

Микология и фитопатология, 2021, T. 55, № 2, стр. 138-147
Видовой состав и способность агарикомицетов к аккумуляции тяжелых металлов в условиях урбоэкосистемы
Д. В. Попыванов 1, *, А. А. Широких 1, **
1 Федеральный аграрный научный центр Северо-Востока им. Н.В. Рудницкого
610007 Киров, Россия
* E-mail: 1fast@mail.ru
** E-mail: aleshirokikh@yandex.ru
Поступила в редакцию 31.03.2020
После доработки 15.04.2020
Принята к публикации 11.05.2020
Аннотация
Впервые составлены видовые списки и охарактеризована эколого-трофическая структура агарикомикоты парково-рекреационных зон г. Кирова и прилегающего к городу природного лесного массива. Установлено, что наибольшей частотой встречаемости в городских парках и скверах характеризуются агарикомицеты ксилотрофых видов, в то время как в лесном массиве подзоны южной тайги виды из разных эколого-трофических групп представлены практически в равном соотношении. Впервые получены сведения о биосорбции Cu, Pb, Zn в базидиомах грибов, собранных в 6 различных экотопах на территории г. Кирова. Установлено, что в условиях урбоэкосистемы наиболее высокое накопление грибами Zn и Pb, в разрезе отдельных эколого-трофических групп, характерно для микоризообразователей, а накопление Cu – для подстилочных сапротрофов.
ВВЕДЕНИЕ
Города, являющиеся крупными промышленными центрами, включают в себя, как правило, достаточное количество рекреационных территорий, которые выполняют культурно-оздоровительные, санитарно-гигиенические, эстетические, почвозащитные и водоохранные функции. В то же время, городская среда испытывает существенную нагрузку от промышленных предприятий, теплоэнергетического комплекса и ежегодно растущего числа автотранспорта.
Возрастающая антропогенная нагрузка на урбоэкосистемы приводит к снижению их устойчивости и биоразнообразия. В связи с этими неутешительными тенденциями остро встает вопрос о способах оценки отдельных экосистемных компонентов с целью принятия управленческих решений для долгосрочного рационального природопользования.
Одними из наиболее опасных в городской среде загрязняющих веществ являются соединения тяжелых металлов (ТМ) по причине их высокой токсичности, подвижности и способности к биоаккумуляции (de Miguel et al., 1997; Dabakhov et al., 2005; Korolev, Boev, 2017; Hussain et al., 2019). В г. Кирове 17 предприятий имеют гальванические цеха (Ashikhmina, 2012), в связи с чем Cu, Zn и Pb являются приоритетными загрязнителями территории города. Способностью обильно накапливать данные соединения характеризуются базидиальные макромицеты (Churakov et al., 2000; Tsvetnova et al., 2001; Shcheglov, Tsvetnova, 2002; Ivanov, Kostychev, 2007; Kostychev, 2009; Otnyukova et al., 2012; Anishchenko et al., 2016), в силу чего они могут представлять интерес для экологического мониторинга состояния среды.
Изучению сообществ агарикомицетов на территории парково-рекреационных зон промышленных городов уделено значительно меньшее внимание, чем на ненарушенных природных территориях. В литературе присутствуют лишь отрывочные сведения о видовом разнообразии агарикомицетов в городской среде (Dremova, 2014; Shilkova, 2015; Palamarchuk, Kirillov, 2017; Savelyev, Kikeeva, 2018), сведений о видах, аккумулирующих ТМ в условиях города, тоже недостаточно (Demirbaş, 2001; Yamaç et al., 2007; Ivanov et al., 2008; Kostychev, 2012; Širić et al., 2017; Abulude, Ndamitso, 2018; Kokkoris et al., 2019).
Содержание ТМ в плодовых телах агарикомицетов, равно как и структура агарикомикоты, ранее на территории г. Кирова практически не изучались.
Целью данной работы являлось выявление особенностей биоаккумуляции тяжелых металлов представителями различных эколого-трофических групп базидиомицетов в условиях городской среды (на примере парково-рекреационных зон г. Кирова), а также определение видового состава агарикомикоты.
МАТЕРИАЛЫ И МЕТОДЫ
Места сбора плодовых тел агарикомицетов, а также отбора почвы и древесины были привязаны к паркам и скверам города, как наиболее характерным местообитаниям грибов в городских условиях. Обследована территория транспортной, промышленной и рекреационной зон г. Кирова, представленная в разной степени преобразованными аналогами естественных дерново-подзолистых почв (Albic retisols), общей площадью 79.8 га (рис. 1). В составе древостоя городских экотопов в основном присутствуют лиственные породы: Acer platanoides, Acer negundo, Tilia cordata, Quercus robur, Betula pendula. В качестве фоновой выбрана территория лесного массива вблизи п. Порошино, на противоположном городу берегу р. Вятки, где древостой представлен хвойными породами: Picea abies, Abies sibirica. Сбор природного материала проводился в осенний период, с сентября по октябрь. Сбор плодовых тел осуществляли в течение трех лет с 2015 по 2017 гг. В общей сложности на территории г. Кирова и фоновой территории было собрано и обработано 145 образцов агарикомицетов, 51 образец почвы, 24 образца древесины, каждый образец – в трехкратной повторности.
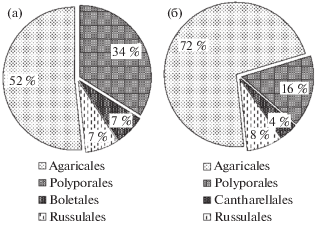
Рис. 1.
Соотношение представителей различных порядков в парках и скверах г. Кирова (а) и в загородном лесу фоновой территории пос. Порошино (б).
Сбор образцов плодовых тел грибов осуществляли маршрутным методом, описание и фиксацию материала проводили по стандартной методике (Bondartsev, Singer, 1950). Морфологические признаки изучали с помощью микроскопа Leica DM 2500 (Германия), с использованием стандартного набора реактивов. При идентификации видов макромицетов были использованы определители по различным таксонам агарикомицетов (Bondartseva, 1998; Nunez, Ryvarden, 2000; Niemelä, 2001; Лессо, 2003; Knudsen, Vesterholt, 2008; Kibby, 2009). Название обнаруженных видов грибов приведены в соответствии с базой данных Index Fungorum (2020).
