

Микология и фитопатология, 2020, T. 54, № 3, стр. 183-194
Влияние метаболических ингибиторов на размеры и структуру гало, индуцированных возбудителем мучнистой росы Blumeria graminis f. sp. tritici на листьях пшеницы
А. В. Бабоша 1, *, Т. В. Аветисян 1, **
1 Главный ботанический сад им. Н.В. Цицина РАН
127276 Москва, Россия
* E-mail: phimmunitet@yandex.ru
** E-mail: tarverdyan_tv@mail.ru
Поступила в редакцию 08.01.2020
После доработки 05.03.2020
Принята к публикации 10.03.2020
Аннотация
Гало – концентрическая зона 5–50 мкм измененной поверхности эпидермиса листьев пшеницы вокруг места внедрения возбудителя мучнистой росы Blumeria graminis. Изучали вариабельность гало в контроле и при обработке метаболическими ингибиторами (3-амино-1,2,4-триазол, бисиндолмалеимид гидрохлорид, дифенилениодониум хлорид, 3-изобутил-1-метилксантин, каталаза, нитрат лантана). При окраске фрагментов эпидермиса амидо черным гало представлены одноцветными округлыми синими, двуцветными сине-красными структурами или системой из нескольких синих, красных и слабоокрашенных колец. Все обработки способствовали заметному снижению доли успешных проникновений патогена в клетки растения с образованием гаусторий. Размеры гало увеличивались при обработке 3-амино-1,2,4-триазолом, ингибитором пероксидазы и каталазы, а также на растворах каталазы – фермента, разрушающего перекись водорода. Обработка нитратом лантана, блокирующим кальциевые каналы, приводила к уменьшению диаметра гало. Гало, ассоциированные с гаусториями, были в 1.5 раза меньше по сравнению с теми, которые наблюдали в отсутствие успешного развития патогена. Их размеры коррелировали с частотой появления гаусторий. Наоборот, средние размеры парных (больших и малых) гало коррелировали между собой и слабо коррелировали с числом гаусторий. Полученные данные свидетельствуют о возможности использования параметров гало для локальной диагностики жизнеспособности отдельных инфекционных единиц B. graminis.
ВВЕДЕНИЕ
Возбудитель мучнистой росы пшеницы Blumeria graminis f. sp. tritici (синоним – Erysiphe graminis) является облигатным грибным патогеном, гифы мицелия которого расположены на поверхности листьев пшеницы, а питание происходит благодаря проникновению в эпидермальные клетки с образованием гаусториев. Поражение посевов может приводить к потерям до 15–20% урожая (Sanin et al., 2011).
Характерной особенностью Blumeria graminis является появление при прорастании конидии 2-х ростковых трубок и, соответственно, контакт с эпидермальными клетками растения, по крайней мере, в 2-х точках (Kunoh et al., 1977). Вокруг места внедрения данного патогена иногда можно наблюдать гало – концентрическую зону измененной поверхности, которая проявляется при цитохимической окраске (Edwards, 1970; Mishina et al., 2001). Малое гало (диаметром 5–15 мкм) образуется в месте контакта с эпидермисом растительной клетки первичной ростковой трубки, большое гало (15–50 мкм) – лопасти аппрессория. Непарные вторичные гало возникают в местах контакта с эпидермисом гиф мицелия. Пространственно и функционально гало связано с папиллой, которая представляет собой утолщение клеточной стенки полусферической или трубчатой формы и занимает его центральную часть. Размеры гало сильно варьируют даже в пределах одного эпидермального препарата. Показано, что диаметр вторичных гало при контакте гифальных лопастей в клетках, не содержащих гаустории, превышал таковую в клетках с гаусториями (Serezhkina et al., 1999). Средний диаметр гало может существенно увеличиваться у растений, обработанных цитокининами (Babosha et al., 2009) или перекисью водорода (Avetisyan, Babosha, 2011). Это позволяет предположить, что размеры и структура гало могут быть связаны с восприимчивостью эпидермальной ткани и характером взаимодействия растения и патогена, как локального, так и системного характера. Поскольку регуляция защитных реакций растения осуществляется при участии комплекса сигнальных систем (Tarchevskiy, 2002), представляет интерес изучение влияния ингибиторов и кофакторов этих систем на образование гало.
Цель работы – изучение влияния ряда метаболических ингибиторов на размеры гало, их морфологию и другие параметры, которые способны нести информацию о зоне контакта растения и патогена.
МАТЕРИАЛЫ И МЕТОДЫ
В работе использовали проростки восприимчивой к мучнистой росе мягкой пшеницы Triticum aestivum L. сорта Тавричанка. Растения выращивали при 20–22°С с 16-часовым фотопериодом (2000–3000 лкс). Популяцию возбудителя мучнистой росы Blumeria graminis f. sp. tritici поддерживали в лабораторных условиях на восприимчивой пшенице. Отделенные первые листья 14-суточных проростков после инфицирования инкубировали в чашках Петри на плаву абаксиальной стороной вверх с использованием водных растворов одного из исследуемых веществ (3-амино-1,2,4-триазол, бисиндолмалеимид гидрохлорид, дифенилениодониум хлорид, 3-изобутил-1-метилксантин, каталаза (ICN Pharmaceuticals Inc.), нитрат лантана 6-водный (отечественного производства). Характер ингибиторной активности и использованная в эксперименте концентрация приведены в табл. 1.
Таблица 1.
