2015) и играют ключевую роль в глобальном цикле углерода, снижая потоки парникового газа СН4 из природных и антропогенных местообитаний в атмосферу (Conrad, 2009). Помимо очевидной биосферной значимости аэробных метанотрофов, эти бактерии имеют большой, но пока лишь частично востребованный биотехнологический потенциал, обусловленный их способностью осуществлять конверсию метана в различные продукты с добавленной стоимостью (Strong et al., 2015; Pieja et al., 2017; Kalyuzhnaya et al., 2020). Растущий интерес к подобным биотехнологиям обусловлен сравнительной дешевизной и высокой доступностью СН4, являющегося основным компонентом природного газа и биогаза, образуемого в результате анаэробной трансформации органических отходов.
Первой реализованной в промышленном масштабе технологией с использованием метанотрофов явилось получение микробной биомассы, пригодной в качестве эффективной белковой добавки в корм сельскохозяйственных животных (“Гаприна”) (Григорян, Горская 1970; Hamer, Harrison, 1980; Егоров и соавт., 1990; Лалов, 1991). Уже более полувека активно изучаются возможности использования метанотрофов для получения разнообразных соединений, таких как каротиноиды, полигидроксибутират и полисахариды (Bothe et al., 2002; Ye et al., 2007), а также для биоремедиации экосистем, загрязненных хлорированными углеводородами (Murrell, Smith, 2010; Smith, Nichol, 2018). В исследованиях последних десятилетий основной упор делается на модификации биохимических путей метанотрофов для получения целевых метаболитов (Kalyuzhnaya et al., 2015; Henard et al., 2016; Garg et al., 2018; Nguyen et al., 2019; Nguyen, Lee, 2019; Pham et al., 2022b). Основные организмы-объекты этих работ по метаболической инженерии, ныне доступный инструментарий для редактирования геномов метанотрофных бактерий, а также возможные мишени направленных модификаций рассмотрены нами ниже.
ФИЛОГЕНЕТИЧЕСКОЕ И МЕТАБОЛИЧЕСКОЕ РАЗНООБРАЗИЕ АЭРОБНЫХ МЕТАНОТРОФОВ
Охарактеризованное разнообразие метанотрофных бактерий. Ныне известные аэробные метанотрофные бактерии формируют три основные филогенетические ветви, две из которых лежат в пределах филума Proteobacteria, а третья – в пределах Verrucomicrobia (рис. 1). Протеобактериальные метанотрофы, принадлежащие к классам Alpha- и Gammaproteobacteria, представлены наибольшим числом изученных культур и охарактеризованных таксонов (четыре семейства, около трех десятков родов и шести десятков видов) (Dedysh, Knief, 2018). Перечень этих таксонов включает организмы с различными фенотипическими характеристиками и предпочтениями к росту при различных температурах, рН среды, солености и проч. Метанотрофы класса Gammaproteobacteria высоко специализированы на использовании С1-соединений и не растут на полиуглеродных субстратах, таких как сахара или органические кислоты. В числе Al-phaproteobacteria, напротив, имеется ряд факультативных метанотрофов, способных помимо метана использовать ограниченное число органических соединений, таких как ацетат, пируват, этанол и некоторые короткоцепочечные алканы (Dedysh, Dunfield, 2010). Метанотрофные представители Verrucomicrobia формируют узкий филогенетический кластер, представленный семейством Methylacidiphilaceae, и являются термофильными или мезофильными ацидофилами, населяющими исключительно геотермальные экосистемы (Op den Camp et al., 2009; Schmitz et al., 2021). В отличие от протеобактериальных метанотрофов, представители Methylacidiphilaceae являются автотрофами и используют метан в качестве источника энергии, окисляя его до CO2, который затем фиксируют в цикле Кальвина (Khadem et al., 2011). Альтернативным источником энергии для этих метанотрофов является Н2 (Schmitz et al., 2021). Более полная информация об охарактеризованных на сегодняшний день аэробных метанотрофах представлена на веб-сайте Methanotroph Commons (http://www.methanotroph.org).
Рис. 1.
Филогенетическая дендрограмма, построенная на основании результатов сравнительного анализа 157 последовательностей геномов аэробных метанотрофных бактерий, представленных в базе данных GTDB (Parks et al., 2022). Анализ выполнен с помощью Genome Taxonomy Database toolkit, версия 2.0.0 (Chaumeil et al., 2020). Филогенетические ветви метанотрофных представителей Verrucomicrobia, Alpha- и Gammaproteobacteria выделены оранжевым, синим и лиловым цветами соответственно. Звездочками обозначены роды метанотрофов, представители которых становились объектами исследований с использованием генетических манипуляций. Уровни поддержки (бутстрепы) рассчитаны методом “maximum-likelihood” путем построения 100 альтернативных дендрограмм. Значения бутстрепов более 70% показаны черными кружками. В качестве корня использованы геномы бактерий филума Calditrichota. Маркер – 0.5 замен на одну аминокислотную позицию.

Метаболическое разнообразие аэробных метанотрофов. Метанотрофы окисляют CH4 до метанола (CH3OH) в присутствии кислорода с образованием одной молекулы воды и потреблением двух электронов и двух протонов. По сравнению с sMMO, pMMO имеет более высокое сродство к метану, ее функционирование коррелирует с более высокой эффективностью роста (Murrell et al., 2000). Для активности sMMO необходим НАДН, а непосредственным донором электронов для рММО является убихинол (UQH2), восстанавливаемый НАДН через комплекс I цепи переноса электронов. Не исключено также, что электроны, образующиеся при окислении метанола, передаются непосредственно на pMMO для окисления метана (Lieven et al., 2018). У метанотрофов, имеющих как растворимую, так и мембранную ММО, экспрессия этих ферментов регулируется ионами меди: рММО экспрессируется во время роста при высоком соотношении меди и биомассы, тогда как sММО ‒ при низком соотношении меди к биомассе (Murrell et al., 2000). Благодаря свойству sММО окислять широкий спектр субстратов, включая оксигенирование моноокиси углерода, некоторых алканов, алкенов, галогенированных метанов и циклических соединений (задокументировано более чем 100 субстратов, см. обзор Murrell, Smith 2010), изучаются возможности использования метанотрофных бактерий в биоремедиации для разложения ряда загрязняющих веществ, таких как трихлорэтилен (ТХЭ) и хлорированные углеводороды (Smith, Nichol, 2018), а также для синтеза гомохиральных эпоксидов (Jiang et al., 1996, 2010; Khider et al., 2021).
Окисление метанола в формальдегид катализирует метанолдегидрогеназа (МДГ). У метанотрофов имеются две формы МДГ, обе содержат пирролохинолинхинон (PQQ) в качестве простетической группы. Гетеротетрамерная MxaFI-МДГ содержит кальций в активном центре, тогда как односубъединичная XoxF-МДГ содержит редкоземельный элемент вместо кальция. Окислительные превращения формальдегида в формиат катализируют ферменты тетрагидрофолат (ТГФ)- и тетрагидрометаноптерин (ТГМП)-зависимых путей. Последнюю стадию окисления метана до CO2 катализирует формиатдегидрогеназа (ФДГ), поставляющая НАДН для монооксигенирования метана или в дыхательную систему для синтеза АТФ. У метанотрофов обнаруживаются до четырех изоформ ФДГ, причем некоторые из них катализируют обратимую реакцию.
Для ассимиляции углерода метанотрофы используют три биохимическиx пути: сериновый цикл, рибулозомонофосфатный (РМФ) путь и цикл Кальвина. Метанотрофы класса Alphaproteobacteria используют сериновый путь, в котором С1-соединение в форме N5,N10-метилен-тетрагидрофолата реагирует с глицином с образованием серина (Chistoserdova, Lidstrom, 2013). Затем аминогруппа серина переносится на глиоксилат с образованием глицина и гидроксипирувата, последний в серии реакций трансформируется в глицеральдегид-3-фосфат и пируват – универсальные метаболиты-предшественники всех компонентов клеток. Метанотрофы класса Gammaproteobacteria ассимилируют углерод посредством РМФ и серинового путей, функционирующих одновременно. Эта избыточность характерна для метанотрофов, но не для не растущих на метане метилотрофов (Бут и соавт., 2017). Первыми соединениями при построении С‒С связи в РМФ пути являются С6-фосфосахара: гексулозо-6-фосфат, образуемый конденсацией формальдегида и рибулозо-5-фосфата, далее изомеризацией гексулозо-6-фосфата образуется фруктозо-6-фосфат. Эти реакции катализируют два специфических фермента ‒ гексулозофосфатсинтаза (ГФС) и фосфогексулоизомераза (ФГИ). Геномный анализ выявил дупликацию, а иногда и трипликацию генов hps у некоторых быстрорастущих метанотрофов (Розова и соавт., 2017), однако влияние количества генов hps на физиологические характеристики штаммов предстоит определить. Для распада фосфосахаров и синтеза С3-соединений эти метанотрофы используют ферменты пентозофосфатного и гликолитического путей, путь Энтнера‒Дудорова и фосфокетолазу (бифидобактериальный шунт). Они имеют полный набор ферментов цикла трикарбоновых кислот (ЦТК), включая обходные пути для некоторых реакций цикла, а также механизмы для восполнения С4 интермедиатов (Fu et al., 2017; Khmelenina et al., 2019). Чрезвычайная гибкость метаболизма гаммапротеобактериальных метанотрофов предоставляет широкие возможности для управления потоками углерода на синтез нужных метаболитов. Представитель этого класса термотолерантный метанотроф Methylococcus capsulatus дополнительно ассимилирует СО2 через цикл Кальвина, служит модельным организмом при изучении метанотрофии как способа питания микроорганизмов и используется для разработки процессов культивирования в промышленном масштабе (Øverland et al., 2010). В 2013 году впервые на примере галотолерантного Methylotuvimicrobium alcaliphilum 20Z показана способность метанотрофов к ферментации формальдегида в условиях лимитирования роста кислородом и, как следствие, к накоплению в среде органических метаболитов (Kalyuzhnaya et al., 2013). Данная работа изменила представления о “токсичности” низкомолекулярных соединений для метанотрофов (Eccleston, Kelly, 1973) и послужила стимулом к более углубленному изучению метаболических путей трансформации метана. Поскольку галоалкалофильные метанотрофы демонстрируют устойчивый рост в широком диапазоне рН, солености, концентраций метанола, при низком парциальном давлении кислорода, а также проявляют повышенную устойчивость к загрязнению, они стали не только перспективными катализаторами биоконверсии метана, но и удобными объектами фундаментальных исследований. Метанотрофы с сериновым путем, будучи представлены как облигатными, так и факультативными формами по отношению к источнику углерода, имеют ряд преимуществ в качестве агентов биоремедиации и промышленной биотехнологии с использованием природного газа (Crombie, Murrell, 2011; Dunfield, Dedysh, 2014; Bordel et al., 2020).
