

Молекулярная биология, 2021, T. 55, № 3, стр. 478-490
Профилактическая вакцинация мезенхимальными стволовыми клетками защищает мышей от летальной инфекции, вызванной вирусом простого герпеса 1 типа
Р. Р. Климова a, *, Н. А. Демидова a, О. В. Масалова a, А. А. Кущ a
a Национальный исследовательский центр эпидемиологии и микробиологии
им. почетного акад. Н.Ф. Гамалеи Минздрава России
123098 Москва, Россия
* E-mail: regi.k@mail.ru
Поступила в редакцию 13.08.2020
После доработки 10.09.2020
Принята к публикации 10.09.2020
Аннотация
Вирусы простого герпеса 1 и 2 типов (ВПГ-1 и ВПГ-2) поражают практически все органы и ткани, вызывают генитальный герпес – самое распространенное заболевание, передающиеся половым путем, ‒ нарушения ЦНС, приводят к тяжелым осложнениям у детей. Несмотря на имеющиеся лекарственные препараты, заболеваемость, вызванная ВПГ-1/2, продолжает расти. Ни одна из профилактических кандидатных вакцин не проявила защитное действие при испытаниях и не получила разрешения на использование в клинической практике. Нами исследованы защитные свойства мезенхимальных стволовых клеток (МСК), выделенных из костного мозга мышей. Проведен сравнительный анализ протективного ответа на введение первичных и модифицированных МСК (мМСК); плазмиды, содержащей ген ВПГ, и инактивированного вируса на модели летальной ВПГ-1-инфекции у мышей. мМСК получали путем трансфекции гена Us6, кодирующего гликопротеин D (gD) ВПГ; плазмида содержала тот же ген. После двукратной иммунизации первичными МСК в периферической крови мышей наблюдали образование антител, взаимодействующих с вирусным антигеном (по данным иммуноферментного анализа) и нейтрализующих инфекционную активность ВПГ-1 в реакции биологической нейтрализации. Кроме того, введение первичных МСК индуцировало продукцию интерферона гамма (ИНФ-γ), детектируемого в периферической крови мышей. После заражения ВПГ-1 у иммунизированных мышей значительно повышались титры вирусспецифических антител, уровень ИФН-γ и они были полностью защищены от летальной ВПГ-1-инфекции. Защитный эффект трех других иммуногенов был ниже и не превышал 50‒65%. Учитывая широкую доступность МСК, доказанную безопасность при внутривенном введении и впервые полученные в работе результаты о способности индуцировать врожденный, адаптивный и протективный иммунитет к ВПГ-1, можно считать МСК перспективной основой для создания новых клеточных вакцин для профилактики герпесвирусных и других вирусных инфекций.
Вирусы простого герпеса 1 и 2 типов (ВПГ-1 и ВПГ-2) – повсеместно распространенные патогены человека: 3.7 млрд инфицировано ВПГ-1 и 417 млн – ВПГ-2 [1‒3]. Согласно данным ВОЗ [4], генитальный герпес признан самой распространенной инфекцией, передающейся половым путем; доказано, что ВПГ-инфекция повышает риск заражения ВИЧ [5]. Оба вируса вызывают неонатальный герпес [6]. ВПГ-1 ‒ ведущий этиологический агент инфекционной слепоты, в мире ежегодно регистрируют 1 млн новых случаев и 9 млн повторяющихся эпизодов ВПГ-инфекции глаз [7]. В настоящее время множество лекарственных препаратов доступно для лечения герпетической инфекции, однако заболеваемость, вызванная ВПГ, продолжает расти [8]. В связи с этим разработка эффективной профилактической вакцины против ВПГ-1/2 признана ВОЗ приоритетным направлением здравоохранения (http://www.who.int/immunization/global_vaccine_ action_plan/GVAP_doc_2011_2020/en/) [9, 10].
В настоящее время разработан целый ряд вакцин против герпесвирусных инфекций, основанных на рекомбинантных ДНК и белках ВПГ, пептидах, живых аттенуированных вирусах и инактивированных цельновирионных препаратах [11‒13]. Однако для большинства кандидатных вакцин не выявлено протективного действия при клинических испытаниях [14], и пока ни одна из профилактических вакцин не получила разрешения на использование в медицине.
Очевидно, что необходимы новые подходы к созданию герпетических вакцин, один из которых изучен в представленной работе. Он основан на использовании мезенхимальных стволовых клеток (МСК), успешно применяемых в разных областях регенеративной медицины. Разрабатываются подходы для получения генетически модифицированных МСК (мМСК), которые экспрессируют целевые гены и могут быть использованы для клеточной терапии раковых, аутоиммунных, нейродегенеративных и инфекционных заболеваний [15‒20]. На основании этих данных можно сделать вывод о пластичности иммуномодуляции МСК, которые могут оказывать как супрессивное, так и иммуностимулирующее действие. Появились первые работы, в которых показана принципиальная возможность использования МСК и мМСК в качестве не традиционных, а инновационных вакцин, усиливающих иммунные реакции против ВИЧ [21] и вируса гепатита С [22].
Цель работы состояла в сравнительном анализе протективного иммунного ответа на введение профилактических кандидатных вакцин на основе МСК, ДНК-плазмиды и инактивированного вируса в отношении летальной ВПГ-1-инфекции мышей.
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
Животные. Мышей линии DBA/2J (H-2d) (самки, 6‒8 недель) получали из Центрального питомника лабораторных животных “Столбовая” (Московская область).
Культуры клеток. Первичную культуру МСК получали из красного костного мозга мышей, как описано ранее [23]. Нами было показано [22, 23], что по адгезивной способности, поверхностным маркерам и дифференцировочному потенциалу эти клетки соответствуют критериям определения МСК Международного общества клеточной терапии (The International Society for Cellular Therapy) [24].
Культивирование МСК проводили в среде DMEM, содержащей 10% эмбриональной телячьей сыворотки (ЭТС; “Gibco”, США), 10 мкг/мл инсулина, 5.5 мкг/мл трансферрина, 6.7 нг/мл селенита, 10 нг/мл фактора роста фибробластов, 2 мМ L-глутамина, 50 мкг/мл гентамицина, в атмосфере 5% СО2 при 37°С.
Культуру клеток почечного эпителия зеленой мартышки (Vero) получили из коллекции клеточных культур ФГБУ “НИЦЭМ им. Н.Ф. Гамалеи” Минздрава России и культивировали в аналогичных условиях, используя среду Игла MEM с добавлением 10% ЭТС (“Biosera”, Франция), 2 мМ L-глутамина, 50 мкг/мл гентамицина. Культуральные среды, сыворотки и компоненты для культивирования клеточных культур, если не указано другое, были приобретены в НПП “ПанЭко” (Россия).