Характер активности и концентрация веществ, использованных в эксперименте
| Вариант | Концентрация, мкМ | Характер активности |
|---|---|---|
| 3-Амино-1,2,4-триазол | 50000 | ингибитор каталазы и пероксидазы (Margoliash, Novogrodsky, 1958) |
| Бисиндолмалеимид | 0.1 | ингибитор протеинкиназы С (Toullec et al., 1991) |
| Дифенилениодониум | 1 | ингибитор NO-синтазы, НАДФ Н-оксидазы и флавинсодержащих оксидаз (Cross, Jones, 1986) |
| 3-изобутил-1-метилксантин | 900 | ингибитор фосфодиэстеразы цАМФ и цГМФ (Beavo et al., 1970) |
| Каталаза | 100 | фермент, разрушающий перекись водорода |
| Нитрат лантана | 2000 | блокатор кальциевых каналов (Pantoja et al., 1992) |
Гало выявляли при окраске фрагментов эпидермиса, снятого с абаксиальной стороны листа через двое суток после инфицирования, 1%-м амидо черным в 7%-й уксусной кислоте. Цифровые фотографии получали с использованием светового микроскопа Zeiss Axioplan 2 (Carl Zeiss, Германия). На микрофотографиях идентифицировали и подсчитывали гало различных типов и гаустории без гало (рис. 1). Размеры гало и расстояние между центрами большого и малого гало определяли при помощи программы Image J. Выборка гало для вычисления долей в разных вариантах составляла 150–400. Для расчета средних значений диаметра гало использованы изображения 20–40 парных гало, поскольку только в этом случае возможна надежная идентификация большого и малого гало. Вычисляли достоверность различий с контролем, применяя t-критерий для средних и долей с поправкой Бонферрони для множественных сравнений с контролем. При этом достоверными считали различия со значениями p < 0.0083. Статистическую обработку полученных результатов проводили с использованием программы Excel.
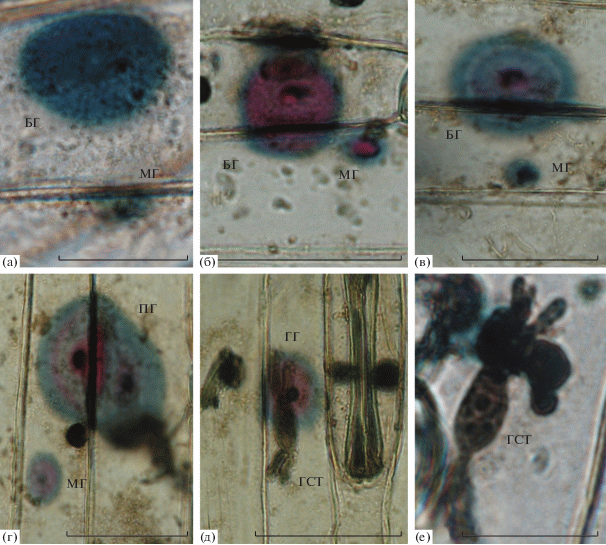
Рис. 1.
Основные типы гало Blumeria graminis на листьях пшеницы: а – одноцветные парные (большое – БГ и малое – МГ) равномерно окрашенные синие гало; б – двуцветные парные красно-синие большое и малое гало; в – трехцветное красно-сине-белое большое гало и малое одноцветное синее гало; г – перекрывающиеся гало (ПГ, перекрываются не менее, чем на треть своего диаметра); д – гало с гаусторией (ГГ); е – гаустория без гало. Масштаб – 25 мкм.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
При окраске амидо черным эпидермиса инфицированных листьев пшеницы гало отличались разнообразием морфологии и окраски (рис. 1–3). Они включали зоны округлой и кольцеобразной формы сине-голубой, пурпурно-красной и сероватой окраски. В центре внутреннего кольца (круга) практически во всех случаях присутствовала папилла, выделяющаяся более насыщенным контуром. По характеру распределения окраски все гало можно разделить на 3 группы: одноцветные равномерно окрашенные, двуцветные и трехцветные (рис. 1). Более 50% от суммы всех вариантов составляли более или менее равномерно окрашенные синие гало без выраженных светлых колец. Примерно треть составляли двуцветные сине-красные гало, у которых в центральной области синяя окраска сменялась красной. Степень такой замены могла различаться, давая оттенки, полученные от совмещения синего и красного, или чисто красную центральную зону. Красная окраска могла захватывать только область папиллы в центре или распространяться почти на все гало, кроме тонкого внешнего контура. Наконец, у части синих или двуцветных гало как в синей, так и в красной области можно было наблюдать дополнительные тонкие слабоокрашенные кольца (трехцветные сине-бело-красные, “слоистые” гало).
Рис. 2.
Варьирование морфологии гало: а – в контрольном варианте (без обработки); б – при обработке бисиндолмалеимидом; в – дифенилениодониумом. Масштаб – 25 мкм.

Рис. 3.
Варьирование морфологии гало при обработке: а – 3-изобутил-1-метилксантином; б – нитратом лантана; в – каталазой; г – 3-амино-1,2,4-триазолом. Масштаб – 25 мкм.

В табл. 2 приведены доли гало каждого из трех типов. Все варианты существенно отличались от контроля. Наиболее интересным представляется увеличение при обработке метаболическими ингибиторами доли трехцветных гало. Даже в вариантах с обработкой дифенилениодониумом и 3-изобутил-1-метилксантином, в которых, согласно использованной статистической методике, вероятность ошибки p > 0.05, доля таких гало была более чем в 3 раза больше контроля. При этом в варианте “Каталаза” их доля превышала одну треть.
Таблица 2.
