

Микология и фитопатология, 2019, T. 53, № 2, стр. 101-107
Embellisia-подобные гифомицеты из России и Казахстана в коллекции чистых культур ВИЗР
Ф. Б. Ганнибал *, Е. Л. Гасич
Всероссийский НИИ защиты растений
196608 Санкт-Петербург, Россия
* E-mail: fgannibal@vizr.spb.ru
Поступила в редакцию 20.12.2017
После доработки 06.02.2018
Принята к публикации 29.05.2018
Аннотация
Род Embellisia – полифилетическая группа, включавшая в себя 24 вида и претерпевшая в последние годы кардинальные таксономические преобразования. Большинство видов Embellisia было перемещено в род Alternaria. Согласно данным литературы из числа Embellisia-подобных грибов в России были обнаружены E. chlamydospora (= Alternaria chlamydosporigena, Pleosporaceae) и E. helianthi (= Alternariaster helianthi, Leptosphaeriaceae). В коллекции живых культур грибов ВИЗР присутствовало 9 изолятов, по предварительным данным имеющих сходство с Embellisia. Целью настоящей работы было провести точную идентификацию данных изолятов, опираясь на морфологические признаки и молекулярные маркеры – последовательности генов Alt a1, cald и gpd. В результате три изолята были идентифицированы как Alternaria chlamydosporigena (мак, Республика Крым; пшеница, Краснодарский край; цикорий, Алматинская обл. Казахстана), два – как A. tellustris (арбуз, Астраханская обл. и мак, Республика Крым), один – как A. embellisia (лук, Псковская обл.), один – как Drechslera biseptata (ситник, Камчатский край), два – как Alternariaster helianthi (подсолнечник, Белгородская обл. и Приморский край). Находки Alternaria embellisia и A. tellustris являются первыми для России. Находки A. chlamydosporigena и Drechslera biseptata расширяют представление о распространении этих видов и их субстратной приуроченности.
Род Embellisia E. G. Simmons был описан для того, чтобы выделить в отдельный таксон атипичный вид Helminthosporium, H. allii Campanile (Simmons, 1971). Пороконидии Embellisia в большинстве случаев представляют собой фрагмоконидии и лишь иногда формируют продольные перегородки, становясь диктиоконидиями. Основное отличие видов, относимых к этому роду от других близких грибов заключается в том, что поперечные перегородки конидий толстые и темнее, чем наружная стенка конидий. Конидии Embellisia эллиптические, вытянутые или почти цилиндрические, иногда немного искривленные; на мицелии нередко формируются хламидоспоры (Simmons, 1971, 1983).
Всего было описано под данным родовым названием или перенесено в этот род из других родов 24 вида (Simmons, 1971, 1983, 1990; de Hoog, Muller, 1973; Muntanjola-Cvetković, Ristanović, 1976; de Hoog et al., 1985; David et al., 2000; Li, Nan, 2007). Род Embellisia морфологически несколько сходен с Alternaria, Drechslera, Ulocladium и Curvularia (Shoemaker, 1959, 1962; Simmons, 1967, 1992; Alcorn, 1988, 1991). Сходство видов Embellisia с представителями других родов и друг с другом, дополняемое широким диапазоном варьирования морфологических признаков внутри одного вида и штамма, затрудняет таксономические исследования и видовую идентификацию, проводимые классическими методами.
Исследования с применением молекулярных методов выявили полифилетичность рода (Pryor, Bigelow, 2003; Lawrence et al., 2012, 2013; Woudenberg et al., 2013). Виды Embellisia оказались в семи кладах. В одной кладе, содержащей типовой вид E. allii, оказалось еще два вида. По 1–2 вида оказались в кладах, содержащих типовые виды Alternaria, Chalastospora, Ulocladium и Stemphylium. Два сестринских кластера, содержащие 8 видов Embellisia и 4 вида Alternaria, не имели связи с типами каких-либо родов. Такая полифилетичность рода Embellisia и некоторых других родственных групп грибов привела к объединению всех альтернариоидных гифомицетов под одним родовым названием Alternaria (Woudenberg et al., 2013). Отдельным кладам был присвоен статус секций. При этом большинство видов Embellisia были перемещены в род Alternaria, но оказались в нескольких секциях. В том числе в Alternaria отнесли и типовой вид E. allii (= A. embellisia Woudenberg et Crous), что автоматически приводит к упразднению рода Embellisia. Название одного вида Embellisia было признано незаконным. Таксономический статус четырех видов пока не определен.
