

Журнал общей биологии, 2019, T. 80, № 1, стр. 68-80
Изучение адаптивной нормы реакции популяций основных лесообразующих видов хвойных в Средней Сибири на основе косвенных данных
И. В. Тихонова 1, *, М. А. Корец 2, 3
1 Западно-Сибириское отделение ИЛ СО РАН,
630082 Новосибирск, ул. Жуковского, 100/1, Россия
2 Институт леса им. В.Н. Сукачева СО РАН,
660036 Красноярск, Академгородок, стр. 50/28, Россия
3 Сибирский федеральный университет,
660041 Красноярск, ул. Киренского, 79, Россия
* E-mail: selection@ksc.krasn.ru
Поступила в редакцию 26.06.2017
После доработки 08.05.2018
Принята к публикации 25.06.2018
Аннотация
С помощью анализа изменчивости метеорологических условий произрастания за последние 50 лет определены климатические пределы нормы адаптивной реакции популяций 5 хвойных видов в Средней Сибири. Выявлены существенные межпопуляционные различия внутри видов по величине межсезонной изменчивости климатических показателей. Установлена видоспецифичная сопряженность плотности заселения территории представителями хвойных видов со среднемноголетними значениями отдельных показателей и с величинами их изменчивости по годам. Наименьшее межпопуляционное разнообразие видов наблюдается при совмещении нескольких климатических характеристик для пихты сибирской и ели сибирской. Результаты проведенного исследования свидетельствуют о необходимости сохранения внутривидовой структуры хвойных для сохранения их внутривидового разнообразия и устойчивости бореальных лесных экосистем. Отмечено, что некоторые популяции видов хвойных из центральной части ареала как и популяции, произрастающие в горах Южной Сибири, обладают потенциальной способностью адаптироваться к новым условиям произрастания при перенесении их на значительное расстояние по широте и долготе.
Одним из достижений науки в ХХ в. было осознание важной средообразующей функции лесов в биосфере (Вернадский, 1940; Протопопов, 1975; Молчанов, 1978; Рахманов, 1984; Davin, Noublet-Ducoudre, 2010). Их состояние в меняющемся климате в условиях растущей антропогенной нагрузки в первую очередь определяется нормой реакции видов. Ввиду большой хозяйственной ценности и экологической значимости хвойных лесов повышение точности прогнозирования их состояния при изменениях климата является необходимым условием неистощимого лесопользования (Кулагин, 1980; Мониторинг биологического разнообразия…, 2008). Это в свою очередь невозможно без установления нормы реакции отдельных популяций внутри видов (Тимофеев-Ресовский и др., 1973; Глотов, Тараканов, 1985; Алтухов, 2004). Их разная способность к акклиматизации была выявлена с помощью закладки серии географических культур хвойных деревьев (Каппер, 1946; Eiche, Andersson, 1974; Ирошников, 1977; Giertych, 1979; Молотков и др., 1982; Jansons, Baumanis, 2005; Шутяев, 2007; Мельник и др., 2009, и др.).
Как известно, изучение нормы реакции видов является одной из основных дискуссионных проблем, решаемых эволюционной биологией (Иогансен, 1933; Шмальгаузен, 1968; Lewontin, 1974; Майр, 1974; Будыко, 1975; Шварц, 1980; Шишкин, 1988; Kirkpatrick, Barton, 1997; Schlichting, Pigliucci, 1998; Петрашов, 2006; Sextonet al., 2009; Hargreaves et al., 2014). Поскольку существуют разные представления о механизмах, обеспечивающих изменение нормы реакции, и разное понимание терминов, дадим краткие определения используемых терминов, опираясь на работу И.И. Шмальгаузена (1968). Под “нормой реакции особи” мы понимаем все потенциально возможные в результате эпигенетических вариаций генотипа модификационные фенотипические изменения особи, под “нормой реакции популяции или вида” – совокупность всех потенциально возможных для всех особей популяции или вида модификационных изменений, связанных с эпигенетическими и рекомбинантными генетическими изменениями, а также нейтральных мутаций, не приводящих к изменению характерной для вида корреляционной системы взаимодействия генетической структуры его генофонда и функций (в широком смысле). “Норма адаптивных реакций особей, популяций, вида” – это соответственно те же изменения, но в пределах ограниченного периода времени с определенными условиями среды. Таким образом, смена “нормы реакции” вида (всего его эколого-генетического потенциала) возможна только наряду с существенными изменениями самого вида, т.е. в результате видообразования, а предлагаемые в литературе механизмы изменения “нормы реакции” видов в результате эпигенеза, рекомбинаций и некоторых мутаций следует отнести к изменениям их “нормы адаптивных реакций”, возможности которых уже заложены в генофонде вида.