Влияние ингибиторов на число гаусторий и различных типов гало в клетках эпидермиса листьев пшеницы через двое суток после инфицирования Blumeria graminis
| Вариант | Всего гало* | Гаустории, %** | Гало с гаусториями, % | Гаустории с гало, % | Гало | ||||
|---|---|---|---|---|---|---|---|---|---|
| Соприкаса- ющиеся, % | Перекрыва- ющиеся, %*** | Одноцветные, % | Двуцветные, % | Трехцветные, % | |||||
| Контроль | 253 | 18.2 | 9.5 | 32.0 | 8.3 | 2.0 | 74.7 | 23.7 | 1.6 |
| 3-Амино-1,2,4-триазол | 210 | 0.0 (0.000) | – | – | 11.9 | 4.8 (0.093) | 69.0 | 20.5 | 10.5 (0.000) |
| Бисиндолмалеимид | 232 | 11.2 (0.026) | 7.8 (0.484) | 40.0 | 6.5 | 6.0 (0.022) | 40.0 | 45.7 | 14.3 (0.000) |
| Дифенилениодониум | 228 | 5.5 (0.000) | 2.2 (0.000) | 27.8 | 10.5 | 3.9 (0.200) | 30.3 | 64.0 | 5.7 (0.015) |
| 3-Изобутил-1-метилксантин | 164 | 8.2 (0.003) | 4.3 (0.031) | 33.3 | 6.1 | 1.2 (0.557) | 49.7 | 44.8 | 5.5 (0.026) |
| Каталаза | 155 | 7.4 (0.006) | 3.2 (0.008) | 29.4 | 11.0 | 11.0 (0.000) | 18.7 | 43.2 | 38.1 (0.000) |
| Нитрат лантана | 382 | 4.4 (0.000) | 2.6 (0.000) | 37.0 | 12.8 | 4.2 (0.127) | 58.0 | 20.1 | 21.9 (0.000) |
Примечание. *Сумма гало всех типов; **доля гаусторий от суммы числа больших гало и гаусторий без гало; ***гало, которые перекрываются не менее, чем на треть своего диаметра. В скобках приведены значения p при сравнении с контролем. В столбцах “Гаустории с гало” и “Соприкасающиеся гало” достоверных различий с контролем не обнаружено. Все варианты отличались от контроля по составу одно-, дву- и трехцветных гало p(χ2) < 0.0002.
Некоторые экспериментальные варианты имели специфические особенности окраски и строения гало. Почти все гало в контроле имели размытые внешние границы, а также границы между различными кольцами. Преобладала синяя окраска, которая накладывалась даже на центральную пурпурную зону (рис. 2, а). Значительная доля гало, сходных с контролем, отмечена при обработке бисиндолмалеимидом (рис. 2, б), 3-изобутил-1-метилксантином (рис. 3, а) и нитратом лантана (рис. 3, б). В варианте с 3-амино-1,2,4-триазолом (рис. 3, г) также преобладала синяя окраска, но у большинства гало она имела очень слабую насыщенность особенно по контуру. Наоборот, каталаза (рис. 3, в) способствовала появлению гало с четкими контурами и четкими более светлыми кольцами. Для варианта обработки каталазой была характерна достаточно высокая доля слившихся, перекрывающихся гало и гало с более, чем одним центром (табл. 2, рис. 3, в). Такие гало, вероятно, возникают в случае, когда аппрессорий образует вторую лопасть при неудаче проникновения в клетку растения-хозяина с первой попытки (Sugai et al., 2010). В наших экспериментах ни одно из таких составных гало не было ассоциировано с гаусторией. Следует отметить, что в данном варианте обработки число соприкасающихся между собой гало статистически не отличалось от контроля. Очевидно, что причины, которые вызывают появление перекрывающихся и соприкасающихся гало сильно различаются, о чем свидетельствует также относительно небольшая (несущественная) величина коэффициента корреляции между ними (R = 0.44, p = 0.39).
Результатом прочного контакта аппрессориальной ростковой трубки с поверхностью эпидермальной клетки растения является попытка проникновения патогена вовнутрь. В случае успеха образуется гаустория, которая обеспечивает поступление питательных веществ и, в дальнейшем, образование колонии. При контакте с поверхностью первичной ростковой трубки проникновения не происходит: может образоваться малое гало, но гаустория всегда отсутствует. Однако и для аппрессориальной ростковой трубки не все попытки проникновения в эпидермальную клетку растения-хозяина заканчиваются успешно, поэтому только относительно небольшая доля больших гало ассоциирована с гаусторией (табл. 2). Наоборот, почти треть обнаруженных гаусторий независимо от варианта имели гало. Отсутствие гало, ассоциированных с гаусторией, может быть связано как с ослабленной реакцией растения, так и с ослаблением окраски с течением времени, которое наблюдали ранее (Mishina et al., 2001).
В первые двое суток инфекционного процесса можно обнаружить только гало в местах контакта с эпидермисом первичной и аппрессориальной (вторичной) ростковых трубок (табл. 2, 3), т.е. малое и большое гало. Одиночные гало, индуцируемые лопастями гиф, отсутствуют, поскольку гифы еще не образовались. Хотя все наблюдаемые гало должны принадлежать либо к классу больших, либо малых, при отсутствии видимых инфекционных структур установить точную принадлежность не всегда возможно. Единственным критерием классификации, не связанным с размерами, является ассоциированность большого и соответствующего ему малого гало с образованием характерной парной структуры. Поскольку каждая из ростковых трубок не обязательно устанавливает прочный контакт с поверхностью с первого раза, число наблюдаемых гало, соответствующее некоторым проросшим конидиям, может превышать число ростковых трубок. Кроме того, сам контакт может быть неполным или отсутствовать вовсе, в этом случае одно из гало (большое или малое) отсутствует. В наших экспериментах число гало в составе парных структур составляло примерно 20% от общего их числа.