В российской микологической литературе упоминания Embellisia почти отсутствуют. Нами ранее сообщалось об обнаружении E. chlamydospora (Hoes, G. W. Bruehl et C. G. Shaw) E. G. Simmons (Gannibal, 2004). Иногда упоминался патоген подсолнечника E. helianthi (Hansf.) Pidopl. (Kozyreva, Primakovskaya, 1981; Tikhonova, Alifirova, 1988). Этот вид в разное время относили к родам Helminthosporium и Alternaria. Комбинация Embellisia helianthi была сделана Н. М. Пидопличко (Pidoplichko, 1977), но не была эффективно опубликована. Она отсутствует в таксономических базах данных MycoBank и Index Fungorum, хотя появлялась иногда в литературе, преимущественно русскоязычной. Недавно этот вид поместили в род Alternariaster (Simmons, 2007). Молекулярно-филогенетические исследования подтверждают справедливость такого таксономического решения. Более того, A. helianthi был отторгнут от семейства Pleosporaceae, куда относятся остальные виды, сходные с Embellisia, и перемещен в Leptosphaeriaceae (Alves et al., 2013).
В коллекции живых культур грибов Всероссийского научно-исследовательского института защиты растений (ВИЗР) присутствует 9 изолятов, по предварительным данным имеющих сходство с Embellisia. Целью настоящей работы было провести точную идентификацию данных изолятов, опираясь на морфологические и молекулярные признаки и используя современную систематику и номенклатуру.
МАТЕРИАЛЫ И МЕТОДЫ
Материалом исследования являлись все изоляты из коллекции чистых культур лаборатории микологии и фитопатологии ВИЗР, которые по результатам проведенного ранее изучения морфологии, были отнесены к роду Embellisia. Семь изолятов были предварительно определены как E. chlamydospora и Embellisia sp. Они были выделены в разные годы разными коллекторами в разных регионах России и одной из областей Казахстана (табл. 1). Еще два изолята представляли вид E. helianthi (теперь Alternariaster helianthi).
Таблица 1.
Происхождение использованных в работе изолятов Embellisia-подобных гифомицетов из России и Казахстана
| Изолят | Вид | Субстрат | Происхождение | Год |
|---|---|---|---|---|
| MF-P041-011 | Alternaria chlamydosporigena | Cichorium intibus, лист арбуза | Алматинская обл. 43°18′, 76°86′ | 2006 |
| MF-P404-011 | “ “ | Papaver sp., корень мака | Республика Крым 44°96′, 34°11′ | 2007 |
| MF-P427-011 | “ “ | Triticumaestivum, корень пшеницы | Краснодарский край 45°, 39° | 2003 |
| MF-P319-011 | A. embellisia | Alliumsativum, лист чеснока | Псковская обл. 56°33′, 30°49′ | 2010 |
| MF-P403-011 | A. tellustris | Papaver sp., корень мака | Республика Крым 44°96′, 34°11′ | 2007 |
| MF-P485-011 | “ “ | Citrullus lanatus, корень арбуза | Астраханская обл. 47°, 47° | 2017 |
| MF-P183-010 | Pyrenophora biseptata | Juncusbufonius, лист ситника | Камчатский край 53°14′, 158°46′ | 2010 |
| MF-P049-011 | Alternariaster helianthi | Helianthusannuus, лист подсолнечника | Приморский край 44°74′, 132°03′ | 2006 |
| MF-P135-011 | “ “ | Helianthusannuus, лист подсолнечника | Белгородская обл. 50°16′, 38°42′ | 2007 |
Для анализа микроскопических признаков изоляты культивировали на картофельно-морковном агаре (КМА, отвар из 20 г картофеля и 20 г моркови, 15 г агар-агара, вода до 1 л) при температуре 24°С и облучении светом люминесцентных ламп дневного света. Наблюдение морфологии и измерение конидий проводили при помощи микроскопа AxioImager M1 c камерой AxioCam MRm и программы Axio Vision (Carl Zeiss, Германия).