Оценивают “нормы адаптивных реакций” популяций, как правило, по прямым признакам, отражающим морфологические, физиологические, поведенческие реакции особей популяций в ответ на изменение внешних и внутренних факторов.
Необходимо отметить, что широко распространенные в Северном полушарии долгоживущие виды хвойных деревьев представляют особый интерес для изучения данной проблемы. Однако с помощью прямых методов исследования популяций древесных растений невозможно охватить значительные по площади и по величине выборки. Для выявления географических закономерностей нормы адаптивных реакций популяций хвойных видов больше подходят косвенные методы, в частности, анализ изменчивости климатических условий произрастания популяций. С их помощью можно примерно оценить искомые характеристики популяций на большой территории. Естественно, чем продолжительнее период наблюдений за погодой, тем надежнее оценки. Так как в данном случае мы имеем дело с экологическими (только климатическими) характеристиками популяций, то для ясности используем другой термин “климатическая норма адаптивных реакций популяции” (КНАР) – пределы климатических изменений за определенный период времени, в которых успешно реализуются адаптивные изменения особей популяции, не приводящие к ее гибели.
В связи с вышесказанным целью проводимых исследований было сравнение климатической нормы адаптивных реакций популяций с помощью анализа пространственно-временной изменчивости метеорологических условий произрастания основных лесообразующих хвойных видов (ели сибирской Picea obovata Ledeb., лиственницы сибирской Larix sibirica Ledeb. с лиственницей Гмелина L. gmelinii (Rupr.) Rupr., сосны сибирской кедровой Pinus sibirica Du Tour., сосны обыкновенной Pinus sylvestris L., пихты сибирской Abies sibirica Ledeb.) в разных природно-климатических зонах Средней Сибири за 1960–2013 гг.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования проводили на территории Средней Сибири в пределах 50–75° с.ш. и 83–129° в.д. Была использована карта “Леса СССР” М 1 : 2 500 000 (1990). Это наиболее полная выполненная по результатам наземных измерений карта, на которой показаны леса таежной зоны, менее поврежденные вырубками и пожарами по сравнению с современными. Для уточнения был использован доработанный в 2013 г. вариант карты растительного покрова России, выполненной по данным спектрорадиометра MODIS М 1 : 2 500 000 (Барталев и др., 2011).
В анализе климатических условий и при построении климатической карты использовали среднемесячные значения приземной температуры воздуха и суммы осадков за 1960–2013 гг. (База данных Всероссийского научно-исследовательского института гидрометеорологической информации (ВНИИГМИ-МЦД)). Из-за недостатка и неравномерности распределения метеостанций по территории за основу была взята база данных UEA (Mitchell, Jones, 2005) в виде матриц экстраполированных значений среднемесячной температуры воздуха с ячейками 0.5° с.ш. × 0.5° в.д., отражающих метеорологические условия России с относительно небольшой погрешностью (Жильцова, Анисимов, 2009). Эти данные уточнялись для исследуемой территории по данным метеостанций ВНИИГМИ-МЦД. Размер ячеек 55.6 км в направлении С–Ю, 27.8–31.9 км в направлении З–В, площадь – 154.6–177.4 тыс. га. Площади, занятые вырубками, гарями, пашнями и населенными пунктами, были исключены из анализа, как и площади под такими природными объектами, как скалы и водоемы. Для каждой ячейки матрицы (выборка составила 3095) были рассчитаны среднемноголетние значения, экстремумы и среднеквадратичные отклонения метеорологических показателей (перечень см. ниже), относительная площадь ячейки, занимаемая каждым из хвойных видов. Матрицы данных были преобразованы в формат растровых изображений ГИС средствами пакета ArcGIS. Территориальную привязку осуществляли с помощью цифровой векторной топоосновы масштаба 1 : 1 000 000 и цифровой модели рельефа (SRTM 90 DEM).
Были использованы следующие метеорологические показатели: среднегодовая температура (Т), годовая сумма осадков (Р), амплитуда среднемесячных температур июля–января (lim), гидротермические коэффициенты за год и за ноябрь месяц, минимумы и максимумы всех показателей. Гидротермические коэффициенты рассчитывали по формулам:
1) гидротермический коэффициент по Де Мартону (Хромов, Мамонтов, 1974) с поправкой на более низкие в Сибири температуры ГТК = Р/(Т + 20), в исходной формуле (Т + 10);
2) гидротермический коэффициент ТГКн = Тн/Рн, за ноябрь для определения степени промерзания почвы в ноябре – количество градусов ниже 0, приходящихся на 1 мм снежного покрова (так как к началу ноября на всей исследуемой территории устанавливается устойчивый снежный покров и отрицательные температуры).
Изменчивость условий произрастания в пространстве и во времени (по годам) оценивали на основании дисперсии (σ) и амплитуды (lim) max–min значений признаков. Данные анализировали с помощью дисперсионного, дискриминантного и кластерного анализов (Боровиков, 2003).