Вирус. ВПГ-1, штамм УС, полученный из Государственной коллекции вирусов ФГБУ “НИЦЭМ им. Н.Ф. Гамалеи” Минздрава России, размножали в клетках Vero, используя стандартные методы культивирования. Множественность инфекции вируса определяли модифицированным методом бляшек in vitro. Готовили серии разведений образцов вируса и вносили в 96-луночные планшеты с монослоем клеток, инкубировали при 37°C в течение 7 сут в присутствии 5% СО2. Очаги инфицированных клеток (бляшки) выявляли и подсчитывали с использованием инвертированного микроскопа Primovert (“Zeiss”, Германия). Титр вируса определяли по формуле: А = = ab/V, где А – число бляшкообразующих единиц на 1 клетку (БОЕ/кл); a – среднее число бляшек на одну лунку; b – разведение вируса; V – объем внесенного вируссодержащего материала.
Определение 50%-ной летальной дозы (ЛД50) ВПГ-1 in vivo проводили путем заражения животных последовательными 10-кратными разведениями вируса внутрибрюшинно. Гибель животных регистрировали в течение 14 сут после заражения. Расчет летальной дозы ВПГ-1 проводили по формуле: ЛД50= 1 × 10Х+[(C1 – 50) /(C1 – C2)], где Х – обратная величина разведения вируса, при которой наблюдали 50%-ную гибель мышей; С1 ‒ доля мышей (%), павших при максимальном разведении, давшем летальность выше 50%; С2 ‒ доля мышей (%), павших при минимальном разведении, давшем летальность ниже 50%.
Получение инактивированного препарата ВПГ-1. Клетки Vero высаживали в культуральные флаконы 175 см2 и по достижении монослоя заражали ВПГ-1. После проявления максимального вирусспецифического цитопатического действия (ЦПД) проводили 1 цикл замораживания‒оттаивания. Вируссодержащую культуральную жидкость центрифугировали (“Jouane”, Франция) при 10 000 об/мин в течение 10 мин для удаления клеточного дебриса и из полученного супернатанта осаждали вирус ультрацентрифугированием при 30 000 об/мин в течение 1 ч при 4°С (ротор SW32; “Beckman”, США). Осадок ресуспендировали в 1 мл 10 мМ Трис-HCl, pH 7.4, и оставляли на ночь при комнатной температуре. В центрифужный флакон наслаивали 2 мл 60%-ной сахарозы, затем 10 мл 15%-ной сахарозы и вируссодержащую суспензию. Ультрацентрифугирование проводили в условиях, описанных выше. Очищенный ВПГ-1 инактивировали при 56°C в течение 30 мин. Инактивированный вирус (иВПГ) использовали для иммунизации животных и в качестве антигена для иммуноферментного анализа (ИФА).
Плазмида. Использовали ДНК-конструкцию pcDNAgD (ДНКgD), полученную на основе коммерческой плазмиды pcDNA-3.1(+) (“Invitrogen”, США), в которую был встроен ген Us6, кодирующий белок gD ВПГ, и маркерный ген neo (неомицин-фосфотрансферазы). Для наработки конструкции использовали клетки Escherichia coli штамм JM109, которые выращивали в питательной среде (LB-бульон, LB-агар и SOB-бульон), состоящей из бактотриптона, дрожжевого экстракта и бактоагара (“Difco”, США). Плазмиду выделяли с использованием коммерческого набора QIAGEN Plasmid Purification Maxi Kit (“QIAGEN”, США) согласно методике фирмы-производителя.
Трансфекция культуры МСК. Для получения генетически модифицированных МСК (мМСК), экспрессирующих ген Us6 ВПГ, использовали первичную культуру МСК на четвертом пассаже. Клетки трансфицировали с помощью Lipofec-tamine 3000 (“Thermo Fisher Scientific”, США) в соответствии с рекомендациями производителя. МСК высаживали в 24-луночный планшет в концентрации 2 × 104 клеток/мл в ростовой среде DMEM без антибиотиков. Через 24 ч в клетки вносили комплекс ДНКgD и Lipofectamine 3000 в соотношении 1 : 2. Селекцию трансформантов проводили в культуральной среде, содержащей генетицин G-418 (“Invitrogene”). Через 5 и 14 суток в клетках, полученных в результате селекции, анализировали экспрессию гена Us6. Клетки фиксировали охлажденным метанолом, обрабатывали моноклональными антителами против белка gD ВПГ (ab6507; “Abсam”, Великобритания) в течение 1 ч при 37°C. Несвязавшиеся антитела отмывали фосфатно-солевым буфером (PBS), наслаивали анти-мышинные антитела, коньюгированные с пероксидазой хрена (anti-mouse PX, PO260; “Dako”, Дания), и инкубировали 30 мин при 37°C. Клетки промывали PBS и наносили раствор 3,3'-диаминобензидина в концентрации 0.05 мг/мл в буфере 0.05 М Трис-HCl (pH 7.4) с добавлением 3% пероксида водорода. Реакцию останавливали через 10 мин внесением в лунки дистиллированной воды. Результаты оценивали с использованием микроскопа AxioScopeA1 (“Zeiss”, Германия). Подсчитывали число окрашенных клеток, содержавших белок gD ВПГ, и представляли в процентах от общего числа клеток в популяции.
Для изучения продукции цитокинов, секретируемых мМСК, в течение 14 сут селекции МСК отбирали культуральную жидкость и определяли в ней концентрацию цитокинов, как описано ниже.
Иммунизация. Животным, разделенным на 5 групп, вводили первичные МСК в концентрации 5 × 105 клеток/мышь внутривенно (МСК; группа 1, n = 15); мМСК (модифицированные МСК, экспрессирующие ген Us6 ВПГ) в концентрации 5 × × 105 клеток/мышь внутривенно (МСКgD; группа 2, n = 10); рекомбинантную ДНК, содержащую ген Us6 ВПГ, в концентрации 100 мкг/мышь внутримышечно (ДНКgD; группа 3, n = 20); инактивированный вирус в концентрации 100 мкг/мышь внутривенно (иВПГ; группа 4, n = 10). В качестве контроля использовали группу 5 (n = 20), где мышам вводили внутривенно физиологический раствор. Иммунизацию проводили дважды с интервалом 14 сут.
Заражение. Мышей групп 1‒5 заражали ВПГ-1 (15 ЛД50 внутрибрюшинно) через 14 суток после последней иммунизации. Наблюдение за животными вели ежедневно в течение 14 сут. Выживаемость животных рассчитывали в процентах (%).
Анализ иммунного ответа. Изучение активности противовирусных и вируснейтрализующих антител, количественное определение уровней провоспалительных цитокинов в культуральной жидкости от мМСК и в сыворотках животных, а также забор спленоцитов проводили до и после заражения ВПГ-1: через 14 сут после последней иммунизации и через 10 сут после заражения.
Определение антител к ВПГ-1. Титры вирусспецифических антител к ВПГ-1 определяли в сыворотках крови мышей методом ИФА, как описано ранее [23]. Для определения ВПГ-специфических иммуноглобулинов субклассов IgG1 и IgG2а 96-луночные планшеты сенсибилизировали иВПГ в объеме 100 мкл/лунка с концентрацией 50 мкг/мл в течение ночи при комнатной температуре. Затем вносили сыворотки животных в разведении 1 : 100 и инкубировали в течение 1 ч при 37°С. Изотип иммуноглобулинов, связавшихся с ВПГ-1, определяли с использованием овечьих анти-мышиных антител, конъюгированных с пероксидазой хрена (115-035-205 и 115-035-206 соответственно; “Jackson Immuno Research Lab”, США).
