

Микология и фитопатология, 2019, T. 53, № 4, стр. 246-253
Динамика расового состава Pyrenophora tritici-repentis в Северо-Кавказском регионе
О. Ю. Кремнева 1, *, Г. В. Волкова 1, **, Н. М. Коваленко 2, ***
1 Всероссийский НИИ биологической защиты растений
350039 Краснодар, Россия
2 Всероссийский научно-исследовательский институт защиты растений
196608 Санкт-Петербург, Россия
* E-mail: kremenoks@mail.ru
** E-mail: vniibzr@mail.kuban.ru
*** E-mail: info@vizr.spb.ru
Поступила в редакцию 01.10.2018
После доработки 10.10.2018
Принята к публикации 21.12.2018
Аннотация
Желтая пятнистость листьев пшеницы (возбудитель Pyrenophora tritici-repentis) является экономически значимым заболеванием во всем мире. В последние годы данное заболевание серьезно влияет на производство пшеницы – как по количеству, так и по качеству производимого зерна. Потери урожая при эпифитотийном развитии достигают 50%. Ранее в наших работах была показана различная вирулентность полученных в Северо-Кавказском регионе изолятов возбудителя. Целью настоящей работы явилось определение расового состава, изменчивости и разнообразия изолятов P. tritici-repentis на Северном Кавказе в период 2010–2014 гг. Материалом исследований послужили 628 моноконидиальных изолятов желтой пятнистости листьев пшеницы, выделенных из трех агроклиматических зон Северного Кавказа: южной предгорной, центральной и западной приазовской. Исследования проводили в условиях теплицы – на растениях пшеницы, выращенных до фазы двух листьев. Расовый состав популяций гриба определяли на дифференцирующем наборе, предложенном Ламари c соавторами: сорта Glenlea, Salamouni и линии 6B662, 6B365. В результате на протяжении пяти лет изучения доминирующими расами во всех зонах являлись 1, 2 и 8. Изоляты гриба расы 1 встречались с частотой от 15 до 60.6%; изоляты расы 2 – от 13.6 до 28.2%, изоляты расы 8 – от 13.8 до 41.9%. Изоляты рас 3 и 4 встречались каждый год, но не во всех агроклиматических зонах, с переменной частотой от 2.7 до 18.5%. Изоляты рас 5, 6 и 7 встречались еще реже, с частотой от 2.6 до 12.0%, а в отдельные годы отсутствовали в популяции патогена. Уровень разнообразия популяции гриба по расовому составу в каждой агроклиматической зоне оказался достаточно высоким (от 1.10 до 1.89). Попарное сравнение выборок P. tritici-repentis изученных зон между собой в отдельные годы значительных различий не выявило (значение индекса Роджерса не превышало 0.3). Это дает основание сделать вывод о единстве популяции P. tritici-repentis по расовому составу во всех изученных агроклиматических зонах Северного Кавказа.
ВВЕДЕНИЕ
Желтая пятнистость листьев пшеницы является экономически значимым заболеванием во всем мире. Ее возбудитель – гомоталличный аскомицет Pyrenophora tritici-repentis (Died.) Drechsler [анаморфа Dreschlera tritici-repentis (Died.) Shoemaker] из порядка Pleosporales. Имеются сообщения о широком распространении болезни в Европе, Юго-Западной Азии, Центральной Азии, Северной и Южной Америке, Африке, Австралии (Maraite et al., 2006; Oliver et al., 2008; Momeni et al., 2014; Kokhmetova et al., 2017).
В России на данный патоген впервые обратили внимание в 1985 г. (Granin et al., 1989). С 2000-х гг. болезнь распространилась по всей России: она обнаружена в Дагестане, Западной Сибири, на Алтае (Mikhailova et al., 2007). В последние 10 лет распространение болезни в Северо-Кавказском регионе на некоторых производственных полях Краснодарского, Ставропольского краев, Ростовской обл. достигало 80–100%, а развитие на восприимчивых сортах – 60–80% (Kremneva et al., 2013, 2014).
В последние годы это заболевание является экономически важным из-за серьезного влияния на производство пшеницы как по количеству, так и по качеству производимого зерна. Средние потери урожая от патогена составляют 5–10%, однако в условиях, благоприятных для развития гриба, наблюдались потери до 50% (Shabeer et al., 1988). Качество зерна снижается из-за образования красных и темных пятен, вызванных инфекцией (Fernandez et al., 1998).
Для возбудителя желтой пятнистости листьев характерно большое генетическое разнообразие (Friesen et al., 2005). Информация о структуре популяции патогена необходима для успешной защиты от болезни с использованием устойчивых сортов, эффективных фунгицидов и средств биологической защиты. Изучение изменчивости популяций фитопатогенных грибов имеет как фундаментальное значение – для развития теории микроэволюции, так и сугубо прикладное, являясь необходимым звеном при создании инфекционных фонов, селекции устойчивых сортов растений, установлении ареалов популяций, территориальном размещении источников и доноров устойчивости (Afanasenko, 1998).