Собранные плодовые тела грибов высушивали при 70°С до постоянного веса и размалывали до однородной массы с помощью лабораторной мельницы. Полученные образцы хранили в zip-пакетах для предотвращения набора влаги. Пробы древесины отбирали при помощи сверла и аккумуляторной дрели, путем высверливания керна на глубину до 7 см. Пробы почвы отбирали из поверхностного слоя 0–5 см методом конверта с площади 100 м2. Навески каждого из субстратов массой 1 г для озоления помещали в муфельную печь при 450°С. Валовое содержание Cu, Zn, Pb в плодовых телах грибов и древесине определяли на атомно-абсорбционном спектрометре Shimadzu-AA-6800 с предварительным растворением зольного остатка в 1 мл 1 М НСl (Otmahov et al., 2004). Пробы почвы анализировали тоже методом атомно-абсорбционной спектрометрии, предварительно экстрагируя воздушно-сухие образцы аммонийно-ацетатным буфером (pH 4.8) (Vorobyova, 2006).
Содержание ТМ в мицелии определяли атомно-абсорбционным методом после озоления в муфельной печи с последующим растворением зольного остатка в 1М HCl. Учет и анализ данных вели при помощи электронного каталога, построенного на основе программы Microsoft Excel. Рассчитывали коэффициенты накопления (Кн) как отношение концентрации ТМ в базидиомах грибов к концентрации его в субстрате (почве или древесине). Кн позволяет определить способность к накоплению ТМ, выявить виды-биоконцентраторы ТМ (при Кн > 1). Частоту встречаемости грибов (%) определяли как отношение числа экотопов, в которых гриб обнаружен к числу всех обследованных экотопов. Для определения степени сходства микобиоты различных экотопов использовали индекс Сёренсена–Чекановского (Ks), рассчитываемый по формуле K = 2c/(a + b), где а и b – число видов, обнаруженных в каждом из сравниваемых экотопов, с – число общих для них видов.
Статистическая обработка экспериментальных данных была выполнена с использованием пакета прикладных программ Microsoft Excel 2007 и Statistica 12.0. При обработке полученных данных применяли одномерный анализ вариационных рядов (средние величины признака и их ошибки) (Lakin, 1990).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
На территории парково-рекреационных зон г. Кирова обнаружено 56 видов агарикомицетов, принадлежащих к 4 порядкам и 25 семействам (табл. 1). На фоновой территории (пос. Порошино) было обнаружено 25 видов агарикомицетов, принадлежащих к четырем порядкам и 15 семействам (рис. 1) (Ainsworth, 2008).
Таблица 1.
Сравнительный анализ таксономического состава агарикомицетов в различных экотопах г. Кирова и фоновой территории пос. Порошино
| Сравниваемый показатель, количество | Экотопы города | Суммарно в городе | Фон п. Порошино | |||||
|---|---|---|---|---|---|---|---|---|
| сквер комбината ИСКОЖ | парк им. Гагарина | парк Победы | Александров-ский сад | аллея на площади Лепсе | парк им. Кирова | |||
| Порядки | 4 | 2 | 4 | 3 | 4 | 4 | 4 | 4 |
| Семейства | 8 | 9 | 12 | 13 | 8 | 15 | 25 | 15 |
| Роды | 10 | 13 | 17 | 17 | 8 | 20 | 41 | 22 |
| Виды | 10 | 13 | 20 | 20 | 8 | 21 | 56 | 25 |
Среди всех обнаруженных видов (73 вида агарикомицетов) только 8 видов встречались одновременно в городских экотопах и фоновой территории (лесного массива пос. Порошино). Наибольшее количество общих видов принадлежит порядку Agaricales [Agaricus bisporus (J.E. Lange) Imbach, Flammulina velutipes (Curtis) Singer, Gymnopus dryophilus (Bull.) Murrill, Hebeloma mesophaeum (Pers.) Quél., Megacollybia platyphylla (Pers.) Kotl. et Pouzar, Pholiota aurivella (Batsch) P. Kumm., Tricholoma terreum (Schaeff.) P. Kumm.]. Из представителей порядка Polyporales общим был вид Daedaleopsis confragosa (Bolton) J. Schröt. Биота агарикомицетов в городе характеризуется более богатым видовым разнообразием, чем биота фоновой территории. С наибольшей частотой в городе встречались микоризообразующие виды Tricholoma terreum (100%), Paxillus involutus (Batsch) Fr. (66%) и ксилотроф Bjerkandera adusta (Willd.) P. Karst. (83%).
Для оценки степени сходства между микобиотой в различных экотопах использовали индекс Сёренсена–Чекановского (Ks). Установлена высокая специфичность таксономического состава агарикомицетов в каждом из экотопов. Кs имели низкие значения, изменяясь от 0.13 до 0.39 (табл. 2).
Таблица 2.
Матрица степени сходства (Кs) видового состава агарикомицетов в различных экотопах города и базидиомикотой фоновой территории
| Сквер комбината ИСКОЖ | Парк им. Гагарина | Парк Победы | Александров-ский сад | Аллея на площади Лепсе | Парк им. Кирова | Пос. Порошино (ФОН) | |
|---|---|---|---|---|---|---|---|
| Сквер комбината ИСКОЖ | 0.19 | 0.20 | 0.13 | 0.22 | 0.39 | 0.11 | |
| Парк им. Гагарина | 0.19 | 0.24 | 0.24 | 0.19 | 0.35 | 0.11 | |
| Парк Победы | 0.20 | 0.24 | 0.30 | 0.29 | 0.29 | 0.13 | |
| Александровский сад | 0.13 | 0.24 | 0.30 | 0.29 | 0.34 | 0.18 | |
| Аллея на площади Лепсе | 0.22 | 0.19 | 0.29 | 0.29 | 0.28 | 0.12 | |
| Парк им. Кирова | 0.39 | 0.35 | 0.29 | 0.34 | 0.28 | 0.22 | |
| Пос. Порошино (ФОН) | 0.11 | 0.11 | 0.13 | 0.18 | 0.12 | 0.22 |
Низкое сходство может объясняться разным возрастом древостоев в обследованных парках и скверах, который варьирует от 50 лет в парке им. С.М. Кирова до 200 лет в Александровском саду. Максимальные различия выявлены между сообществами агарикомицетов фоновой территории и агарикомикотой парка им. Ю.А. Гагарина (Ks = 0.11), а также сквера комбината ИСКОЖ (Ks = 0.11).