Реакция биологической нейтрализации. Нейтрализующую активность – способность антител подавлять репликацию ВПГ-1 – определяли на культуре клеток Vero. Сыворотки крови мышей в различных разведениях инкубировали с ВПГ-1 (множественность инфекции = 0.01 БОЕ/клетка) в течение 2 ч при 37°С. Затем реакционную смесь наносили на монослой клеток Vero. Результаты регистрировали через 48 ч, когда в инфицированной культуре, не обработанной сыворотками, детектировали максимальную величину ЦПД. За активность вируснейтрализующих антител принимали обратные величины максимальных разведений сывороток, при которых наблюдали 50%-ное подавление вирусспецифического ЦПД.
Реакция бласттрансформации (РБТ). О стимуляции пролиферации Т-лимфоцитов судили по изменению морфологии клеток в РБТ in vitro – по образованию бластов. Спленоциты мышей каждой группы пулировали и помещали в 24-луночные планшеты по 105 клеток в лунку. Культивировали в ростовой среде (Рс) RPMI-1640, содержащей 20% ЭТС (“Invitrogene”), 4.5 мг/мл глюкозы, 2 мМ глутамин, 0.2 ед./мл инсулина, 50 мкг/мл гентамицина. В лунки вносили иВПГ, который использовали в качестве специфического стимулятора, в конечной концентрации 50 мкг/мл, а для оценки спонтанной пролиферации вносили Рс без стимуляторов. Митоген конкавалин А (ConA, 5 мкг/мл; “Sigma”, США) использовали как положительный контроль. Клетки инкубировали 5 сут при 37°С в атмосфере 5% СО2, после чего подсчитывали число бластов в каждой лунке (по 3 лунки на вариант) и рассчитывали средние значения. Результаты представляли как индекс стимуляции пролиферации (ИСП), который рассчитывали по формуле ИСП = a/b, где a – число антигенспецифичных бластов, b – число неспецифичных (спонтанных) бластов.
Количественный анализ цитокинов. Интерлейкины 2 и 6 (ИЛ-2 и ИЛ-6 соответственно), интерферон-γ (ИНФ-γ) и фактор некроза опухолей-α (ФНО-α) определяли в культуральных жидкостях от первичных МСК и мМСК в динамике, используя метод ИФА. Секрецию ИЛ-6, ИНФ-γ и ФНО-α в сыворотках крови мышей оценивали через 14 сут после последней иммунизации и через 14 сут после заражения ВПГ-1. Использовали тест-системы Mouse IL-2 ELISA development kit (HRP), Mouse IL-6 ELISA development kit (HRP), Mouse IFN-γ ELISA development kit (HRP) и Mouse TNF-α ELISA development kit (HRP) (“Mabtech”, Швеция). Чувствительность анализа составила 20 пг/мл для ИЛ-6, 6 пг/мл для ФНО-α и 4 пг/мл для ИФН-γ и ИЛ-2. Концентрации цитокинов рассчитывали по соответствующим калибровочным кривым согласно инструкциям производителя.
Статистическая обработка результатов. Для статистической обработки результатов использовали программу GraphPadPrism 5.1 и Statistica 6.0. Для сравнения параметрических количественных данных использовали t-тест (критерий Стьюдента), непараметрических – U-тест (критерий Манна‒Уитни). Межгрупповые различия относительных показателей анализировали с помощью критерия χ2 (хи-квадрат). Различия показателей считали статистически значимыми при р < 0.05. Для оценки гипотезы о связи двух переменных проводили корреляционный анализ и рассчитывали коэффициент корреляции Спирмена (r). Значение r > 0.7 рассматривали как свидетельство высокой связи.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
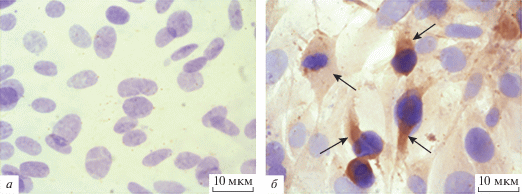
Экспрессию гена Us6 ВПГ определяли иммуноцитохимически с использованием моноклональных антител к белку gD через 5 и 14 сут после трансфекции первичных МСК плазмидой ДНКgD и селекции в присутствии генетицина G-418. В результате подсчета окрашенных клеток показано, что через 5 сут 75 ± 5% клеток содержали вирусный белок (рис. 1а, 1б), а через 14 сут все клетки продуцировали белок gD ВПГ. Полученные мМСК, стабильно продуцирующие белок gD ВПГ (МСКgD), использовали для иммунизации животных.
Рис. 1.
Иммунноцитохимическая детекция белка gD ВПГ в генетически модифицированной культуре МСК, трансфицированной плазмидой ДНКgD. а – Первичная культура МСК; б – модифицированные МСК. Стрелками указано накопление белка gD в цитоплазме трансфицированных клеток (коричневое окрашивание). Ядра окрашены гематоксилином (синее окрашивание). Увеличение 400×.
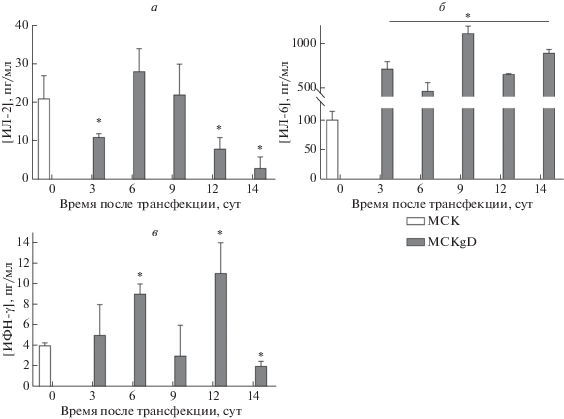
Концентрацию цитокинов ИЛ-2, ИЛ-6, ИНФ-γ и ФНО-α, секретируемых МСК в динамике после трансфекции и селекции in vitro, определяли в культуральных жидкостях, начиная с 3 сут после трансфекции и заканчивая 14 сут ‒ перед введением мМСК мышам. Уровень ИЛ-2 на третьи сутки после трансфекции снижался, затем на 6‒9 сут повышался, но к 14 суткам снова сильно снижался – приблизительно в 10 раз по сравнению с уровнем первичных МСК (р < 0.05) (рис. 2а). Секреция ИЛ-6, напротив, увеличивалась уже на 3 сутки после трансфекции и оставалась на высоком уровне, превышая к 14 суткам уровень в первичных МСК в 9 раз (р < 0.05) (рис. 2б). Концентрация ИФН-γ значительно увеличивалась на 6 и 12 сутки после трансфекции, затем к 14 сут снижалась и была ниже значения для первичных МСК (р = 0.02) (рис. 2в). ФНО-α не выявлен в культуральной жидкости от первичных МСК и мМСК.