Для экстракции ДНК изоляты выращивали на среде Чапека в течение 7–10 сут. при 23 ± 1°С. Отбирали 50–150 мг воздушного мицелия, помещали в 1.5 мл микропробирку и замораживали при –80°С. Экстракцию проводили по стандартному “ЦТАБ-хлороформ” протоколу (Doyle, Doyle, 1987). Амплификация генов основного аллергена Alternaria alternata (Alt a 1), глицеральдегид-3-фосфат дегидрогеназы (gpd) и кальмодулина (cald) была проведена с использованием праймеров Alt-for/Alt-rev, gpd1/gpd2 и CaldF1/CaldR1 (Hong et al., 2005; Berbee et al., 1999; Lawrence et al., 2013, соответственно). Для идентификации предполагаемых штаммов Alternariaster helianthi был секвенирован только ген gpd. Определение нуклеотидных последовательностей было осуществлено на автоматическом капиллярном генетическом анализаторе ABI Prism 3500 (ABI-Hitachi, Япония) с использованием набора реактивов с флуоресцентно меченными дезоксинуклеозидтрифосфатами BigDye Terminator v3.1 Cycle Sequencing Kit (ABI, США). Полученные последовательности были проверены и отредактированы в программе ContigExpress пакета Vector NTI Suite 8.0 и затем депонированы в базе данных GenBank (https://www.ncbi.nlm.nih.gov).
Полученные нуклеотидные последовательности при помощи программного модуля BLASTn (Altschul et al., 1990) сравнили с базой данных GenBank. После выявления видов, находящихся по отношению к изучаемым изолятам в наибольшем родстве, были построены филогенетические деревья. Для сравнения видов по степени сходства нуклеотидных последовательностей из базы данных GenBank были взяты последовательности референсных штаммов четырех видов бывшего рода Embellisia (табл. 2). В качестве внешней группы был использован вид Alternaria papavericola (Brachycladium papaveris). Нуклеотидные последовательности выравнивали с помощью программы ClustalX 1.8 (Thompson et al., 1997). Дендрограммы сходств были построены методом наибольшего правдоподобия (maximum likelihood – ML) с использованием программного обеспечения RAxML (v. 7.2.8, Stamatakis et al., 2006). Надежность топологии дендрограммы была оценена с помощью бутстреп-анализа c 500 повторностей.
Таблица 2.
Учетные номера GenBank для последовательностей трех генов эталонных штаммов, использованных для идентификации видов и реконструкции молекулярной филогении
| Вид | Штамм | Учетные номера GenBank | ||
|---|---|---|---|---|
| Alt a1 | cald | gpd | ||
| Alternaria didymospora (Embellisia didymospora) | CBS 766.79 | JN383506 | JQ646130 | JN383470 |
| A. tellustris (E. telluster) | EGS 33-026 | AY563325 | JQ646128 | JN383475 |
| A. chlamydosporigena (E. chlamydospora) | EGS 33-022 | JN383504 | JQ646129 | JN383468 |
| A. embellisia (E. allii) | EGS 38-073 | AY563322 | JQ646127 | AY278827 |
| A. papavericola | P351 | JN383501 | JQ646121 | FJ357298 |
РЕЗУЛЬТАТЫ
Были установлены 23 нуклеотидные последовательности генов Alt a 1 (около 450 п.н.), cald (около 750 п.н.) и gpd (около 600 п.н.), которые были депонированы в GenBank. Сравнение полученных нуклеотидных последовательностей с уже имеющимися в базе данных выявило наибольшее сходство трех изолятов с Alternaria chlamydosporigena Woudenberg et Crous [E. chlamydospora (Hoes, G.W. Bruehl et C.G. Shaw) E.G. Simmons], двух – с A. tellustris (E.G. Simmons) Woudenberg et Crous (= E. telluster E.G. Simmons) и одного – с A. embellisia Woudenberg et Crous [= E. allii (Campan.) E.G. Simmons].