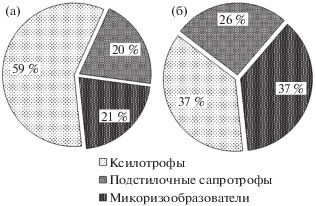
Согласно классификации, предложенной А.Е. Коваленко (Kovalenko, 1980) с некоторыми дополнениями О.В. Морозовой (Morozova, 2001) выявленные виды относятся к следующим трофическим группам: Le – ксилотрофы, Mr – симбиотрофы, St – подстилочные сапротрофы. Наиболее многочисленной по количеству видов в городской среде была группа ксилотрофов (59%) (рис. 2).
Рис. 2.
Соотношение видов из разных эколого-трофических групп в парках и скверах г. Кирова (а) и в загородном лесу фоновой территории пос. Порошино (б).
Большее распространение ксилотрофов объясняется наиболее благоприятными условиями для их произрастания: возрастные насаждения лиственных пород, ослабленные антропогенной нагрузкой, позволяют ксилотрофам без труда колонизировать доступный субстрат. Кроме того, для многих ксилотрофных грибов характерна субстратная специализация, которая выражается в их приуроченности к древесным остаткам определенных видов деревьев.
Виды-симбиотрофы доминируют в хвойных и хвойно-широколиственных лесах, образуя микоризу с древесными доминантами, что также отмечено другими авторами (Burova, 1986; Malysheva, Malysheva, 2008).
Листовой опад, являющийся субстратом для подстилочных сапротрофов, в городах удаляется коммунальными службами, в связи с чем данная группа представлена в городах небольшим количеством видов. В естественном лесу фоновой территории распределение видов по эколого-трофическим группам, напротив − относительно равномерное. Значительное количество симбиотрофных грибов в таежных лесах определяется разнообразием микотрофных древесных пород (ель, сосна, пихта, лиственница, береза, осина и др.), а также смещенными к пессимуму условиями их произрастания.
Изученные виды грибов принимают участие в биологической аккумуляции тяжелых металлов в условиях городских зеленых насаждений. Определение валового содержания Cu, Zn и Pb в плодовых телах грибов, собранных на территории парково-рекреационных зон, показало, что величина сорбции варьирует в широких пределах в зависимости от экотопа, вида гриба и от природы металла.
Наиболее высоким содержанием ТМ (317.9 мкг/г) отличались плодовые тела грибов-микоризообразователей, собранных в промышленной зоне (аллея на пл. Лепсе) (табл. 3).
Таблица 3.
Средние значения суммарного накопления ТМ (мкг/г) в плодовых телах грибов из разных функциональных зон
| Экотоп | Эколого-трофические группы | ||
|---|---|---|---|
| ксилотрофы | микоризообразователи | подстилочные сапротрофы | |
| Промышленная зона | |||
| Аллея на пл. Лепсе | 118.7 | 317.9 | н/о |
| Парк Победы | 108.0 | 238.7 | 151.5 |
| Рекреационная зона | |||
| Александровский сад | 105.0 | 233.0 | 112.2 |
| Парк им. Гагарина | 95.5 | 71.7 | 143.0 |
| Транспортная зона | |||
| Сквер комб. ИСКОЖ | 269.7 | 281.4 | 303.8 |
| Парк им. Кирова | 111.5 | 196.9 | 153.0 |
| Фоновая территория | |||
| Пос. Порошино (ФОН) | 78.2 | 152.3 | 214.8 |
Сопоставление данных по содержанию ТМ в плодовых телах грибов из различных эколого-трофических групп и субстрате (почве) позволяет заключить, что суммарное накопление ТМ увеличивается в плодовых телах грибов всех эколого-трофических групп в направлении с юга и юго-запада на северо-восток, аналогично ранее отмеченному для почв (табл. 4).
Таблица 4.
Содержание подвижных форм ТМ (мкг/г) в почве, 0–5 см
| Экотоп | Подвижные формы, мкг/г | |||
|---|---|---|---|---|
| Cu2+ | Pb2+ | Zn2+ | Суммарно | |
| Промышленная зона | ||||
| Аллея на площади Лепсе | 2.1 ± 0.2 | 0.7 ± 0.2 | 16.5 ± 0.4 | 19.3 |
| Парк Победы | 0 | 0 | 9.1 ± 2.8 | 9.1 |
| Рекреационная зона | ||||
| Александровский сад | 0.5 ± 0.2 | 3.1 ± 0.6 | 27.1 ± 0.4 | 30.8 |
| Парк им. Гагарина | 0.5 ± 0.1 | 4.7 ± 0.2 | 13.9 ± 1.9 | 19.1 |
| Транспортная зона | ||||
| Сквер комбината ИСКОЖ | 0.1 ± 0.1 | 11.0 ± 2.1 | 14.5 ± 0.1 | 25.6 |
| Парк им. Кирова | 0.7 ± 0.2 | 2.7 ± 0.6 | 13.1 ± 2.7 | 16.5 |
| Фоновая территория | ||||
| Лес в Порошино | 0 | 0 | 4.8 ± 2.1 | 4.8 |
Уровень накопления ТМ в базидиомах для разных металлов был не одинаков. Так, для видов всех эколого-трофических групп наиболее высокие значения аккумуляции были отмечены в отношении Zn (рис. 3). В меньших количествах плодовые тела грибов сорбировали из городской среды Cu и Pb. Данная закономерность наблюдалась во всех обследованных экотопах, в том числе, и на фоновой территории.
Рис. 3.
Усредненные данные по биоаккумуляции ТМ базидиомицетами (мкг/г в.-с. массы). Группа микоризообразователей характеризуется в среднем более высокими концентрациями Zn и Pb (163.7 и 18.1 мкг/г), чем ксилотрофы и подстилочные сапротрофы.
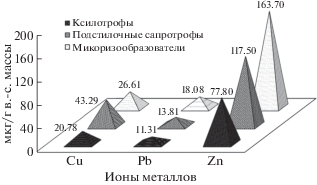
Максимальные значения Zn отмечены у следующих микоризообразующих видов (мкг/г): Paxillus involutus – до 293.3, Tricholoma terreum – до 286.0, Hebeloma crustuliniforme (Bull.) Quél. – 253.8; Pb: Tricoloma terreum – 147.8; T. sciodes (Pers.) C. Martín – 60.2; Cu: Infundibulicybe geotropa (Bull.) Harmaja – 74.1, Paxillus involutus – 71.6. Виды, накапливающие высокие концентрации Zn из группы подстилочных сапротрофов (мкг/г): Clitocybe rivulosa (Pers.) P. Kumm. – 276.1, C. phyllophila (Pers.) P. Kumm. – 210.1, Agaricus bisporus – 201.5.