Рис. 2.
Уровни цитокинов, секретируемых первичными и генетически модифицированными МСК in vitro: ИЛ-2 (а), ИЛ-6 (б) и ИФН-γ (в). Результаты представлены как среднее ± SD, рассчитанные по трем повторам. *p < 0.05 по сравнению с МСК (t-тест).
Гуморальный ответ на иммунизацию различными иммуногенами изучали через 14 сут после второй иммунизации, а также через 10 сут после заражения ВПГ-1. В результате двукратной иммунизации (рис. 3а) титр антител к ВПГ-1 в группе 1 (МСК) был относительно невысоким (1 : 200), их максимальная продукция выявлена у мышей группы 4, которых иммунизировали иВПГ (титр 1 : 14 080). У мышей группы 2 (МСКgD) содержание антител было статистически значимо выше, чем в группе 3 (ДНКgD): титры 1 : 6400 против 1 : 1600 (р < 0.001). После иммунизации в группах 1 (МСК) и 4 (иВПГ) преобладали антитела субкласса IgG1 (рис. 3б), тогда как в группах 2 и 3, иммунизированных ДНК или мМСК, соотношение антител субклассов IgG2a к IgG1 было значительно выше. После заражения у животных, иммунизированных иВПГ, титры антител снизились, а в остальных группах, особенно в группе 1, значительно возросли.
Рис. 3.
Гуморальный иммунный ответ на иммунизацию до и после заражения мышей летальными дозами ВПГ-1. a – Титры противовирусных антител (ИФА); б – относительные уровни ВПГ-1-специфичных антител субклассов IgG у иммунизированных животных: группа 1 (МСК), группа 2 (МСКgD), группа 3 (ДНКgD), группа 4 (иВПГ) и группа 5 (контроль). Результаты ИФА представлены как значения оптической плотности при 450 нм (OD450). §Соотношение IgG2a/IgG1. в – Титры вируснейтрализующих антител (метод биологической нейтрализации); *p < 0.05 между группами (t-test); #p < 0.05 по сравнению со всеми группами (t-тест).
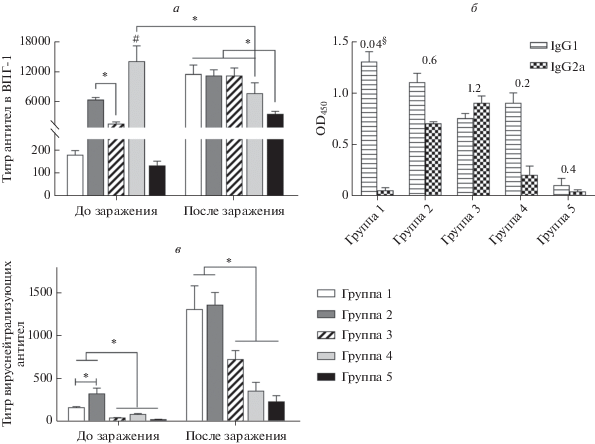
Что касается нейтрализующей активности противовирусных антител, вырабатываемых двукратно иммунизированными животными, их титры в группах 1 и 2 (МСК и мМСК соответственно) были невысокими, но статистически значимо выше, чем в других группах (р < 0.05). Такую же тенденцию, но при гораздо более высоких титрах антител, выявили в сыворотках крови зараженных животных.
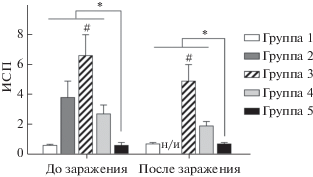
Клеточный ответ изучали в реакции бласттрансформации лимфоцитов in vitro. Для этого спленоциты, выделенные из селезенок мышей, обрабатывали иВПГ для стимуляции специфического ответа, ConA использовали как положительный контроль; ростовую среду, в которой культивировали спленоциты без стимуляторов, – для оценки спонтанной (неспецифической) пролиферации. Установлено (табл. 1), что число ВПГ-1-специфических бластов было статистически значимо выше во всех иммунизированных группах по сравнению с контролем (группа 5, р < < 0.05), при этом максимальное количество бластов стимулировала ДНК-иммунизация (группа 3). Значения ИСП (рис. 4) для этой группы: 6.6 до заражения и 4.9 после заражения ‒ значимо отличались от ИСП в других группах (p < 0.05). Следует отметить, что уровень спонтанной пролиферации спленоцитов у животных, иммунизированных первичными МСК (группа 1), как до заражения, так и после, был статистически значимо выше по сравнению с остальными изученными группами (р < < 0.05) (табл. 1).
Таблица 1.
Пролиферативная активность спленоцитов в ответ на введенные иммуногены до и после заражения животных летальными дозами ВПГ-1
| Группа | Число бластовa | |||||
|---|---|---|---|---|---|---|
| до заражения | после заражения | |||||
| стимуляторы | ||||||
| ConA | иВПГ | Рсb | ConA | иВПГ | Рс | |
| 1 (МСК) |
135 ± 6 | 32 ± 12 | 50 ± 17c | 271 ± 76 | 92 ± 18 | 138 ± 28c |
| 2 (МСКgD) |
132 ± 6 | 54 ± 5 | 14 ± 2 | н/иd | ||
| 3 (ДНКgD) |
122 ± 12 | 58 ± 4 | 9 ± 2 | 252 ± 41 | 130 ± 24 | 27 ± 8 |
| 4 (иВПГ) |
118 ± 8 | 31 ± 4 | 12 ± 3 | 238 ± 36 | 67 ± 20 | 35 ± 19 |
| 5 (контроль) |
125 ± 19 | 9 ± 2c | 8 ± 3 | 212 ± 52 | 22 ± 7c | 25 ± 9 |
Рис. 4.
Индекс стимуляции пролиферации спленоцитов до и после заражения мышей летальными дозами ВПГ-1. Результаты представлены как среднее ± SD, рассчитанные по трем повторам у иммунизированных животных: группа 1 (МСК), группа 2 (МСКgD), группа 3 (ДНКgD), группа 4 (иВПГ) и группа 5 (контроль); н/и – не исследовано. *p < 0.05 между группами (t-test); #p < 0.05 по сравнению со всеми группами (t-тест).
Цитокины ИЛ-6, ИФН-γ и ФНО-α определяли в сыворотках иммунизированных мышей в группах 1, 2, 3 и 5 до и после заражения ВПГ-1. Как видно из данных, представленных в табл. 2, после иммунизации уровень ИФН-γ был значительно выше в группе 1 (МСК) по сравнению с остальными группами (р < 0.05), а в группе 2 (мМСК) превышал таковые для групп 3 (ДНКgD) и 5 (контроль) (р < 0.05). После заражения уровень ИФН-γ в группе 1 оставался максимальным, хотя в группе 3 его концентрация возросла в 23 раза, а в контроле (группа 5) в 12 раз. Мы не исследовали уровни цитокинов у животных группы 4, так как согласно литературным данным иммунизация иВПГ без адъювантов не стимулирует значимого повышения уровня ИФН-γ или количества Т-клеток, продуцирующих этот цитокин [24, 25].