Таблица 3.
Влияние ингибиторов на размеры гало в клетках эпидермиса отделенных листьев пшеницы через двое суток после инфицирования возбудителем мучнистой росы (среднее и его ошибка, значения p приведены для сравнения с контролем)
| Вариант | Большое гало, мкм | Малое гало, мкм | Расстояние между большим и малым гало, мкм | Гало с гаусторией, мкм | ||||
|---|---|---|---|---|---|---|---|---|
| среднее, х ± m | p | среднее, х ± m | p | среднее, х ± m | p | среднее, х ± m | p | |
| Контроль | 30.3 ± 1.4 | – | 12.5 ± 0.6 | – | 24.5 ± 1.5 | – | 20.9 ± 1.2 | – |
| 3-Амино-1,2,4-триазол | 41.1 ± 3.1 | 0.008 | 15.7 ± 1.6 | 0.101 | 39.2 ± 4.2 | 0.013 | – | – |
| Бисиндолма-леимид | 29.2 ± 1.0 | 0.493 | 11.0 ± 0.5 | 0.064 | 28.1 ± 1.5 | 0.095 | 19.8 ± 1.3 | 0.533 |
| Дифенилен-иодониум | 26.8 ± 1.1 | 0.053 | 12.8 ± 0.7 | 0.691 | 27.1 ± 1.6 | 0.239 | 16.2 ± 2.4 | 0.122 |
| 3-Изобутил-1-метил-ксантин | 29.5 ± 1.1 | 0.644 | 11.9 ± 0.8 | 0.549 | 25.8 ± 1.6 | 0.546 | 18.4 ± 1.7 | 0.233 |
| Каталаза | 40.7 ± 1.6 | 0.000 | 15.0 ± 1.0 | 0.043 | 31.3 ± 2.3 | 0.018 | 14.6 ± 2.1 | 0.042 |
| Нитрат лантана | 24.2 ± 0.8 | 0.000 | 9.7 ± 0.3 | 0.001 | 20.9 ± 1.0 | 0.056 | 15.7 ± 1.0 | 0.002 |
Все обработки способствовали заметному снижению доли успешных проникновений патогена в клетки растения с образованием гаусторий (табл. 2). Наименьшей доля успешных проникновений была в вариантах обработки 3-амино-1,2,4-триазолом и нитратом лантана (0 и 4.4%, соответственно). Только в этих вариантах через 7 дней после инфицирования не наблюдали спороносящих колонии. Очевидно, что изменение числа гаусторий отражает изменение устойчивости (восприимчивости) растения при обработке изучаемыми веществами.
Как видно из табл. 3, диаметр большого гало в трех вариантах (бисиндолмалеимид, дифенилениодониум, 3-изобутил-1-метилксантин) не имел достоверных отличий от контроля. При обработке 3-амино-1,2,4-триазолом и каталазой размеры большого гало заметно увеличивались. Следует отметить, что оба варианта предполагают изменения в окислительном обмене, но противоположного характера: увеличение уровня перекиси водорода в первом случае и уменьшение во втором. Обработка нитратом лантана, блокирующим кальциевые каналы, приводила к заметному уменьшению диаметра больших гало. Во многом аналогичные различия получены для диаметра малого гало и расстояния между гало в паре, хотя для этих параметров не во всех этих вариантах различия достигали уровня достоверности.
Гало, ассоциированные с гаусториями, гомологичны большим гало, однако парное малое гало в этом случае, как правило, отсутствует. Во всех вариантах обработки их размеры были существенно и более, чем в 1.5 раза меньше по сравнению с большими гало без гаусторий в тех же вариантах (табл. 3). Это согласуется с аналогичными наблюдениями (Serezhkina et al., 1999) над более поздними одиночными гифальными гало. Эти гало также имели меньшие размеры, если были ассоциированы с гаусторией. Во всех вариантах с обработкой ингибиторами гало с гаусториями показали тенденцию к уменьшению размеров по сравнению с контролем, однако только в варианте с нитратом лантана эти различия были достоверны. Меньшая статистическая выраженность различий между опытными вариантами и контролем может быть обусловлена низкой частотой гало этого типа (табл. 2), и, соответственно, недостаточной повторностью.
Показатель “расстояние между большим и малым гало”, очевидно, определяется длиной аппрессориальной ростковой трубки: в результате увеличения или уменьшения ее длины места контакта с растением первичной ростковой трубки и аппрессория оказываются разнесенными на большее или меньшее расстояние. Достоверное и более чем 1.5-кратное увеличение этого расстояния происходило при обработке 3-амино-1,2,4-триазолом (табл. 3). Заметное увеличение наблюдали также в варианте обработки каталазой. Нитрат лантана уменьшал как расстояние между большим и малым гало, так и их размеры. Известно, что прорастание конидии B. graminis иногда имеет аномальный характер с появлением очень длинных ростковых трубок длиной 50–60 мкм. Однако во всех вариантах наших экспериментов расстояние между гало было близко длине ростковых трубок нормальной морфологии (табл. 3). Вероятно, это обусловлено неспособностью аномальных структур образовать оба типа гало.