Информация об изменчивости популяций возбудителя желтой пятнистости листьев пшеницы представлена во многих работах. Ряд авторов (Schilder et al., 1990; Ali et al., 2003; Kremneva et al., 2007; Mikhailova et al., 2007) изучали изменчивость вирулентности в популяциях патогена, и в своих работах они отмечали, что в природной популяции P. tritici-repentis существуют группы изолятов с разной вирулентностью к сортам пшеницы.
Основываясь на способности P. tritici-repentis вызывать некрозы и хлорозы на трех дифференцирующих линиях (Glenlea, 6В365 и 6В662) было описано 8 рас (Lamari et al., 2003; 2010). Хозяинспецифичные токсины Ptr ToxA, Ptr ToxB и Ptr ToxC дифференциально продуцируются расами P. tritici-repentis и служат факторами патогенности гриба. Изоляты гриба рас 2, 3 и 5 производят по одному токсину: Ptr ToxA, Ptr ToxC или Ptr ToxB, соответственно. Изоляты рас 1, 6 и 7 производят по два токсина каждый: Ptr ToxA + Ptr ToxC, Ptr ToxB + Ptr ToxC и Ptr ToxA + Ptr ToxB, соответственно. Изоляты расы 8 производят все три токсина, в то время как изоляты расы 4 не производят никаких известных токсинов и считаются непатогенными (Lamari et al., 2003; 2010).
В большинстве регионов мира, включая Америку и Австралию, расы 1 и 2 P. tritici-repentis являются доминирующими, встречаются с частотой от 40 до 90% (Friesen et al., 2005; Ali et al., 2003; Gamba et al., 2012; Aboukhaddour et al., 2013). В регионах, которые охватывают центры разнообразия пшеницы, такие как Азербайджан, Сирия и Северная Африка разнообразие по расовому составу является наибольшим (Lamari et al., 2005). В этих регионах обнаружены шесть рас – 1, 2, 3, 5, 7, 8, встречающихся с различной частотой.
Л.А. Михайлова с соавторами (Mikhailova et al., 2014, 2015) проводила изучение расового состава популяций P. tritici-repentis, распространенных на территории России. С использованием канадских дифференциаторов среди изолятов P. tritici-repentis были определены 6 рас. Расы 1, 2 и 8 доминировали и встречались во всех изученных популяциях. Было показано, что омская популяция отличалась от уфимской, северокавказской и северо-западной популяций по расовому составу, по частоте изолятов, вирулентных к сортам-дифференциаторам, и величине клональной фракции.
Северо-Кавказский регион является основным производителем зерна в России – до 20% от общероссийского объема. Только в Краснодарском крае высевается более 100 генетически разнородных сортов пшеницы (Romanenko et al., 2017). Для создания и использования сортов со стабильной и эффективной устойчивостью к болезни необходимо знание структуры популяции гриба.
Целью настоящих исследований было определение расового состава, изменчивости и разнообразия популяции возбудителя желтой пятнистости листьев в Cеверо-Кавказском регионе России.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования по изучению расового состава проводили с 2010 по 2014 гг. Ежегодно были обследованы производственные и селекционные посевы озимой пшеницы в фазу колошения – молочной спелости в разных агроклиматических зонах Северного Кавказа (Краснодарский и Ставропольский края), отличающихся по влаго- и теплообеспеченности (Batova, 1966). Листья с симптомами желтой пятнистости были собраны с 3–5 полей (каждое поле ≥1 га) из каждого района. В общей сложности было собрано по 60 листьев с каждого поля с интервалом не менее 1 м друг от друга. Изоляты, включенные в это исследование, были собраны с сортов озимой мягкой и твердой пшеницы российской селекции НЦЗ им. П.П. Лукьяненко, Научного центра “Донской”.
Было проанализировано 628 моноконидиальных изолятов P. tritici-repentis, выделенных из трех агроклиматических зон Северного Кавказа. 204 изолята выделены из 6 районов южной предгорной зоны (Лабинский, Курганинский, Андроповский, Предгорный, Советский, Георгиевский), 237 изолятов из 6 районов центральной зоны (Тимашевский, Динской, Усть-Лабинский, Кавказский, Гулькевичский, Новоалександровский) и 187 – из 4 районов западной приазовской зоны (Ейский, Каневский, Кущевский, Щербиновский). Ежегодно изучали от 100 до 130 изолятов.