Высокие значения Zn в плодовых телах Paxillus involutus отмечены также в работах других авторов, так, установлено, что концентрация Zn может достигать 540 мкг/г, Pb – 4.7 мкг/г, Cu – 120 мкг/г (Brzostowski et al., 2011). Плодовые тела Tricholoma terreum, согласно данным других авторов, накапливают Zn до 179 мкг/г, Cu – до 51 мкг/г, Pb – до 4.4 мкг/г (Sazanova et al., 2017).
Подстилочные сапротрофы накапливали также наибольшие концентрации Cu (мкг/г): Agaricus bisporus – 173.8; Lycoperdon perlatum Pers. – 129.8. Pb в максимальных концентрациях в группе был отмечен у вида Melanoleuca polioleuca (Fr.) Kühner et Maire – 121.1 мкг/г. Agaricus bisporus в работах других исследователей аккумулировал Cu до 107 мкг/г, Pb – до 29.7 мкг/г, Zn до 57.2 мкг/г (Sazanova et al., 2017). Способность Lycoperdon perlatum к активному накоплению Cu – до 70 мкг/г отмечена и другими исследователями (Sarikurkcu et al., 2015). По литературным данным, виды рода Melanoleuca накапливают в плодовых телах Pb – 2.68–3.74 мкг/г, Zn – 75–150 мкг/г (Kula et al., 2011).
Несмотря на то, что в среднем ксилотрофы уступали видам других эколого-трофических групп в накоплении Zn среди них также встречались виды с высоким содержанием этого элемента в базидиомах (мкг/г): Bjerkandera adusta – 247.6; Heterobasidion annosum (Fr.) Bref. – 243.5; Pb: Armillaria ostoyae (Romagn.) Herink – 53.0; Cu: Trametes hirsuta (Wulfen) Lloyd – 90.6; Coprinellus disseminatus (Pers.) J.E. Lange – 72.5.
Расчет Кн металлов для представителей разных эколого-трофических групп показал, что все агарикомицеты в городской среде активнее всего концентрируют из субстратов Cu, затем Zn и, в последнюю очередь ─ Pb (рис. 4).
Рис. 4.
Усредненные данные по коэффициентам накопления (Кн) ТМ базидиомицетами из разных эколого-трофических групп.
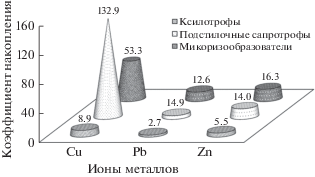
В разрезе отдельных эколого-трофических групп ксилотрофы характеризовались меньшими значениями коэффициентов накопления ТМ (Кн от 0 до до 15.29 в зависимости от металла), по сравнению с микоризообразователями (Кн от 12 до 53) и подстилочными сапротрофами (Кн от 5 до 110). При анализе Кн у микоризообразующих видов отмечали наибольшую способность к биоаккумуляции Cu, несмотря на ее низкую, в сравнении с другими металлами, концентрацию в плодовых телах. Максимальные Кн Cu имели виды Paxillus involutus – 511.4; Tricholoma terreum – 148.3; Hebeloma crustuliniforme – 139.1.
По способности сорбировать из субстратов Zn микоризообразователи (Кн 16) и подстилочные сапротрофы (Кн 13) в среднем различались несущественно. Наибольшие Кн Zn отмечены у видов Tricholoma fulvum (DC.) Bigeard et H. Guill. – 45.7; Hebeloma mesophaeum – 33.8; Tricoloma album (Schaeff.) P. Kumm. – 33.8. Кроме того, микоризообразующие виды характеризовались, по сравнению с видами других эколого-трофических групп, в 2.5–4 раза более высокими Кн Pb: T. terreum – 97.2; T. sciodes – 84.8; Paxillus involutus – 76.4. Подстилочные сапротрофы отличались от других эколого-трофических групп агарикомицетов самыми высокими Кн Cu (в среднем 110) (Agarius bisporus – 1241.7; Melanoleuca polioleuca – 452.1; Coprinus comatus (O.F. Müll.) Pers. – 135.5 и Lycoperdon perlatum – 129.8). Среди часто встречающихся в городской среде видов, максимальным содержанием Cu отличался Agaricus bisporus (173.8 мкг/г), Pb – Tricholoma terreum (147.7 мкг/г), Zn – Paxillus involutus (293.3 мкг/г) (табл. 5).
Таблица 5.