СИСТЕМЫ РЕДАКТИРОВАНИЯ ГЕНОМОВ МЕТАНОТРОФОВ
Спонтанный мутагенез. По сравнению с традиционными объектами работ по геномному редактированию, такими, например, как Escherichia coli или Corynebacterium, манипуляции с метанотрофами существенно более длительны и трудоемки вследствие сложности культивирования этих бактерий. Первые работы по получению устойчивых к антибиотикам мутантов Methylococcus и Methylomonas основывались на использовании ультрафиолета или химических агентов (например, нитрозогуанидина, этилметансульфоната) (Harwood et al., 1972; Williams et al., 1977). Однако применение подхода спонтанного мутагенеза оказалось неэффективным, что объяснялось функционированием у метанотрофов высокоточных систем репарации ДНК. Однако впоследствии путем адаптивной лабораторной эволюции были получены штаммы, устойчивые к дихлорметану (Nicolaidis, Sargent, 1987) и рифамицину (Puri et al., 2015). Эти эксперименты продемонстрировали необходимость отработки подходящих методов скрининга и закрепления вносимых мутаций. Достаточно успешными оказались попытки транспозонного мутагенеза метанотрофов, которые завершились получением мутантов с делециями генов фиксации азота (Toukdarian, Lidstrom, 1984) или штаммов-продуцентов каротиноидов (Sharpe et al., 2007).
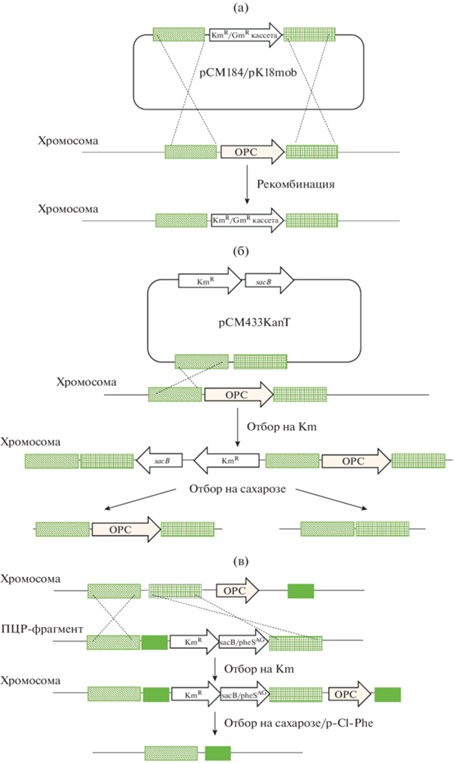
Двойная гомологичная рекомбинация. На сегодняшний день наиболее распространенным методом инактивации генов у метанотрофов является двойная гомологичная рекомбинация, сопровождаемая инсерцией кассеты устойчивости к антибиотикам, которая замещает ген-мишень (рис. 2а). Такая методология успешно применялась для Mm. alcaliphilum 20Z (Mustakhimov et al., 2010; Бут и соавт., 2013; Henard et al., 2018; But et al., 2020), Mc. capsulatus (Csaki et al., 2003; Henard et al., 2021), Methylosinus trichosporium OB3b (Stafford et al., 2003). Фрагменты ДНК, фланкирующие гены-мишени (“плечи” гена), клонируются в суицидальные (т.е. не реплицирующиеся в клетках реципиента) векторы pCM184 или pK18mob, по этим фрагментам происходит рекомбинация.
Рис. 2.
Способы генетического редактирования метанотрофов (схемы внесения модификаций в хромосому метанотрофов): а ‒ двойная гомологичная рекомбинация с использованием суицидальной плазмиды; б ‒ использование плазмиды с геном контрселекции sacB для получения безмаркерных мутаций; в ‒ использование ПЦР-фрагментов с генами контрселекции sacB или pheSAG для получения безмаркерных мутаций.
Методы трансформации. Для доставки плазмид в клетки метанотрофов используются два основных метода ‒ трансформация на основе конъюгации и трансформация на основе электропорации (Crombie, Murrell, 2011; Yan et al., 2016; Ro, Rosenzweig, 2018). Трансформацию с помощью конъюгации проводят посредством двухродительского скрещивания со штаммом E. coli S-17-1. Большое значение для генетических манипуляций имеет разработка эффективных методов электропорации с целью замены трудоемких методов конъюгации. Для повышения эффективности электропорации у метанотрофов, имеющих одновременно pММО и sММО, их выращивают на минеральной среде с пониженным содержанием CuSO4 (<10 мкМ), определяющей пониженный уровень ВЦМ (Ro, Rosenzweig, 2018; Nguyen et al., 2019). В недавних исследованиях эффективность электропорации была повышена посредством оптимизации нескольких ключевых параметров, таких как подготовка компетентных клеток, условия электропорации, время восстановления и концентрация антибиотика (Hu et al., 2021). Основываясь на гомологичной рекомбинации и эффективной трансформации, делеция гена и экспрессия гетерологичного гена достигнуты одновременно путем прямой электропорации линейных фрагментов ДНК, полученных с помощью ПЦР. Использование этого подхода позволило сконструировать продуцент изобутиральдегида на основе Mm. buryatense 5GB1 путем замены эндогенного гена fadE, кодирующего ацил-КоА-дегидрогеназу, участвующую в расщеплении жирных кислот, гетерологичным геном декарбоксилазы 2-кетоизовалериата kivD (Hu et al., 2021).
Направленный мутагенез может быть достигнут не только с помощью плазмид, но и линейных ПЦР-фрагментов, содержащих “плечи” гена-мишени и кассету устойчивости к антибиотику. Это позволяет сократить время создания конструкций для генетического редактирования (Puri et al., 2015, 2016; Yan et al., 2016; Ro, Rosenzweig, 2018). В двухэтапной процедуре делетируемый из хромосомы ген сначала заменяется кассетой устойчивости к антибиотикам, которая затем удаляется с помощью Cre рекомбиназы бактериофага Р1 (Sternbergand, Hamilton, 1981), выщепляющей маркерный ген, клонированный между сайтами связывания loxP (Marx, Lidstrom, 2002; Crombie, Murrell, 2011). Этим методом был получен мутант Methylocella silvestris BL2 по изоцитратлиазе, неспособный расти на С1-соединениях. Данная методика была также успешно применена для Methylomonas sp. DH-1, при этом при высеве на агаризованную среду без антибиотика для оптимальной работы Cre рекомбиназы вносили 4 мМ NiCl2 (Lee et al., 2021b).
Система безмаркерного аллельного обмена. Разработка протоколов контрселекции, не предполагающих повторного введения антибиотических маркеров, упрощает процесс получения множественных мутаций. Одним из эффективных методов редактирования геномов является использование гена sacB в качестве контрселектируемого маркера. Кодируемая геном sacB левансахараза превращает сахарозу в леван, приводя к лизису клеток в присутствии определенной концентрации сахарозы (Recorbet et al., 1993). Метод успешно применялся для получения безмаркерных мутаций у метилотрофов (Marx, 2008). При этом редактирование проводится в два этапа. Сначала происходит встраивание плазмиды в хромосому по одному из гомологичных плечей, а затем при селекции на сахарозе может восстановиться аллель дикого типа или выщепиться целевой ген (рис. 2б). Этим методом с использованием плазмид, несущих ген sacB, у Mm. buryatense 5GB1 были инактивированы гены синтеза гликогена (Puri et al., 2015), а у Mm. alcaliphilum 20Z ‒ нокаутированы одновременно два кластера генов синтеза и распада гликогена и ген sps, кодирующий сахарозофосфатсинтазу (Бут и соавт., 2020). Подавление синтеза гликогена у метанотрофов имеет практическое значение, поскольку они способны накапливать гликоген до 30% сухой массы клеток в условиях лимитирования роста азотом (Khmelenina et al., 1999; Ешинимаев и соавт., 2002), снижая эффективность преобразования углерода. Интересно, что при инактивации путей синтеза сахарозы и гликогена в клетках Mm. alcaliphilum увеличилось относительное содержание белка, но несколько снизилась скорость роста на метане. Таким образом, была показана принципиальная возможность относительно стабильного роста метанотрофа в отсутствие синтеза запасных соединений углерода.
Маркер SacB применялся при получении штаммов Mm. alcaliphilum, способных синтезировать 2,3-бутандиол, рассматриваемый в качестве биотоплива будущего (Nguyen et al., 2018), и путресцин (1,4-диаминобутана), используемый в фармацевтике и агрохимии (Nguyen, Lee, 2019), а также при конструировании пути биосинтеза пигмента астаксантина у Methylomonas sp. 16а (Ye et al., 2007) и для скрининга двойных рекомбинантов у Ms. trichosporium OB3b при получении сайт-направленных мутаций в sMMO (Borodina et al., 2007). С использованием этого метода была изучена функция пентациклических тритерпеноидных липидов у метанотрофов. Было установлено, что 3-метилгопаноидметилаза участвует в стабилизации ВЦМ у Mс. capsulatus Bath (Welander, Summons, 2012).