Выделение моноспоровых изолятов проводили по методике, описанной Л.А. Михайловой с соавторами (Mikhailova et al., 2002). Из пластинки листа вырезали сегменты, на которых располагалось одно инфекционное пятно и прилегающий к нему участок зеленой ткани. Сегменты листьев помещали в марлевые мешочки, которые промывали в проточной воде в течение часа. Стерилизацию отрезков проводили в 0.1%-м р-ре AgNO3 30 сек. Простерилизованные сегменты помещали в чашку Петри со средой V-4 (4 части сока свеклы, 3 – сока сельдерея, 2 – сока моркови и 1 – томата, 850 мл воды, 1.5 г СаСО3). Для получения конидиального спороношения культуру патогена выдерживали в течение 5–6 суток при температуре 22–24°С под бактерицидными эритемными лампами ЛЭ-40 (освещенность 5000 лк). Затем чашки Петри с культурой помещали в холодильник при температуре 4°С на 1–2 суток для формирования конидий. Отдельные конидии иглой переносили на чистую чашку со средой.
Расовый состав популяций гриба определяли на дифференцирующем наборе, предложенном Ламари c соавторами (Lamari et al., 2005): сорта Glenlea, Salamouni и линии 6B662, 6B365. При проведении опытов растения пшеницы выращивали в набитых песком 25-миллилитровых пластиковых вазонах на гидропонике с применением питательного раствора Кнопа. Растения выращивали до фазы двух листьев в условиях теплицы при средней температуре 22°C с 16-часовым фотопериодом. В каждый вазон помещали по 5 растений каждого сорта. Опыт проводили в 3-кратной повторности. Каждую повторность инокулировали водно-конидиальной суспензией моноспорового изолята гриба.
Для приготовления конидиальной суспензии использовали 8-суточную культуру гриба. Фильтрацию осуществляли через 2–3 слоя стерильной марли. Титр спор готовой суспензии составлял 3000 спор/мл. Перед инокуляцией с растений снимали восковой налет и заражали их суспензией спор путем опрыскивания из пульверизатора. Инокулированные растения выдерживали во влажной камере в течение 11–12 ч при температуре 22°С. Учет степени развития заболевания проводили на 7-е сутки по шкале Rees et al. (1987). Согласно этой шкале изоляты с проявлением болезни 1–2 балла относили к авирулентным, с типом реакции 3–5 баллов – к вирулентным.
Анализ частоты расового состава популяций проводили между выборками изолятов гриба из разных агроклиматических зон и по годам. Уровень разнообразия выборок изолятов P. tritici-repentis по расовому составу оценивали с помощью индекса Shannon: ${{Н }_{w}} = - \sum {pi\ln (pi)} $, где pi – частота i-го фенотипа в данной популяции (Kolmer et al., 2001).
Различия между выборками патогена по расовому составу оценивали с помощью расстояния Роджерса: ${{H}_{r}} = 1{\text{/}}2\sum {(pi1 - pi2),} $ где pi1 – частота i-го фенотипа в первой популяции; pi2 – частота i-го фенотипа во второй популяции (Rogers, 1972). Различия в популяциях патогенов считаются существенными при значении индекса Роджерса более 0.3.
РЕЗУЛЬТАТЫ
Результаты определения расового состава образцов популяций возбудителя желтой пятнистости P. tritici-repentis в трех агроклиматических зонах Северного Кавказа за 2010–2014 гг. представлены в табл. 1.
Таблица 1.
Частота рас Pyrenophora tritici-repentis в различных агроклиматических зонах Северного Кавказа (2010–2014 гг.)
| Агроклиматическая зона | Частота рас, % | |||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
| 2010 | ||||||||
| Южная предгорная | 60.6 | 18.2 | 0.0 | 3.0 | 0.0 | 0.0 | 3.0 | 15.2 |
| Центральная | 40.5 | 13.6 | 8.1 | 2.7 | 0.0 | 0.0 | 2.7 | 32.4 |
| Западная приазовская | 44.8 | 20.8 | 17.2 | 0.0 | 0.0 | 0.0 | 3.4 | 13.8 |
| 2011 | ||||||||
| Южная предгорная | 34.4 | 22.9 | 0.0 | 5.7 | 2.8 | 0.0 | 2.8 | 31.4 |
| Центральная | 29.8 | 21.6 | 2.7 | 8.1 | 2.7 | 0.0 | 5.4 | 29.7 |
| Западная приазовская | 25.9 | 14.8 | 0.0 | 18.5 | 7.4 | 0.0 | 3.7 | 29.6 |
| 2012 | ||||||||
| Южная предгорная | 40.0 | 5.0 | 0.0 | 10.0 | 5.0 | 0.0 | 0.0 | 40.0 |
| Центральная | 48.3 | 13.7 | 6.9 | 10.3 | 0.0 | 0.0 | 0.0 | 20.7 |
| Западная приазовская | 27.2 | 22.7 | 4.5 | 4.5 | 9.1 | 0.0 | 0.0 | 31.8 |
| 2013 | ||||||||
| Южная предгорная | 20.0 | 13.3 | 0.0 | 16.7 | 6.7 | 6.7 | 3.3 | 33.3 |
| Центральная | 25.8 | 16.1 | 3.2 | 6.5 | 0.0 | 3.2 | 3.2 | 41.9 |
| Западная приазовская | 16.0 | 20.0 | 0.0 | 16.0 | 12.0 | 4.0 | 0.0 | 32.0 |
| 2014 | ||||||||
| Южная предгорная | 15.0 | 25.0 | 0.0 | 0.0 | 10.0 | 5.0 | 5.0 | 40.0 |
| Центральная | 15.4 | 28.2 | 12.8 | 10.3 | 7.7 | 2.6 | 5.1 | 17.9 |
| Западная приазовская | 20.8 | 25.0 | 0.0 | 8.3 | 0.0 | 8.3 | 0.0 | 37.5 |
Очевидно, что на протяжении пяти лет изучения доминирующими расами во всех зонах являлись 1, 2 и 8. В 2010 г. изоляты гриба 1 расы встречались с частотой от 40.5 (центральная зона) до 60.6% (южная предгорная зона). Но с годами наблюдалось постепенное снижение частоты изолятов этой расы до 15.0% (южная предгорная зона).