Данные табл. 2 и 3 позволяют предположить взаимосвязь между некоторыми из представленных в них показателями. В табл. 4 представлена корреляционная матрица с результатами попарных расчетов (параметры из табл. 2 и 3, не давшие достоверных значений корреляции ни с одним из других, из табл. 4 исключены), которые свидетельствуют о наличии трех корреляционных плеяд взаимозависимых параметров. Первая включает размеры большого гало, малого гало и расстояние между ними со значениями корреляции R, близкими к 0.9 и достоверными при p < 0.05. Расстояние между гало, как сказано выше, обусловлено длиной аппрессориальной ростковой трубки. Таким образом, изменение метаболизма растения при обработке метаболическими ингибиторами, вероятно, оказывает сходное влияние как на синтез полимеров клеточной стенки в зоне обоих гало, так и на рост аппрессориальной трубки гриба. Если в табл. 4 корреляция рассчитана с использованием в качестве повторности средних значений для каждого из 7 вариантов, коэффициенты в табл. 5 получены с использованием измерений в каждой паре “большое – малое гало”. Корреляция размеров гало является статистически значимой в 4 из 7 вариантов и в общей выборке, однако последняя заметно меньше полученной с использованием средних значений в табл. 4. Корреляция диаметра малого гало и расстояния между большим и малым гало не была достоверной ни в одном из вариантов, и только в объединенной выборке p = 0.049 < 0.05. Аналогичная корреляция с расстоянием между гало для большого гало в общей выборке также была заметно меньше, а в отдельных вариантах наблюдали корреляционную зависимость противоположной направленности. Таким образом, на уровне отдельной инфекционной единицы взаимосвязь между размерами гало и инфекционной трубки менее выражена, чем та, которая представлена в табл. 4 для средних по вариантам. Это может быть связано как с локальными различиями, так и с неоднозначным действием некоторых ингибиторов на разные инфекционные единицы, которые нивелируются при вычислении средних значений. Поскольку на листьях злаков индуцированные изменения устойчивости к мучнистой росе в одной клетке растения-хозяина с трудом передаются даже в соседнюю клетку (Ouchi et al., 1976), гипотетическая карта восприимчивости отдельных клеток на поверхности листьев злаков могла бы выглядеть достаточно пестрой. Та же закономерность способствует положительному для патогена балансу при повторном проникновении с помощью второй лопасти того же аппрессория при неуспехе первой (Sugai et al., 2010). При этом интенсивность роста аппрессориальной ростковой трубки, по видимому, более автономна от процессов в клеточной стенке растения, влияющих на величину гало, чем это может показаться при учете только корреляции средних величин, представленных в табл. 4.
Таблица 4.
Корреляция* между некоторыми параметрами гало и гаусторий
| Показатель | БГ | МГ | РГ | ГГ | ДГ | ПГ | 3ЦВ |
|---|---|---|---|---|---|---|---|
| Большое гало (БГ) | 1 | 0.0004 | 0.01 | 0.27 | 0.56 | 0.22 | 0.42 |
| Малое гало (МГ) | 0.92 | 1 | 0.01 | 0.47 | 0.65 | 0.39 | 0.77 |
| Расстояние между большим и малым гало (РГ) | 0.87 | 0.87 | 1 | 0.42 | 0.75 | 0.37 | 0.74 |
| Гало с гаусторией (ГГ) | 0.53 | 0.37 | 0.41 | 1 | 0.01 | 0.97 | 0.72 |
| Доля гаусторий (ДГ)** | 0.3 | 0.24 | 0.17 | 0.94 | 1 | 0.73 | 0.39 |
| Перекрывающиеся гало (ПГ) | 0.53 | 0.39 | 0.40 | 0.02 | –0.20 | 1 | 0.01 |
| Трехцветные гало (3ЦВ) | 0.37 | 0.14 | 0.15 | –0.19 | –0.44 | 0.86 | 1 |
Примечание. *Коэффициенты корреляции представлены в нижнем (левом) треугольном блоке корреляционной матрицы, соответствующие значения p (вероятность отличия корреляции от нулевого значения) в правом верхнем. Исходные данные для расчета корреляции взяты из табл. 2 и 3. Повторность – n = 7 (каждый вариант – одна повторность, кроме коэффициентов с участием показателя “доля гаусторий” n = 6: здесь исключены данные варианта с 3-амино-1,2,4-триазолом, в котором не было обнаружено гаусторий). **Доля гаусторий (ДГ) рассчитана по формуле ДГ = arcsin$\sqrt \Gamma $, где Г – доля гаусторий (число гаусторий, деленное на сумму числа больших гало и гаусторий без гало).
Таблица 5.
Попарные коэффициенты корреляции между диаметрами большого (БГ) и малого (МГ) гало и расстоянием (РГ) между ними, рассчитанные для данных каждого из вариантов обработки метаболическими ингибиторами
| Вариант | Повторность* | БГ/МГ | p** | МГ/РГ | p | БГ/РГ | p |
|---|---|---|---|---|---|---|---|
| Контроль | 14 | 0.62 | 0.02 | 0.30 | 0.30 | 0.43 | 0.13 |
| 3-Амино-1,2,4-триазол | 6 | –0.16 | 0.76 | –0.25 | 0.63 | 0.83 | 0.04 |
| Бисиндолмалеимид | 20 | 0.55 | 0.01 | 0.42 | 0.06 | 0.55 | 0.01 |
| Дифенилениодониум | 19 | 0.70 | 0.00 | 0.22 | 0.38 | 0.29 | 0.23 |
| 3-Изобутил-1-метилксантин | 16 | 0.45 | 0.08 | –0.02 | 0.94 | 0.17 | 0.53 |
| Каталаза | 14 | 0.36 | 0.20 | 0.04 | 0.89 | 0.14 | 0.62 |
| Нитрат лантана | 19 | 0.54 | 0.02 | –0.43 | 0.07 | –0.52 | 0.02 |
| Все варианты | 108 | 0.59 | 0.00 | 0.19 | 0.05 | 0.37 | 0.00 |
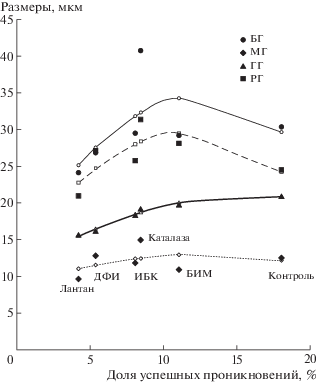
Вторая корреляционная плеяда таблицы 4 включает коррелирующие между собой размеры гало с гаусторией и частоту гаусторий (R = 0.94). Графики зависимости от частоты гаусторий размеров разных типов гало и расстояния между ними на рис. 4 позволяют предположить, что зависимость может быть нелинейной. Апроксимация полиномом второго порядка (Y = aX2 + bX + c) зависимости размеров гаусториального гало (Y) от частоты успешных проникновений патогена (доля гаусторий, X) для 6 вариантов обработки дала достоверные (p < 0.02) коэффициенты регрессии: a = –0.04* ± 0.01, b = 1.3* ± 0.2 и c = 10.7* ± 0.9. Для других параметров рис. 4 вследствие большого разброса экспериментальных точек коэффициенты регрессии не были значимыми, т.е. установить форму зависимости не удалось.