Анализируя данные, мы исходили из того, что показатели изменчивости условий произрастания в одной или нескольких ячейках позволяют исследовать климатическую норму адаптивных реакций видов на уровне отдельных популяций.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Необходимо отметить, что значительная часть лесов Сибири подвержена влиянию хозяйственной деятельности человека, что является основной причиной сокращения числа хвойных видов в пригородных лесах и появления лесостепей в основной части ареалов видов (Леса СССР, 1969, с. 318; Бузыкин, Пшеничникова, 2008). Сокращение площади хвойных лесов на исследуемой территории на 9.2%, отмеченное нами при сравнении карт 1990 и 2013 гг., является частью общей длительной тенденции уменьшения площади лесов в Сибири (Калашников, 1998). Наличие вырубок, гарей, пашни и населенных пунктов несомненно отразилось на статистических показателях распределения популяций по климатическим характеристикам (уменьшилась доля популяций из наиболее благоприятной для хвойных подзоны южной тайги). Однако там, где климат ограничивает распространение хвойных лесов, влияние человека пока невелико, поэтому в анализ вошли все предельные климатические характеристики местообитаний популяций.
Кроме того, на текущее распространение хвойных видов влияет также множество других факторов, таких как история расселения, сукцессии, межвидовая конкуренция (Сочава, 1956; Колесников, 1961; Бобров, 1978; Sextonet al., 2009; Козин, 2013; Hargreaves et al., 2014), что в целом тоже ведет к сокращению площади фактически занимаемых видами местообитаний по сравнению с потенциально возможными. В частности, известно, что перечисленные виды хвойных сформировались 5–33 (до 50) млн лет назад (Бобров, 1978; Разумовский, 1981; Gernandt et al., 2008; Leslie et al., 2012). На территории России хвойные леса в современном их виде существуют с начала голоцена, без существенных изменений – последние несколько тысяч лет (Разумовский, 1981) и, следовательно, пережили несколько периодов похолоданий, в том числе так называемый малый ледниковый период. Известно также, что в ходе сукцессии в лесах европейской части страны до Урала под пологом сосны чаще всего поселяется ель, а восточнее – пихта, либо пихта в смеси с елью и кедром, что также, по-видимому, является следствием разной послеледниковой истории расселения видов в европейской и азиатской частях ареалов. Поэтому полученные диапазоны климатических норм адаптивных реакций для популяций видов и их распределения могут отличаться от потенциально возможных в современном климате, не сокращенных влиянием других факторов, отделить и компенсировать которое в настоящее время не представляется возможным не только для исследуемых, но и для множества других видов живых организмов. Как справедливо считают некоторые исследователи (Lewontin, 1974; Глотов, Тараканов, 1985), даже если мы не в состоянии охватить все популяции, все генотипы и все возможные проявления генотипов вида, нужно искать разные подходы к решению проблемы и при ограниченных возможностях.
Сравнивая статистические величины изменчивости метеорологических показателей, отметим, что дисперсии (σ) отражают достаточно регулярную климатическую норму адаптивных реакций популяций хвойных видов (КНАР1), произрастающих в данных условиях, а пределы экстремумов (max–min, в отдельные годы) – нерегулярную (КНАР2). Поэтому первые можно использовать в прогнозировании способности популяций видов адаптироваться к постепенным изменениям климата, а вторые – к быстрым изменениям, не выходящим за норму реакции видов. Так, например, для хвойных лесов в целом интервал среднемноголетних значений температуры (lim Т) составил 17°С, а с учетом экстремумов – 23°С; по сумме осадков за год (lim Р) соответственно 1010 и 1400 мм; по ГТК – 9.7 и 41.0; по ТГКн – 5.3 и 31.2. В основной части ареала пяти хвойных видов на территории Средней Сибири там, где площадь хвойных лесов уменьшилась больше всего, те же показатели варьировали в следующих пределах: среднемноголетние Т –6.5…1.2°С (в отдельные годы до –8.9…2.9°С), Р – 360–850 мм (в отдельные годы до 270–1250 мм, но без ГТК не используется), ГТК от 2.1 до 5.8 (с экстремумами по годам от 1.3 до 8.9), ТГКн – от –1.7 до –0.1 (в разные годы от –5.7 до –0.01).