Таблица 2.
Уровни провоспалительных цитокинов в сыворотках иммунизированных мышей до и после заражения ВПГ-1
| Группа | Концентрация цитокина, медиана [минимум; максимум], пг/мл | |||||
|---|---|---|---|---|---|---|
| до заражения | после заражения ВПГ-1 | |||||
| ИЛ-6 | ИФН-γ | ФНО-α | ИЛ-6 | ИФН-γ | ФНО-α | |
| 1 (МСК) |
11 [4; 68 ] |
209a [98; 396] |
24 [4; 93 ] |
13 [4; 55 ] |
236a [103; 468] |
38 [12; 120] |
| 2 (МСКgD) |
20 [4; 83 ] |
63b [35; 132] |
11 [4, 20] |
н/иc | н/и | н/и |
| 4 (иВПГ) |
н/ис | |||||
| 3 (ДНКgD) |
<4a | 4 [1, 18] |
19 [10;55 ] |
<4a | 93 [36;129] |
10a [8, 25] |
| 5 (контроль) |
<4a | 2 [1, 6] |
5a [1, 11] |
14 [4; 67 ] |
48 [1; 98] |
28 [1; 102] |
Уровень ФНО-α до заражения был статистически значимо меньше в группе 5 (контроль) по сравнению с опытными группами (р < 0.05), у иммунизированных животных концентрации этого цитокина статистически значимо не различались (р > 0.05). После заражение у мышей группы 3 наблюдали снижение уровня ФНО-α почти в 2 раза, однако различия не были статистически значимыми (р > 0.05). Следует отменить, что концентрация цитокина в этой группе исходно была значительно ниже, чем в других группах (р < 0.05).
До заражения уровни ИЛ-6 были низкими в группах 1 и 2 и недетектируемыми в группах 3 и 5. У мышей группы 3 ИЛ-6 в сыворотках крови практически отсутствовал как до, так и после заражения.
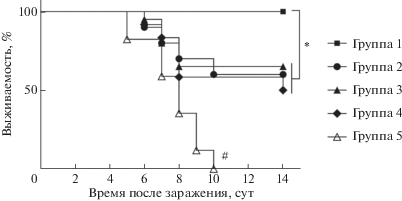
Протективный эффект, индуцируемый различными иммуногенами, изучали в течение 14 суток после заражения ВПГ-1 иммунизированных мышей на модели генерализованной инфекции. В контрольной группе характерные признаки герпетической инфекции с поражением центральной нервной системы: вялость, взъерошенность шерсти, парезы задних конечностей, судороги ‒ наблюдали с пятых суток после заражения, а к 10 суткам все животные погибли (рис. 5). Выживаемость всех иммунизированных групп была статистически значимо выше по сравнению с этим показателем в контроле (р < 0.05). В группе 1 (МСК) у двух мышей на 6 сут после заражения появились неврологические симптомы, однако к 14 суткам все животные выжили (100%-ная выживаемость). При введении мМСК (группа 2) выжило 60% мышей, при ДНК-иммунизации (группа 3) ‒ 65% (13/20), при иммунизации иВПГ (группа 4) – 50%. В результате проведенного статистического анализа показано, что выживаемость животных в группе 1 была значимо выше, чем в других группах (р < < 0.05), и напрямую коррелировала с высокой концентрацией ИФН-γ (табл. 2) и титром вируснейтрализующих антител (рис. 3) (r = 0.93/0.79 соответственно). Таким образом, можно заключить, что двукратная внутривенная инъекция первичных МСК защищает 100% животных от заражения 15 летальными дозами ВПГ-1.
Рис. 5.
Протективный ответ при экспериментальной генерализованной ВПГ-1-инфекции у мышей, иммунизированных МСК (группа 1), мМСК (группа 2), ДНКgD (группа 3) или иВПГ (группа 4), и в контрольной группе (группа 5). *p < 0.05, #p < 0.05 выживаемость в контроле по сравнению с иммунизированными группами (критерий Каплана‒Мейера).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Широкое распространение герпесвирусных инфекций на фоне использования вирусспецифической терапии означает, что для контроля этих заболеваний необходимо применять вакцины. Однако 60-летние исследования в этой области пока не увенчались успехом. Из этого следует, что необходимо внедрение новых стратегий для решения проблем, связанных с заболеваниями герпесвирусной этиологии. Нами исследована возможность использования МСК в качестве кандидатной профилактической вакцины против ВПГ-инфекции. МСК – гетерогенная популяция стволовых стромальных клеток, которая может быть получена в большом количестве из разных доступных источников: костного мозга, жировой ткани, пупочного канальца, зубной пульпы, менструальной крови и даже из кожи. Мультипотентность, иммунорегуляторные и регенеративные свойства МСК, а также отсутствие иммуногенности ‒ эти характеристики инициировали исследования по использованию МСК в терапии заболеваний разной этиологии. Доказана безопасность терапии МСК при их внутривенном введении [27]. Такие международные организации, как Европейское агентство по лекарственным средствам (EMA) и Управление по контролю за пищевыми продуктами и лекарственными средствами США (FDA), одобрили использование некоторых продуктов на основе МСК для лечения болезни Крона и посттрансплантационных осложнений типа “трансплантат против хозяина” [28, 29].
В работах, направленных на создание вакцин против герпеса, с использованием моделей ВПГ-инфекции был сделан вывод, что между “иммунитетом и защитой” нет прямой корреляции [30], имея в виду, что высокий иммунный ответ не может быть гарантией протективного действия. Учитывая эти выводы, мы иммунизировали мышей выбранными иммуногенами и оценивали профилактический эффект in vivo – после заражения животных летальными дозами ВПГ-1. Для сравнения использовали: 1) первичные (нативные) МСК; 2) МСК, трансфицированные плазмидой, содержащей ген Us6, кодирующий белок gD ВПГ; 3) рекомбинантную ДНК, содержащую ген Us6 ВПГ, и 4) цельновирионный инактивированный ВПГ-1. Следует отметить, что два последних иммуногена входят в состав кандидатных вакцин, проходящих испытания в настоящее время: ДНК-вакцина, стимулирующая клеточный ответ [31]; инактивированная вирусная вакцина, предотвращающая инфицирование ВПГ-2 и заболевание морских свинок [32]. В результате проведенных экспериментов нами показано, что полную защиту от летальной ВПГ-1-инфекции обеспечивает двукратное введение первичных МСК, все другие иммуногены оказались менее эффективными – защищали от гибели 50‒65% животных (рис. 5).