В качестве маркера контрселекции используется также ген pheS, кодирующий α-субъединицу фенилаланил-тРНК-синтетазы. Введением двух точечных мутаций в ген pheS из Mс. capsulatus Bath получен вариант PheSAG, который может включать в белки p-хлорфенилаланин, вызывая гибель клеток. Посредством отбора с помощью p-хлорфенилаланина был получен безмаркерный мутант xoxF с гораздо большей эффективностью, чем ко-нтрселекцией на основе SacB (Ishikawa et al., 2018). Варианты pheS использовались в метанотрофных штаммах-хозяевах, включая Mm. buryatense 5GB1C, Mm. alcaliphilum 20Z и Methylomonas sp. LW13 (Liu et al., 2020, 2021). Путем объединения pheSAG и гена устойчивости к зеоцину была сконструирована позитивно-контрселективная кассета PZ (Liu et al., 2020). Стратегия, основанная на PZ и ПЦР (рис. 2б), была успешно использована при создании немаркированной делеции glgA1 или всего оперона, кодирующего sММО, в Mm. buryatense 5GB1C и Mm. alcaliphilum 20Z.
Система генетического редактирования CRI-SPR/Cas9 (clustered regularly interspaced short palindromic repeats). В исследованиях последних лет высоко востребована система генетического редактирования, основанная на использовании эндонуклеазы Cas9, являющейся частью иммунной системы прокариот. Эта система позволяет с высокой специфичностью редактировать геномы как про-, так и эукариотических организмов. Эндонуклеаза Cas9 вносит двухцепочечный разрыв в нить ДНК, при этом таргетирование осуществляется с помощью так называемой “single guide” РНК (gRNA), содержащей протоспейсер из 20 п.н., комплементарный целевой последовательности, прилегающей к PAM (protospacer-adjacent motif) мотиву 5'-NGG-3'. Двухцепочечный разрыв может быть восстановлен с помощью специально выбранной “заплатки”, что позволяет ввести желаемые генетические модификации (Garneau et al., 2010; Gasiunas et al., 2012; Jinek et al., 2012). К настоящему времени, однако, выполнена лишь одна работа по редактированию геномов метанотрофов с помощью системы CRISPR/Cas9 (Tapscott et al., 2019). У Mc. capsulatus Bath была успешно внесена точечная мутация в ген гидроксилазного компонента sMMO на плазмиде, ген mmoX инактивирован в хромосоме введением в открытую рамку считывания стоп-кодона, что нарушило функцию sMMO. При этом стоит отметить низкую эффективность модификации (2%), что, вероятно, обуслaвливает непопулярность данного подхода в отношении метанотрофов.
Плазмиды ‒ ценный генетический инструмент, который можно применять для экспрессии нескольких копий гена и быстрого тестирования генетических конструкций. Репликоны широкого круга хозяев, которые были успешно использованы у метанотрофов, включают репликон RP4/RK2, определяющий группу несовместимости P (IncP) (Ali, Murrell, 2009), а также репликоны RSF1010 (IncQ) (Lloyd et al., 1999) и pBBR1 (Welander et al., 2012). На основе этих репликонов были сконструированы плазмиды pAWP78, pAWP79 и pAWP89, примененные для экспрессии гетерологичных белков и репортерных генов в Mс. capsulatus Bath (Ali, Murrell, 2009), Methylomonas sp. 16a, DH-1 (Sharpe et al., 2007), Ms. trichosporium OB3b (Lloyd et al., 1999), Mm. buryatense 5GB1C (Puri et al., 2015) и Mm. alcaliphilum 20Z (Pham et al., 2022). При этом векторы pAWP79 и pAWP89 получили широкое распространение и стали универсальными для различных групп метанотрофов (табл. 1; Nguyen et al., 2019, 2020a, 2020b), а pAWP78 стал основой для создания вектора pCAH01, используемого для контролируемой экспрессии генов (Henard et al., 2016). Экспрессируемый ген в pCAH01 находится под контролем тетрациклинового репрессора/оператора. Репрессия гена снимается при добавлении ангидротетрациклина – аналога тетрациклина, не обладающего антибиотической активностью (Henard et al., 2010).
Таблица 1.
Примеры применения методов редактирования геномов для метанотрофных бактерий и достигнутые результаты
| Генетические инструменты | Метанотроф | Цель манипуляций | Результат | Ссылка |
|---|---|---|---|---|
| CRISPR/Cas9 | Methylococcus capsulatus | Введение стоп-кодона в открытую рамку считывания гена mmoX | Инактивация растворимой метанмонооксигеназы (sMMO) | Tapscott et al., 2019 |
| Безмаркерный мутагенез | ||||
| pheSAG | Mc.capsulatus (Bath) | Делеция гена xoxF Инактивация La2+-зависимой метанолдегидрогеназы Xox |
Создание нового метода контрселекции для Mс. capsulatus с использованием pheSAG | Ishikawa et al., 2018 |
| Mm. buryatense 5GB1C Mm. alcaliphilum 20Z |
Делеция 10-т.п.н. smmo оперона, кодирующего sMMO. Выключение гена гликогенсинтазы Выключение гена гликогенсинтазы |
Создание нового метода контрселекции для Mm. buryatense и Mm. alcaliphilum с использованием pheSAG | Liu et al., 2020 | |
| sacB | Mc. capsulatus Bath | Делеция гена hpnR, кодирующего метилазу гопаноидов | Выяснение роли метилгопаноидов в стабилизации мембран | Welander, Summons, 2012 |
| Mm. buryatense 5GB1C | Инсерция PmxaF-xylE в хромосому | Доказательство репрессии гена mxaF редкоземельными металлами | Chu, Lidstrom, 2016 | |
| Mm. buryatense 5GB1C Methylomonas sp. LW13 Methylobacter tundripaludum 21/22 |
ΔglgA1, ΔmmoD, ΔsMMO Делеция генов гликогенсинтазы (glgA1), регулятора экспрессии рММО (mmoD) и локуса, кодирующего гена рММО (~10 т.п.н.) |
Разработка протоколов метаболической инженерии метанотрофов типа I на основе электропорации, контрселекции sacB или сайт-специфической рекомбинации FLP-FRT | Yan et al., 2016 | |
| Mm. alcaliphilum 20Z | ΔspsΔglg Делеция кластера генов синтеза и распада гликогена и сахарозы |
Получение штамма, не синтезирующего гликоген и сахарозу | Бут с соавт., 2020 | |
| ΔldhΔackΔspeE1::argDJ Выключение генов ldh лактатдегидрогеназы и ацетаткиназы ack, замена гена спермидинсинтазы spe на ацетилорнитинаминотрансферазу argD и орнитинацилтрансферазу argJ |
Создание штамма, способного к синтезу путресцина | Nguyen, Lee, 2019 | ||
| Δldh Δmdh Δack Тройной мутант по генам лактатдегидрогеназы, ацетаткиназы и малатдегидрогеназы |
Повышение уровня биосинтеза 2,3-бутандиола | Nguyen et al., 2018 | ||
| Flp-рекомбиназа | Mm. buryatense 5GB1C | Делеция генов xoxF, mxaF, mxaI с помощью ПЦР-фрагмента с зеоциновой кассетой с последующим выщеплением последней с помощью Flp-рекомбиназы | Показана преобладающая функция La3+-зависимой XoxF-МДГ по сравнению с Ca2+-зависимой MxaF-МДГ у Mm. buryatense | Chu, Lidstrom, 2016 |
| ΔmxaY | Установлена функция гистидинкиназы MxaY в качестве регулятора транскрипции генов метанолдегидрогеназы MxaFI | Chu et al., 2016 | ||
| Маркерные модификации | ||||
| pCM184 (Km) | Methylomonas sp. DH-1 | Делеция гена сукцинатдегидрогеназы sdh | Получение штамма, синтезирующего сукцинат | Nguyen et al., 2019 |
| pCM184 (Gm) | Mm. alcaliphilum 20Z | Δpdh::GmR. Делеция пируватдегидрогеназы | Получение штамма, синтезирующего лактат | Henard et al., 2016 |
| Mc.capsulatus Bath | Δpyc, Δsga, Δgcv Делеция пируваткарбоксилазы, серинглиоксилатаминотрансферазы, фермента расщепления глицина |
Новые знания о роли цикла Кальвина в метаболизме метанотрофов Х типа | Henard et al., 2021 | |
| pAWP89 | Methylomonas sp. DS(Δsdh) | Внесение генов глиоксилатного шунта из E. coli MG1655 под контролем PmxaF промотора | Увеличение продукции сукцината из метана | Nguyen et al., 2019 |
| Mm. alcaliphilum 20Z | Ptet::asbF-aroY-catA внесение на плазмиде дегидрошикиматдегидратазу (asbF) из Bacillus thuringensis, протокатезуатдекарбоксилазу (aroY) из Enterobacter cloacae и катехолдиоксигеназы (catA) из Acinetobacter baylyi |