Образование гаустории – один из ключевых этапов патогенеза мучнистой росы злаков, поэтому доля успешных проникновений с образованием гаустории может служить количественным параметром для оценки устойчивости к патогену. В данной работе впервые установлено, что диаметр аппрессориальных гало, ассоциированных с гаусторией, коррелирует с частотой гаусторий и также может служить для количественной оценки устойчивости (восприимчивости) в изученной патосистеме.
Рис. 4.
Зависимость диаметра большого гало (БГ), малого гало (МГ), гало, ассоциированных с гаусторией (ГГ), и расстояния между большим и малым гало (РГ) от доли успешных проникновений патогена с образованием гаусторий (%) для 6 вариантов обработки: лантан, дифенилениодониум (ДФИ), 3-изобутил-1-метилксантин (ИБК), каталаза, бисиндолмалеимид (БИМ), контроль.
Третью корреляционную плеяду образуют коррелирующие доли “перекрываюшихся гало” и “трехцветных гало”. По мнению Мишиной и соавторов (Mishina et al., 2001), гало со сложной “слоистой” структурой из нескольких колец могут возникать при наличии определенного уровня защитных реакций растения. Перекрываюшиеся гало или гало с двумя центрами, вероятно, образуются при последовательных попытках проникновения с помощью двух лопастей аппрессория, что также связано с неудачным для B. graminis взаимодействием с растением (Sugai et al., 2010). В нашем исследовании не наблюдали образования гаусторий поблизости от слившихся или трехцветных гало. При этом коэффициент корреляции доли трехцветных гало и частоты гаусторий относительно не велик, хотя и имеет отрицательный знак. Вопрос о связи гало со сложной структурой с защитными реакциями растения требует дополнительного исследования.
Таким образом, ряд параметров, характеризующих размеры большого и малого гало, а также их окраска специфически изменяются при обработке растения некоторыми метаболическими ингибиторами. При обработке каталазой наблюдали существенное увеличение доли перекрывающихся гало с более чем одним центром. При обработке 3-амино-1,2,4-триазолом, каталазой и нитратом лантана наблюдали существенное увеличение среднего диаметра большого гало. При этом даже в пределах одного варианта обработки можно было наблюдать гало, сильно различающиеся по размерам и структуре. По-видимому, такая вариабельность свидетельствует об одновременном существовании участков ткани с очень разным метаболизмом и, соответственно, с различным уровнем восприимчивости к инфекции.
Полученные данные свидетельствуют о преимущественной роли в формировании гало оксидазной и кальциевой сигнальных систем. Считается, что перекись водорода, наряду с ее ролью в индукции защиты от патогена в качестве одного из мессенджеров оксидазной сигнальной системы имеет значение как реагент, при участии пероксидазы сшивающий полимеры клеточной стенки между собой (Shetty et al., 2008). Некоторые активные формы кислорода были обнаружены в области папиллы и гало (Thordal-Christensen et al., 1997; Hückelhoven et al., 1999; Mellersh, Heath, 2001; Trujillo et al., 2004). Очевидно, что локальное укрепление клеточной стенки может приводить к изменению способности ее полимеров взаимодействовать с красителями и тем самым влияет на характер окраски зоны гало. Поэтому действие ингибитора пероксидазы 3-амино-1,2,4-триазола на размеры и окрашиваемость гало достаточно ожидаемо. Ранее было показано, что обработка 3-амино-1,2,4-триазолом и перекисью водорода не только ингибирует развитие патогена, но и приводит к появлению более крупных гало необычной окраски (Avetisyan, Babosha, 2011). Ингибирование ферментов, которые разрушают перекись водорода, должно приводить к повышению ее содержания в тканях и развитию оксидативного стресса. Учитывая возможное участие механизма взаимной активации синтеза активных форм кислорода и кальциевого сигналинга (Demidchik, 2018; Pottosin, Zepeda-Jazo, 2018), прерывание данного цикла блокатором кальциевых каналов (нитрат лантана) должно вызывать противоположную реакцию, т.е. уменьшение размеров гало, что и наблюдали в наших экспериментах. Неожиданным оказалось то, что обработка каталазой, которая разрушает перекись водорода, также приводила к увеличению размеров гало. Однако необходимо принять во внимание известные данные об активации каталазы кальций-кальмодулиновой системой (Yang, Poovaiah, 2002). Если подобная связка актуальна в исследуемой нами модельной системе, добавление блокатора кальция и вызванное им снижение активности эндогенной каталазы должно уменьшать размеры гало, а добавление самого фермента – увеличивать размеры гало, что соответствует результатам эксперимента. Известно, что экспрессию гена каталазы наблюдали при обработке перекисью водорода (Xing et al., 2008). В этом случае именно этот фермент может быть ответственен за появление гало больших размеров во всех тех вариантах, в которых это происходило, поскольку действие 3-амино-1,2,4-триазола опосредовано перекисью водорода и, следовательно, 3-амино-1,2,4-триазол также способен индуцировать актиоксидантную защиту, включая и увеличение активности каталазы. В согласии с предложенным механизмом возможное уменьшение генерации активных форм кислорода в присутствии дифенилениодониума никак не может увеличивать активность каталазы и, соответственно, размеры гало, что и обнаружено в эксперименте. Можно предположить, что точки приложения ингибиторных свойств бисиндолмалеимида и 3-изобутил-1-метилксантина, далеки от системы регуляции активности каталазы. Их использование не оказало влияния на величину гало. Конечно, это не препятствует возможному их влиянию на другие процессы, к которым чувствительно развитие B. graminis.