Представляло интерес сравнить параметры иммунного ответа животных. В результате иммунизации (до заражения) иВПГ (группа 4) стимулировал антительный ответ к ВПГ-1 и слабый клеточный ответ, тогда как ДНКgD (группа 3) – слабый гуморальный и сильный Т-клеточный ответ. Эти данные согласуются с результатами других авторов, полученными ранее на разных экспериментальных моделях при сравнении профилактического действия субъединичных/вирусных и ДНК-вакцин (см. обзор [33]). Первичные МСК после двукратного введения индуцировали продукцию ВПГ-1-специфических антител, правда в невысоких титрах, и стимулировали врожденный иммунитет, судя по высокому уровню ИФН-γ. мМСК, экспрессирующие ген Us6 ВПГ, индуцировали как антительный ответ на вирусный антиген, так и клеточный иммунный ответ. В то же время при иммунизации мМСК титр антител был ниже, чем при иммунизации иВПГ, и уровень пролиферации лимфоцитов (ИСП) был ниже, чем при ДНК-иммунизации (ДНКgD). После заражения антительный ответ возрос у всех животных, кроме группы 4, иммунизированной иВПГ, в которой уровень антител снизился. Важно, что у животных, иммунизированных МСК и мМСК, титры вируснейтрализующих антител были значительно выше, чем в других группах. Самые большие различия между группами обнаружены в концентрации ИФН-γ, которая оказалась максимальной в крови животных, иммунизированных первичными МСК. Следует обратить внимание на тот факт, что после иммунизации первичными МСК отсутствовал статистически значимый пролиферативный ответ Т-клеток на ВПГ-1 на фоне повышенного уровня спонтанного (неспецифического) ответа ‒ как до, так и после заражения ВПГ-1. В то же время в обоих случаях зарегистрирован высокий уровень продукции ИФН-γ (табл. 1 и 2). Неспецифический ответ Т-клеток после двукратного введения МСК, возможно, связан с функциональными особенностями этих клеток, генерацию и пролиферацию которых индуцируют цитокины окружающей среды, причем независимо от чужеродного антигена [34]. Эти клетки имеют фенотип Т-клеток памяти или подобных им – “memory-like”. Показано, что ИФНы I типа (ИФН-α/β), продуцируемые при гомеостатических условиях или при инфекции, способствуют пролиферации и функционированию “memory-like” T-клеток CD8+ [35]. Т-клетки с более широкой специфичностью ‒ с неспецифическими, но полезными эффектами ‒ могут значительно улучшить иммунный ответ, что следует учитываться при создании вакцин ‒ как составляющих гетерологичного иммунного ответа [34].
Следует отметить, что антитела, обнаруженные после иммунизации первичными МСК (группа 1), были в основном IgG1-субтипа (маркер Тh2-ответа) и соотношение противовирусных антител субтипов IgG2a (маркер Th1-ответа)/IgG1 в этой группе было минимальным. Согласно сложившимся представлениям, это означает, что иммунный ответ смещен в сторону Тh2-клеток и направлен на синтез противовоспалительных цитокинов (ИЛ-4, ИЛ-10 и др.). В результате проведенных экспериментов нами выявлены высокие концентрации провоспалительных цитокинов, особенно ИФН-γ, продуцируемого главным образом клетками Th1-фенотипа. Можно предположить, что это “противоречие” объясняется тем, что МСК секретируют большой набор как про-, так и противовоспалительных цитокинов [36], которые способны активировать широкий спектр иммунных клеток. Кроме того, индуцируемые МСК иммунные клетки могут модифицировать тип ответа: “гуморальный” Th2 на “клеточный” Th1 и стимулировать продукцию ИФН-γ [37]. По-видимому, благодаря перечисленным свойствам МСК, после заражения животных, ими иммунизированных, активизируется как Т-клеточный, так и В-клеточный ответ к ВПГ-1, а также формируется защита от летальной инфекции. Более мощный защитный эффект первичных МСК по сравнению с мМСК может быть обусловлен тем, что в результате трансфекции и последующего отбора клеток, экспрессирующих введенный ген Us6 ВПГ, в мМСК изменяется in vitro секреция провоспалительных цитокинов: значительно снижается продукция ИФН-γ и ИЛ-2 и существенно повышается уровень ИЛ-6 (рис. 2). Введение селектированных мМСК приводило к изменению концентрации цитокинов также и в крови иммунизированы мышей: приблизительно в 2 раза снижалась концентрация ИФН-γ и в 2 раза возрастала концентрация ИЛ-6. Повышение уровня ИЛ-6 регистрируют при различных воспалительных заболеваниях и рассматривают как маркер развития утяжеленных форм заболеваний, в том числе герпесвирусной этиологии [38] и при COVID-19 [39]. По-видимому, такие изменения в цитокиновом паттерне вносят вклад в снижение протективной активности мМСК-иммуногена по сравнению с МСК на экспериментальной модели ВПГ-1-инфекции, так как при вакцинации первичными МСК, индуцирующими 100%-ную выживаемость мышей, концентрация ИЛ-6 не изменялась, а ИФН-γ возрастала.
В большинстве опубликованных работ описаны иммуносупрессорные свойства МСК (см. обзор [37]), благодаря которым они подавляют избыточные воспалительные реакции, цитокиновый шторм, отторжение трансплантата. В других исследованиях показано, что МСК обладают скорее иммунорегуляторными, чем иммуносупрессорными свойствами и могут не только подавлять, но и стимулировать иммунные реакции [40]. В организме МСК присутствуют практически во всех тканях, экспрессируют множество типов рецепторов, поэтому различия в направленности выполняемых ими регуляторных функций можно объяснить различиями в их микроокружении [41]. В условиях воспаления, при высоких уровнях провоспалительных цитокинов ИФН-γ и ФНО-α, МСК активируются и приобретают иммуносупрессорный фенотип – МСК2. При отсутствии в микроокружении признаков воспаления (низкие уровни ФНО-α и ИФН-γ) МСК приобретают провоспалительный, МСК1, фенотип и усиливают Т-клеточный ответ. Активацию МСК изучали в динамике ‒ после добавления poly(I:C) как аналога двухцепоцечной вирусной РНК [42]. Авторы установили, что свойства МСК в ходе активации могут изменяться с провоспалительных на противовоспалительные, и определили это как зависимость поляризации от времени действия МСК – “time-dependent MSC polarization”. В проведенных нами экспериментах МСК вводили здоровым животным, и это было одно микроокружение, а после заражения вирусом микроокружение МСК изменилось, что привело к стимуляции В-клеточного ответа и, особенно важно, к значительному повышению уровня вируснейтрализующих антител. Эти результаты хорошо согласуются с данными, полученными Glenn & Whartenby [43] при изучении действия МСК на В-клетки in vitro. В условиях, имитирующих вирусную инфекцию, в присутствии МСК повышались выживаемость В-клеток, их дифференцировка и уровни продуцируемых ими антител.
Заражение мышей ВПГ-1 после иммунизации МСК, по-видимому, запускает множество механизмов, включая как антигензависимые, так и антигеннезависимые. Молекулярные и клеточные механизмы, которые в этих условиях обеспечивают противовирусное действие, активно изучают [44‒46] и, можно надеется, будут раскрыты в дальнейших исследованиях.