Для последовательности гена gpd изолята MF-P183-010 ближайшим гомологом оказался сиквенс Pyrenophora biseptata (Sacc. et Roum.) Crous [= Drechslera biseptata (Sacc. et Roum.) M.J. Richardson et E.M. Fraser]. Гомологичные сиквенсы генов Alt a 1 и cald для этого штамма найдены не были, вероятно, из-за того, что эти гены у Pyrenophora biseptata ранее не секвенировали.
Поиск гомологичных последовательностей гена gpd изолятов Alternariaster helianthi (Hansf.) E.G. Simmons выявил таковые, сходные на 100 или почти 100% с сиквенсами депонированными в GenBank под тем же видовым названием. Только последовательность изолята MF-P135-011 отличалась от репрезентативного штамма (CBS 119672), имея одну нуклеотидную замену на 592 п.н.
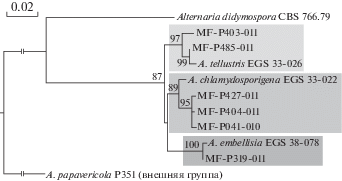
Таким образом, шесть изолятов были отнесены к трем видам секции Embellisia рода Alternaria, два – к роду Alternariaster, один – к Pyrenophora. Для видов Alternaria была построена дендрограмма (рис. 1), на которой видна степень сходства исследуемых изолятов, с типовыми штаммами соответствующих видов.
Рис. 1.
Дендрограмма сходств нуклеотидных последовательностей генов Alt a1, gpd и cald видов рода Alternaria секции Embellisia, построенная методом наибольшего правдоподобия (maximum likelihood). Цифры над узлами ветвления означают уровень бутстреп-поддержки (%).
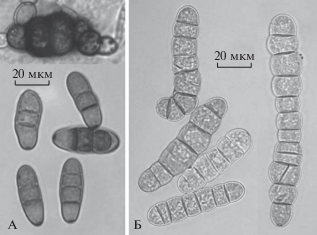
Ревизия морфологии изолятов подтвердила правильность идентификации, сделанной в ходе анализа, выполненного посредством BLAST. Конидии у всех изолятов были вытянуто-овальными, иногда слегка искривленными (ассиметричными), коричневыми, с толстыми темными поперечными перегородками. Морфологические признаки изолята MF-P319-011, выделенного из листьев чеснока, не противоречили описанию Alternaria embellisia (Simmons, 1971). Конидии 25–44 × 10–13(15) мкм с 3–5(6) поперечными перегородками и иногда одной продольной (рис. 2А). Конидии располагались по 3–8 или более на коленчатых или извилистых нередко разветвленных конидиеносцах. Изолят формировал более или менее округлые хламидоспоры.
Рис. 2.
Морфология видов Alternaria из секции Embellisia: А – A. embellisia (конидиеносцы и конидии); Б – A. tellustris (конидии, конидии на конидиеносце и хламидоспоры); В – A. chlamydosporigena (конидиеносцы, конидии и хламидоспоры).

Три изолята имели форму и размер конидий и хламидоспор (рис. 2В), соответствующие описанию A. chlamydosporigena (Simmons, 1971). Конидии 18–34 × 7–9 мкм с (2)3–5(6) поперечными толстыми темными перегородками и иногда одной продольной. Конидии располагались по 1–3 на конидиеносце, в исключительных случаях – по 4–5. Наблюдались единичные случаи формирования цепочек из двух спор. Все изоляты формировали интеркалярные хламидоспоры, имеющие вид одной утолщенной овальной клетки, цепочки таких клеток или конгломерата округлой, удлиненно-овальной или неправильной формы.