Получение штамма, синтезирующего муконовую кислоту | Henard et al., 2019 | |
| Ptac:: glpFKDgspA Экспрессия НАД-зависимой глицеролдегидрогеназы (glpA), дигидроксиацетонкиназы (glpK), транспортера глицерина (glpF) из E. coli и гомологичной глицерол-3-фосфатдегидрогеназы (gpsA) |
Получение штамма, растущего на глицерине | Le et al., 2021 | ||
| pAWP89 | Mm. alcaliphilum 20Z | Внесение единым опероном генов триптофаназа (tna) из E. coli и флавинсодержащей монооксигеназы (fmo) из Methylophaga aminisulfidivorans, AroGfbr(D146N) и TrpEfbr(S40F) из E. coli | Синтез красителя индиго | Pham et al., 2022 |
| Внесение единым опероном генов afs, ispA и оптимизированного nDXP пути (ribB-DSAG-dxr) | Синтез α-фарнезина | |||
| Ptac::zss1-ispA-dxs-ispG-zwf-pgd Внесение на плазмиде гумуленсинатазы (zss1), фарнезилпирофосфатсинтазы (ispA), 1-дезоксиксилулозо-5-фосфатсинтазы (dxs), гидроксиметилбутенилдифосфатсинтазы (ispG), глюкозо-6-фосфатдегидрогеназы (zwf), фосфоглюконатдегидрогеназы (pgd) |
Синтез α-гумулена | Nguyen et al., 2020b | ||
| Mm. alcaliphilum 20Z XYL | Внесение на плазмиде кластера budABC из Klebsiella pneumoniae, кодирующего ацетолактатдекарбоксилазу, ацетолактатсинтазу и ацетоинредуктазу, под контролем Рtac промотора в штамм метанотрофа, растущий на ксилозе | Синтез 2,3-бутандиола | Nguyen et al., 2021 | |
| Внесение на плазмиде 2-диметил-4-деоксигадузолсинтазы, О-метилтрансферазы, АТФ-лигазы, аланин-аланин лигазы и транскетолазы из Nostoc punctiform под контролем Рtac промотера в штамм метанотрофа, растущий на ксилозе | Синтез шинорина | |||
| pAWP89 | Mm. alcaliphilum 20Z XYL | Ptac::phdphaABtesB Внесение на плазмиде генов тиолазы (phaA), ацетоацетил КоА редуктазы (phaB) из Ralstonia eutropha H16, тиоэстеразы (tesB) из E. coli и нативной пируватдегидрогеназы Mm. alcaliphilum 20Z под контролем Рtac промотора в штамм, растущий на ксилозе |
Синтез 3-гидроксибутирата | Nguyen et al., 2021 |
| Mm. alcaliphilum 20ZE4A (ΔldhΔackΔspeE1::argDJ) |
Внесение на плазмиде pAWP89 под контролем Ptac промотора гена орнитиндекарбоксилазы speF из Ms. trichosporium OB3b | Усиление синтеза путресцина | Nguyen, Lee, 2019 | |
| Mm. alcaliphilum 20ZM3 (Δldh Δmdh Δack) |
Внесение на плазмиде гена budA из Klebsiellapneumoniae и budB из Bacillus subtilis под контролем Рtac промотора | Синтез 2,3-бутандиола | Nguyen et al., 2018 | |
| Ms. trichosporium OB3b | Ptac::mcr-mmc-pepc Внесение на плазмиде малонил-КоА редуктазы (mcr) из Chloroflexus aurantiacus, метилмалонил-КоА карбоксилтрансферазы (mmc) из Propionibacterium freudenreichii и гомологичной ФЕП карбоксилазы (pepc) |
Синтез 3-гидроксипропионовой кислоты | Nguyen et al., 2020a | |
| pCAH01 | Mm. buryatense 5GB1C | Внесение гена лактатдегидрогеназы (ldh) из Lactobacillus helveticus под контролем тетрациклинового промотора | Индуцируемый синтез лактата | Henard, Guarnieri, 2016 |
| pAWP87 | M. buryatense 5GB1C | PmxaF::synthetic RBS::atoB-hbd-crt-ydiI Внесение на плазмиде генов тиолазы (atoB), 3-гидроксиацил-КоА дегидрогеназы (hdb), тиоэстеразы (ydiI) из E. coli, еноил-КоА гидратаза из Clostridium acetobutylicum |
Синтез кротоновой кислоты | Garg et al., 2018 |
| pK18mobSacB | Ms. trichosporium OB3b | mxaF::Gmr | Изучение регуляции экспрессии Xox-МДГ и Mxa-МДГ | Haque et al., 2016 |
| ПРЦ-фрагмент | Mm. buryatense 5GB1C | ΔglgA1::KmR, ΔmmoD::KmR, ΔsMMO::KmR Замена генов гликогенсинтазы (glgA1), регулятора экспрессии sММО (mmoD) или локуса, кодирующего ген рММО (~10 т.п.н.) на канамициновую кассету |
Создание метода контрселекции | Yan et al., 2016 |
| Δsga, ΔfumA, ΔfumC, Δlpd, Δakdc, Δkor, ΔlpdΔakdc Делеция серин-глиоксилатаминотрансферазы (sga), фумараз (fumA, fumС), 2-альфакетоглутаратдегидрогеназы (lpd), альфакетоглутаратдекарбоксилазы (akdc), альфакетоглутаратферридоксиноксидоредуктазы (kor) |
Изучение функции генов | Fu et al., 2017 | ||
| Methylomonas sp. LW13 | ΔxoxF::KmR, ΔxoxG::KmR, ΔmxaF::KmR, ΔxoxG::KmR Делеция структурных генов метанолдегидрогеназ XoxF и MxaF и цитохрома XoxG(4) | Выяснение функции цитохрома XoxG(4) в метаболизме метанотрофов | Zheng et al., 2018 | |
| pCM351-glgА (Gm) | Mm. alcaliphilum 20Z | Ptac:: xylABrpe Ведение генов утилизации ксилозы в glgA локус хромосомы: ксилозоизомеразы (xylA) и ксилулозокиназы (xylB), а также рибулозо-фосфат-3-эпимеразы из E. coli (rpe) под контролем Рtac промотора |
Получение штамма метанотрофа, растущего на ксилозе | Nguyen et al., 2021 |
| Mm. alcaliphilum 20Z | Замена glgA1 кластером budABC из Klebsiella pneumoniae под контролем Рtac промотера в штамм, растущий на глицерине за счет экспрессии кластера Ptac::glpFKDgspA | Синтез 2,3-бутандиола | Le et al., 2021 | |
Недавно у Methylomonas sp. DH-1 была обнаружена цитозинметилтрансфераза, которая помогает чужеродным плазмидам обходить систему рестрикции‒модификации (РM) (Ren et al., 2020), что приводит к более высокой эффективности трансформации. Цитозинметилтрансфераза распознает последовательность GGCC для метилирования. Ген, кодирующий цитозинметилтрансферазу Methylomonas sp. DH-1, был введен в E. coli JM110 с дефицитом генов метилазы dam и dcm, тем самым была создана синтетическая система РM. Кишечная палочка, экспрессирующая цитозинметилтрансферазу, метилировала чужеродные плазмиды, которые после выделения из E. coli JM110 показали повышенную эффективность трансформации в Methylomonas sp. DH-1 по сравнению с неметилированными плазмидами. С использованием этого подхода в клетках Methylomonas sp. DH-1, трансформированных плазмидами, несущими гены синтеза каротиноидов, продукция пигмента была увеличена на 26% (Ren et al., 2020).
В бактериальной биотехнологии для генетических манипуляций или физиологических модификаций возможно доставлять функциональные белки в клетки с использованием пептидов в качестве переносчиков. Библиотеку проникающих в клетку пептидов и метод доставки белков, конъюгированных c пептидами, тестировали, вырезая маркерный ген в Methylomonas sp. DH-1 (Lie et al., 2021). Предложенная библиотека проверена на отсутствие значительной цитотоксичности проникающих пептидов в отношении E. coli.
Элиминация нативной плазмиды у Mm. buryatense 5GB1. Системы экспрессии генов на основе плазмид позволяют быстро тестировать несколько генетических конструкций. Ранее для манипулирования штаммами неметанотрофных метилотрофов была разработана система с использованием плазмиды широкого круга хозяев pCM66 на основе IncP размером 7.6 т.п.н. (Marx, Lidstrom, 2001). Однако исходный штамм Mm. buryatense 5GB1 содержит плазмиду размером 80 т.п.н., несущую функции стабильности и поддержания, которые приводят к несовместимости между плазмидами. Элиминация нативной плазмиды была проведена путем введения IncP-плазмиды pVK100 и последующим избавлением от нее с помощью пересевов без антибиотика (Puru et al., 2015). Как результат, получены штаммы Mm. buryatense 5GB1S и 5GB1C, которые способны конъюгировать с малыми векторами и поддаются современным методам клонирования.
Библиотеки промоторов для экспрессии генов метанотрофов и репортерные гены. В метаболической инженерии сверхэкспрессия генов может не привести к увеличению выхода продукции, поскольку клеточные ресурсы расходуются чрезмерно на синтез белка, что приводит к снижению ростовых параметров. Кроме того, неоптимизированная экспрессия генов может способствовать накоплению токсичных промежуточных метаболитов, что снизит выход продукции. Для тонкой настройки уровней экспрессии генов используются промоторы различной силы.