Доля контактов растения с инфекционными структурами патогена, которые имели результатом появление питающих структур патогена – гаусторий, является очевидной оценкой величины устойчивости (или восприимчивости). В наших экспериментах все обработки в той или иной мере, достоверно или в виде тенденции снижали число гаусторий по сравнению с контролем. При этом в 2-х вариантах (обработка 3-амино-1,2,4-триазолом и нитратом лантана) с наибольшим по величине снижением частоты гаусторий развития колоний не наблюдали вовсе.
Совокупность приведенных выше литературных данных о развитии B. graminis дает возможность на основании, как правило, косвенных данных полагать, что размеры гало, длина аппрессориальной трубки и такие особенности гало, как сложная структура с дополнительными кольцами, могут быть связаны с протеканием защитных реакций. В наших исследованиях нашла подтверждение только взаимосвязь между размерами гало, ассоциированных с гаусториями, и числом гаусторий. При этом взаимосвязи между протеканием защитных реакций и размерами большого гало, длины аппрессориальной трубки или долей сложных гало с кольцевой зональностью более сложны и требуют более глубоких исследований.
Таким образом, изучение размеров и структуры гало представляет значительный интерес, поскольку они могут нести важную информацию о процессах, в т.ч. защитных реакциях, происходящих непосредственно в зоне контакта растения и патогена. Полученные данные свидетельствуют о потенциальной возможности их использования в качестве новых параметров диагностики состояния патосистемы в тестах на устойчивость в селекционной работе и при поиске соединений с иммуномодулирующей активностью.
Работа выполнена в рамках госзадания ГБС РАН № 118021490111-5.
Список литературы
Avetisyan G.A., Babosha A.V. Role of oxidative stress in pathogenesis of wheat powdery mildew. Byulleten Glavnogo botanicheskogo sada. 2011. V. 196. P. 157–164 (in Russ.).
Babosha A.V., Ryabchenko A.S., Avetisyan T.V. Effect of exogenous cytokinins on dynamics of development and differentiation of infectious structures of the pathogen of wheat powdery mildew. Cell and Tissue Biology. 2009. V. 3 (4). P. 387–396. https://doi.org/10.1134/S1990519X09040117
Beavo J.A., Rogers N.L., Crofford O.B., Hardman J.G., Sutherland E.W., Newman E.V. Effects of xanthine derivatives on lipolysis and on adenosine 3',5'-monophosphate phosphodiesterase activity. Molecular pharmacology. 1970. V. 6 (6). P. 597–603.
Cross A.R., Jones O.T.G. The effect of the inhibitor diphenylene iodonium on the superoxide-generating system of neutrophils. Specific labelling of a component polypeptide of the oxidase. Biochem. J. 1986. V. 237 (1). P. 111–116. https://doi.org/10.1042/bj2370111
Demidchik V. ROS-activated ion channels in plants: Biophysical characteristics, physiological functions and molecular nature. Int. J. Molec. Sci. 2018. V. 19 (4). P. 1263. https://doi.org/10.3390/ijms19041263
Edwards H.H. A basic staining material associated with the penetration process in resistant and susceptible powdery mildewed barley. New Phytol. 1970. V. 69. P. 299–301. https://doi.org/10.1111/j.1469-8137.1970.tb02429.x
Hückelhoven R., Fodor J., Preis C., Kogel K.H. Hypersensitive cell death and papilla formation in barley attacked by the powdery mildew fungus are associated with hydrogen peroxide but not with salicylic acid accumulation. Plant Physiol. 1999. V. 119. P. 1251–1260. https://doi.org/10.1104/pp.119.4.1251
Kunoh H., Ishizaki H., Nakaya K. Cytological studies of early stages of powdery mildew in barley and wheat leaves: (II) significance of the primary germ tube of Erysiphe graminis on barley leaves. Physiol. Plant Pathol. 1977. P. 10. P. 191–199. https://doi.org/10.1016/0048-4059(77)90022-4
Margoliash E., Novogrodsky A. A study of the inhibition of catalase by 3-amino-1:2:4-triazole. Biochem. J. 1958. V. 68 (3). V. 468–475. https://doi.org/10.1042/bj0680468
Mellersh D.G., Heath M.C. Plasma membrane – cell wall adhesion is required for expression of plant defense responses during fungal penetration. Plant Cell. 2001. V. 13. P. 413–424. https://doi.org/10.1105/tpc.13.2.413
Mishina G.N., Serezhkina G.V., Avetisian T.V., Ryabchenko A.S., Andreev L.N. Specific features of the formation of a halo during pathogenesis as a response of cereal epidermal cells to penetration of powdery mildew causative agents. Biology Bulletin. 2001. V. 28. P. 354–360. https://doi.org/10.1023/A:1016666906210
Ouchi S., Oku H., Hibino C. Localization of induced resistance and susceptibility in barley leaves inoculated with the powdery mildew fungus. Phytopathol. 1976. V. 66. P. 901–905. https://doi.org/10.1094/Phyto-66-901
Pantoja O., Gelli A., Blumwald E. Voltage-dependent calcium channels in plant vacuoles. Science. 1992. V. 255 (5051). P. 1567–1570. https://doi.org/10.1126/science.255.5051.1567
Pottosin I., Zepeda-Jazo I. Powering the plasma membrane Ca2+-ROS self-amplifying loop. J. Exp. Bot. 2018. V. 69 (14). P. 3317–3320. https://doi.org/10.1093/jxb/ery179
Sanin S.S., Motovilin A.A., Korneva L.G. Chemical protection of wheat from the diseases under the conditions of intensive grain production. Zashchita i karantin rasteniy. 2011. V. 8. P. 3–10 (in Russ.).