Морфология изолятов MF-P403-011 и MF-P485-010 (рис. 2Б) совпала с описанием A. tellustris (Simmons, 1983). Конидии 18–33 × 6–10 мкм с (2)3–5 поперечными толстыми темными перегородками располагались по 1–3 на конидиеносце. Изолят формировал хламидоспоры более или менее округлые, удлиненно-овальные или неправильной формы.
Морфологическое исследование изолята MF-P183-010 и сравнение с описанием Pyrenophora biseptata (Ellis, 1971; Sivanesan, 1987) подтвердило его принадлежность этому виду. Конидии были вытянуто-овальными или широко булавовидными, иногда незначительно ассиметричными, от светло-коричневых до коричневых, с (1)2–3 поперечными перегородками того же цвета, что и стенка конидии (рис. 3А). Размер конидий 20–35(40) × × 12–17 мкм. Конидии располагались по 4–12 на относительно длинных коленчато изогнутых конидиеносцах. Особенностью изолята была способность формировать хламидоспоры, более или менее округлые, овальные или неправильной формы.
Рис. 3.
Морфология Embellisia-подобных гифомицетов, не относящихся к роду Alternaria: А – Drechslera biseptata (сверху – хламидоспоры, снизу – конидии); Б – Alternariaster helianthi (конидии).
Оба изолята Alternariaster helianthi соответствовали морфологическим критериям данного вида (Simmons, 2007). Конидии одиночные, бледно-серо-желтые или бледно-коричневые, узкоовальные, эллипсоидальные или почти цилиндрические, до 80–130(160) × 18–23(30) мкм с 7–12 поперечными перегородками (рис. 3б).
ОБСУЖДЕНИЕ
Большинство Embellisia-подобных гифомицетов известно по единичным находкам. По всей видимости, Alternaria embellisia и A. chlamydosporigena – наиболее широко распространенные виды рода. Образцы чеснока, содержащие A. embellisia, были собраны в разных странах Европы, Австралии, Африки, Северной и Южной Америки (Simmons, 1971; David, 1991; Corlett, 1996). По сообщению E.G. Simmons (1971) A. embellisia мог быть с легкостью выделен из луковиц чеснока, купленных в штате Массачусетс на местных рынках. Недавно этот вид был найден и в восточной Азии (Lee et al., 2002). Вид A. chlamydosporigena также был известен в виде изолятов из Европы, Северной Америки, Передней Азии и с севера Африки (Simmons, 1971). Данный вид был выделен из корней, колосьев и семян зерновых культур, почвы, стеблей льна и из моркови. Вид A. tellustris был обнаружен в нескольких разнообразных почвенных образцах из Северной Америки, в юго-западной Азии и Антарктиды (Simmons, 1983).
Все три обсуждаемых вида Alternaria относятся к одной секции – Embellisia (Woudenberg et al., 2013). Перевод в род Alternaria сопровождался для двух видов присвоением новых названий из-за того, что их эпитеты были уже ранее использованы для обозначения видов Alternaria (A. allii и A. chlamydospora). Виды из других семи секций, включающих виды упраздненного рода Embellisia (главным образом секция Embellisioides), нами обнаружены не были.
Вид Pyrenophora biseptata не имеет четкой субстратной приуроченности. Был обнаружен на разных континентах (Европа, Австралия, Северная Америка) на злаках и растениях других семейств, в т.ч. на споровых (Ellis, 1971; Sivanesan, 1987). Cделанная нами находка этого вида расширяет представление о его ареале и подтверждает его способность поражать широкий спектр растений.
Alternariaster helianthi распространен весьма широко и ассоциирован исключительно с подсолнечником и вызывает его вредоносное заболевание – темно-бурую пятнистость (альтернариоз, эмбеллизия). Принято считать, что этот патоген распространен почти на всей территории возделывания культуры (Tikhonova, Alifirova, 1988). Нами он ранее наблюдался в Белгородской обл., Республике Адыгея и Приморском крае (Gannibal, 2011).