Для поиска подходящих промоторов, обеспечивающих конститутивную или регулируемую экспрессию целевых генов, используются репортерные белки. Применение канонических флуоресцирующих белков (таких как GFP, dTomato) считалось неэффективным способом оценки силы промоторов у метанотрофов вследствие наличия у них обширной сети ВЦМ и/или S-слоев на поверхности клеток. Трудоемкость оценки флуоресценции в клеточных экстрактах сводит на нет преимущество в использовании репортерных генов (Ali, Murrel, 2009; Puri et al., 2015). Для определения силы промоторов у Methylomonas sp. DH-1 тандем промотор-gfp был интегрирован в некодирующую область генома, а флуоресценцию тестировали методом проточной цитометрии (Lee et al., 2021a). У Mm. buryatense 5GB1S эндогенные промоторы, управляющие экспрессией гена метанолдегидрогеназы mxaF и сигма-факторов rpoD, были протестированы с помощью красного флуоресцентного белка dTomato в качестве репортера (Shaner et al., 2004). Обнаружено, что промотор гена mxaF неактивен в E. coli, хотя это был самый сильный из протестированных промоторов в Mm. buryatense. Наряду с генами флуоресцирующих белков, с этой целью применяются гены катехол-2,3-диоксигеназы xylE и β-галактозидазы lacZ (Ali, Murrell, 2009). Силу промоторов ключевых генов у Mm. alcaliphilum 20Z тестировали в штамме метанотрофа, дефектном по глюкокиназе (Мустахимов и соавт., 2016). Ген, кодирующий глюкокиназу, вносили на плазмиде под контролем тестируемых промоторов, индикатором уровня экспрессии с промотора служила активность глюкокиназы в бесклеточных экстрактах. Это исследование подтвердило высокую активность промотора гена метанолдегидрогеназы mxaF в метанотрофных бактериях.
Оценка уровней экспрессии генов у Mm. buryatense 5GB1 недавно была проведена с помощью наборов данных RNA-seq (Wilson, 2021). Штамм культивировали в различных условиях и анализировали данные секвенирования РНК. Была идентифицирована группа генов с высокой экспрессией и выявлены соответствующие промоторные области-кандидаты. Проверку предполагаемых промоторов проводили с использованием репортера XylE.
Другой подход применяли при выборе оптимальных промоторов для производства кадаверина в Methylomonas sp. DH-1 (Lee et al., 2021). Были проанализированы участки 100 п.н. выше сайта начала транскрипции всех генов, кроме гипотетических белков. Параллельно у Methylomonas sp. DH-1 идентифицировали высоко экспрессируемые белки с помощью 2D-гель-электрофореза и MALDI-TOF. В результате была получена библиотека из 38 промоторов и проверена c использованием gfp. Основываясь на полученных данных, была проведена гетерологичная экспрессия генов cadA и cadB в Methylomonas sp. DH-1. Титр кадаверина достиг наивысшего значения 18.12 мг/л в штамме, экспрессирующем гены cadA и cadB под промотором, соответственно, рибосомного белка RpmB (L28) и белка-инициатора репликации DnaA.
Модификация метаболизма с целью получения целевого продукта. Внесением на плазмиде pCAH01 гена лактатдегидрогеназы из Lactobacillus helveticus в клетки Mm. buryatense был получен штамм, который выделял в среду лактат в концентрации 800 мг/л (Henard et al., 2016). Существенный вклад в выход целевого продукта вносит также настройка трансляции генов, т.е. подбор наилучших рибосом-связывающих последовательностей (RBS). Следует отметить, что использование определенного RBS может быть удачным в одном окружении (т.е. с определенным протомотором и геном) и неэффективным в другом. Оптимальный дизайн RBS разрабатывали с помощью программы RBS калькулятор 2.0 (Salis et al., 2009; Espah Borujeni et al., 2014), что позволило значительно увеличить выход лактата у Mm. buryatense 5GB1C (Garg et al., 2018), а также шинорина у Mm. alcaliphilum 20Z (Nguyen et al., 2021). Примечательно, что выход шинорина при использовании сконструированного метанотрофного биокатализатора был сопоставим с таковыми у промышленных продуцентов.
Протестирована способность Mm. buryatense синтезировать кротоновую кислоту (выход 50 мг/л) путем внесения генов обратного пути β-окисления из E. coli MG1655 с помощью плазмидного вектора (Garg et al., 2018). Для успешного результата при модификации метанотрофов часто используют кодон-оптимизированные гены (Pham et al., 2022b). Для модификации Mm. alcaliphilum с целью получения путресцина, в штамм c делецией генов лактатдегидрогеназы, ацетаткиназы и спермидинсинтазы на плазмиде pAWP89 под taс промотором были внесены кодон-оптимизированные гены ацетилорнитинаминотрансферазы argD и орнитинацилтрансферазы argJ из Ms. trichosporium OB3b, а также ген орнитиндекарбоксилазы speC. В результате удалось достичь концентрации путресцина 98.08 мг/л (Nguyen et al., 2019). Благодаря использованию кодон-оптимизированных генов триптофаназы tna из E. coli и флавинсодержащей монооксигеназы fmo из Methylophaga aminisulfidivorans, на основе Mm. alcaliphilum был получен продуцент пигмента индиго (3.9 мкг/л). После внесения генов ксилозоизомеразы (xylA) и ксилулокиназы (xylB) из E. coli стал возможен миксотрофный рост Mm. alcaliphilum в среде с добавлением 2 г/л ксилозы, что увеличило выход красителя до 6.3 мкг/л (Pham et al., 2022b).
Среди нейтрофильных негалофильных метанотрофов наиболее податливыми к модификациям оказались представители рода Methylomonas. Так, замена гена сукцинатдегидрогеназы (sdh) у Methylomonas sp. DH-1 на канамициновую кассету привела к накоплению в среде сукцината, а введение на плазмиде pAWP89 генов глиоксилатного шунта позволило увеличить выход сукцината до 134 мг/л (Nguyen et al., 2019). Попытки перенаправить поток углерода посредством выключения генов ацетаткитназы (ack) и фосфоацетилтрансферазы (pta) или пируват-формиатлиазы (pfl) не привели к увеличению выхода сукцината.
Метаболическое моделирование. Еще одним современным подходом, активно используемым в исследованиях молекулярных основ метано- и метилотрофии, является метаболическое моделирование. Моделирование метаболических путей позволяет прогнозировать направление потока углерода при генетической модификации, конструировать новые пути, ведущие к синтезу нужного продукта, а также предсказывать влияние окружающей среды или генетических манипуляций на поведение системы. На сегодняшний день метаболические модели разработаны для Mm. buryatense 5GB1 (de la Torre et al., 2015), Mc. capsulatus Bath (Lieven et al., 2018), Mm. alcaliphilum 20Z (Akberdin et al., 2018) и Ms. trichosporium OB3b (Naizabekov, Lee, 2020). Интересно, что модели для Methylotuvimicrobium и Methylococcus предполагают значительные различия в механизмах переноса электронов. Метаболическое моделирование необходимо для прогнозирования фенотипов in silico, что может определять стратегии биоинженерии.
Совместное выращивание на метане и полиуглеродных субстратах. Большинство метанотрофов не могут использовать полиуглеродные субстраты и имеют относительно низкую скорость роста на метане ввиду низкой растворимости СН4. Кроме того, для монооксигенирования инертной молекулы метана нужны восстановительные эквиваленты. Использование полиуглеродного соединения в качестве ко-субстрата представляется многообещающей стратегией при производстве целевых продуктов. Первая попытка демонстрации способности гаммапротеобактериального метанотрофа к использованию для роста сахаров была осуществлена введением в Мm. alcaliphilum 20Z ксилозоизомеразы (xylA) и ксилулозокиназы (xylB) из E. coli наряду со сверхэкспрессией собственной рибулозофосфат-3-эпимеразы (rpe) (Nguyen et al., 2021). Сконструированный штамм мог расти на ксилозе как на единственном источнике углерода, но с довольно продолжительной лаг-фазой. Скорость роста модифицированного штамма на метане с ксилозой была в 1.5 раза выше по сравнению со скоростью роста только на метане, а продукция шинорина (производное седогептулозо-7-фосфата) возросла в 2.3 раза. Кроме того, синтез 2,3-бутандиола, ацетоина и 3-гидроксимасляной кислоты значительно усиливался при росте в среде, содержащей метан и ксилозу.
В Mm. alcaliphilum 20Z были введены гены, ответственные за утилизацию глицерина, включая транспортер глицерина (glpF), глицеролкиназу (glpK) и эндогенную глицерол-3-фосфатдегидрогеназу (gpsA) (Lee et al., 2021). Cконструированный штамм рос на глицерине при низкой концентрации 0.1%, но эта способность была улучшена адаптивной эволюцией. Глицерин, как дополнительный к метану источник углерода, в 2 раза увеличивал скорость роста и в 3.9 раза продукцию 2,3-бутандиола. Поскольку метаболизм глицерина обеспечивает синтез дополнительного НАДН, метанотроф при миксотрофном росте может конвертировать метан в метанол без добавления формиата в качестве донора электронов. Биокатализатор для превращения метана в метанол, работающий без ингибиторов метанолдегидрогеназы (хлорида аммония, этилендиаминтетраацетатного или фосфатного буфера) можно создать на основе мутанта с дефицитом метанолдегидрогеназы (Hur et al., 2017; Patel et al., 2020). Новая концепция одностадийной биоконверсии метана в метанол в метанотрофных бактериях с использованием глицерина в качестве восстановительной силы может быть расширена для превращения других алканов в соответствующие им первичные спирты.
Применение метанотрофов в агротехнологиях. Одним из примеров в этой области исследований является сконструированный штамм Methylomonas sp. LW13, способный разлагать широко используемый гербицид бенсульфурон-метил (БСМ) (Liu et al., 2021). Для этого в Methylomonas sp. LW13 была экспрессирована гидролаза SulE из Hansschlegelia zhihuaiae S113 под промотором Ptac с помощью стратегии безмаркерного гена на основе p-Cl-Phe. Фермент деэтерифицирует БСМ до бенсульфуроновой кислоты и метанола. Сконструированный штамм полностью конвертировал 40 мг/л БСМ в бенсульфуроновую кислоту через 96 ч.