Накопление ТМ в плодовых телах базидиомицетов, характеризующихся высокой (≥50%) частотой встречаемости в городских экотопах
| Виды грибов | Содержание элементов, мкг/г в.-с. массы | Коэффициент накопления (среднее) | ||||
|---|---|---|---|---|---|---|
| Cu | Pb | Zn | Cu | Pb | Zn | |
| Tricholoma terreum (*М) | 17.3/10.9–23.6 | 35.0/0.0–147.7 | 185.3/17.0–286.0 | 36.06 | 17.46 | 16.79 |
| Bjerkandera adusta (*К) | 14.3/1.2–36.0 | 10.8/0.0–23.7 | 96.7/23.1–247.6 | 1.48 | 0.43 | 3.43 |
| Paxillus involutus (М) | 40.3/13.1–71.6 | 18.8/1.0–54.2 | 224.1/139.4–293.3 | 123.16 | 16.65 | 16.81 |
| Agaricus bisporus (*П) | 79.5/38.1–173.8 | 15.2/0.9–20.0 | 116.1/21.7–201.5 | 360.68 | 4.26 | 12.00 |
| Trametes hirsuta (К) | 33.99/5.45–90.63 | 3.65/0.0–10.95 | 55.25/39.61–74.95 | 3.72 | 0.14 | 3.31 |
| Cerioporus squamosus (К) | 18.6/7.4–41.0 | 13.5/3.4–22.0 | 65.1/10.8–192.3 | 1.70 | 0.56 | 1.17 |
| Heterobasidion annosum (К) | 23.0/1.7–46.3 | 16.4/0.0–58.8 | 130.5/41.0–243.5 | 2.20 | 0.65 | 3.62 |
| Postia stiptica (К) | 8.9/3.0–13.6 | 13.0/0.0–19.7 | 54.9/12.1–136.9 | 0.87 | 0.52 | 1.17 |
| Trametes gibbosa (К) | 15.9/8.0–29.6 | 7.3/0.0–12.4 | 46.0/9.1–80.0 | 1.47 | 0.19 | 3.33 |
| T. versicolor (К) | 9.2/2.5–12.7 | 12.5/0.0–30.2 | 80.3/62.8–97.7 | 0.93 | 0.51 | 4.65 |
| Melanoleuca polioleuca (П) | 52.5/40.3–53.6 | 58.2/11.5–121.1 | 156.2/129.6–172.3 | 292.73 | 8.79 | 13.13 |
| Pluteus cervinus (П) | 16.9/12.3–19.3 | 8.8/0.0–9.7 | 103.7/40.9–145.2 | 19.47 | 2.57 | 9.91 |
| Trametes hirsuta (К) | 34.0/5.4–90.6 | 3.6/0.0–11.0 | 55.3/39.6–75.0 | 3.72 | 0.14 | 3.31 |
Коэффициенты корреляции между содержанием ТМ в субстрате и их суммарным накоплением в базидиомах микоризообразователей и ксилотрофов составили соответственно 0.31 и 0.46, что говорит о наличии слабой положительной связи между этими величинами, тогда как для подстилочных сапротрофов такая связь не установлена (r = –0.04). Установлена положительная корреляция между содержанием Pb в базидиомах из различных экотопов Tricholoma terreum (r = 0.8), а также незначительная корреляция по Zn ─ в Paxillus involutus (r = 0.52) и содержанием металлов в субстрате (табл. 6). Данный факт свидетельствует о дополнительных источниках поступления ТМ в плодовые тела, среди которых можно рассматривать аэрогенный путь поступления загрязнителей.
Таблица 6.
Коэффициенты корреляции накопления тяжелых металлов агарикомицетами с высокой частотой встречаемости (≥50%) и содержанием этих металлов в субстрате
| Виды | Коэффициенты корреляции | ||
|---|---|---|---|
| Cu | Zn | Pb | |
| Ксилотрофы | |||
| Bjerkandera adusta | –1.00 | 0.21 | –0.26 |
| Cerioporus squamosus | 0.05 | –0.31 | 0.01 |
| Heterobasidion annosum | –0.10 | –0.52 | 0.47 |
| Trametes gibbosa | –0.48 | 0.57 | –0.37 |
| T. versicolor | 0.50 | 0.22 | –0.92 |
| T. hirsuta | –0.50 | –0.75 | 0.50 |
| Postia stiptica | –0.49 | –0.26 | 0.59 |
| Микоризообразователи | |||
| Tricholoma terreum | 0.11 | 0.07 | 0.80 |
| Paxillus involutus | 0.32 | 0.60 | 0.17 |
| Подстилочные сапротрофы | |||
| Agaricus bisporus | –0.71 | 0.35 | –0.05 |
| Melanoleuca polioleuca | –0.92 | –0.60 | 0.71 |
| Pluteus cervinus | 0.41 | –0.62 | 0.84 |
Показана значимость эколого-трофической специализации агарикомицетов как фактора, способного влиять на состав элементов-загрязнителей в плодовых телах агарикомицетов (рис. 5).
Рис. 5.
Усредненные концентрации ТМ в плодовых телах базидиомицетов из разных эколого-трофических групп (мкг/г).
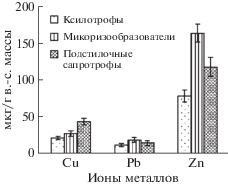
Способность накапливать Cu возрастала в ряду ксилотрофы – микоризообразователи – подстилочные сапротрофы. Ксилотрофные виды агарикомицетов накапливают меньшее количество Cu, Pb, Zn, чем представители других эколого-трофических групп. Микоризообразователи же, наоборот, способны накапливать Zn и Pb в больших количествах, чем другие виды.
Способность к накоплению ТМ плодовыми телами выражена в разной степени во всех эколого-трофических группах исследованных агарикомицетов и носит специфический характер. Также установлено, что аккумулирование некоторых из изученных элементов базидиомами макромицетов может определяться их таксономической принадлежностью (рис. 6).
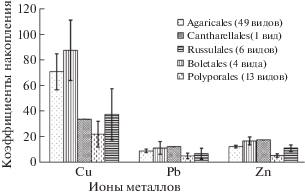
ЗАКЛЮЧЕНИЕ
Анализ полученных данных свидетельствует, что в эколого-трофической структуре агарикомицетов в городской среде доминируют ксилотрофы (59%), тогда как в структуре биоты агарикомицетов фоновой территории ксилотрофы, подстилочные сапротрофы и микоризообразователи представлены приблизительно в равных долях (37, 26 и 37% соответственно).
Выявлено отсутствие четко выраженной связи между аккумуляцией ТМ (Cu, Zn и Pb) плодовыми телами большинства базидиальных макромицетов (за исключением Paxillus involutus, Tricholoma terreum) и содержанием металла в субстрате. Проведенный корреляционный анализ между величиной накопления ТМ в плодовых телах, собранных в различных экотопах, и содержанием этих же металлов в субстрате, не выявил тесной связи (r = = 0.12–0.17) между этими показателями. Это совпадает с результатами работ ряда зарубежных исследователей (Kokkoris et al., 2019). Возможным объяснением тому может служить слабая подвижность ионов тяжелых металлов в щелочной среде (Gäbler, 1997), которая характерна для большинства городов, благодаря промышленному и транспортному загрязнению (Ashikhmina, 2012). Кислотность почв г. Кирова изменяется от pH 7.5 до рН 8.2 на территории парково-рекреационных зон (Efremova et al., 2013), что не способствует высокой подвижности ионов металлов (Gäbler, 1997). Кроме того, на концентрацию элементов в плодовых телах оказывает влияние суммарное накопление металлов (Kokkoris et al., 2019; Borovička et al., 2019), а также аэрогенный способ поступления указанных поллютантов.
На основе расчета коэффициентов накопления выявлены виды-биоконцентраторы Cu (Кн): Agaricus bisporus – 1241.67, Paxillus involutus – 511.43, Melanoleuca polioleuca – 452.14, T. terreum –148.33, Zn: T. fulvum – 45.73, Clitocybe phyllophila – 43.41, Gymnopus dryophilus – 37.24, Tricholoma fulvum – 45.73, Hebeloma mesophaeum – 33.82, Tricholoma album – 33.76 и Pb: T. terreum – 97.21, T. sciodes – 84.75, Paxillus involutus – 76.36, Armillaria ostoyae – 74.58, Pholiota aurivella – 40.42. Полученные сведения о видах-биоконцентраторах в настоящее время представляют интерес для использования грибов в микоремедиации от загрязнения ТМ различных объектов и сред (Bharath et al., 2019; Demasi et al., 2017; Alves et al., 2017).
Список литературы
Abulude F.O., Ndamitso M.M. Assessment of heavy metals in mushroom species obtained in akure, Nigeria AU – eJournal of Interdisciplinary Research. 2018. V. 3 (2). P. 194–201.
Ainsworth G.C. Ainsworth and Bisby’s dictionary of the fungi. Cabi, 2008.
Alves R.P., Bolson S.M., de Albuquerque M.P. et al. A potencial use of edible mushrooms Pleurotus ostreatoroseus Singer (Pleurotaceae) and Lentinus sajor-caju (Fr.) Fr. (Polyporaceae) in metal remediation processes. Revista De Biologia Neotropica. 2017. V. 14 (2). P. 82–90.
nishchenko L.N., Ladnova G.G., Frolova N.V. Features of the accumulation of heavy metals by the fruiting bodies of macromycetes under conditions of combined radiation-chemical pollution. Siberian J. Life Sciences and Agriculture. 2016. № 8 (80). P. 81–96 (in Russ.)
Ashikhmina T.Ya. Features of urban ecosystems of the subzone of the southern taiga of the European Northeast. Kirov, 2012 (in Russ.).
Bharath Y., Singh S.N., Keerthiga G. et al. Mycoremediation of contaminated soil in MSW sites. In: Waste management and resource efficiency. Singapore, 2019, pp. 321–329.
Bondartsev A.S., Singer R. A handbook on collection of higher basidial fungi for their scientific study. Trudy Botanicheskogo instituta imeni V.L. Komarova AN SSSR. 1950. V. 2 (6). P. 499–543 (in Russ.).
Bondartseva M.A. Keybook to the fungi of Russia. The order Aphyllophorales. Issue 2. St. Petersburg, Nauka, 1998 (in Russ.).
Borovička J., Braeuer S., Sácký J. et al. Speciation analysis of elements accumulated in Cystoderma carcharias from clean and smelter-polluted sites. Science of the total environment. 2019. V. 648. P. 1570–1581. https://doi.org/10.1016/j.scitotenv.2018.08.202
Brzostowski A., Falandysz J., Jarzyńska G. et al. Bioconcentration potential of metallic elements by Poison Pax (Paxillus involutus) mushroom. J. Environmental Science and Health. Pt A. 2011. V. 46. № 4. P. 378–393. https://doi.org/10.1080/10934529.2011.542387
Burova L.G. Ecology of macromycetes. Nauka, Moscow, 1986 (in Russ.).
Churakov B.P., Lisov E.S., Evseeva N.A. et al. Mycoindication of heavy metal pollution of forest ecosystems. Mikologiya i fitopatologiya. 2000. V. 34 (2). P. 57–61 (in Russ.).
Dabakhov M.V., Dabakhova E.V., Titova V.I. Ecotoxicology and rationing problems. N. Novgorod, 2005 (in Russ.).
de Miguel E., Llamas J.F., Chacón E. et al. Origin and patterns of distribution of trace elements in street dust: unleaded petrol and urban lead. Atmospheric Environment. 1997. V. 31 (17). P. 2733–2740. https://doi.org/ 00101-5https://doi.org/10.1016/S1352-2310(97)
Demasi S., Berruti A., Ajmone Marsan F. et al. Role of mycorrhization in the phytoremediation of heavy metals in urban soils. In: International symposium on greener cities for more efficient ecosystem services in a climate changing world 1215–2017. P. 311–314. https://doi.org/10.17660/ActaHortic.2018.1215.57
Demirbaş A. Heavy metal bioaccumulation by mushrooms from artificially fortified soils. Food Chemistry. 2001. V. 74. № 3. P. 293–301. https://doi.org/10.1016/S0308-8146(01)00155-8
Dremova N.A. Xylotrophic basidiomycetes (Basidiomycota) as an indicator of the ecological state of the urban environment. Izvestiya Orenburgskogo gosudarstvennogo agrarnogo universiteta. 2014. № 2. P. 150–155 (in Russ.).
Efremova V.A., Kondakova L.V., Dabakh E.V. Chemical and biological assessment of urban soils. Sibirskiy ekologicheskiy zhurnal. 2013. V. 20 (5). P. 741–750 (in Russ.).
Gäbler H.E. Mobility of heavy metals as a function of pH of samples from an overbank sediment profile contaminated by mining activities. J. Geochem. Exploration. 1997. V. 58 (2–3). P. 185–194. https://doi.org/10.1016/S0375-6742(96)00061-1
Hussain M., Ullah S.H., Baqi A. et al. Study of heavy metals (Cd, Cu, Ni, Pb and Zn) in some medicinal plant species (Hertia intermedia, Cardaria chalepense, Scorzonera ammophila, Tamarix karelini, Astragalus auganus) at Pishin area in Balochistan, Pakistan. Pure and Applied Biology. 2019. V. 8 (1). P. 995–1007. https://doi.org/10.19045/ bspab.2019.80040
Index Fungorum. 2020. http://www.indexfungorum.org/Names/Names.asp. Accessed 08.01.2020.
Ivanov A.I., Kostychev A.A. Character of accumulation of some metals and arsenic in basidiomes of fungi of the order Boletales. Mikologiya i fitopatologiya. 2007. V. 41 (6). P. 500–505 (in Russ.).
Ivanov A.I., Kostychev A.A., Skobanev A.V. Accumulation of heavy metals and arsenic by basidioms of macromycetes of various ecological-trophic and taxonomic groups. Povolzhskiy ekologicheskiy zhurnal. 2008. № 3. P. 190–199 (in Russ.).
Kibby G. Atlas of mushrooms: key to species. SPb., Amfora, 2009 (in Russ.).
Knudsen H., Vesterholt J. Funga Nordica. Agaricoid, boletoid and cyphelloid genera. Nordsvamp, Copenhagen, Denmark, 2008.
Kokkoris V., Massas I., Polemis E. et al. Accumulation of heavy metals by wild edible mushrooms with respect to soil substrates in the Athens metropolitan area (Greece). Science of the Total Environment. 2019. V. 685. P. 280–296. https://doi.org/10.1016/j.scitotenv.2019.05.447
Korolev A.N., Boev V.A. Heavy metals in soils and vegetable crops in the zone of influence of Semey cement plant (Republic of Kazakhstan). Vestnik Omskogo gosudarstvennogo agrarnogo universiteta. 2017. № 3 (27). P. 74–79 (in Russ.).
Kostychev A.A. Accumulation of lead and arsenic by fruit bodies of wild mushrooms in the Penza region. In: Sovremennaya mikologiya v Rossii. Moscow, 2012. P. 187 (in Russ.).
Kostychev A.A. Possibility of using basidial macromycetes as bioindicators of environmental pollution by heavy metals and arsenic. Vestnik OGU. 2009. № 1. P. 108–112 (in Russ.).
Kovalenko A.E. Ecological review of fungi from the orders of Polyporales s.str., Boletales, Agaricales s.str., Russulales in the mountain forests of the central part of the Northwest Caucasus. Mikologiya i fitopatologiya. 1980. V. 14 (4). P. 300–314 (in Russ.).
Kula I., Solak M.H., Uğurlu M. et al. Determination of mercury, cadmium, lead, zinc, selenium and iron by ICP-OES in mushroom samples from around thermal power plant in Muğla, Turkey. Bulletin of environmental contamination and toxicology. 2011. V. 87 (3). P. 276–281. https://doi.org/10.1007/s00128-011-0357-1
Lakin G.F. Biometrics. Vysshaya shkola, Moscow, 1990 (in Russ.).
Lesso T. Mushrooms: keybook. Moscow, 2003 (in Russ.).
Malysheva V.F., Malysheva E.F. Higher basidiomycetes of forest and meadow ecosystems of Zhiguli. Tovarishchestvo nauchnykh izdaniy KMK, Moscow, SPb., 2008 (in Russ.).
Morozova O.V. Agaricoid basidiomycetes of the southern taiga subzone of the Leningrad Region. Abstract of Cand. Sci thesis. SPb., 2001 (in Russ.).
Niemelä T. Tinder mushrooms of Finland and the adjacent territory of Russia. Finnish museum of Natural History, Helsinki, 2001 (in Russ.).
Nunez M., Ryvarden L. East Asian polypores. Vol. 1: Ganodermataceae and Hymenochaetaceae. Fungiflora, Oslo, 2000.
Otmakhov V.I., Petrova E.V., Pushkareva T.N. et al. Atomic emission method of analysis of mushrooms on the content of heavy metals and its use for the purposes of environmental monitoring. Izvestiya Tomskogo politekhnicheskogo universiteta. 2004. V. 307 (6). P. 44–46 (in Russ.).
Otnyukova T.N., Zhizhayev A.M., Kutafyeva N.P. et al. Macromycetes as bioindicators of environmental pollution in the territory of the city of Krasnoyarsk and its vicinities. Vestnik Krasnoyarskogo gosudarstvennogo agrarnogo universiteta. 2012. № 11. P. 101–113 (in Russ.).
Palamarchuk M.A., Kirillov D.V. Agaricoid basidiomycetes of Syktyvkar and the surrounding area (Republic of Komi). Mikologiya i fitopatologiya. 2017. V. 51. № 3. P. 137–146 (in Russ.).
Sarikurkcu C., Tepe B., Kocak M.S. et al. Metal concentration and antioxidant activity of edible mushrooms from Turkey. Food Chem. 2015. V. 175. P. 549–555. https://doi.org/10.1016/j.foodchem.2014.12.019
Savelev L.A., Kikeeva A.V. Agaricoid basidiomycetes of green spaces in Petrozavodsk (Republic of Karelia). Lesotekhnicheskiy zhurnal. 2018. V. 8 (1). P. 50–68 (in Russ.).
Sazanova K.V., Velikova V.D., Stolyarova N.V. The accumulation of heavy metals by mushrooms. Ecological and species specificity, accumulation mechanisms, potential danger to humans. Medline.ru. 2017. V. 18. P. 336–361 (in Russ.).
Shcheglov A.I., Tsvetnova O.B. Fungi as bioindicators of technogenic pollution. Priroda. 2002. № 11. P. 7–16 (in Russ.).
Shilkova T.A. Agaricoid basidiomycetes of the city of Perm: taxonomic diversity and ecology. Cand. Biol. Thesis. Perm, 2015 (in Russ.).
Širić I., Kasap A., Bedeković D. et al. Lead, cadmium and mercury contents and bioaccumulation potential of wild edible saprophytic and ectomycorrhizal mushrooms, Croatia. J. Environm. Sci. and Health. Pt B. 2017. V. 52 (3). P. 156–165. https://doi.org/10.1080/03601234.2017.1261538
Tsvetnova O.B., Shatrova N.E., Shcheglov A.I. Accumulation of radionuclides and heavy metals by the mushroom complex of forest ecosystems. Sbornik nauchnykh trudov IYaI. Kiev. 2001. № 3 (5). P. 171–176 (in Russ.).
Vorobyova L.A. Theory and practice of soil chemical analysis. Geos, Moscow, 2006 (in Russ.).
Yamaç M., Yıldız D., Sarıkürkcü C. et al. Heavy metals in some edible mushrooms from the Central Anatolia, Turkey. Food Chem. 2007. V. 103 (2). P. 263–267. https://doi.org/10.1016/j.foodchem.2006.07.041
Zaytsev G.N. Methods of biometric calculations. Nauka, Moscow, 1963 (in Russ.).
Анищенко Л.Н., Ладнова Г.Г., Фролова Н.В. (Anishchenko et al.) Особенности накопления тяжелых металлов плодовыми телами макромицетов в условиях сочетанного радиационно-химического загрязнения // Siberian Journal of Life Sciences and Agriculture. 2016. № 8 (80). С. 81–96.
Ашихмина Т.Я. (Ashikhmina) Особенности урбоэкосистем подзоны южной тайги Европейского Северо-Востока. Киров: Изд-во ВятГГУ, 2012. 282 с.