Процентное содержание изолятов патогена, принадлежащих к расе 2, было более стабильным на протяжении периода изучения, но к 2014 г. наблюдалось постепенное увеличение частот таких изолятов – их доля в центральной зоне составила 28.2%, в южной предгорной и западной приазовской – по 25.0%.
Изоляты P. tritici-repentis расы 8 продуцируют все три токсина, и их содержание в выборках популяций фитопатогена было достаточно высоким (в среднем 25–35%) во всех агроклиматических зонах за годы изучения. Максимальная частота расы 8 – 40.0% – отмечена в южной предгорной зоне в 2012, 2014 гг. и 41.9% – в центральной зоне в 2013 г. Динамика частот доминирующих рас северокавказской популяции P. tritici-repentis с 2010 по 2014 гг. представлена на рис. 1.
Рис. 1.
Динамика частот доминирующих рас северокавказской популяции Pyrenophora tritici-repentis (2010–2014 гг.).
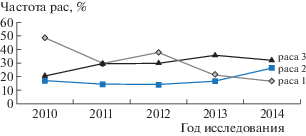
Изоляты расы 4 встречались каждый год, но не во всех агроклиматических зонах, с переменной частотой от 2.7% (центральная зона, 2010 г.) до 18.5% (западная приазовская зона, 2011 г.). Изоляты 3, 5 и 7 рас встречались еще реже и в отдельные годы отсутствовали в популяции патогена. А изоляты расы 6 в популяциях 2010–2012 гг. не выявлены, но в 2013–2014 гг. наблюдалось их появление во всех агроклиматических зонах Северного Кавказа с частотой от 2.6 до 8.3%.
Уровень разнообразия популяции гриба по расовому составу представлен в табл. 2. Несмотря на отсутствие в отдельные годы некоторых рас в северокавказской популяции P. tritici-repentis, в целом уровень разнообразия в каждой агроклиматической зоне остается довольно высоким. Самые низкие показатели индекса разнообразия отмечены в 2010 г., а в 2013–2014 гг. популяция патогена была наиболее разнообразной. В 2014 г. был достигнут максимальный из полученных показатель индекса разнообразия в центральной зоне (Sh = 1.89).
Таблица 2.
Индекс разнообразия Шеннона популяции Pyrenophora tritici-repentis в различных агроклиматических зонах Северного Кавказа (2010–2014 гг.)
| Агроклиматическая зона | Годы сбора | |||||
|---|---|---|---|---|---|---|
| 2009 | 2010 | 2011 | 2012 | 2013 | 2014 | |
| Южная предгорная | 1.56 | 1.11 | 1.43 | 1.26 | 1.73 | 1.53 |
| Центральная | 1.64 | 1.40 | 1.60 | 1.37 | 1.52 | 1.89 |
| Западная приазовская | 1.32 | 1.38 | 1.62 | 1.55 | 1.66 | 1.45 |
| Среднее значение индекса разнообразия | 1.51 | 1.29 | 1.55 | 1.39 | 1.63 | 1.63 |
Агроклиматические зоны Северного Кавказа отличаются по влаго- и теплообеспеченности, как известно, это – один из основных факторов, влияющих на развитие желтой пятнистости листьев. Ранее в наших работах было показано различие выборок изолятов из этих зон по признаку вирулентности (Kremneva et al., 2007). Из южной предгорной зоны изоляты P. tritici-repentis характеризовались наибольшей вирулентностью в сравнении с изолятами из центральной и западной приазовской зон.
Западная приазовская зона отдалена центральной зоной от южной предгорной. Расстояние от граничных точек каждой зоны составляло минимум 100 км. Сортимент высеваемых сортов также в этих зонах различен. Поэтому представлялось интересным, отличается ли северокавказская популяция патогена в различных агроклиматических зонах по расовому составу. Для этого было проведено попарное сравнение по индексу Роджерса выборок изолятов P. tritici-repentis из изученных зон между собой в годы исследований (табл. 3).
Таблица 3.