Метанотрофы в природе не синтезируют фитогормоны. Тем не менее, связь между растениями и метанотрофами подтверждена многочисленными наблюдениями (Hanson, Hanson, 1996; Bosse, Frenzel, 1997; Davamani et al., 2020). В частности, интродукция метанотрофных бактерий была успешно использована для снижения выбросов метана в атмосферу из затопляемых рисовых чеков (Davamani et al., 2020). Задача создания метанотрофных бактерий, обладающих способностью синтезировать фитогормоны для стимуляции роста растений и для снижения эмиссии метана, была реализована на примере Mm. alcaliphilum 20Z (Pham et al., 2022c). L-триптофан, незаменимая для живых организмов аминокислота, синтезируется из фосфоенолпирувата и эритрозо-4-фосфата – центральных интермедиатов гликолиза ‒ доминирующего пути распада фосфосахаров у метанотрофов с РМФ циклом ассимиляции углерода. Триптофан также является предшественником ауксина (индолил-3-уксусной кислоты, ИУК). Гетерологичной экспрессией в Mm. alcaliphilum 20Z ферментов синтеза триптофана из E. coli, устойчивых к ингибированию по принципу обратной связи, получен штамм-продуцент триптофана. Дополнительным введением генов, кодирующих ферменты синтеза ИУК, сконструирован рекомбинантный штамм, синтезирующий ауксин и способный стимулировать прорастание семян, длину побегов и длину корней пшеницы на ранней стадии роста в засоленных щелочных условиях (Pham et al., 2022b). Таким образом, предложен подход для создания метанотрофов, способных повысить продуктивность и рост растений и одновременно сократить выбросы метана, образующегося в результате сельскохозяйственной деятельности.
ВЫВОДЫ И ПЕРСПЕКТИВЫ
Последнее десятилетие продемонстрировало быстрое развитие генетических инструментов и их адаптацию для применения в работе с метанотрофными бактериями. Секвенирование геномов, многочисленные примеры успешной генной инженерии в сочетании с быстро растущим пониманием фундаментального метаболизма метанотрофов дают надежду на реализацию, по крайней мере, ряда рациональных стратегий направленной модификации этих уникальных бактерий. Это относительно новая область исследований для метанотрофных бактерий; все еще существует потребность в более глубоких исследованиях, нацеленных на устранение таких ограничений, как низкий поток углерода к целевым продуктам, эффективность окисления метана и уровни экспрессии ключевых ферментов. В данном обзоре не были представлены примеры работ по синтетической биологии – преобразование метана неметанотрофными микроорганизмами, что также может внести значительный вклад в повышение эффективности производства и выхода целевых продуктов.
Список литературы
Бут С.Ю., Дедыш С.Н., Попов В.О., Пименов Н.В., Хмеленина В.Н. Конструирование метанотрофа I типа с пониженной способностью аккумулировать гликоген и сахарозу // Прикл. биохимия и микробиология. 2020. Т. 56. С. 465–471.
But S.Yu., Dedysh S.N., Popov V.O., Pimenov N.V., Khmelenina V.N. Construction of a Type-I metanotroph with reduced capacity for glycogen and sucrose accumulation // Appl. Biochem. Microbiol. 2020. V. 56. P. 538–543.
Бут С.Ю., Егорова С.В., Хмеленина В.Н., Троценко Ю.А. Биохимические свойства и филогения гидроксипируватредуктаз метанотрофных бактерий, реализующих различные пути С1 ассимиляции // Биохимия. 2017. Т. 82. С. 1647–1656.
But S.Y., Egorova S.V., Khmelenina V.N., Trotsenko Y.A. Biochemical properties and phylogeny of hydroxypyruvate reductases from methanotrophic bacteria with different C1-assimilation pathways // Biochemistry (Moscow). 2017. V. 82. P. 1295–1303.
Гальченко В.Ф. Метанотрофные бактерии. М.: ГЕОС, 2001. 500 с.
Григорян А.Н., Горская Л. Использование природного газа для микробиологического синтеза. М.: ОНТИ Микробиопром, 1970. 90 с.
Егоров И., Купина Л., Аксюк И., Муртазаева Р. Гаприн – источник белка // Птицеводство. 1990. Т. 8. С. 25‒27.
Ешинимаев Б.Ц., Хмеленина В.Н., Сахаровский В.Г., Сузина Н.Е., Троценко Ю.А. Физиолого-биохимические и цитологические особенности галоалкалотолерантного метанотрофа при росте на метаноле // Микробиология. 2002. Т. 71. С. 690–700.
Eshinimaev B.T., Khmelenina V.N., Sakharovskii V.G., Suzina N.E., Trotsenko Y.A. Physiological, biochemical, and cytological characteristics of a haloalkalitolerant methanotroph grown on methanol // Microbiology (Moscow). 2002. V. 71. P. 512‒518.
Лалов В.В. Анализ и синтез энерготехнологических систем производства кормового белка из природного газа. Автореф. дис. … докт. биол. наук. Москва, 1991.
Мустахимов И.И., Бут С.Ю., Решетников А.С., Хмеленина В.Н., Троценко Ю.А. Использование гомо- и гетерологичных репортерных белков для оценки активности промоторов у Methylomicrobium alcaliphilum 20Z // Прикл. биохимия и микробиология. 2016. Т. 52. С. 279‒286.
Mustakhimov I.I., But S.Y., Reshetnikov A.S., Khmelenina V.N., Trotsenko Y.A. Homo- and heterologous reporter proteins for evaluation of promoter activity in Methylomicrobium alcaliphilum 20Z // Appl. Biochem. Microbiol. 2016. V. 52. P. 279‒286.
Akberdin I.R., Thompson M., Hamilton R., Desai N., Alexander D., Henard C.A., Guarnieri M.T., Kalyuzhnaya M.G. Methane utilization in Methylomicrobium alcaliphilum 20ZR: a systems approach // Sci. Rep. 2018. V. 8. Art. 2512.
Ali H., Murrell J.C. Development and validation of promoter-probe vectors for the study of methane monooxygenase gene expression in Methylococcus capsulatus Bath // Microbiology (SGM. Reading, Engl.). 2009. V. 155. P. 761‒771.
Bordel S., Crombie A.T., Muñoz R., Murrell J.C. Genome Scale Metabolic Model of the versatile methanotroph Methylocella silvestris // Microb. Cell Fact. 2020. V. 19. P. 144.
Borodina E., Nichol T., Dumont M.G., Smith T.J., Murrell J.C. Mutagenesis of the “leucine gate” to explore the basis of catalytic versatility in soluble methane monooxygenase // Appl. Environ. Microbiol. 2007. V. 73. P. 6460–6467.
Bosse U., Frenzel P. Activity and distribution of methane-oxidizing bacteria in flooded rice soil microcosms and in rice plants (Oryza sativa) // Appl. Environ. Microbiol. 1997. V. 63. P. 1199–1207.
Bothe H., Moller Jensen K., Mergel A., Larsen J., Jorgensen C., Bothe H., Jorgensen L. Heterotrophic bacteria growing in association with Methylococcus capsulatus (Bath) in a single cell protein production process // Appl. Microbiol. Biotechnol. 2002. V. 59. P. 33‒39.
But S.Y., Egorova S.V., Khmelenina V.N., Mustakhimov I.I. Malyl-CoA lyase provides glycine/glyoxylate synthesis in type I methanotrophs // FEMS Microbiol. Lett. 2020. V. 367. fnaa207.
Chaumeil P.-A., Mussig A.J., Hugenholtz P., Parks D.H. GTDB-Tk: a toolkit to classify genomes with the Genome Taxonomy Database // Bioinformatics. 2020. V. 36. P. 1925–1927.
Chistoserdova L., Lidstrom M.E. Aerobic methylotrophic prokaryotes // The Prokaryotes: Prokaryotic Physiology and Biochemistry. 2013. P. 267–285.
Chu F., Beck D.C., Lidstrom M.E. MxaY regulates the lanthanide-mediated methanol dehydrogenase switch in Methylomicrobium buryatense // PeerJ. 2016. V. 4. Art. e2435.
Chu F., Lidstrom M.E. XoxF acts as the predominant methanol dehydrogenase in the type I methanotroph Methylomicrobium buryatense // J. Bacteriol. 2016. V. 198. P. 1317–1325.
Conrad R. The global methane cycle: recent advances in understanding the microbial processes involved // Environ. Microbiol. Rep. 2009. V. 1. P. 285–292.
Crombie A., Murrell J.C. Development of a system for genetic manipulation of the facultative methanotroph Methylocella silvestris BL2 // Methods Enzymol. 2011. V. 495. P. 119‒133.
Csaki R., Bodrossy L., Klem J., Murrell J.C., Kovacs K.L. Genes involved in the copper-dependent regulation of soluble methane monooxygenase of Methylococcus capsulatus (Bath): cloning, sequencing and mutational analysis // Microbiology (SGM). 2003. V. 149. P. 1785–1795.
Davamani V., Parameswari E., Arulmani S. Mitigation of methane gas emissions in flooded paddy soil through the utilization of methanotrophs // Sci. Total Environ. 2020. V. 726. Art. 138570.
De la Torre A., Metivier A., Chu F., Laurens L.M., Beck D.A., Pienkos P.T., Lidsrom M.E., Kalyuzhnaya M.G. Genome-scale metabolic reconstructions and theoretical investigation of methane conversion in Methylomicrobium buryatense strain 5G(B1) // Microb. Cell Factories. 2015. V. 14. P. 188.
Dedysh S.N., Dunfield P.F. Facultative methane oxidizers // Handbook of Hydrocarbon and Lipid Microbiology / Ed. Timmis K.N. Berlin: Springer-Verlag, 2010. P. 1967‒1976.
Dedysh S.N., Knief C. Diversity and phylogeny of described aerobic methanotrophs // Methane Biocatalysis: Paving the Way to Sustainability. 2018. P. 17–42.
Dunfield P.F., Dedysh S.N. Methylocella: a gourmand among methanotrophs // Trends Microbiol. 2014. V. 22. P. 368‒369.