Serezhkina G.V., Mishina G.N., Andreev L.N., Avetisyan T.V., Lapochkina I.F. Cytophysiological characteristics of resistance of Aegilops speltoides Tausch, and disomic addition wheat – Aegilops lines (2N = 44) at the stage of seedlings against powdery mildew. Biology Bulletin 1999. V. 26. P. 344–350.
Shetty N.P., Jørgensen H.J.L., Jensen J.D., Collinge D.B., Shetty H.S. Roles of reactive oxygen species in interactions between plants and pathogens. Eur. J. Plant Pathol. 2008. V. 121. P. 267–280. https://doi.org/10.1007/s10658-008-9302-5
Sugai K., Masaoka H., Penjore K., Hanboonsong Y., Nishiguchi M., Yamaoka N. The time and spatial strategy of Blumeria graminis f. sp. hordei for surviving after failure of first infection. Physiol. Mol. Plant Pathol. 2010. V. 74 (5). P. 346–350. https://doi.org/10.1016/j.pmpp.2010.05.005
Tarchevskiy I.G. Signal systems of plant cells. Moscow, Nauka, 2002 (in Russ.).
Thordal-Christensen H., Zhang Z., Wei Y., Collinge D.B. Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley–powdery mildew interaction. Plant J. 1997. V. 11. V. 1187–1194. https://doi.org/10.1046/j.1365-313X.1997.11061187.x
Toullec D., Pianetti P., Coste H., Bellevergue P., Grand-Perret T., Ajakane M., Baudet V., Boissin P., Boursier E., Loriolle F., Duhamel L., Charon D., Kirilovsky J. The bisindolylmaleimide GF 109203X is a potent and selective inhibitor of protein kinase C. J. Biol. Chem. 1991. V. 266 (24). P. 15771–15781.
Trujillo M., Kogel K.-H., Hückelhoven R. Superoxide and hydrogen peroxide play different roles in the nonhost interaction of barley and wheat with inappropriate formae speciales of Blumeria graminis. Mol. Plant-Microbe Interact. 2004. V. 17. P. 304–312. https://doi.org/10.1094/MPMI.2004.17.3.304
Xing Y., Jia W., Zhang J. AtMKK1 mediates ABA-induced CAT1 expression and H2O2 production via AtMPK6-coupled signaling in Arabidopsis. Plant J. 2008. V. 54. P. 440–451. https://doi.org/10.1111/j.1365-313X.2008.03433.x
Yang T., Poovaiah B.W. Hydrogen peroxide homeostasis: Activation of plant catalase by calcium/calmodulin. Proc. Natl. Acad. Sci. USA. 2002. V. 99. P. 4097–4102. https://doi.org/10.1073/pnas.052564899
Аветисян Г.А., Бабоша А.В. (Avetisyan, Babosha) Роль окислительного стресса в патогенезе возбудителя мучнистой росы пшеницы // Бюллетень ГБС. 2011. Т. 196. С. 157–164.
Бабоша А.В., Рябченко А.С., Аветисян Т.В. (Babosha et al.) Влияние экзогенных цитокининов на динамику развития и дифференциацию инфекционных структур возбудителя мучнистой росы пшеницы // Цитология. 2009. Т. 51(7). С. 602–611.
Мишина Г.Н., Сережкина Г.В., Аветисян Т.В., Рябченко А.С., Андреев Л.Н. (Mishina et al.) Особенности формирования гало в процессе патогенеза как ответная реакция эпидермальных клеток злаков на проникновение возбудителей мучнистой росы // Известия РАН. Сер. биол. 2001. № 4. С. 424–430.
Санин С.С., Мотовилин А.А., Корнева Л.Г., Жохова Т.П., Полякова Т.М., Акимова Е.А. (Sanin et al.) Химическая защита пшеницы от болезней при интенсивном зернопроизводстве // Защита и карантин растений. 2011. № 8. С. 3–10.
Сережкина Г.В., Мишина Г.Н., Андреев Л.Н., Аветисян Т.В., Лапочкина И.Ф. (Serezhkina et al.) Цитофизиологическая характеристика устойчивости к мучнистой росе Aegilops speltoides Tausch. и дисомнодополненных пшенично-эгилопсных линий (2n = 44) в стадии проростков // Известия РАН. Сер. биол. 1999. № 4. С. 417–424.
Тарчевский И.Г. (Tarchevskiy) Сигнальные системы клеток растений. М.: Наука, 2002.
Дополнительные материалы отсутствуют.
Инструменты
Микология и фитопатология