Таким образом, в России и Казахстане нами было обнаружено пять Embellisia-подобных гифомицетов. Три вида относились к Alternaria секции Embellisia. Первыми находками для России являются следующие: A. embellisia на чесноке в Псковской обл. и A. tellustris на маке в Крыму и арбузе в Астраханской области. Гриб A. chlamydosporigena выявлен на корнях пшеницы и впервые на листьях цикория и корнях мака в Краснодарском крае, Алматинской обл. Казахстана и Крыму (первая находка для Крыма). Подтверждено обнаружение Alternariaster helianthi на подсолнечнике в Приморском крае и Белгородской области. На листьях ситника жабьего на Камчатке обнаружен вид Drechslera biseptata, что можно считать первой находкой для региона и первым обнаружением на данном растении-хозяине.
Авторы благодарны коллегам из Лаборатории микологии и фитопатологии ВИЗР И.А. Казарцеву, А.А. Варехиной и М.М. Гомжиной за помощь, оказанную при секвенировании ДНК. Работа была выполнена при финансовой поддержке Российского научного фонда (проект № 14-26-00067).
Список литературы
Alcorn J.L. The taxonomy of “Helminthosporium” species. Ann. Rev. Phytopathol. 1988. V. 26. P. 37–56.
Alcorn J.L. New combinations and synonymy in Bipolaris and Curvularia, and a new species of Exserohilum. Mycotaxon. 1991. V. 41. P. 329–343.
Altschul S.F., Gish W., Miller W., Myers E.W., Lipman D.J. Basic local alignment search tool. J. Mol. Biol. 1990. V. 215. P. 403–410.
Alves J.L., Woudenberg J.H.C., Duarte L.L., Crous P.W., Barreto R.W. Reappraisal of the genus Alternariaster (Dothideomycetes). Persoonia. 2013. V. 31 (12). P. 77–85.
Berbee M.L., Pirseyedi M., Hubbard S. Cochliobolus phylogenetics and the origin of known, highly virulent pathogens, inferred from ITS and glyceraldehydes-3-phosphate dehydrogenase gene sequences. Mycologia. 1999. V. 91 (6). P. 964–977.
Corlett M. Embellisia allii. Can. J. Plant Pathol. 1996. V. 18. P. 486–487.
David J.C. Embellisia allii. IMI Descriptions of Fungi and Bacteria No. 1078. Mycopathologia. 1991. V. 116. P. 59–60.
David J.C., Coles K., Fisher J., Moss S.T. A new species of Embellisia from soil with high levels of heavy metals. Mycoscience. 2000. V. 41. P. 533–537.
Doyle J.J., Doyle J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987. V. 19. P. 11–15.
Ellis M.B. Dematiaceous Hyphomycetes. Kew, CAB International, 1971.
Gannibal Ph.B. Small-spored species of the genus Alternaria on grasses. Mikologiya i fitopatologiya. 2004. V. 38 (3). P. 19–28 (in Russ.).
Gannibal Ph.B. Species composition, systematics and geography of agents of sunflower alternarioses in Russia. Vestnik zashchity rasteniy. 2011. V. 1. P. 13–19 (in Russ.).
Hong S.G., Robert A.C., Lawrence C.B., Pryor B.M. Alt a 1 allergen homologs from Alternaria and related taxa: analysis of phylogenetic content and secondary structure. Fungal Genet. Biol. 2005. V. 42. P. 119–129.
Hoog de G.S., Muller P.J. A new species of Embellisia associated with skin disease of hyacinths. Neth. J. Plant Path. 1973. V. 79. P. 85–93.
Hoog de G.S., Seigle-Murandi F., Steinman R., Eriksson K.E. A new species of Embellisia from the North Sea. J. Microbiol. 1985. V. 51. P. 409–413.
Kozyreva E.P., Primakovskaya M.A. Embellisia of sunflower. Zashchita rasteniy. 1981. V. 8. P. 38 (in Russ.).
Lawrence D.P., Gannibal Ph.B., Peever T.L., Pryor B.M. The Sections of Alternaria: Formalizing species-group concepts. Mycologia. 2013. V. 105 (3). P. 530–546.
Lawrence D.P., Park M.S., Pryor B.M. Nimbya and Embellisia revisited, with nov. comb for Alternaria celosiae and A. perpunctulata. Mycol. Progress. 2012. V. 11. P. 799–815.
Lee H.B., Chang-Jin Kim C.-J., Yu S.H. First Report of Bulb Canker of Garlic Caused by Embellisia allii in Korea. Mycobiology. 2002. V. 30 (4). P. 240–243.
Li Y., Nan Z. A new species, Embellisia astragali sp. nov., causing standing milk-vetch disease in China. Mycologia. 2007. V. 99. P. 406–411.
Muntanjola-Cvetković M., Ristanović B. A new species of Embellisia isolated from seawater. Mycologia. 1976. V. 68. P. 47–51.
Pidoplichko N.M. Fungi – parasites of cultivated plants. Identification key. Vol. 2. Fungi Imperfecti. Kiev, Naukova Dumka, 1977 (in Russ.).
Pryor B.M., Bigelow D.M. Molecular characterization of Embellisia and Nimbya species and their relationship to Alternaria, Ulocladium and Stemphylium. Mycologia. 2003. V. 95. P. 1141–1154.
Shoemaker R.A. Nomenclature of Drechslera and Bipolaris, grass parasites segregated from “Helminthosporium”. Can. J. Bot. 1959. V. 37. P. 879–887.
Shoemaker R.A. Drechslera Ito. Can. J. Bot. 1962. V. 40. P. 1143–1150.
Simmons E.G. Typification of Alternaria, Stemphylium, and Ulocladium. Mycologia. 1967. V. 59. P. 67–92.
Simmons E.G. Helminthosporium allii as type of a new genus. Mycologia. 1971. V. 63. P. 380–386.
Simmons E.G. An aggregation of Embellisia species. Mycotaxon. 1983. V. 17. P. 216–241.
Simmons E.G. Embellisia and related teleomorphs. Mycotaxon. 1990. V. 38. P. 251–265.
Simmons E.G. Alternaria taxonomy: current status, viewpoint, challenge. In: J. Chełkowski, A. Visconti. (eds). Alternaria. Biology, plant diseases and metabolites. Amsterdam, Elsevier, 1992. P. 1–36.
Simmons E.G. Alternaria: an identification manual. Utrecht, 2007.
Sivanesan A. Graminicolous species of Bipolaris, Curvularia, Drechslera, Exserohilum and their teleomorphs. Kew, CAB International, 1987.
Stamatakis A. Raxml-vi-hpc: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics. 2006. V. 22. P. 2688–2690.
Thompson J.D., Gibson T.J., Plewniak F., Jeanmougin F., Higgins D.G. The ClustalX windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucl. Acids Res. 1997. V. 24. P. 4876–4882.
Tikhonova O.I., Alifirova T.P. Harmfulness of sunflower embellisia. Bolezni podsolnechnika. Krasnodar, VNIIMK, 1988. P. 19–23 (in Russ.).
Woudenberg J., Groenewald J., Binder M., Crous P. Alternaria redefined. Stud. Mycol. 2013. V. 75. P. 171–212.
Ганнибал Ф.Б. (Gannibal) Мелкоспоровые виды рода Alternaria на злаках // Микология и фитопатология. 2004. Т. 38. № 3. С. 19–28.
Ганнибал Ф.Б. (Gannibal) Видовой состав, систематика и география возбудителей альтернариозов подсолнечника в России // Вестник защиты растений. 2011. Т. 1. С. 13–19.
Козырева Е.П., Примаковская М.А. (Kozyreva, Primakovskaya) Эмбеллизия подсолнечника // Защита растений. 1981. № 8. С. 38.
Пидопличко Н.М. (Pidoplichko) Грибы-паразиты культурных растений. Определитель. Т. 2. Грибы несовершенные. Киев: Наукова Думка, 1977. 300 с.
Тихонова О.И., Алифирова Т.П. (Tikhonova, Alifirova) Вредоносность эмбеллизии подсолнечника // Болезни подсолнечника. Краснодар: ВНИИМК, 1988. С. 19–23.
Дополнительные материалы отсутствуют.
Инструменты
Микология и фитопатология