Согласно последним данным, успешное терапевтическое действие МСК может быть длительным [47], что предполагает индукцию Т-клеток памяти, необходимых для протективного действия вакцины [48]. Можно предположить, что после двукратной иммунизации МСК в организме мышей сформировались клетки памяти, функционирование которых способствовало полной защите животных от последующей летальной вирусной инфекции.
Таким образом, благодаря уникальной способности МСК индуцировать врожденный, адаптивный и протективный иммунитет, а также их доступности и безопасности, эти клетки в перспективе можно рассматривать в качестве главных компонентов вакцин нового поколения (на основе клеточных технологий) против герпетических и других инфекций.
Работа выполнена при финансовой поддержке гранта Российского фонда фундаментальных исследований (№ 19-04-01218).
Все исследования выполняли согласно “Пpавилам пpоведения pабот с иcпользованием экспериментальных животных” (приказ № 266 Минздрава России от 19.06.2003).
Авторы заявляют об отсутствии конфликта интересов.
Список литературы
Looker K.J., Magaret A.S., May M.T., Turner K.M., Vickerman P., Gottlieb S.L. (2015) Global and regional estimates of prevalent and incident herpes simplex virus type 1 infections in 2012. PLoS One. 10(10), e0140765.
Looker, K. J., Magaret A.S., Turner K.M., Vickerman P., Newman L.M. (2015) Global estimates of prevalent and incident herpes simplex virus type 2 infections in 2012. PLoS One. 10, e114989.
Burn Aschner C., Knipe D.M., Herold B.C. (2020) Model of vaccine efficacy against HSV-2 superinfection of HSV-1 seropositive mice demonstrates protection by antibodies mediating cellular cytotoxicity. NPJ. Vaccines. 5, 35.
Gottlieb S.L., Giersing B., Boily M.C., Chesson H., Looker K.J., Schiffer J., Spicknall I., Hutubessy R., Broutet N.; WHO HSV Vaccine Impact Modelling Meeting Working Group. (2019) Modelling efforts needed to advance herpes simplex virus (HSV) vaccine development: key findings from the World Health Organization consultation on HSV vaccine impact modelling. Vaccine. 37(50), 7336‒7345.
Looker K.J., Welton N.J., Sabin K.M., Dalal S., Vickerman. P, Turner K.M.E., Boily M.C., Gottlieb S.L. (2020) Global and regional estimates of the contribution of herpes simplex virus type 2 infection to HIV incidence: a population attributable fraction analysis using published epidemiological data. Lancet Infect. Dis. 20(2), 240‒249.
James SH, Kimberlin DW. (2015) Neonatal herpes simplex virus infection: epidemiology and treatment. Clin. Perinatol. 42, 47‒59.
Tsatsos M., MacGregor C., Athanasiadis I., Moschos M.M., Hossain P., Anderson D. (2016) Herpes simplex virus keratitis: an update of the pathogenesis and current treatment with oral and topical antiviral agents. Clin. Exp. Ophthalmol. 44, 824–837.
Whitley R., Baines J. (2018) Clinical management of herpes simplex virus infections: past, present, and future. F1000Res. 7, F1000 Faculty Rev-1726. https://doi.org/10.12688/f1000research.16157.1
Gottlieb S.L., Giersing B.K., Hickling J., Jones R., Deal C., Kaslow D.C. (2019) Meeting report: initial World Health Organization consultation on herpes simplex virus (HSV) vaccine preferred product characteristics, March 2017. Vaccine. 37(50), 7408‒7418.
Looker K.J., Johnston C., Welton N.J., James. C, Vickerman P., Turner K.M.E., Boily M.C., Gottlieb S.L. (2020) The global and regional burden of genital ulcer disease due to herpes simplex virus: a natural history modelling study. BMJ Glob. Health. 5(3), e001875.
Johnston C., Gottlieb S.L., Wald A. (2016) Status of vaccine research and development of vaccines for herpes simplex virus. Vaccine. 34(26), 2948‒2952.
Ghaffarifar F. (2018) Plasmid DNA vaccines: where are we now? Drugs Today (Barc.). 54(5), 315‒333.
Ike A.C., Onu C.J., Ononugbo C.M., Reward E.E., Muo S.O. (2020) Immune response to herpes simplex virus infection and vaccine development. Vaccines (Basel). 8(2), 302.
Egan K., Hook L.M., Latourette P., Desmond A., Awasthi S., Friedman H.M. (2020) Vaccines to prevent genital herpes. Transl. Res. 220, 138‒152.
Spees J.L., Lee R.H., Gregory C.A. (2016) Mechanisms of mesenchymal stem/stromal cell function. Stem Cell Res. Ther. 7(1), 125.
Han J., Liu Y., Liu H., Li Y. (2019) Genetically modified mesenchymal stem cell therapy for acute respiratory distress syndrome. Stem Cell Res. Ther. 10(1), 386.
Fu X., Chen Y., Xie F.N., Dong P., Liu W.B., Cao Y., Zhang W.J., Xiao R. (2015) Comparison of immunological characteristics of mesenchymal stem cells derived from human embryonic stem cells and bone marrow. Tissue Eng. Part A. 21(3‒4), 616‒626.
Cagliani J., Grande D., Molmenti E.P., Miller E.J., Rilo H.L.R. (2017) Immunomodulation by mesenchymal stromal cells and their clinical applications. J. Stem Cell Regen. Biol. 3(2), .https://doi.org/10.15436/2471-0598.17.022
Sage E.K., Thakrar R.M., Janes S.M. (2016) Genetically modified mesenchymal stromal cells in cancer therapy. Cytotherapy. 18(11), 1435‒1445.
Jibin Han, Yuxiang Liu, Hong Liu, Yuanyuan Li. (2019) Genetically modified mesenchymal stem cell therapy for acute respiratory distress syndrome. Stem Cell Res. Ther. 10(1), 386.
Tomchuck S.L., Norton E.B., Garry R.F., Bunnell B.A., Morris C.A., Freytag L.C., Clements J.D. (2012) Mesenchymal stem cells as a novel vaccine platform. Front. Cell. Infect. Microbiol. 2, 140.
Masalova O.V., Lesnova E.I., Klimova R.R., Momotyuk E.D., Kozlov V.V., Ivanova A.M., Payushina O.V., Butorina N.N., Zakirova N.F., Narovlyansky A.N., Pronin A.V., Ivanov A.V., Kushch A.A. (2020) Genetically modified mouse mesenchymal stem cells expressing non-structural proteins of hepatitis C virus induce effective immune response. Vaccines (Basel). 8(1), 62.
Климова Р.Р., Момотюк Е.Д., Демидова Н.А., Ярыгина Е.И., Кущ А.А. (2018) Мезенхимальные стволовые клетки: усиление иммунного ответа и защита мышей от летальной герпесвирусной инфекции. Вопросы вирусологии. 63(6), 261‒267.
Roizman B., Knipe D.M., Whitley R.J. (2007) Herpes simplex viruses. In: Fields Virology, 5th edition. Eds Knipe D.M., Howley P.M. Lippincott Philadelphia: Williams & Wilkins, 2501–2601.