Бондарцев А.С., Зингер Р. (Bondartsev, Singer) Руководство по сбору высших базидиальных грибов для научного их изучения // Труды Ботанического института им. В.Л. Комарова. Сер. 2. 1950. № 6. С. 499–543.
Бондарцева М.А. (Bondartseva) Определитель грибов России. Порядок афиллофоровые. Вып. 2. СПб.: Наука, 1998. 391 с.
Бурова Л.Г. (Burova) Экология грибов макромицетов. М.: Наука, 1986. 224 с.
Воробьева Л.А. (Vorobyova) Теория и практика химического анализа почв. М.: Геос, 2006. 400 с.
Дабахов М.В., Дабахова Е.В. Титова В.И. (Dabakhov et al.) Экотоксикология и проблемы нормирования. Н. Новгород: Нижегородская гос. с.-х. академия, 2005. 165 с.
Дремова Н.А. (Dremova) Ксилотрофные базидиомицеты (Basidiomycota) как показатель экологического состояния городской среды // Известия Оренбургского государственного аграрного университета. 2014. № 2. С. 150–155.
Ефремова В.А., Кондакова Л.В., Дабах Е.В. (Efremova et al.) Химико-биологическая оценка состояния городских почв. Сибирский экологический журнал. 2013. Т. 20. № 5. С. 741–750.
Зайцев Г.Н. (Zaytsev) Методика биометрических расчетов. М.: Наука, 1963. 256 с.
Иванов А.И., Костычев А.А. Характер накопления некоторых металлов и мышьяка в базидиомах грибов порядка Boletales // Микология и фитопатология. 2007. Т. 41. № 6. С. 500–505.
Иванов А.И., Костычев А.А., Скобанев А.В. (Ivanov et al.) Аккумуляция тяжелых металлов и мышьяка базидиомами макромицетов различных эколого-трофических и таксономических групп // Поволжский экологический журнал. 2008. № 3. С. 190–199.
Кибби Дж. (Kibby) Атлас грибов: Определитель видов. СПб.: Амфора, 2009. 269 с.
Коваленко А.Е. (Kovalenko) Экологический обзор грибов из порядков Polyporales s. str., Boletales, Agaricales s. str., Russulales в горных лесах центральной части Северо-Западного Кавказа // Микология и фитопатология. 1980. Т. 14. № 4. С. 300–314.
Королев А.Н., Боев В.А. (Korolev, Boev) Тяжелые металлы в почвах и овощных культурах в зоне влияния цементного завода города Семей (Республика Казахстан) // Вестник Омского государственного аграрного университета. 2017. № 3 (27). С. 74–79.
Костычев А.А. (Kostychev) Возможность использования базидиальных макромицетов в качестве биоиндикаторов загрязнения окружающей среды тяжелыми металлами и мышьяком // Вестник ОГУ. 2009. № 1. С. 108–112.
Костычев А.А. (Kostychev) Накопление свинца и мышьяка плодовыми телами дикорастущих грибов в условиях Пензенской области. Современная микология в России: материалы 3-го Съезда микологов России. М.: Национальная академия микологии, 2012. С. 187.
Лакин Г.Ф. (Lakin) Биометрия. М.: Высшая школа, 1990. 351 с.
Лессо Т. (Lesso) Грибы: Определитель. М.: Издательство АСТ, 2003. 304 с.
Малышева В.Ф., Малышева Е.Ф. (Malysheva, Malysheva) Высшие базидиомицеты лесных и луговых экосистем Жигулей. М.; СПб.: Товарищество научных изданий КМК, 2008. 242 с.
Морозова О.В. (Morozova) Агарикоидные базидиомицеты подзоны южной тайги Ленинградской области. Автореф. дисс. … канд. биол. наук. СПб.: БИН РАН, 2001. 22 с.
Ниемеля Т. (Niemelä) Трутовые грибы Финляндии и прилегающей территории России. Хельсинки, 2001. 120 с.
Отмахов В.И., Петрова Е.В., Пушкарева Т.Н. и др. (Otmakhov et al.) Атомно-эмиссионная методика анализа грибов на содержание тяжелых металлов и использование ее для целей экомониторинга // Известия Томского политехнического университета. 2004. Т. 307. № 6. С. 44–46.
Отнюкова Т.Н., Жижаев А.М., Кутафьева Н.П. и др. (Otnyukova et al.) Макромицеты как биоиндикаторы загрязнения окружающей среды территории г. Красноярска и его окрестностей // Вестник Красноярского государственного аграрного университета. 2012. № 11. С. 101–113.
Паламарчук М.А., Кириллов Д.В. (Palamarchuk, Kirillov) Агарикоидные базидиомицеты Сыктывкара и его окрестностей (Республика Коми) // Микология и фитопатология. 2017. Т. 51. № 3. С. 137–146.
Савельев Л.А., Кикеева А.В. (Savelev, Kikeeva) Aгарикоидные базидиомицеты зеленых насаждений города Петрозаводска (Республика Карелия) // Лесотехнический журнал. 2018. Т. 8. № 1 (29). С. 50–68.
Сазанова К.В., Великова В.Д., Столярова Н.В. (Sazanova et al.) Накопление тяжелых металлов грибами. Экологическая и видовая специфичность, механизмы аккумуляции, потенциальная опасность для человека // Medline.ru. 2017. Т. 18. С. 336–361.
Цветнова О.Б., Шатрова Н.Е., Щеглов А.И. (Tsvetnova et al.) Накопление радионуклидов и тяжелых металлов грибным комплексом лесных экосистем // Сборник науч. трудов ИЯИ. Киев. 2001. № 3 (5). С. 171–176.
Чураков Б.П., Лисов Е.С., Евсеева Н.А. и др. (Churakov et al.) Микоиндикация загрязнения лесных экосистем тяжелыми металлами // Микология и фитопатология. 2000. Т. 34 (2). С. 57–61.
Шилкова Т.А. (Shilkova) Агарикоидные базидиомицеты города Перми: таксономическое разнообразие и экология. Автореф. дисc. … канд. биол. наук. Пермь, 2015.
Щеглов А.И., Цветнова О.Б. (Shcheglov, Tsvetnova) Грибы – биоиндикаторы техногенного загрязнения // Природа. 2002. № 11. С. 7–16.
Дополнительные материалы отсутствуют.
Инструменты
Микология и фитопатология