Eccleston M., Kelly D.P. Assimilation and toxicity of some exogenous C1 compounds, alcohols, sugars and acetate in the methane-oxidizing bacterium Methylococcus capsulatus // J. Gen. Microbiol. 1973. V. 75. P. 211‒221.
Espah Borujeni A., Channarasappa A.S., Salis H.M. Translation rate is controlled by coupled trade-offs between site accessibility, selective RNA unfolding and sliding at upstream standby sites // Nucleic Acids Res. 2014. V. 42. P. 2646‒2659.
Fu Y., He L., Reeve J., Beck D.A.C., Lidstrom M.E. Core metabolism shifts during growth on methanol versus methane in the methanotroph Methylomicrobium buryatense 5GB1 // MBio. 2019. V. 10. P. e00406–19.
Fu Y., Li Y., Lidstrom M. The oxidative TCA cycle operates during methanotrophic growth of the Type I methanotroph Methylomicrobium buryatense 5GB1 // Metab. Eng. 2017. V. 42. P. 43–51.
Garg S., Wu H., Clomburg J.M., Bennett G.N. Bioconversion of methane to C-4 carboxylic acids using carbon flux through acetyl-CoA in engineered Methylomicrobium buryatense 5GB1C // Metab. Eng. 2018. P. 48. P. 175‒183.
Garneau J.E., Dupuis M.-È., Villion M., Romero D.A., Barrangou R., Boyaval P., Fremaux C., Horvath P., Magadán A.H., Moineau S. The CRISPR/Cas bacterial immune system cleaves bacteriophage and plasmid DNA // Nature 2010. V. 468. P. 67‒71.
Gasiunas G., Barrangou R., Horvath P., Siksnys V. Cas9-crRNA ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria // Proc. Natl. Acad. Sci. USA. 2012. V. 109. P. E2579–E2586.
Haque M.F.U., Gu W., DiSpirito A.A., Semrau J.D. Marker exchange mutagenesis of mxaF, encoding the large subunit of the Mxa methanol dehydrogenase, in Methylosinus trichosporium OB3b // Appl. Environ. Microbiol. 2016. V. 82. P. 1549–1555.
Hamer G., Harrison D.E.F. Single cell protein: the technology, economics and future potential // Hydrocarbons in Biotechnology / Eds. Harrison D.E.F., Higgins I.J., London W.R. London: Heyden Institute of Petroleum, 1980. P. 59–73.
Hanson R.S., Hanson T.E. Methanotrophic bacteria // Microbiol. Rev. 1996. V. 60. P. 439‒471.
Harwood J.H., Williams E., Bainbridge B.W. Mutation of the methane oxidizing bacterium Methylococcus capsulatus // J. Appl. Bacteriol. 1972. V. 35. P. 99–108.
Henard C.A., Franklin T.G., Youhenna B., But S., Alexander D., Kalyuzhnaya M.G., Guarnieri M.T. Biogas biocatalysis: methanotrophic bacterial cultivation, metabolite profiling, and bioconversion to lactic acid // Front. Microbiol. 2018. V. 9. Art. 2610.
Henard C.A., Smith H., Dowe N., Kalyuzhnaya M.G., Pienkos P.T., Guarnieri M.T. Bioconversion of methane to lactate by an obligate methanotrophic bacterium // Sci. Rep. 2016. V. 6. Art. 21585.
Henard C.A., Wu C., Xiong W., Henard J.M., Davidheiser-Kroll B., Orata F.D., Guarnieri M.T. Ribulose-1,5-bisphosphate carboxylase/oxygenase (RubisCO) is essential for growth of themethanotroph Methylococcus capsulatus strain Bath // Appl. Environ. Microbiol. 2021. V. 87. P. e00881-21.
Henard C.A., Bourret T.J., Song M., Vázquez-Torres A. Control of redox balance by the stringent response regulatory protein promotes antioxidant defenses of Salmonella // J. Biol. Chem. 2010. V. 285. P. 36785–36793.
Henard C.A., Akberdin I.R., Kalyuzhnaya M.G., Guarnieri M.T. Muconic acid production from methane using rationally-engineered methanotrophic biocatalysts // Green Chem. 2019. V. 21. P. 6731–6737.
Hu L., Guo S., Yan X., Zhang T., Xiang J., Fei Q. Exploration of an efficient electroporation system for heterologous gene expression in the genome of methanotroph // Front. Microbiol. 2021. V. 12. Art. 717033.
Ishikawa M., Yokoe S., Kato S., Hori K. Efficient counterselection for Methylococcus capsulatus (Bath) by using a mutated pheS gene // Appl. Environ. Microbiol. 2018. V. 84. P. e01875-18.
Jiang D., Kim C.S., Hanson R.S., Wood T.K. Optimization of trichloroethylene degradation using soluble methane monooxygenase of Methylosinus trichosporium OB3b expressed in recombinant bacteria // Biotechnol. Bioeng. 1996. V. 51. P. 349–359.
Jiang H., Chen Y., Jiang P., Zhang C., Smith T.J., Murrell J.C., Xing X.H. Methanotrophs: multifunctional bacteria with promising applications in environmental bioengineering // Biochem. Eng J. 2010. V. 49. P. 277‒288.
Jinek M., Chylinski K., Fonfara I., Hauer M., Doudna J.A., Charpentier E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity // Science 2012. V. 337. P. 816–821.
Kalyuzhnaya M.G., Kumaresan D., Heimann K., Caetano N.S., Visvanathan C., Parthiba Karthikeyan O. Editorial: Methane: a bioresource for fuel and biomolecules // Front. Environ. Sci. 2020. V. 8. Art. 9.
Kalyuzhnaya M.G., Puri A.W., Lidstrom M.E. Metabolic engineering in methanotrophic bacteria // Metab. Engin. 2015. V. 29. P. 142‒152.
Kalyuzhnaya M.G., Yang S., Rozova O.N., Smalley N.E., Clubb J., Lamb A., Gowda G.N., Raftery D., Fu Y., Bringel F. Highly efficient methane biocatalysis revealed in a methanotrophic bacterium // Nat. Commun. 2013. V. 4. P. 2785.
Khadem A.F., Pol A., Wieczorek A., Mohammadi S.S., Francoijs K.J., Stunnenberg H.G., Jetten M.S.M., Op den Camp H.J.M. Autotrophic methanotrophy in verrucomicrobia: Methylacidiphilum fumariolicum SolV uses the Calvin‒Benson‒Bassham cycle for carbon dioxide fixation // J. Bacteriol. 2011. V. 193. P. 4438–4446.
Khider M.L.K., Brautaset T., Irla M. Methane monooxygenases: central enzymes in methanotrophy with promising biotechnological applications // World J. Microbiol. Biotechnol. 2021. V. 37. P. 72.
Khmelenina V.N., But S.Y., Rozova O.N., Trotsenko Y.A. Metabolic features of aerobic methanotrophs: news and views // Curr. Issues Mol. Biol. 2019. V. 33. P. 85‒100.
Khmelenina V.N., Kalyuzhnaya M.G., Sakharovsky V.G., Suzina N.E., Trotsenko Y.A., Gottschalk G. Osmoadaptation in halophilic and alkaliphilic methanotrophs // Arch. Microbiol. 1999. V. 172. P. 321‒329.
Knief C. Diversity and habitat preferences of cultivated and uncultivated aerobic methanotrophic bacteria evaluated based on pmoA as molecular marker // Front. Microbiol. 2015. V. 6. P. 1346.
Le H.T.Q., Nguyen A.D., Park Y.R., Lee E.Y. Sustainable biosynthesis of chemicals from methane and glycerol via reconstruction of multi-carbon utilizing pathway in obligate methanotrophic bacteria // Microb. Biotechnol. 2021. V. 14. P. 2552‒2565.
Lee H.M., Ren J., Tran K.M., Jeon B.M., Park W.U., Kim H., Lee K.E., Oh Y., Choi M., Kim D.S., Na D. Identification of efficient prokaryotic cell-penetrating peptides with applications in bacterial biotechnology // Commun. Biol. 2021a. V. 4. P. 205.
Lee H.M., Ren J., Yu M.S., Kim H., Kim W.Y., Shen J., Yoo S.M., Eyun S.I., Na D. Construction of a tunable promoter library to optimize gene expression in Methylomonas sp. DH-1, a methanotroph, and its application to cadaverine production // Biotechnol. Biofuels. 2021b. V. 14. P. 228.
Lee J.K., Kim S., Kim W., Kim S., Cha S., Moon H., Hur D.H., Kim S.Y., Na J.G., Lee J.W., Lee E.Y., Hahn J.S. Efficient production of d-lactate from methane in a lactate-tolerant strain of Methylomonas sp. DH-1 generated by adaptive laboratory evolution // Biotechnol. Biofuels. 2019. V. 12. P. 234.
Lieven C., Petersen L.A.H., Jørgensen S.B., Gernaey K.V., Herrgard M.J., Sonnenschein N. A genome-scale metabolic model for Methylococcus capsulatus (Bath) suggests reduced efficiency electron transfer to the particulate methane monooxygenase // Front. Microbiol. 2018. V. 9. Art. 2947.
Liu Y., He X., Zhu P., Cheng M., Hong Q., Yan X. pheSAG based rapid and efficient markerless mutagenesis in Methylotuvimicrobium // Front. Microbiol. 2020. V. 11. Art. 441.
Liu Y., Zhang H., He X., Liu J. Genetically engineered methanotroph as a platform for bioaugmentation of chemical pesticide contaminated soil // ACS Synth. Biol. 2021. V. 10. P. 487–494.
Lloyd J.S., Finch R., Dalton H., Murrell J.C. Homologous expression of soluble methane monooxygenase genes in Methylosinus trichosporium OB3b // Microbiology (SGM. Reading). 1999. V. 145. P. 461‒470.