Morello C.S., Kraynyak K.A., Levinson M.S., Chen Z., Lee K.-F., Spector D.H. (2012) Inactivated HSV-2 in MPL/alum adjuvant provides nearly complete protection against genital infection and shedding following long term challenge and rechallenge. Vaccine. 30(46), 6541‒6550.
Dominici M., Le Blanc K., Mueller I., Slaper-Cortenbach I., Marini F., Krause D., Deans R., Keating A., Prockop Dj., Horwitz E. (2006) Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy. 8(4), 315‒317.
Thompson M., Mei S.H.J., Wolfe D., Champagne J., Fergusson D., Stewart D.J. (2020) Cell therapy with intravascular administration of mesenchymal stromal cells continues to appear safe: an updated systematic review and meta-analysis. EClinicalMedicine. 19, 100249. https://doi.org/10.1016/j.eclinm.2019.100249
Romano B., Elangovan S., Erreni M., Sala E., Petti L., Kunderfranco P. (2019) TNF-stimulated gene-6 is a key regulator in switching stemness and biological properties of mesenchymal stem cells. Stem Cells. 37(7), 973‒987.
Saldanha-Araujo F., Garcez E.M., Silva-Carvalho A.E., Carvalho J.L. (2020) Mesenchymal stem cells: a new piece in the puzzle of COVID-19 treatment. Front. Immunol. 11, 1563.
Khodai T., Chappell D., Christy C., Cockle P., Eyles J., Hammond D., Gore K., McCluskie M.J., Evans D.M., Lang S., Loudon P.T., Townend T., Wright P., West K., Bright H. (2011) Single and combination herpes simplex virus type 2 glycoprotein vaccines adjuvanted with CpG oligodeoxynucleotides or monophosphoryl lipid A exhibit differential immunity that is not correlated to protection in animal models. Clin. Vaccine Immunol. 18(10), 1702‒1709.
Dutton J.L., Woo W.P., Chandra J., Xu Y., Li B., Finlayson N., Griffin P., Frazer I.H. (2016) An escalating dose study to assess the safety, tolerability and immunogenicity of a herpes simplex virus DNA vaccine COR-1. Hum. Vaccin. Immunother. 12, 3079–3088.
Hensel M.T., Marshall J.D., Dorwart M.R., Heeke D.S., Rao E., Tummala P., Yu L., Cohen G.H., Eisenberg R.J., Sloan D.D. (2017) Prophylactic herpes simplex virus 2 (HSV-2) vaccines adjuvanted with stable emulsion and Toll-like receptor 9 agonist induce a robust HSV-2-specific cell-mediated immune response, protect against symptomatic disease, and reduce the latent viral reservoir. J. Virol. 91(9), e02257-16.
Ike A.I., Onu C.J., Ononugbo C.M., Reward E.E., Muo S.O. (2020) Immune response to herpess virus infection and vaccine development. Vaccines (Basel). 8(2), 302.
Lauvau G., Goriely S. (2016) Memory CD8+ T cells: orchestrators and key players of innate immunity? PLoS Pathog. 12(9), e1005722.
Martinet V., Tonon S., Torres D., Azouz A., Nguyen M., Kohler A. (2015) Type I interferons regulate eomesodermin expression and the development of unconventional memory CD8(+) T cells. Nat. Commun. 6, 7089.
Tomchuck S.L., Zwezdaryk K.J., Coffelt S.B., Waterman R.S., Danka E.S., Scandurro A.B. (2008) Toll-like receptors on human mesenchymal stem cells drive their migration and immunomodulating responses. Stem Cells. 26(1), 99‒107.
Shammaa R., El-Kadiry A.E., Abusarah J., Rafei M. (2020) Mesenchymal stem cells beyond regenerative medicine. Front. Cell Dev. Biol. 8, 72.
Khazan M., Nasiri S., Riahi S.M., Robati R.M., Hedayati M. (2020) Measurement of melatonin, indole-dioxygenase, IL-6, IL-18, ferritin, CRP, and total homocysteine levels during herpes zoster. J. Med. Virol. 92(8), 1253‒1259.
Henry B.M., Santos de Oliveira M.C., Benoit S., Plebani M., Lippi G. (2020) Hematologic, biochemical and immune biomarker abnormalities associated with severe illness and mortality in coronavirus disease 2019 (COVID-19): a meta-analysis. Clin. Chem. Lab. Med. 58(7), 1021‒1028.
Najar M., Raicevic G., Fayyad-Kazan H., Bron D., Toungouz M., Lagneaux L. (2016) Mesenchymal stromal cells and immunomodulation: a gathering of regulatory immune cells. Cytotherapy. 18(2), 160‒171.
Waterman R.S, Tomchuck S.L., Henkle S.L., Betancourt A.M. (2010) A new mesenchymal stem cell (MSC) paradigm: polarization into a pro-inflammatory MSC1 or an immunosuppressive MSC2 phenotype. PLoS One. 5, e10088.
Petri R.M., Hackel A., Hahnel K., Dumitru C.A., Bruderek K., Flohe S.B., Paschen A., Lang S., Brandau S. (2017) Activated tissue-resident mesenchymal stromal cells regulate natural killer cell immune and tissue-regenerative function. Stem Cell Reports. 9(3), 985‒998.
Glenn J.D., Whartenby K.A. (2014) Mesenchymal stem cells: emerging mechanisms of immunomodulation and therapy. World J. Stem Cells. 6(5), 526‒539.
Rasmussen S.B., Sørensen L.N., Malmgaard L., Ank N., Baines J.D., Chen Z.J., Paludan S.R. (2007) Type I interferon production during herpes simplex virus infection is controlled by cell-type-specific viral recognition through Toll-like receptor 9, the mitochondrial antiviral signaling protein pathway, and novel recognition systems. J. Virol. 81(24), 13315‒13324.
Suarez-Ramirez J.E., Tarrio M.L., Kim K., Demers D.A., Biron C.A. (2014) CD8 T cells in innate immune responses: using STAT4-dependent but antigen-independent pathways to gamma interferon during viral infection. mBio. 5(5), e01978-14.
Welch W. J., Mizzen L. A. (1988) Characterization of the thermotolerant cell. II. Effects on the intracellular distribution of heat-shock protein 70, intermediate filaments, and small nuclear ribonucleoprotein complexes. J. Cell Biol. 106(4), 1117‒1130.
Lopez-Santalla M., Hervas-Salcedo R., Fernandez-Garcia M., Bueren J.A., Garin M.I. (2020) Cell therapy with mesenchymal stem cells induces an innate immune memory response that attenuates experimental colitis in the long-term. J. Crohns Colitis. 14(10), 1424‒1435.
Shin H., Iwasaki A. (2012) A vaccine strategy that protects against genital herpes by establishing local memory T cells. Nature. 491(7424), 463‒467.
Дополнительные материалы отсутствуют.
Инструменты
Молекулярная биология