Популяции хвойных видов на исследуемой территории распределились следующим образом по среднемноголетним значениям (СМ) климатических показателей (рис. 1) и по КНАР (рис. 2, 3). Значения КНАР1 (σ) по Т в местообитаниях популяций варьировали в пределах 0.8–1.7°С, по ГТК – 0.3–9.8, по ТГКн – 0.15–7.8. КНАР2 по Т (амплитуды значений – lim Т) в местах произрастания отдельных популяций изменялись от 3.4 до 7.0°С. По этому показателю популяции внутри видов различались в 1.7–2 раза. Так же в 2–4 раза местообитания популяций различались по средним амплитудам Рвег (вегетационного сезона, май–август) (74–255 мм). ГТК в местах произрастания популяций темнохвойных видов изменялся в пределах 1.8–7.8, у сосны обыкновенной – 1.5–6.4, у видов лиственницы – 1.7–8.5 (до 37.3 в горных популяциях субарктики). Наибольшие различия между популяциями по КНАР2 наблюдались по амплитудам ГТК – 1.3–34.3 (в популяциях внутри видов в 3–23 раза) и ТГКн – 0.6–33.2 (соответственно в 4–35 раз). При этом виды лиственницы L. sibirica и L. gmelinii (самостоятельность которых на наш взгляд справедливо ставится под сомнение (Авров, 1998) из-за небольших морфологических и генетических различий (следует также из работ (Semerikov al., 1999; Семериков и др., 2007; Polezhaeva et al., 2010, и др.), отличались не только наибольшими пределами СМ значений показателей тепло- и влагообеспеченности популяций, но и наибольшими амплитудами их изменчивости в популяциях по годам (КНАР1 и КНАР2). Очевидно, что такое расширение климатической нормы реакций стало возможным благодаря переходу к листопадности у видов р. Larix.
Рис. 1.
Распределение популяций пихты сибирской (1), сосны сибирской кедровой (2), ели сибирской (3), сосны обыкновенной (4), видов лиственницы (5) по Т (а), ГТК (б), ТГКн (в), ТГКн min (г).
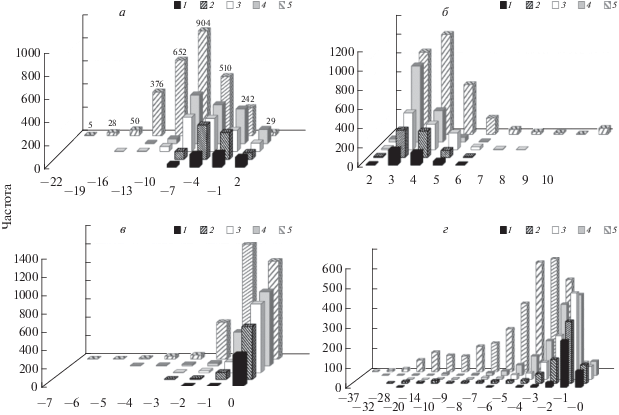
Рис. 2.
Распределение частот встречаемости популяций по величине КНАР1 (σ): пихты сибирской (1), сосны сибирской кедровой (2), ели сибирской (3), сосны обыкновенной (4), видов лиственницы (5), определенных по: Т (а), ГТК (б), ТГКн (в).
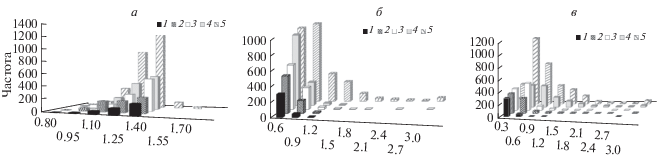
Рис. 3.
Распределение по величине КНАР2 (lim) частот встречаемости популяций пихты сибирской (1), сосны сибирской кедровой (2), ели сибирской (3), сосны обыкновенной (4), видов лиственницы (5), определенных по: Т (а), ГТК (б), ТГКн (в).
На втором месте по величине КНАР была Picea obovata, которая характеризовалась высокими внутривидовыми пределами СМ значений Т и ГТК и высокими амплитудами популяционных значений КНАР1 и КНАР2 по всем учтенными показателям. Для Pinus sylvestris отмечен широкий диапазон средних значений Т и ТГКн и более узкий – ГТК, и широкие интервалы популяционных значений КНАР1 и КНАР2 по ТГКн. Меньшей климатической нормой адаптивных реакций отличались популяции Pinus sibirica и особенно Abies sibirica. В то же время необходимо отметить, что небольшая часть популяций Pinus sibirica и отдельные популяции Abies sibirica приспособлены к выживанию в крайне суровых условиях произрастания, где ТГКн в отдельные годы опускается до –20…–29 (рис. 1г, 2в, 3в), с широкими КНАР1 и КНАР2. Более высокие значения F-критерия для Picea obovata, Pinus sylvestris и особенно Larix spp. (табл. 1) объясняются их распространением в более суровых климатических условиях (ели и лиственницы – в условиях низких температур, сосны и лиственницы – недостатка влаги), где их относительная площадь сильно варьирует.
Таблица 1.
Оценка вклада изменчивости климатических условий в дисперсию относительной площади распространения хвойных видов в лесах Средней Сибири
| Вид | Климатические показатели | MSf | MSе | F |
|---|---|---|---|---|
| Abies sibirica | Т | 3143.0 | 33.61 | 93.52 |
| Pinus sibirica | 13 850.7 | 94.98 | 145.84 | |
| Picea obovata | 4039.5 | 91.62 | 44.09 | |
| Pinus sylvestris | 36 967.2 | 289.25 | 175.17 | |
| Larix spp. | 204 303.8 | 674.82 | 302.8 | |
| Abies sibirica | $\frac{{{\text{К Н А Р 1}}\,{\text{Т }}}}{{{\text{К Н А Р 2}}\,{\text{Т }}}}$ | $\frac{{342.5}}{{532.2}}$ | $\frac{{40.58}}{{50.49}}$ | $\frac{{8.44}}{{10.54}}$ |
| Pinus sibirica | $\frac{{905.9}}{{2709.8}}$ | $\frac{{125.34}}{{122.91}}$ | $\frac{{7.23}}{{22.05}}$ | |
| Picea obovata | $\frac{{4538.3}}{{3748.2}}$ | $\frac{{97.69}}{{97.02}}$ | $\frac{{46.45}}{{38.64}}$ | |
| Pinus sylvestris | $\frac{{3140.6}}{{4118.1}}$ | $\frac{{289.25}}{{290.04}}$ | $\frac{{10.74}}{{14.18}}$ | |
| Larix spp | $\frac{{43\,977.4}}{{37\,071.3}}$ | $\frac{{1107.87}}{{1100.71}}$ | $\frac{{39.70}}{{33.68}}$ | |
| Abies sibirica | ГТК | 437.3 | 40.00 | 10.92 |
| Pinus sibirica | 1263.6 | 124.26 | 10.17 | |
| Picea obovata | 1408.0 | 98.44 | 14.30 | |
| Pinus sylvestris | 48 348.9 | 216.41 | 223.42 | |
| Larix spp. | 50 934.2 | 1054.98 | 48.28 | |
| Abies sibirica | $\frac{{{\text{К Н А Р 1}} \cdot {\text{Г Т К }}}}{{{\text{К Н А Р 2}} \cdot {\text{Г Т К }}}}$ | $\frac{{1415.0}}{{1100.6}}$ | $\frac{{38.95}}{{39.30}}$ | $\frac{{36.33}}{{28.01}}$ |
| Pinus sibirica | $\frac{{3834.2}}{{4198.9}}$ | $\frac{{121.60}}{{120.89}}$ | $\frac{{31.53}}{{34.70}}$ | |
| Picea obovata | $\frac{{3060.0}}{{2843.1}}$ | $\frac{{96.96}}{{97.05}}$ | $\frac{{31.56}}{{29.29}}$ | |
| Pinus sylvestris | $\frac{{68\,072.5}}{{50\,382.3}}$ | $\frac{{186.10}}{{229.45}}$ | $\frac{{365.79}}{{219.58}}$ | |
| Larix spp. | $\frac{{160\,851.0}}{{96\,940.1}}$ | $\frac{{872.02}}{{1011.25}}$ | $\frac{{184.67}}{{95.86}}$ | |
| Abies sibirica | ТГКн | 1950.1 | 38.79 | 50.27 |
| Pinus sibirica | 10 021.3 | 116.49 | 86.03 | |
| Picea obovata | 13 320.5 | 87.73 | 151.84 | |
| Pinus sylvestris | 43 422.8 | 252.33 | 172.09 | |
| Larix ssp. | 295 705.5 | 849.69 | 348.01 | |
| Abies sibirica | $\frac{{{\text{К Н А Р 1}} \cdot {\text{Т Г }}{{{\text{К }}}_{{\text{н }}}}}}{{{\text{К Н А Р 2}} \cdot {\text{Т Г }}{{{\text{К }}}_{{\text{н }}}}}}$ | $\frac{{2824.6}}{{1853.7}}$ | $\frac{{34.14}}{{37.71}}$ | $\frac{{82.73}}{{49.16}}$ |
| Pinus sibirica | $\frac{{4821.0}}{{5308.1}}$ | $\frac{{115.13}}{{117.81}}$ | $\frac{{41.88}}{{45.10}}$ | |
| Picea obovata | $\frac{{8344.2}}{{9004.3}}$ | $\frac{{82.28}}{{86.14}}$ | $\frac{{101.41}}{{104.53}}$ | |
| Pinus sylvestris | $\frac{{24\,930.8}}{{22\,744.3}}$ | $\frac{{240.60}}{{257.85}}$ | $\frac{{103.62}}{{88.21}}$ | |
| Larix spp. | $\frac{{157\,314.4}}{{155\,236.3}}$ | $\frac{{838.47}}{{886.15}}$ | $\frac{{187.60}}{{175.18}}$ |
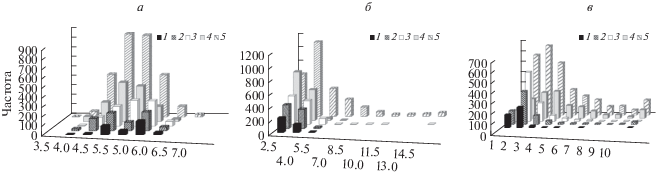
Сравнивали также распределения популяций видов в разных сочетаниях климатических характеристик на плоскости двух–трех координат. Приведем некоторые из них (рис. 4). В парах координат, начиная от исходных признаков (Т и Р) до расчетных гидротермических коэффициентов и особенно при сочетании последних, наблюдается существенное сокращение рассеивания точек популяций – сужение видовой КНАР в 2–3-мерном признаковом пространстве, в результате чего изменилась последовательность видов по КНАР (в порядке убывания: Larix spp. → Pinus sibirica → Pinus sylvestris → Picea obovata → Abies sibirica). Особенно заметное ограничение в распространении двух темнохвойных видов (Abies sibirica и Picea obovata) выявлено по ТГКн (высоте снежного покрова и степени промерзания почвы с наступлением холодов), виды сосен по сравнению с ними отличались большим разнообразием популяций по обобщенным климатическим характеристикам.
Рис. 4.
Распределение популяций пихты сибирской (1), ели сибирской (2), сосны сибирской кедровой (3), видов лиственницы (4) по парным КНАР1: Т–Р – 1; ГТК–ТГК н – 2.

Более высокая чувствительность популяций пихты сибирской к изменениям климата подтверждается наибольшим среди других темнохвойных видов распространением очагов усыхания пихтовых лесов, регистрируемых повсеместно в последние десятилетия в связи с ухудшением состояния деревьев и распространением инвазионных видов насекомых-фитофагов, произошедшем в результате климатических изменений (Forest decline…, 1993; Манько, Гладкова, 1995; Бажина, 2010; Allen et al., 2010; Van Mantgem et al., 2009; Павлов, 2015). Существенное сокращение в России площади темнохвойных лесов с 1988 по 2009 год (кедровых сосен – на 3.5%, ели – на 7.2% и пихты – на 9.9%) выявлено также по данным учета лесного фонда (Замолодчиков, 2011), что согласуется с приведенной выше последовательностью видов по комплексным характеристикам КНАР. О значимости внутрисезонной изменчивости температур (КНАР2) для елово-пихтовых древостоев свидетельствует также трехкратное снижение фитомассы деревьев (т/га) при возрастании индекса континентальности климата с 50 до 80 (Усольцев и др., 2016).
Для того чтобы выяснить, насколько исследуемые виды разделяются по всем учтенным нами климатическим характеристикам, был проведен дискриминантный анализ. Установлено, что 4 достоверных канонических корня объясняют 55.6% изменчивости площади популяций в зависимости от СМ значений климатических показателей в местообитаниях видов (F = 190.64, р < 0.0001), 36.2% изменчивости в связи с КНАР1 (3 достоверных канонических корня F = 125.12, р < 0.0001), 33.5% изменчивости в связи с КНАР2 (4 достоверных канонических корня F = 116.98, р < 0.0001), 44.0% изменчивости КНАР1 и КНАР2 популяций видов вместе объясняют 4 достоверных канонических корня (F = 75.95, р < 0.0001). Во всех случаях дистанции Махаланобиса между видами были достоверными (табл. 2).
Таблица 2.
Результаты дискриминантного анализа
| Признаки | Стандартизированные коэффициенты переменных при канонических корнях | F | |||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | ||
| Среднемноголетние значения | |||||
| T | 0.834 | –0.617 | 0.850 | 0.463 | 214.6 |
| Р | 0.235 | 1.169 | 0.426 | –0.577 | 88.7 |
| ГТК | 0.352 | –0.099 | –0.076 | –1.230 | 8.6 |
| ТГКн | 0.132 | –0.287 | –1.464 | 0.267 | 23.1 |
| A. sibirica | 1.319 | 0.261 | –0.252 | –0.187 | 10.8–310.0 |
| P. sibirica | 0.930 | 0.287 | 0.363 | –0.039 | 10.8–354.4 |
| P. obovata | 0.465 | 0.123 | –0.231 | –0.018 | 38.5–231.1 |
| P. sylvestris | 0.354 | –0.481 | 0.024 | 0.271 | 56.3–292.0 |
| Larix spp. | –0.683 | 0.072 | 0.018 | 0.104 | 231.1–354.4 |
| χ2 | 2740.5 | 446.5 | 72.9 | 4.88 | |
| р | 0.000000 | 0.000000 | 0.000000 | 0.035 | |
| КНАР1 | |||||
| σ T | –0.076 | 0.478 | 0.892 | 38.2 | |
| σ Р | 0.537 | 0.861 | –0.123 | 139.6 | |
| σ ГТК | –0.533 | 0.511 | –0.184 | 111.0 | |
| σ ТГКн | –0.474 | 0.166 | –0.189 | 69.4 | |
| A. sibirica | 1.107 | 0.565 | 0.059 | 40.6–225.2 | |
| P. sibirica | 0.615 | 0.011 | –0.356 | 40.2–190.8 | |
| P. obovata | 0.390 | 0.029 | 0.259 | 27.1–156.1 | |
| P. sylvestris | 0.283 | –0.353 | 0.026 | 27.1–180.0 | |
| Larix spp. | –0.537 | 0.074 | –0.017 | 156.1–225.2 | |
| χ2 | 1898.0 | 414.7 | 143.8 | ||
| р | 0.000000 | 0.000000 | 0.000000 | ||
| КНАР2 | |||||
| lim T | 0.265 | 0.687 | 0.507 | –0.592 | 55.23 |
| lim Р | 0.488 | 0.601 | –0.630 | 0.151 | 113.59 |
| lim ГТК | –0.670 | 0.438 | 0.028 | 0.6958 | 152.48 |
| limТГКн | –0.373 | 0.164 | –0.409 | –0.912 | 42.93 |
| A. sibirica | 1.067 | 0.546 | –0.089 | 0.066 | 32.0–210.7 |
| P. sibirica | 0.516 | 0.052 | –0.164 | –0.059 | 14.2–146.2 |
| P. obovata | 0.443 | 0.065 | 0.216 | –0.019 | 14.2–163.9 |
| P. sylvestris | 0.291 | –0.394 | –0.024 | 0.022 | 29.3–185.1 |
| Larix spp. | –0.521 | 0.072 | –0.008 | 0.003 | 146.2–210.7 |
| χ2 | 1780.97 | 369.01 | 66.34 | 4.65 | |
| р | 0.000000 | 0.000000 | 0.000000 | 0.031 | |
Согласно полученным в трех вариантах анализа матрицам дистанций Махаланобиса (D) степень различий между видами возрастает от КНАР2 к КНАР1, и наибольшая дифференциация наблюдается при сравнении видов на основе СМ значений метеорологических показателей (рис. 5). Виды Larix во всех случаях выделяются в отдельный кластер. Сильно варьирует положение Abies sibirica – по КНАР1 этот вид выделяется в отдельный кластер, но обнаруживает большее сходство с Picea obovata по КНАР2, либо с Pinus sibirica (по СМ характеристикам). Picea obovata от КНАР2 (минимальные расстояния D с Abies sibirica) к двум другим вариантам анализа обнаруживает большее сходство с видами р. Pinus. Необходимо также отметить наибольшее сходство по КНАР между Pinus sylvestris и P. sibirica, сильно различающимися по экологическим характеристикам местообитаний. Причем виды сосен обнаруживают максимальное сходство по способности адаптироваться к резким изменениям климата (экстремальным событиям). Очевидно, что это свидетельствует о сходстве их чувствительности, так как чувствительность является одной из характеристик живых организмов, тесно связанной с их нормой реакции, и одним из наиболее высоко наследуемых признаков (Тихонова и др., 2012).
Рис. 5.
Дендрограмма сходства видов для трех вариантов сочетания климатических показателей: а – по среднемноголетним значениям Т, Р, ГТК, ТГКн; б – по КНАР1; в – по КНАР2, D – дистанции Махаланобиса.
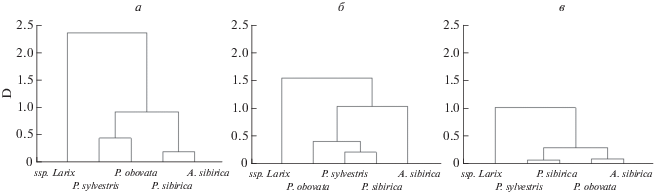
Необходимо отметить, что полученные классификации в общем согласуются с выводами ботаников, палеоботаников и молекулярных генетиков о происхождении и родстве хвойных видов в сем. Pinaceae (Чавчавадзе, Яценко-Хмелевский, 1978; Бобров, 1978; Willyard et al., 2007; Farjon, 2010; Leslie et al., 2012, и др.). Поэтому предлагаемый нами подход к исследованию КНАР видов после доработки и уточнения, по-видимому, можно будет в зависимости от решаемых задач использовать для реконструкции филогенетических связей между таксонами и наоборот – условий их эволюции в дополнение к уже используемым методам. Подчеркнем, что если бы мы оперировали только СМ характеристиками местообитаний, то не заметили бы такой возможности.
Поскольку все сравниваемые виды различаются по экологическим функциям и появляются на разных этапах сукцессии, то определенный интерес представляет тот факт, что свойство листопадности сформировалось у одного из пионерных видов (родов), участвующих в первых этапах формирования лесных сообществ. Более высокая чувствительность к климатическим изменениям популяций видов, относящихся к последним стадиям сукцессии (так называемых климаксовых сообществ), является, по-видимому, вынужденной платой сем. Pinacea (все исследуемые виды относятся к одному семейству) за специализацию, сочетающую высокую теневыносливость, конкурентоспособность и рост в более благоприятных микроклиматических и эдафических условиях чем те, что были у светолюбивых видов первых этапов сукцессий (сосны обыкновенной и лиственницы сибирской). Добавим к этому, что по мнению В. Гранта (1991), узкоспециализированные виды в условиях сильной межвидовой конкуренции больше подвержены опасности вымирания при изменении среды. Вероятно именно поэтому у представителей темнохвойных видов отмечена наибольшая негативная реакция на современные климатические изменения.
При этом повышение устойчивости бореальных лесных сообществ на каждом этапе сукцессии к изменениям климата может обеспечиваться тем, что в каждой из сукцессионных групп произрастающих в Сибири лесообразующих видов хвойных имеется хотя бы один вид с очень широкой КНАР. Так, среди пионерных светлохвойных видов виды лиственницы адаптированы к широкому спектру суровых условий (низкие температуры, избыточное увлажнение, засухи), а сосна обыкновенная больше приспособлена к выживанию в условиях засушливого климата. Из темнохвойных видов большей КНАР отличаются сосна сибирская (по устойчивости к низким температурам почвы и значительным колебаниям показателя ТГКн в местообитаниях с невысоким снежным покровом) и ель сибирская (по устойчивости к колебанию условий увлажнения КНАР1 и КНАР2 ГТК), но ее меньшая устойчивость к низкой температуре почвы компенсируется расселением на более влажных почвах по долинам рек. Отметим, что в многомерном пространстве показателей КНАР облака видов расширяются в следующем порядке: Abies sibirica – Picea obovata – Pinus sylvestris – Pinus sibirica – Larix sibirica, т.е. темнохвойные и светлохвойные виды частично перекрываются, кроме того, темнохвойные виды не образуют отдельного кластера ни на одной из приведенных на рис. 5 дендрограмм.
Сопоставляя полученные оценки сходства и различия между хвойными видами по КНАР1, КНАР2 и СМ значениям можно также предположить, что различия по СМ характеристикам мест произрастания популяций видов, классификация по которым больше соответствует сходству/различиям между видами по их географическим ареалам и приуроченности к возрастным этапам развития лесных сообществ, в меньшей степени генетически детерминированы по сравнению с КНАР, точнее, могут быть связаны с менее глубокими генетическими перестройками в процессе эволюции хвойных видов. Это предположение может подтверждаться многими примерами быстрого изменения видов и “наиболее сильных эволюционных преобразований” при появлении свободных экологических ниш (Шварц, 1980; Медников, 1986). Более того, по утверждению С.С. Шварца (1980), каждый таксон – это своя экологическая ниша. Ответить на этот вопрос, однако, можно только с помощью междисциплинарной систематизации знаний, сопоставив степени морфологических и генетических (по комплексу разных маркеров) различий между таксонами и определив “генетический вес” разных таксонов и морфологических признаков, определив таким образом внутренние или внешние факторы являются наиболее значимыми в процессе эволюции (пока априори отдается предпочтение внешним).
В завершение оценим степень дифференциации популяций внутри видов. Но прежде дадим краткую характеристику природно-климатических зон (ПКЗ), в которых они произрастают. Горные районы с субарктическим, суббореальным и субаридным типами поясности наряду с высокой изменчивостью метеорологических условий произрастания хвойных лесов в пространстве отличаются меньшей изменчивостью климатических показателей по годам (для всех меньшей по Т, а в горах Южной Сибири с суббореальным типом поясности – также и по ГТК и ТГКн). В других ПКЗ метеорологические величины распределяются в пространстве более равномерно, но более изменчивы во времени. Наибольшей межсезонной изменчивостью ТГКн отличались местообитания хвойных видов в горах с субаридным климатом.
Установлено, что в среднетаежной подзоне нередко наблюдаются температуры, характерные для северной границы сплошного произрастания лесов. В частности, в окрестностях метеостанций “Байкит” и “Ванавара” температуры до –7.1…–9.5°С составляют более 30% учтенных лет. В некоторых местообитаниях 3–5 совместно произрастающих хвойных видов в горах с суббореальным и бореальным типами поясности на высотах от 1000 м над ур. м. до верхней границы леса среднемноголетние температуры составляют –5.5…–6.9°С, с минимумами –7.6…–9.1°С. Большим разнообразием характеризуются условия произрастания хвойных видов в горах Южной Сибири – Западного и Восточного Саян, Прибайкалья, