

Микология и фитопатология, 2020, T. 54, № 6, стр. 426-435
Мониторинг агарикоидных подстилочных сапротрофов в сосновых лесах Пермского края
В. С. Боталов 1, *, Л. Г. Переведенцева 2, **
1 Пермский государственный аграрно-технологический университет им. акад. Д.Н. Прянишникова
614000 Пермь, Россия
2 Пермский государственный национальный исследовательский университет
614990 Пермь, Россия
* E-mail: vitalywc@yandex.ru
** E-mail: perevperm@mail.ru
Поступила в редакцию 03.04.2020
После доработки 03.05.2020
Принята к публикации 11.05.2020
Аннотация
Подведены итоги 40-летнего мониторинга биоты агарикоидных базидиомицетов в сосновых лесах Пермского края в подзоне южной тайги. Исследования проводились стационарным методом на учетных площадях размером 50 × 20 м, заложенных по одной в сосняках: лишайниково-вейниковом, брусничном, сфагновом и чернично-сфагновом. Работа осуществлялась в три этапа: I – 1975–1977 гг., II – 1994–1996 гг., III – 2010–2012 гг. В каждый год наблюдений по 4 раза с интервалом в 10 дней собирались все плодовые тела агарикоидных грибов, учитывалось их число и воздушно-сухая биомасса для каждого вида, отбирались образцы для последующей идентификации. К настоящему времени установлено, что число видов подстилочных сапротрофов в сосновых лесах колеблется от 9 (сосняк сфагновый) до 45 (сосняк брусничный). Большинство выявленных видов относится к семейству Tricholomataceae (68.3–88.9% от общего числа видов в разных сосновых лесах). Показано ежегодное накопление выявляемых видов грибов, описываемое уравнением y = 4.283x + 22.3 (R2 = 0.97), причем наибольшее число видов выявлялось только в какой-либо один год наблюдений. Отмечена относительная стабильность видового состава высших сосудистых растений (J = 69–96) во времени и зафиксированы существенные изменения видового состава грибов (J = 0–67). Наиболее близкими по видовому составу грибов оказались биогеоценозы со схожим режимом влажности (JСЧС–ССФ = 53; JСЛВ–СБР = 46), а большее разнообразие агарикоидных подстилочных сапротрофов и большая выровненность численностей их видов установлены для сосняка брусничного (H = 1.13). Изучена связь между разнообразием и продуктивностью подстилочных сапротрофов в августе с основными климатическими факторами. Установлено, что осадки в июле положительно влияют на число базидиом агарикоидных грибов и их суммарную биомассу в сосняках: лишайниково-вейниковом, брусничном и сфагновом (rs ≈ 0.7…0.8; p < 0.05). Влияние климатических показателей на число видов подстилочных сапротрофов, а также температуры воздуха на их “урожайность”, не выявлено.
ВВЕДЕНИЕ
Сосновые леса в пределах района южнотаежных пихтово-еловых лесов Пермского края являются коренными сообществами, встречающимися, главным образом, на песчаных берегах р. Камы. Сукцессионные процессы в данных типах леса протекают медленно, а флора отличается бедным видовым составом (Ovesnov, 2000). Важнейшим гетеротрофным компонентом лесных экосистем являются грибы, выполняющие роль главного регулятора вещественно-энергетических процессов, однако работы, посвященные изучению динамики агарикомицетов в лесных сообществах, немногочисленны (Straatsma el al., 2001; Sidorova et al., 2011; Zmitrovich et al., 2015; Avis et al., 2017; Gange et al., 2018; Karavani et al., 2018), а для Урала и вовсе отсутствуют.
Подстилочные сапротрофы в лесных сообществах выполняют ведущую роль в разложении подстилки и участвуют в формировании гумусового горизонта почвы. Данная группа грибов характеризуется наибольшей вариабельностью видового состава, численности базидиом и их биомассы, так как помимо сложных сочетаний субстрата, их мицелий способен выдерживать резкие колебания температуры и влажности, что приводит к определенным сложностям и недостаточности кратковременных исследований при их изучении.
В связи с этим, для установления закономерностей влияния экологических факторов на гетеротрофный компонент биогеоценозов и выявления скрытого разнообразия агарикоидных базидиомицетов в сообществах, не подверженных антропогенной нагрузке, нами проводятся многолетние стационарные наблюдения. Целью работы является изучение многолетней динамики структуры и продуктивности агарикоидных базидиомицетов, относящихся к группе подстилочных сапротрофов, в 4 типах сосновых лесов Пермского края. Для реализации поставленной цели решались следующие задачи: 1 – выявить видовой состав подстилочных сапротрофов в разных типах сосновых лесов; 2 – установить особенности таксономической структуры изучаемой группы грибов; 3 – проанализировать зависимость плодоношения подстилочных сапротрофов от количества осадков и температуры воздуха.
МАТЕРИАЛЫ И МЕТОДЫ
Район исследований расположен на востоке Восточно-Европейской равнины в подзоне южной тайги (Пермский край, ООПТ “Верхняя Кважва”). В соответствии с ботанико-географическим районированием Пермского края территория относится к району южнотаежных пихтово-еловых лесов (Specially…, 2002). Заметную роль в формировании климата данной территории играет Камское водохранилище, сглаживая суточные и сезонные изменения температуры, а также повышая влажность воздуха в летний период. За год выпадает в среднем 557 мм осадков. Среднегодовая температура воздуха составляет +1.5 °С (Shkliaev, Shkliaeva, 2006).
Исследования проводились стационарным методом в сообществах, не подверженных антропогенной нагрузке. Учетные площади размером 50 × × 20 м были заложены в сосняках: лишайниково-вейниковом, брусничном, сфагновом и чернично-сфагновом (по одной в каждом типе леса). Работа осуществлялась в три этапа: I – 1975–1977 гг., II – 1994–1996 гг., III – 2010–2012 гг. В период наибольшего появления базидиом (август) с интервалом в 10 дней собирались все плодовые тела агарикоидных грибов, учитывалось их число и воздушно-сухая биомасса по каждому виду, отбирались образцы для последующей идентификации. В сентябре отбирались образцы для учета видового состава грибов (одно посещение) (Perevedentseva, 1983, 1999; Perevedentseva, Botalov, 2015).
Принадлежность агарикоидных грибов к эколого-трофическим группам устанавливалась по наблюдениям в природе и литературным данным. За основу взята шкала эколого-трофических групп А.Е. Коваленко (Kovalenko, 1980) с дополнениями (Stolyarskaya, Kovalenko 1996), адаптированная для многолетних исследований Л.Г. Переведенцевой (Perevedentseva, 1999). Названия и объем таксонов (семейств, родов) приведены по системе, принятой M. Moser (1983) с некоторыми изменениями (Perevedentseva, 1999). Геоботанические описания выполнялись согласно В.Н. Сукачеву и Е.В. Зонну (Sukachev, Zonn, 1961).
Степень сходства биогеоценозов по видовому составу грибов и растений установлена с помощью коэффициента Жаккара (J × 100) (Shmidt, 1973). Видовое разнообразие агарикоидных подстилочных сапротрофов вычислялось с помощью индекса Шеннона (H) (Leontyev, 2008).
При оценке влияния погодных условий на видовой состав и урожайность подстилочных сапротрофов использовался коэффициент ранговой корреляции Спирмена (rs) (Trukhacheva, 2012). Статистическая обработка осуществлялась при помощи программ Microsoft Office Excel 2013 и StatSoft Statistica 10. Корреляционный анализ проводился в сумме для всех сосновых лесов, раздельно для каждого сосняка, а также для сообществ, объединенных в две группы в соответствии с особенностями гидротермического режима. Рассматривалось влияние климатических показателей на следующие характеристики подстилочных сапротрофов: число базидиом и их суммарная воздушно-сухая биомасса за август, число видов за август–сентябрь. В качестве климатических показателей использованы данные метеостанции г. Добрянка (1975–2012 гг.) (Meteorological.., 1975–1977, 1994–1996; Weather.., 2012): сумма положительных температур за май–август (°C), средняя месячная температура воздуха за май–сентябрь (°C), месячная сумма осадков (май–сентябрь) (мм), сумма осадков за май–август (мм).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Эколого-ценотическая характеристика исследуемых биогеоценозов
Сосняк брусничный (58°37.97 с.ш.; 56°39.36 в.д.) сформировался на месте выгоревшего соснового леса и располагается на дюнных всхолмлениях третьей боровой террасы реки Кама. Состав древостоя 10С, возраст около 95 лет. Сомкнутость крон 0.6. В подросте Pinus sylvestris L., в подлеске Sorbus aucuparia L. Проективное покрытие травяно-кустарничкового яруса составляет около 50%, преобладают Vaccinium vitis-idaea L. и V. myrtillus L. Моховой покров представлен видами родов Dicranum, Polytrichum, Pleurozium, Hylocomium. Валежника мало. Почва дерново-подзолистая, супесчаная.
Сосняк лишайниково-вейниковый (58°37.58′ с.ш.; 56°40.75′ в.д.) сформировался на месте частично выгоревшего соснового леса и располагается на дюнных всхолмлениях третьей боровой террасы реки Кама. Состав древостоя 9С1Б, возраст около 95 лет. Сомкнутость крон 0.4. В подросте Pinus sylvestris и Larix sibirica Ledeb. Подлесок не выражен. Проективное покрытие травяно-кустарничкового яруса 20%, преобладают Calamagrostis epigeios (L.) Roth и C. arundinacea (L.) Roth. Лишайниковый покров почти сплошной, состоит, в основном, из видов рода Cladonia. Изредка встречаются зеленые мхи: Dicranum sp., Polytrichum sp. Валежника очень мало. Почва дерново-подзолистая, супесчаная.
Сосняк чернично-сфагновый (58°39.37′ с.ш.; 56°36.72′ с.ш.) представляет собой облесенное верховое болото, коренное сообщество, располагается на равнинном плато. Состав древостоя 10С, возраст около 100 лет. Сомкнутость крон 0.4. В подросте встречается Pinus sylvestris. Подлесок не выражен. В кустарниковом ярусе Chamaedaphne calyculata (L.) Moench и Ledum palustre L. Травяно-кустарничковый ярус состоит из Vaccinium vitis-idaea L. и Oxycoccus palustris Pers., встречаются различные виды осок, проективное покрытие около 20%. Моховой покров образован сфагновыми мхами, сплошной. Валежника мало. Почва торфянистая, мощностью более 120 см.
Сосняк сфагновый (58°36.99′ с.ш.; 56°37.32′ в.д.) – облесенное верховое болото, коренное сообщество. Состав древостоя 10С+Б, возраст около 80 лет. Сомкнутость крон 0.4. Подрост состоит из Pinus sylvestris. Подлесок не выражен. В кустарниковом ярусе преобладает Chamaedaphne calyculata. В травяно-кустарничковом ярусе Vaccinium myrtillus L., Eriophorum vaginatum L., и Oxycoccus palustris. Моховой покров состоит из сфагновых мхов, сплошной. Валежника очень мало. Почва торфянистая, мощностью более 120 см.
По периодам исследований в сосновых лесах выявлялось от 12 (сосняки: сфагновый и чернично-сфагновый) до 33 (сосняк брусничный) видов высших сосудистых растений. Их видовой состав со временем оставался относительно стабильным (J = 69–96). Состав древостоя не изменялся. В сосняках: лишайниково-вейниковом и брусничном во II период отмечено увеличение обилия зеленых мхов до 20%. В сосняке чернично-сфагновом во II период зафиксировано усиление болотообразовательного процесса: снижение проективного покрытия травяно-кустарничкового яруса с 50% в I период до 20% во II и III, застаивание воды и увеличение обилия сфагновых мхов.
Аннотированный список видов агарикоидных подстилочных сапротрофов. Видовой состав подстилочных сапротрофов сосновых лесов Пермского края не отличается большим разнообразием; далее приводится его список. Названия и объем семейств и родов, а также латинские названия видов указаны по системе, принятой M. Moser (1983) с некоторыми изменениями (Perevedentseva, 1999). Современные латинские названия видов и сокращения авторов приведены в скобках после названия вида, в соответствии с номенклатурной базой данных “Mycobank” (2020). Римскими цифрами обозначены месяцы вегетационного периода. Арабскими цифрами обозначены периоды выявления вида: 1 – 1975–1977 гг., 2 – 1994–1996 гг., 3 – 2010–2012 гг. Место нахождения: СЛВ – сосняк лишайниково-вейниковый, СБР – сосняк брусничный, ССФ – сосняк сфагновый, СЧС – сосняк чернично-сфагновый.
AGARICALES
Agaricaceae
Cystoderma adnatifolium (Peck) Harmaja – VIII, СБР (3).
C. amianthinum (Scop.) Fayod – VIII–IX, СЛВ (1–3), СБР (1–3).
C. carcharias (Pers.) Fayod – VIII–IX, СЛВ (1).
C. granulosum (Batch) Fayod – VII–VIII, СЛВ (2, 3), СБР (1–3).
C. rugosoreticulatum (F. Lorinser) Wasser [C. amianthinum f. rugosoreticulatum (F. Lorinser) A.H. Sm. et Singer] – VIII–IX, СЛВ (3), СБР (1–3).
C. terryi (Berk. et Broome) Harmaja – VII–VIII, СЛВ (2, 3), СБР (1–3).
Entolomataceae
Entoloma cetratum (Fr.) M.M. Moser – VIII–IX, СБР (1–3), СЧС (1, 2), ССФ (1–3).
E. conferendum (Britzelm.) Noordel. – VIII, СБР (2).
E. griseorubidum Kühner ex Noordel. [E. indutoides var. griseorubidum (Kühner ex Noordel.) Noordel.] – VIII, СЛВ (1).
E. infula (Fr.) Noordel. – VIII, СБР (3).
E. juncinum (Kühner et Romagn.) Noordel. – VIII, СЛВ (2), СЧС (1).
E. minutum (P. Karst.) Noordel. – VIII, СЛВ (2).
E. sericeum Quél. – VI–IX, СЛВ (1–3), СБР (1, 3).
E. turbidum (Fr.) Quél. [= Entocybe turbida (Fr.) T.J. Baroni, V. Hofstetter et Largent] – VIII, СЛВ (2), СБР (2, 3).
E. vernum S. Lundell – V, СЛВ (1).
Tricholomataceae
Baeospora myosura (Fr.) Singer – VIII, СЛВ (3), СБР (2, 3).
Cantharellula umbonata (J.F. Gmel.) Singer – VIII–IX, СЛВ (1–3), СБР (1–3), СЧС (1), ССФ (1, 3).
Clitocybe brumalis (Fr.) Quél. – VIII, СБР (3).
C. candicans (Pers.) P. Kumm. [= Leucocybe candicans (Pers.) Vizzini, P. Alvarado, G. Moreno et Consiglio] – VIII, СЛВ (1–3), СБР (1–3).
C. cerussata (Fr.) P. Kumm. [C. phyllophila (Pers.) P. Kumm.] – VIII–IX, СЛВ (1–3), СБР (1).
C. clavipes (Pers.) P. Kumm. [= Ampulloclitocybe clavipes (Pers.) Redhead, Lutzoni, Moncalvo et Vilgalys] – VIII–IX, СЛВ (1–3), СБР (1–3).
C. dealbata (Sowerby) P. Kumm. – VIII–IX, СБР (1).
C. diatreta (Fr.) P. Kumm. – VIII, СЛВ (3), СБР (3).
C. ericetorum Quél. – VIII–IX, СЛВ (3), СБР (3).
C. gibba (Pers.) P. Kumm. [= Infundibulicybe gibba (Pers.) Harmaja] – VII–IX, СБР (1).
C. metachroa (Fr.) P. Kumm. – VIII–IX, СЛВ (1, 3), СБР (1–3).
C. squamulosa (Pers.) P. Kumm. – VIII, СБР (1).
C. vibecina (Fr.) Quél. – VIII–IX, СЛВ (1–3), СБР (1, 2).
Collybia dryophila (Bull.) P. Kumm. [= Gymnopus dryophilus (Bull.) Murrill] – VII–VIII, СЛВ (3), СБР (1–3).
C. exsculpta (Fr.) Gillet [= Gymnopus ocior (Pers.) Antonín et Noordel.] – VIII–IX, СБР (1–3).
C. maculata (Alb. et Schwein.) P. Kumm. [= Rhodocollybia maculata (Alb. et Schwein.) Singer] – VIII, СЧС (1), ССФ (1, 2).
C. putilla (Fr.) Singer – IX, СБР (3).
C. tergina (Fr.) S. Lundell [= Gymnopus terginus (Fr.) Antonín et Noordel.] – VIII, СЛВ (3).
Marasmius androsaceus (L.) Fr. [= Gymnopus androsaceus (L.) J.L. Mata et R.H. Petersen] – VII–IX, СЛВ (1–3), СБР (1–3), СЧС (1–3), ССФ (1–3).
M. epiphyllus (Pers.) Fr. – VIII, СЧС (1).
M. scorodonius (Fr.) Fr. [= Mycetinis scorodonius (Fr.) A.W. Wilson et Desjardin] – VII–IX, СЛВ (1–3), СБР (1–3).
Micromphale perforans (Hoffm.) Gray [= Paragymnopus perforans (Hoffm.) J.S. Oliveira] – VIII, СЛВ (1–3), СБР (1–3), СЧС (1, 2), ССФ (1–3).
Mycena amicta (Fr.) Quél. – VIII, СЛВ (3).
M. clavicularis (Fr.) Gillet – VIII, СЛВ (3), СБР (3).
M. epipterygia (Scop.) Gray – VI–VIII, СБР (3), СЧС (1).
M. epipterygia var. epipterygioides (A. Pearson) Kühner [M. epipterygioides A. Pearson] – VIII, СБР (3).
M. epipterygia var. viscosa (Maire) Ricken – VIII–IX, СЛВ (3).
M. filopes (Bull.) P. Kumm. – VIII, СЛВ (3), СБР (3).
M. flavoalba (Fr.) Quél. [= Atheniella flavoalba (Fr.) Redhead] – VII–VIII, СБР (2), СЧС (1).
M. galopus (Pers.) P. Kumm. – VI–VIII, СЧС (1, 3), ССФ (1–3).
M. galopus var. nigra Rea – VIII–IX, ССФ (2, 3).
M. metata (Fr.) P. Kumm. – VIII–IX, СЛВ (3).
M. polygramma (Bull.) Gray – VI–VIII, СЛВ (1).
M. psammicola (Berk. et Broome) Sacc. – VIII, СЛВ (2, 3).
M. pura (Pers.) P. Kumm. – VII–IX, СЛВ (3), СБР (1–3).
M. rorida (Fr.) Quél. [= Roridomyces roridus (Fr.) Rexer] – VI–IX, СЛВ (1, 2), СБР (1–3), СЧС (1), ССФ (1).
M. rosella (Fr.) P. Kumm. – VIII, СЛВ (2).
M. simia Kühner – VIII–IX, СЛВ (3), СБР (3).
M. vitilis (Fr.) Quél. – VIII–IX, СБР (3), СЧС (2).
M. vulgaris (Pers.) P. Kumm. – VIII, СЛВ (3).
Strobilurus stephanocystis (Kühner et Romagn. ex Hora) Singer – V–VI, VIII–IX, СЛВ (1), СБР (1), СЧС (1), ССФ (1, 3).
Tephrocybe confusa (P.D. Orton) P.D. Orton [= Lyophyllum confusum (P.D. Orton) Gulden] – VIII, СЛВ (3), СБР (2, 3).
T. rancida (Fr.) Donk [= Lyophyllum rancidum (Fr.) Singer] – VIII, СБР (2).
Xeromphalina cauticinalis (Fr.) Kühner et Maire – IX, СБР (2, 3).
X. cornui (Quél.) J. Favre – VIII–IX, СЧС (1, 2).
X. fellea Maire et Malençon – VIII, СБР (3).
Hygrophoraceae
Gliophorus laetus (Pers.) Herink – VIII, СБР (2).
Таксономический состав агарикоидных базидиомицетов. В сосновых лесах за все время исследований (1975–1977, 1994–1996, 2010–2012 гг.) выявлено 64 вида и внутривидовых таксона агарикоидных грибов, относящихся к группе подстилочных сапротрофов и принадлежащих к 15 родам и 6 семействам (табл. 1).
Таблица 1.
Таксономический состав подстилочных сапротрофов за все время наблюдений (1975–1977, 1994–1996, 2010–2012 гг.)
| Семейство (число родов/число видов) | Род (общее число видов) | В том числе, видов | |||
|---|---|---|---|---|---|
| СЛВ | СБР | ССФ | СЧС | ||
| Agaricaceae (1/6) | Cystoderma (6) | 5 | 5 | – | – |
| Coprinaceae (1/1) | Psathyrella (1) | 1 | – | – | – |
| Entolomataceae (1/9) | Entoloma (9) | 6 | 5 | 1 | 2 |
| Hygrophoraceae (1/1) | Gliophorus (1) | – | 1 | – | – |
| Strophariaceae (1/1) | Pholiota (1) | 1 | 1 | – | – |
| Tricholomataceae (10/46) | Baeospora (1), Cantharellula (1), Clitocybe (11), Collybia (5), Marasmius (3), Micromphale (1), Mycena (18), Strobilurus (1), Tephrocybe (2), Xeromphalina (3) | 28 | 33 | 8 | 12 |
| 6 | 15 (64) | 41 | 45 | 9 | 14 |
Агарикоидные базидиомицеты изучаемых сосновых лесов, относящиеся к группе подстилочных сапротрофов, характерны для подзоны южной тайги, что согласуется с данными, имеющимися для северных широт и лесной зоны Голарктики (Moser, 1983; Funga Nordica, 2012; Zmitrovich et al., 2015). Большинство выявленных видов относится к семейству Tricholomataceae (68.3–88.9% от общего числа видов в разных сосняках). Меньшим числом видов представлены семейства Entolomataceae (11.1–14.6%) и Agaricaceae (11.1–12.2%). Остальные семейства содержат 4.5–4.9% выявленных видов грибов в разных биогеоценозах (рис. 1). В спектре ведущих отмечены роды: Mycena (28.1% от всех видов подстилочных сапротрофов в сосновых лесах), Clitocybe (17.2%), Entoloma (14.1%). Указанные роды представлены 38 видами.
Рис. 1.
Соотношение ведущих семейств подстилочных сапротрофов в исследуемых биогеоценозах за все время наблюдений (в процентах от общего числа видов за 1975–2012 гг. для каждого из ценозов): СЛВ – сосняк лишайниково-вейниковый; СБР – сосняк брусничный; ССФ – сосняк сфагновый; СЧС – сосняк чернично-сфагновый; 1 – Tricholomataceae; 2 – Entolomataceae; 3 – Agaricaceae; 4 – остальные семейства.
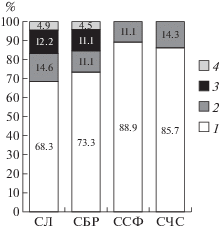
Видовой состав подстилочных сапротрофов зависит от качественной и количественной характеристики подстилки, а также условий увлажнения (Burova, 1986; Perevedentseva, 1999). По годам наблюдений выявлялось 7–89% от общего числа видов подстилочных сапротрофов для разных сосновых лесов (в среднем за год 38.5%). Большинство видов отмечено в сосняках: лишайниково-вейниковом (41 вид) и брусничном (45), в сравнении с переувлажненными биогеоценозами (9–14). Бедность видового состава сапротрофов характерна для сосновых лесов, отличающихся специфичностью субстрата, однообразием его химического состава, а также, значительной суточной амплитудой колебаний температуры и влажности воздуха (Rysin, Savelyeva, 2008). В сосняке чернично-сфагновом выявляемое число видов подстилочных сапротрофов с каждым периодом снижалось, в сосняке сфагновом оставалось стабильным, а в остальных биогеоценозах – увеличивалось.
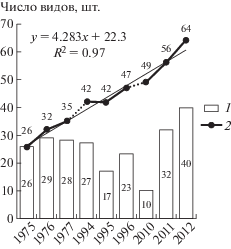
В связи с тем, что скрытое разнообразие грибов не поддается учету, все ранее зафиксированные виды грибов включались в общий список. В каждый сезон исследований для каждого из сосняков выявлялись новые виды подстилочных сапротрофов, то есть происходило увеличение общего выявленного разнообразия грибов как для каждого из биогеоценозов, так и в сумме для сосняков, что подчеркивает несоответствие наблюдаемого и скрытого разнообразия биоты агарикоидных базидиомицетов (Tedersoo et al., 2014). Динамика выявления скрытого разнообразия данной группы грибов описывается уравнением y = 4.283x + 22.3 (R2 = 0.97) (рис. 2). Кривая накопления выявляемого разнообразия грибов сохраняет восходящий тренд, что свидетельствует о необходимости продолжения стационарных наблюдений для наиболее полного выявления структуры грибного компонента данных сообществ.
Рис. 2.
Изменение видового разнообразия агарикоидных подстилочных сапротрофов по годам наблюдений в сумме для всех сосновых лесов: 1 – число видов за год наблюдений; 2 – накопление общего выявленного разнообразия.
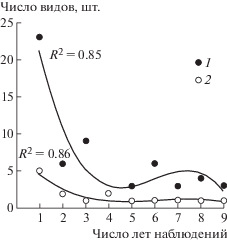
Наибольшее число видов грибов, как для каждого из ценозов, так и в сумме для сосняков, появлялось в ценозе не ежегодно, а выявлялось только один раз – в какой-либо один сезон наблюдений (из 64 видов, таких было 20, что составляет 31.5%). Близкая динамика частоты выявления видов на учетной площади отмечена для ценозов с похожим режимом увлажнения: сосняка лишайниково-вейникового и брусничного, а также для заболоченных сообществ (сосняки: сфагновый и чернично-сфагновый) (рис. 3). Меньше всего видов встречалось ежегодно и одновременно во всех типах леса.
Рис. 3.
Частота выявления видов подстилочных сапротрофов в сосновых лесах по годам наблюдений в зависимости от продолжительности стационарных исследований для биогеоценозов с умеренным режимом влажности (1) и переувлажненных (2).
Общими для всех сосновых лесов являются 5 видов: Marasmius androsaceus, Micromphale perforans, Cantharellula umbonata, Mycena rorida и Strobilurus stephanocystis. Cantharellula umbonata и Mycena rorida массово встречались в сосняке брусничном, а в заболоченных ценозах – единично. Entoloma cetratum выявлена во всех ценозах, за исключением сосняка лишайниково-вейникового. К видам, установленным как в одном из ценозов с умеренным режимом влажности, так и в переувлажненном, относятся Entoloma juncinum (СЛВ, СЧС), Mycena epipterygia (СБР, СЧС), M. flavoalba (СБР, СЧС) и M. vitilis (СБР, СЧС). Зачастую, данные виды в сосняках с умеренным режимом влажности встречались в дождливые годы, а в переувлажненных – в засушливые. Остальные 54 вида выявлены либо в сосняках с близким гидротермическим режимом, либо в каком-либо одном ценозе.
Общими видами для сосняков: лишайниково-вейникового и брусничного, и встречающимися только в этих ценозах, являются 22 вида: Baeospora myosura, Clitocybe candicans, C. cerussata, C. clavipes, C. diatreta, C. ericetorum, C. metachroa, C. vibecina, Collybia dryophila, Cystoderma amianthinum, C. granulosum, C. rugosoreticulatum, C. terryi, Entoloma sericeum, E. turbidum, Marasmius scorodonius, Mycena clavicularis, M. filopes, M. pura, M. simia, Pholiota spumosa, Tephrocybe confusa. В каждый период в сообществах выявляются новые виды, увеличивается суммарное число общих видов и происходит сближение ценозов по этому показателю. Из указанных видов, в I период выявлено 10, во II их список, с учетом ранее выявленных, увеличился до 12 и в III – до 22. Общими видами для заболоченных сосняков, и встречающимися только в этих ценозах, оказались Collybia maculata и Mycena galopus.
Для разных типов леса также были установлены постоянные виды. В сосняке лишайниково-вейниковом ежегодно выявляли Marasmius androsaceus. Кроме того, в данном ценозе 9 видов отмечено не ежегодно, но в каждый период наблюдений: Cantharellula umbonata, Cystoderma amianthinum, Clitocybe candicans, C. clavipes, C. cerussata, C. vibecina, Entoloma sericeum, Marasmius scorodonius, Micromphale perforans. Постоянные виды данного биогеоценоза были приурочены, главным образом, к моховым парцеллам.
Для сосняка брусничного, в сравнении с другими ценозами, характерно наличие подлеска из Sorbus aucuparia, большее богатство видового состава растений травяно-кустарничкового яруса и наибольшее обилие зеленых мхов, что обусловливает сложность и разнообразие субстрата, а также относительное постоянство гидротермического режима подстилки и верхнего слоя почвы и, как следствие, благоприятствует развитию подстилочных сапротрофов. В данном типе леса ежегодно выявляли такие виды как: Cantharellula umbonata, Marasmius androsaceus и Mycena rorida. В каждый период, помимо указанных видов, отмечены еще 14: Cystoderma amianthinum, C. granulosum, C. rugosoreticulatum, C. terryi, Clitocybe candicans, C. clavipes, C. metachroa, Collybia dryophila, C. exsculpta, Entoloma cetratum, Marasmius scorodonius, Micro-mphale perforans, Mycena pura, Pholiota spumosa.
В заболоченных ценозах зафиксировано немного видов подстилочных сапротрофов. В сосняке сфагновом ежегодно встречался Marasmius androsaceus. Из постоянных и выявляемых в каждый период наблюдений видов отмечены: Entoloma cetratum, Micromphale perforans и Mycena galopus. Для сосняка чернично-сфагнового ежегодно встречающимся видом также является Marasmius androsaceus.
По периодам наблюдений, в каждом из ценозов, происходило изменение видового состава грибов. Наиболее стабильным видовой состав подстилочных сапротрофов оказался в сосняках: сфагновом (J = 56–67) и брусничном (J = 44–58). Более существенны изменения в сосняке лишайниково-вейниковом (J = 34–42) и особенно – в сосняке чернично-сфагновом (J = 0–29). Виды, характерные для ценозов с умеренным режимом влажности или переувлажненных, в разные годы наблюдений встречаются и в других сосняках. Наибольшее сходство по видовому составу подстилочных сапротрофов установлено между сосняками: сфагновым и чернично-сфагновым (J = 53), лишайниково-вейниковым и брусничным (J = 46).
По видовому составу грибов значения индекса Шеннона для сосняка брусничного составили 1.13, для сосняка лишайниково-вейникового – 0.57, для сосняка сфагнового – 0.43 и для сосняка чернично-сфагнового – 0.71. Следовательно, для сосняка брусничного, в сравнении с другими ценозами, характерно несколько большее разнообразие агарикоидных подстилочных сапротрофов, большая выровненность численностей их видов и, в целом, условия данного ценоза более благоприятны для развития подстилочных сапротрофов.
За время исследований (1975–1977, 1994–1996, 2010–2012 гг.), по данным метеостанции г. Добрянка, отмечен повышенный (относительно нормы) фон температур воздуха и значительные колебания количества осадков. Самыми неблагоприятными для развития грибов оказались наиболее засушливые 1975 г. (количество осадков за июль – август ниже нормы на 26–31 мм, а среднемесячная температура воздуха ниже нормы на 0.56–0.65°C) и 2010 г. (количество осадков в июле ниже нормы на 64 мм, в августе все количество осадков выпало в третью декаду; среднемесячная температура воздуха в июле–августе выше нормы на 2.0–3.2°C), а также 1994 г. как наиболее холодный и дождливый (количество осадков за июль–август выше нормы на 27–83 мм, среднемесячная температура воздуха ниже нормы на 1.0–2.6°C).
В исследуемых сосновых лесах по периодам наблюдений установлено разное число базидиом и их биомасса (в августе). Наибольшие “урожаи” подстилочных сапротрофов по числу базидиом и их воздушно-сухой биомассе выявлены в сосняке брусничном (2001 шт./га; 2.4 кг/га), а наименьшие – в сосняке чернично-сфагновом (13 шт./га; 0.01 кг/га). Варьирование “урожаев” по обоим показателям оказалось близким для сосняков со схожим гидротермическим режимом: лишайниково-вейникового и брусничного, сфагнового и чернично-сфагнового.
Для сосняков с умеренным режимом влажности установлено не только наибольшее число видов подстилочных сапротрофов, но и максимальные колебания их численности по годам наблюдений. На изменение видового состава подстилочных сапротрофов сосняка лишайниково-вейникового, как наиболее ксерофитного биогеоценоза, в некоторой степени влияют интенсивность и количество осадков, определяющие влажность субстрата. В сосняке сфагновом выявлялось немного видов по годам и периодам наблюдений, кроме того, их видовой состав оставался наиболее стабильным, что связано со средообразующей ролью сфагновых мхов. В сосняке чернично-сфагновом на изменение видового состава подстилочных сапротрофов значительно повлияло усиление болотообразовательного процесса ко II периоду наблюдений, что привело к снижению количества доступного субстрата и его консервации. Многие виды грибов в таких условиях существовать не могут (Burova, 1986; Perevedentseva, 1999; Wieder, Vitt, 2006).
На плодоношение агарикоидных грибов в значительной степени влияют погодные условия текущего года. Установленные зависимости оказались для факторов, предшествующих учетам грибов за один месяц, так как реагирует на них мицелий путем формирования базидиом (Straatsma el al., 2001; Avis et al., 2017). Так, с увеличением количества осадков в июле, установлено повышение общего числа базидиом подстилочных сапротрофов и их суммарной биомассы в августе в сосняках: лишайниково-вейниковом и брусничном (rs ≈ 0.7; p < 0.05), а также сосняке сфагновом (rs ≈ ≈ 0.8; p < 0.05) (рис. 4). В рамках проводимого исследования, значимых коэффициентов корреляции между средней месячной температурой воздуха и плодоношением подстилочных сапротрофов не установлено. Влияние климатических показателей на число появляющихся видов подстилочных сапротрофов текущего года также не выявлено, что согласуется с литературными данными о главенствующей роли особенностей субстрата в видовом разнообразии данной группы грибов (Burova, 1986; Perevedentseva, 1999).
Рис. 4.
Диаграммы рассеяния суммы осадков за июль и характеристик подстилочных сапротрофов за август: числа базидиом, шт./га (1) и их воздушно-сухой биомассы, г/га (2) в сосняке лишайниково-вейниковом (rs1 = 0.72, rs2 = 0.77; p < 0.05) и сосняке брусничном (rs1 = 0.72, rs2 = 0.77; p < 0.05).
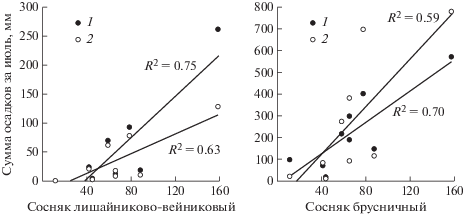
Исследуемые ценозы были объединены в две группы, в соответствии с особенностями гидротермического режима. Для сосняков: лишайниково-вейникового и брусничного число и суммарная биомасса базидиом подстилочных сапротрофов положительно коррелируют с суммой осадков за июль (rs ≈ 0.5; p < 0.05). Для заболоченных ценозов (сосняки: сфагновый и чернично-сфагновый) статистически значимых коэффициентов корреляции изучаемых характеристик подстилочных сапротрофов со средними месячными температурами воздуха и количеством осадков не установлено.
ЗАКЛЮЧЕНИЕ
В результате многолетнего мониторинга биоты агарикоидных грибов в сосновых лесах Пермского края отмечено 64 вида и внутривидовых таксона агарикоидных базидиомицетов, относящихся к группе подстилочных сапротрофов и принадлежащих к 15 родам и 6 семействам. Во всех ценозах наиболее разнообразным и богатым в видовом отношении является семейство Tricholomataceae (68.3–88.9% от общего числа видов в разных сосняках). Наиболее стабильным за все время наблюдений оказался видовой состав подстилочных сапротрофов в сосняках: сфагновом (J = 56–67) и брусничном (J = 44–58). Значительные изменения отмечены в сосняке лишайниково-вейниковом (J = 34–42) и особенно – в сосняке чернично-сфагновом (J = 0–29). Наиболее близкими по видовому составу подстилочных сапротрофов оказались биогеоценозы со схожим режимом влажности (JСЧС–ССФ = 53; JСЛВ–СБР = 46), а большее разнообразие агарикоидных подстилочных сапротрофов и большая выровненность численностей их видов установлены для сосняка брусничного (HСБР = 1.13; HСЛВ = 0.57; HССФ = 0.43; HСЧС = 0.71). Кривая накопления выявляемого разнообразия грибов сохраняет восходящий тренд и описывается уравнением y = 4.283x + 22.3 (R2 = = 0.97).
Продуктивность подстилочных сапротрофов в сосновых лесах различается по годам наблюдений и зависит от изменения погодных условий текущего года. Установлено положительное влияние осадков в июле на плодоношение агарикомицетов в августе. Для сосняков: лишайниково-вейникового, брусничного и сфагнового обильные осадки в июле способствуют увеличению числа базидиом агарикоидных грибов и их суммарной биомассы в августе (rs ≈ 0.7…0.8; р < 0.05). Влияния климатических показателей на число появляющихся видов подстилочных сапротрофов текущего года, а также температуры воздуха на число видов и их урожайность не выявлено.
Список литературы
Avis P.G., Gaswick W.C., Tonkovich G.S. et al. Monitoring fungi in ecological restorations of coastal Indiana, U.S.A. Restoration Ecology. 2017. V. 25 (1). P. 92–100.
Burova L.G. Ecology of macromycetes fungi. Moscow, Nauka, 1986 (in Russ.).
Funga Nordica. Edit. H. Knudsen et J. Vesterholt. Copenhagen, Nordsvamp, 2012. V. 1, 2.
Gange A.C., Heegaard E., Boddy L. et al. Trait-dependent distributional shifts in fruiting of common British fungi. Ecography. 2018. V. 41. I. 1. P. 51–61. https://doi.org/10.1111/ecog.03233
Karavani A., de-Miguel C., Martinez de Aragon J. et al. Effect of climatic and soil moisture conditions on mushroom productivity and related ecosystem services in Mediterranean pine stands facing climate change. Agricultural and Forest Meteorology. 2018. V. 248. P. 432–440. https://doi.org/10.1016/j.agrformet.2017.10.024
Kovalenko A.E. Ecological survey of the fungi of the orders Polyporales s. str., Boletales, Agaricales s. str., Russulales in the mountain forests of the Central North-West Caucasus. Mikologiya i fitopatologiya. 1980. V. 14 (4). P. 300–314 (in Russ.).
Leontyev D.V. Floristic analysis in mycology. Kharkiv, PP Ranok-NT, 2008 (in Russ.).
Meteorological monthly. Uralskoe UGMS, Sverdlovsk. 1975–1977, 1994–1996, I. 9 (in Russ.).
Moser M. Die Rohrlinge und Blätterpilze (Polyporales, Boletales, Agaricales, Russulales). Kleine Kryptogamenflora. Bd. 2b/2. Stuttgart, New York, Fischer, 1983.
MycoBank Database. http://www.mycobank.org. Accessed 15.08.2020.
Ovesnov S.A. Botanical-geographical zoning of the Perm region. Vestnik Permskogo universiteta. Biology. 2000. I. 2. P. 13–21 (in Russ.).
Perevedenceva L.G. Ground macromycetes in some types of pine forests of the Kama region. In: Mycorrhiza and other forms of consorive relationships in the nature. Perm, 1983, pp. 63–67 (in Russ.).
Perevedentseva L.G. Biota and ecology of agaricoid basidiomycetes of Perm region. Dr. Sci. Thesis. Moscow, 1999 (in Russ.).
Perevedentseva L.G., Botalov V.S. Monitoring of the species composition of agarics in some types of pine forests of the Perm territory (southern taiga subzone). Problems of forest phytopathology and mycology. Minsk, 2015, pp. 163–166 (in Russ.).
Rysin L.P., Savelyeva L.I. Pine forests of Russia. Moscow, KMK, 2008 (in Russ.).
Specially protected natural territories of the Perm Region. Edit. S.A. Ovesnov. Knizhnyy mir, Perm, 2002 (in Russ.).
Shkliaev V.A., Shkliaeva L.S. Climatic resources of the Ural Kama Region. Geograficheskij vestnik. 2006. N 2. P. 97–100 (in Russ.).
Shmidt V.M. Mathematical Methods in Botany. Leningrad, Nauka, 1973 (in Russ.).
Sidorova I.I., Voronina E.Yu., Aleksandrova A.V. Long-term dynamics of species diversity of agaricomycetes in the Moscow Region. In: The role of botanical gardens and protected nature areas in investigation and protection of biodiversity of plants and fungi. Yaroslavl, 2011, pp. 152–161 (in Russ.).
Stolyarskaya M.V., Kovalenko A.E. Mushrooms of the Nizhnesvirsky reserve. I. 1. Macromycetes: annotated lists of species. Saint Petersburg, NGPZ, 1996 (in Russ.).
Straatsma G., Ayer F., Egli S. Species richness, abundance, and phenology of fungal fruit bodies over 21 years in a Swiss forest plot. Mycol. Res. 2001. V. 105 (5). P. 515–523. https://doi.org/10.1017/S0953756201004154
Sukachev V.N., Zonn E.V. Methodical instructions for studying of forest types. 2 ed. Moscow, Izdatelstvo AN SSSR, 1961 (in Russ.).
Tedersoo L., Bahram M., Põlme S. et al. Global diversity and geography of soil fungi. Science. 2014. V. 346 (6213). P. 1256688. https://doi.org/10.1126/science.1256688
Trukhacheva N.V. Mathematical statistics in biomedical research using the Statistica package. Moscow, GEOTAR-Media, 2012 (in Russ.).
Weather and climate. Reference and information portal. 2012. http://www.pogodaiklimat.ru. Accessed 01.12.2012 (in Russ.).
Wieder R.K., Vitt D.H. Boreal Peatland Ecosystems. Berlin, Springer, 2006.
Zmitrovich I.V., Stolyarskaya M.V., Kalinovskaya N.I. et al. Macromycetes of Nizhne-Svirsky Reserve (annotated checklist). Saint Petersburg, Svoyo Izdatelstvo, 2015 (in Russ.).
Бурова Л.Г. (Burova) Экология грибов макромицетов. М.: Наука, 1986. 224 с.
Змитрович И.В., Столярская М.В., Калиновская Н.И., Попов Е.С., Мясников А.Г., Морозова О.В., Волобуев С.В., Большаков С.Ю., Светашева Т.Ю., Бондарцева М.А., Коваленко А.Е. (Zmitrovich et al.) Макромицеты Нижне-Свирского заповедника (аннотированный список видов). СПб.: ООО Свое издательство, 2015. 185 с.
Коваленко А.Е. (Kovalenko) Экологический обзор грибов из порядков Polyporales s. str., Boletales, Agaricales s. str., Russulales в горных лесах Центральной части Северо-Западного Кавказа // Микология и фитопатология. 1980. Т. 14. № 4. С. 300–314.
Леонтьев Д.В. (Leontyev) Флористический анализ в микологии. Харьков: ПП Ранок-НТ, 2008. 110 с.
Метеорологический ежемесячник (Meteorological). Уральское УГМС. Свердловск, 1975–1977, 1994–1996. Вып. 9.
Овеснов С.А. (Ovesnov) Ботанико-географическое районирование Пермской области // Вестник Пермского университета. 2000. № 2. Биология. С. 13–21.
Особо охраняемые природные территории Пермской области (Specially…): Реестр / Отв. ред. С.А. Овеснов. Пермь: Книжный мир, 2002. 464 с.
Переведенцева Л.Г. (Perevedentseva) Напочвенные макромицеты в некоторых типах сосновых лесов Прикамья // Микориза и другие формы консортивных связей в природе. 1983. С. 63–67.
Переведенцева Л.Г. (Perevedentseva) Биота и экология агарикоидных базидиомицетов Пермской области. Автореф. дис. … докт. биол. наук. М., 1999. 48 с.
Переведенцева Л.Г., Боталов В.С. (Perevedentseva, Botalov) Мониторинг видового состава агарикоидных базидиомицетов в некоторых типах сосновых лесов Пермского края (подзона южной тайги) // Проблемы лесной фитопатологии и микологии: материалы 9-й Международной конференции. Минск, 2015. С. 163–166.
Погода и климат (Weather…). Справочно-информационный портал. 2012. http://www.pogodaiklimat.ru. Дата обращения: 01.12.2012.
Рысин Л.П., Савельева Л.И. (Rysin, Savelyeva) Сосновые леса России. М.: КМК, 2008. 289 с.
Сидорова И.И., Воронина Е.Ю., Александрова А.В. (Sidorova et al.) Многолетняя динамика видового разнообразия агарикомицетов Московской области // Роль ботанических садов и охраняемых природных территорий в изучении и сохранении разнообразия растений и грибов: материалы Всероссийской научной конференции с Международным участием. Ярославль, 2011. С. 152–161.
Столярская М.В., Коваленко А.Е. (Stolyarskaya, Kovalenko) Грибы Нижнесвирского заповедника. Вып. 1. Макромицеты: аннотированные списки видов. СПб.: НГПЗ, 1996. 59 с.
Сукачев В.Н., Зонн Е.В. (Sukachev, Zonn) Методические указания к изучению типов леса. 2-е изд. М.: Изд-во АН СССР, 1961. 144 с.
Трухачева Н.В. (Trukhacheva) Математическая статистика в медико-биологических исследованиях с применением пакета Statistica. М.: ГЭОТАР-Медиа, 2012. 384 с.
Шкляев В.А., Шкляева Л.С. (Shkliaev, Shkliaeva) Климатические ресурсы Уральского Прикамья // Географический вестник. 2006. №. 2. С. 97–100.
Шмидт В.М. (Shmidt) Математические методы в ботанике. Л.: Наука, 1973. 263 с.
Дополнительные материалы отсутствуют.
Инструменты
Микология и фитопатология