Различие между популяциями Pyrenophora tritici-repentis разных агроклиматических зон Северного Кавказа по индексу Роджерса (2010–2014 гг.)
| Годы исследования | Сравниваемые пары популяций патогена различных агроклиматических зон Северного Кавказа | ||
|---|---|---|---|
| Южная предгорная – центральная | Южная предгорная – западная приазовская | Центральная – западная приазовская | |
| 2010 | 0.2 | 0.3 | 0.2 |
| 2011 | 0.1 | 0.2 | 0.2 |
| 2012 | 0.2 | 0.3 | 0.3 |
| 2013 | 0.2 | 0.1 | 0.3 |
| 2014 | 0.3 | 0.2 | 0.3 |
В целом, по всем зонам значительных различий не обнаружено (значение индекса Роджерса не превышало 0.3) даже между географически отдаленными зонами – южной предгорной и западной приазовской. Это дает основание сделать вывод о единстве северокавказской популяции P. tritici-repentis по расовому составу во всех изученных агроклиматических зонах.
Для того, чтобы более детально проследить динамику расового состава в каждой агроклиматической зоне на протяжении всего периода изучения, нами было проведено сравнение между годами исследований изолятов патогена, собранных в каждой из трех зон Северного Кавказа (табл. 4). Максимальными были различия между изолятами гриба западной приазовской зоны 2010 г. и остальными годами исследований. Изоляты P. tritici-repentis южной предгорной зоны 2010 г. также показывают существенные различия с остальными годами исследований.
Таблица 4.
Различие между популяциями Pyrenophora tritici-repentis разных агроклиматических зон Северного Кавказа по годам исследований
| Сравниваемые годы | Различие по индексу Роджерса | ||
|---|---|---|---|
| Южная предгорная | Центральная | Западная приазовская | |
| 2010–2011 | 0.3 | 0.2 | 0.4 |
| 2010–2012 | 0.4 | 0.2 | 0.3 |
| 2010–2013 | 0.4 | 0.2 | 0.5 |
| 2010–2014 | 0.5 | 0.4 | 0.4 |
| 2011–2012 | 0.2 | 0.3 | 0.2 |
| 2011–2013 | 0.2 | 0.2 | 0.2 |
| 2011–2014 | 0.3 | 0.3 | 0.3 |
| 2012–2013 | 0.3 | 0.3 | 0.2 |
| 2012–2014 | 0.4 | 0.5 | 0.2 |
| 2013–2014 | 0.1 | 0.4 | 0.2 |
В целом, при сравнении выборок патогена одних и тех же агроклиматических зон между годами: значения индекса Роджерса колебались от 0.1 (южная предгорная зона 2013–2014 гг.) до 0.5 (южная предгорная зона 2010–2014 гг.). Наибольшие различия во всех зонах получены при сравнении популяций P. tritici-repentis 2010 и 2014 гг.
ОБСУЖДЕНИЕ
В результате изучения расового состава в Северо-Кавказском регионе России выявлено доминирование рас 1, 2 и 8. К 2014 г. изоляты расы 1, которые заметно доминировали на протяжении первых трех лет изучения, уступили по частоте изолятам рас 2 и 8. Расы 5 и 6, ответственные за развитие хлорозов и индукцию Ptr ТохС и Ptr ТохВ, с 2010 по 2012 гг. не встречались или встречались с низкой частотой. В 2013 и 2014 гг. эти расы отмечали с частотой до 12.0%. Можно предположить, что изоляты, синтезирующие согласно фитопатологическому тесту Ptr ТохВ, стали занимать больший ареал и вытеснять изоляты расы 1, продуцирующие Ptr ТохА и Ptr ТохС. К 2014 г. северокавказская популяция P. tritici-repentis представлена всеми 8 известными расами.
Аналогичный анализ расового состава желтой пятнистости листьев выполнили J. Šarova с соавторами (Sarova et al., 2005). Они определили в чешской популяции гриба расы 1, 2 и 4 с частотой 50, 3 и 5%, соответственно. Таким образом, раса 2 в Чехии является редкой, тогда как на европейской территории России она доминирует наряду с расой 1 (Mikhailova et al., 2015).
Имеются примеры, когда раса 2 является доминирующей или, напротив, встречается редко. С высокой частотой (36%) она была найдена в популяции Канадских Прерий и, вместе с тем, практически отсутствовала в популяции Великих Равнин. При этом расы 3, 4 и 5 в обеих популяциях встречались редко (Ali et al., 2003; Aboukhaddour et al., 2013). В Иране, расположенном близко к центру разнообразия пшеницы, изоляты гриба были сгруппированы только в две расы. Раса 1 была преобладающей и представляла 90% тестируемых изолятов, в то время как остальные 10% были классифицированы как раса 2 (Momeni et al., 2014). По данным Л.А. Михайловой и соавторов (Mikhailova et al., 2015), расы 5, 6, 7 на европейской территории РФ являются редкими, однако раса 8 встречается с высокой частотой до 40%.