Marx C.J., Lidstrom M.E. Broad-host-range cre-lox system for antibiotic marker recycling in gram-negative bacteria // Biotechniques. 2002. V. 33. P. 1062‒1067.
Marx C.J., Lidstrom M.E. Development of improved versatile broad-host-range vectors for use in methylotrophs and other Gram-negative bacteria // Microbiology. 2001. V. 147. P. 2065‒2075.
Marx C.J. Development of a broad-host-range sacB-based vector for unmarked allelic exchange // BMC Res. Notes. 2008. V. 1. Art. 1. https://doi.org/10.1186/1756-0500-1-1
Murrell J.C., Gilbert B., McDonald I.R. Molecular biology and regulation of methane monooxygenase // Arch. Microbiol. 2000. V. 173. P. 325–332.
Murrell J.C., Smith T.J. Biochemistry and Molecular Biology of Methane Monooxygenase. Handbook of Hydrocarbon and Lipid Microbiology. Berlin‒Heidelberg: Springer, 2010. P. 1045–1055.
Mustakhimov I.I., Reshetnikov A.S., Glukhov A.S., Khmelenina V.N., Kalyuzhnaya M.G., Trotsenko Y.A. Identification and characterization of EctR1, a new transcriptional regulator of the ectoine biosynthesis genes in the halotolerant methanotroph Methylomicrobium alcaliphilum 20Z // J. Bacteriol. 2010. V. 192. P. 410–417.
Naizabekov S., Lee E.Y. Genome-scale metabolic model reconstruction and in silico investigations of methane metabolism in Methylosinus trichosporium OB3b // Microorganisms. 2020. V. 8. P. 437.
Nguyen A.D., Chau T.H.T., Lee E.Y. Methanotrophic microbial cell factory platform for simultaneous conversion of methane and xylose to value-added chemicals // Chem. Eng. J. 2021. V. 420. P. 127632.
Nguyen A.D., Hwang I.Y., Lee O.K., Kim D., Kalyuzhnaya M.G., Mariyana R., Hadiyati S., Kim M.S., Lee E.Y. Systematic metabolic engineering of Methylomicrobium alcaliphilum 20Z for 2,3-butanediol production from methane // Metab. Engin. 2018. V. 47. P. 323‒333.
Nguyen D., Lee O.K., Lim C., Lee J., Na J.-G., Lee E.Y. Metabolic engineering of type II methanotroph, Methylosinus trichosporium OB3b, for production of 3-hydroxypropionic acid from methane via a malonyl-CoA reductase-dependent pathway // Metab. Engin. 2020a. V. 59. P. 142‒150.
Nguyen D.T.N., Lee O.K., Hadiyati S., Affifah A.N., Kim M.S., Lee E.Y. Metabolic engineering of the type I methanotroph Methylomonas sp. DH-1 for production of succinate from methane // Metab. Engin. 2019. V. 54. P. 170‒179.
Nguyen L.T., Lee E.Y. Biological conversion of methane to putrescine using genome-scale model-guided metabolic engineering of a methanotrophic bacterium Methylomicrobium alcaliphilum 20Z // Biotechnol. Biofuels. 2019. V. 12. P. 147.
Nguyen A.D., Kim D., Lee E.Y. Unlocking the biosynthesis of sesquiterpenoids from methane via the methylerythritol phosphate pathway in methanotrophic bacteria, using α‑humulene as a model compound // Metab. Engin. 2020b. V. 61. P. 69–78.
Op den Camp H.J.M., Islam T., Stott M.B., Harhangi H.R., Hynes, A., Schouten S., Jetten M.S.M., Birkeland N.K., Pol A., Dunfield P.F. Environmental, genomic and taxonomic perspectives on methanotrophic Verrucomicrobia // Environ. Microbiol. Rep. 2009. V. 1. P. 293–306.
Øverland M., Tauson A.H., Shearer K., Skrede A. Evaluation of methane-utilising bacteria products as feed ingredients for monogastric animals // Arch. Anim. Nutr. 2010. V. 64. P. 171‒189.
Parks D.H., Chuvochina M., Rinke C., Mussig A.J., Chaumeil P.-A., Hugenholtz P. GTDB: an ongoing census of bacterial and archaeal diversity through a phylogenetically consistent, rank normalized and complete genome-based taxonomy // Nucleic Acids Res. 2022. V. 50. P. D785–D794.
Pham D.N., Nguyen A.D., Lee E.Y. Outlook on engineering methylotrophs for one-carbon-based industrial biotechnology // Chem. Engin. J. 2022a. V. 449. P. 137769.
Pham D.N., Nguyen A.D., Oh S.H., Lee E.Y. Bypassing the bottlenecks in the shikimate and methylerythritol phosphate pathways for enhancing the production of natural products from methane in Methylotuvimicrobium alcaliphilum 20Z // Green Chem. 2022b. V. 24. P. 2893‒2903.
Pham D.N., Mai D.H.A., Nguyen A.D., Chau T.H.T., Lee E.Y. Development of an engineered methanotroph-based microbial platform for biocatalytic conversion of methane to phytohormone for sustainable agriculture // Chem. Engin. J. 2022c. V. 429. Art. 132522.
Pieja A.J., Morse M.C., Cal A.J. Methane to bioproducts: the future of the bioeconomy? // Curr. Opin. Chem. Biol. 2017. V. 41. P. 123–131.
Puri A.W., Owen S., Chu F., Chavkin T., Beck D.A.C., Kalyuzhnaya M.G. Genetic tools for the industrially promising methanotroph Methylomicrobium buryatense // Appl. Environ. Microbiol. 2015. V. 81. P. 1775–1781.
Recorbet G., Robert C., Givaudan A., Kudla B., Normand P., Faurie G. Conditional suicide system of Escherichia coli released into soil that uses the Bacillus subtilis sacB gene // Appl. Environ. Microbiol. 1993. V. 59. P. 1356‒1365.
Ren J., Lee H.‑M., Thai T.D., Na D. Identification of a cytosine methyltransferase that improves transformation efficiency in Methylomonas sp. DH‑1 // Biotechnol. Biofuels. 2020. V. 13. P. 200.
Ro S.Y., Rosenzweig A.C. Recent advances in the genetic manipulation of Methylosinus trichosporium OB3b // Methods Enzymol. 2018. V. 605. P. 335‒349.
Salis H.M., Mirsky E.A., Voigt C.A. Automated design of synthetic ribosome binding sites to control protein expression // Nat. Biotechnol. 2009. V. 27. P. 946‒950.
Schmitz R.A., Peeters S.H., Versantvoort W., Picone N., Pol A., Jetten M.S.M., Op Den Camp H.J.M. Verrucomicrobial methanotrophs: ecophysiology of metabolically versatile acidophiles // FEMS Microbiol. Rev. 2021. V. 45. fuab007.
Shaner N.C., Campbell R.E., Steinbach P.A., Giepmans B.N.G., Palmer A.E., Tsien R.Y. Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein // Nat. Biotechnol. 2004. V. 22. P. 1567‒1572.
Stafford G.P., Scanlan J., McDonald I.R., Murrell J.C. rpoN, mmoR and mmoG, genes involved in regulating the expression of soluble methane monooxygenase in Methylosinus trichosporium OB3b // Microbiology (SGM). 2003. V. 149. P. 1771–1784.
Sternberg N., Hamilton D. Bacteriophage P1 site-specific recombination: I. Recombination between loxP sites // J. Mol. Biol. 1981. V. 150. P. 467‒486.
Strong P.J., Xie S., Clarke W.P. Methane as a resource: can the methanotrophs add value? // Environ Sci. Technol. 2015. V. 49. P. 4001–4018.
Tapscott T., Guarnieri M.T., Henard C.A. Development of a CRISPR/Cas9 system for Methylococcus capsulatus in vivo gene editing // Appl. Environ. Microbiol. 2019. V. 85. e00340-19.
Trotsenko Y.A., Murrell J.C. Metabolic aspects of aerobic obligatemethanotrophy // Adv. Appl. Microbiol. 2008. V. 63. P. 183–229.
Welander P.V., Summons R.E. Discovery, taxonomic distribution, and phenotypic characterization of a gene required for 3-methylhopanoid production // Proc. Natl. Acad. Sci. USA. 2012. V. 109. P. 12905–12910.
Williams E., Shimmin M.S., Bainbridge B.W. Mutation in the obligate methylotrophs Methylococcus capsulatus and Methylomonas albus // FEMS Microbiol. Lett. 1977. V. 2. P. 293–296.
Wilson E.H., Groom J.D., Sarfatis M.C., Ford S.M., Lidstrom M.E., Beck D.A., A computational framework for identifying promoter sequences in nonmodel organisms using RNA-seq data sets // ACS Synth. Biol. 2021. V. 10. P. 1394–1405.
Yan X., Chu F., Puri A.W., Fu Y., Lidstrom M.E. Electroporation-based genetic manipulation in type I methanotrophs // Appl. Environ. Microbiol. 2016. V. 82. P. 2062–2069.
Ye R.W., Yao H., Stead K., Wang T., Tao L., Cheng Q., Sharpe P.L., Suh W., Nagel E., Arcilla D., Dragotta D., Miller E.S. Construction of the astaxanthin biosynthetic pathway in a methanotrophic bacterium Methylomonas sp. strain 16a // Ind. Microbiol. Biotechnol. 2007. V. 34. P. 289‒299.
Zheng Y., Huang J., Zhao F., Chistoserdova L. Physiological effect of XoxG(4) on lanthanide-dependent methanotrophy // mBio. 2018. V. 9. e02430-17.
Дополнительные материалы отсутствуют.
Инструменты
Микробиология