Предыдущие работы показали, что популяции P. tritici-repentis имеют наибольшее разнообразие по вирулентности и расовому составу в регионах, охватывающих центр разнообразия пшеницы (Lamari et al., 2005). В этих регионах обнаружены изоляты, представляющие разнообразие рас и комбинации всех известных токсинов. Это подтверждает тот факт, что центр разнообразия растения также является центром наибольшей изменчивости патогенов этих растений (Vavilov, 1964).
Предпосылкой распространения желтой пятнистости на территории России могло быть широкое возделывание сортов озимой пшеницы Безостая 1 (Mikhailova et al., 2015). Сорт был создан в начале 1960-х гг. в Краснодарском НИИСХ, он был устойчив к бурой ржавчине, которая в то время являлась широко распространенной на территории России. С его участием создано большое число современных сортов пшеницы, широко районированных в южном регионе страны (Romanenko et al., 2017). Период возделывания сорта Безостая 1 и его производных длился 30 лет и был назван “эрой Безостая 1”. Возможно в период доминирования в посевах на Северном Кавказе и прилегающих регионах сорта Безостая 1 и его производных, создались благоприятные условия для накопления инфекционного материала болезни и возрастания численности популяции возбудителя.
В Краснодарском крае в Гулькевичском районе существует Кубанская опытная станция Всероссийского научно-исследовательского института растениеводства им. Н.И. Вавилова (в настоящее время ФИЦ “Всероссийский институт генетических ресурсов растений имени Н.И. Вавилова”). На станции высевается более 2000 образцов озимой и яровой пшеницы, собранной со всего мира и поступившей из коллекции этого института. Это место, по нашему мнению, может также играть большую роль в генетической разнородности и разнообразии популяции P. tritici-repentis, распространенной на Северном Кавказе. А широкий ассортимент высеваемых на юге России сортов мягкой и твердой пшеницы также оказывает влияние на формирование популяции патогена.
Выявленные нами различия между годами в исследуемых выборках патогена можно объяснить отличительными погодными условиям и сортиментом высеваемых сортов. 2010 г. характеризовался как засушливый. За период активной вегетации пшеницы выпало всего 20% осадков от нормы. Количество высеваемых сортов на территории Краснодарского и Ставропольского краев с 2010 к 2014 г. увеличилось. Но в пределах каждого года различий не выявлено. Это дает основание предположить, что северокавказская популяция патогена является единой на территории изученных агроклиматических зон, географически отдаленных и различных по тепло- и влагообеспеченности. В то же время популяция P. tritici-repentis является гибкой системой, быстро реагирующей на изменения различных факторов окружающей среды, что диктует необходимость ее постоянного мониторинга.
Авторы выражают благодарность научному сотруднику лаборатории иммунитета зерновых культур к грибным болезням ФГБНУ ВНИИБЗР, к.б.н. О.А. Кудиновой за помощь в проведении статистической обработки экспериментальных данных. Исследования выполнены согласно Государственному заданию № 075-00376-19-00 Министерства науки и высшего образования РФ в рамках НИР по теме № 0686-2019-0008.
Список литературы
Aboukhaddour R., Turkington K., Strelkov S. Race structure of Pyrenophora tritici-repentis (Tan spot of wheat) in Alberta. Can. J. Plant Pathol. 2013. N 35. V. 2. P. 256–268. https://doi.org/10.1080/07060661.2013.782470
Afanasenko O.S. Methods for the analysis of populations of causal agents of barley leaf blights. Collection of methodical recommendations for plant protection. 1998. P. 127–133 (in Russ.).
Ali S., Francl L.J. Population race structure of Pyrenophora tritici-repentis prevalent on wheat and noncereal grasses in the Great Plains. Plant Disease. 2003. V. 87. P. 418–422. https://doi.org/10.1094/PDIS.2003.87.4.418
Batova V.M. Agroclimatic resources of the North Caucasus. 1966. P. 132–143 (in Russ.).
Fernandez M.R., DePauw R.M., Clarke J.M., Fox S.L. Discoloration of wheat kernels by Pyrenophora tritici-repentis. Can. J. Plant Pathol. 1988. V. 20. P. 380–383. https://doi.org/10.1080/07060669809500407
Friesen T.L., Ali S., Klein K.K., Rasmussen J.B. Population genetic analysis of a global collection of Pyrenophora tritici-repentis, causal agent of tan spot of wheat. J. Phytopathol. 2005. V. 95. P.1144–1150. https://doi.org/10.1094/PHYTO-95-1144
Gamba F.M., Strelkov S.E., Lamari L. Virulence of Pyrenophora tritici-repentis in the Southern Cone Region of South America. Can. J. Plant Pathol. 2012. V. 34. P. 545–550. https://doi.org/10.1080/07060661.2012.695750
Granin E.F., Monastirnaya E.I., Kraeva G.A., Kochubei K.Yu. Pirenophorosis of winter wheat in the North Caucasus. Zashchita rasteniy. 1989. P. 12–21 (in Russ.).
Kokhmetova A., Kremneva O., Volkova G., Atishova M., Sapakhova Z. Evaluation of wheat cultivars growing in Kazakhstan and Russia for resistance to tan spot. J. Plant Pathology. № 99. V. 1. P. 161–167. https://doi.org/10.4454/jpp.v99i1.3812
Kolmer J.A., Long D.L., Kosman E., Hughes M.E. Physiologic specialization of Puccinia triticina on wheat in the United States in 2001. J. Plant Dis. 2001. V. 87. P. 859–866. https://doi.org/10.1007/s10658-018-1549-x
Kremneva O.Yu., Volkova G.V. Structure of the Pyrenophora tritici-repentis population in the North Caucasus by virulence, and racial composition, morphology and culture characteristics. Mikologiya i fitopatologiya. 2007. V. 41. N 4. P. 356–361 (in Russ.).
Kremneva O.Yu., Volkova G.V., Popov I.B. Tan spot of wheat and the influence of the predecessor on its development. Trudi Kubanskogo gosudarstvennogo agrarnogo universiteta. 2014. V. 47. P. 79–84 (in Russ.).
Kremneva O.Yu., Volkova G.V., Segeda E.S. Resistance of winter wheat varieties to the causative agent of Tan spot. Politematicheskiy setevoi elektronnyy nauchnyy zhurnal Kubanskogo gosudarstvennogo agrarnogo universiteta. 2013. N 07. V. 091. P. 1741–1751 (in Russ.).
Lamari L., Strelkov S.E. The wheat – Pyrenophora tritici-repentis interaction: progress towards an understanding of tan spot disease. Can. J. Plant Pathol. 2010. V. 32. P. 4–10. https://doi.org/10.1007/s00122-016-2670-x
Lamari L., Strelkov S.E., Yahyaoui A., Amedov M., Saidov M., Djunusova M., Koichibayev M. Virulence of Pyrenophora tritici-repentis in the countries of the Silk Road. Can. J. Plant Pathol. 2005. V. 27. P. 383–388. https://doi.org/10.1080/07060660509507236
Lamari L., Strelkov S.E., Yahyaoui A., Orabi J., Smith R.B. The identification of two new races of Pyrenophora tritici-repentis from the host centre of diversity confirms a one to one relationship in tan spot of wheat. Phytopathology. 2003. V. 93. P. 391–396. https://doi.org/10.5423/PPJ.OA.10.2016.0214
Maraite H., Berny J.F., Coffin A. Epidemiology of tan spot in Belgium. Advances in tan spot, North Dakota Agriculture Experimental Station. 1992. P. 73–79. https://doi.org/10.3103/S1068367411040094.
Maraite H., Mercado Vergnes D., Renard M.E., Zhanarbekova A., Duveiller E. The importance of a variety of pathogens in combating yellow spot and burn of wheat leaves in central Asia. Agromeridian. 2006. V. 3. N 2. P. 105–114. https://doi.org/10.1007/s10681-007-9380-z
Mikhailova L.A., Gultyaeva E.I., Kokorina I.M. Laboratory methods of cultivation of wheat tan spot causal agent Pyrenophora tritici-repentis. Mikologiya i fitopatologiya. 2002. V. 36. N 1. P. 63–67 (in Russ.).
Mikhailova L.A., Kovalenko N.M., Mironenko N.V., Rosseeva L.P. Populations of Pyrenophora tritici-repentis in the territory of Russia. Mikologiya i fitopatologiya. 2015. V. 49. N 4. P. 257–261 (in Russ.).
Mikhailova L.A., Mironenko N.V., Kovalenko N.M. Populations of Pyrenophora tritici-repentis in the north Caucasus and north-west Russia: the racial composition and dynamics of virulence. Mikologiya i fitopatologiya. 2014. V. 48. N 6. P. 393–400 (in Russ.).
Mikhailova L.A., Ternyuk I.G., Mironenko N.V. Population structure of Pyrenophora tritici-repentis from the European part of Russia on the virulence. Mikologiya i fitopatologiya. 2007. V. 41. N 3. P. 269–275 (in Russ.).
Momeni H., Aboukhaddour R., Javan-Nikkhah M., Razavi M., Naghavi M.R., Akhavan A., Strelkov S.E. Race identification of Pyrenophora tritici-repentis in Iran. J. Plant Pathology. 2014. N 96. V. 2. P. 287–294. https://doi.org/10.4454/JPP.V9612/036
Oliver R.P., Lord M., Rybak K., Faris J.D., Solomon P.S. Emergence of tan spot disease caused by toxigenic Pyrenophora tritici-repentis in Australia is not associated with increased deployment of toxin sensitive cultivars. Phytopathology. 2008. V. 98. P. 488–491. https://doi.org/10.1094/PHYTO-98-5-0488
Rees R.G., Platz G.J., Mayer R.J. Susceptibility of Australian wheats to Pyrenophora tritici-repentis. Aust. J. Agric. Res. 1987. V. 39. P. 141–151. https://doi.org/10.1007/BF00032151
Rogers J.S. Measures of genetic similarity and genetic distance. Studies in Genetics. University of Texas. Austin. 1972. P. 143–145.
Romanenko A.A., Bespalova L.A., Lavrenchuk N.F. et al. Catalogue. Varieties and hybrids KNIISH them P.P. Lukyanenko. Rossia, Krasnodar, 2017 (in Russ.).
Sarova J., Hanzalova A., Bartos P. Races of Pyrenophora tritici-repentis in the Czech Republic. Acta. Agrobotanica. 2005. V. 58. P. 73–78. https://doi.org/10.5586/aa.2005.011
Schilder A.M.C., Bergstrom G.C. Variation in virulence and aggressiveness within the population of Pyrenophora tritici-repentis in New York. J. Phytopathol. 1990. N 80. V. 1. P. 84–89. https://doi.org/10.1094/Phyto-80-84
Shabeer A., Bockus W.W. Tan spot effects on yield and yield components relative to growth stage in winter wheat. Plant Disease. 1988. V. 72. P. 599–602. https://doi.org/10.1094/PD-72-0599
Singh P.K., Singh R.P., Duveiller E., Mergoum M., Adhikari T.B., Elias E.M. Genetics of wheat – Pyrenophora tritici-repentis interactions. Euphytica. 2010. V. 171. P. 1–13. https://doi.org/10.1007/s10681-009-0074-6
Vavilov N.I. Immunity of plants to infectious diseases. Selected Works. Moskva, 1964. P. 132–313 (in Russ.).
Афанасенко О.С. (Afanasenko) Методы анализа популяций возбудителей пятнистостей листьев ячменя // Сборник методических рекомендаций по защите растений. СПб.: ВИЗР, 1998. С. 127–133.
Батова В.М. (Batova) Агроклиматические ресурсы Северного Кавказа. Ленинград, 1966. С. 132–143.
Вавилов Н.И. (Vavilov) Иммунитет растений к инфекционным заболеваниям. Избранные труды. Т. 5. Вып. 1. Москва. 1964. С. 132–313.
Гранин Е.Ф., Монастырная Э.И., Краева Г.А., Кочубей К.Ю. (Granin et al.) Пиренофороз озимой пшеницы на Северном Кавказе // Защита растений. 1989. Вып. 12. С. 21.
Кремнева О.Ю., Волкова Г.В. (Kremneva et al.) Структура популяции Pyrenophora tritici-repentis на Северном Кавказе по вирулентности, и расовому составу, морфологии и культуральным характеристикам // Микология и фитопатология 2007. Т. 41. № 4. С. 356–361.
Кремнева О.Ю., Волкова Г.В., Попов И.Б. (Kremneva et al.) Желтая пятнистость листьев пшеницы и влияние предшественника на ее развитие // Труды Кубанского государственного аграрного университета. 2014. Вып. 47. С. 79–84.
Кремнева О.Ю., Волкова Г.В., Сегеда Е.С. (Kremneva et al.) Устойчивость сортов озимой пшеницы к возбудителю желтой пятнистости листьев // Политематический сетевой электронный научный журнал Кубанского государственного аграрного университета. 2013. Т. 7. Вып. 91. С. 1741–1751.
Михайлова Л.А., Коваленко Н.М., Мироненко Н.В., Россеева Л.П. (Mikhailova et al.) Популяции Рyrenophora tritici-repentis на территории России // Микология и фитопатология. 2015. Т. 49. № 4. С. 257–261.
Михайлова Л.А., Гультяева Е.И., Кокорина И.М. (Mikhailova et al.) Лабораторные методы культивирования возбудителя желтой пятнистости пшеницы Pyrenophora tritici-repentis // Микология и фитопатология. 2002. Т. 36. № 1. С. 63–67.
Михайлова Л.А., Мироненко Н.В., Коваленко Н.М. (Mikhailova et al.) Популяции Рyrenophora tritici-repentis на Северном Кавказе и северо-западе России: расовый состав и динамика вирулентности // Микология и фитопатология. 2014. Т. 48. № 6. С. 393–400.
Михайлова Л.А., Тернюк И.Г., Мироненко Н.В. (Mikhailova et al.) Структура популяции Pyrenophora tritici-repentis в европейской части России по вирулентности // Микология и фитопатология. 2007. Т. 41. № 3. С. 69–275.
Романенко А.А., Беспалова Л.А., Лавренчук Н.Ф. и др. (Romanenko et al.) Каталог. Сорта и гибриды КНИИСХ им. П.П. Лукьяненко. Краснодар, 2017. 126 с.
Дополнительные материалы отсутствуют.
Инструменты
Микология и фитопатология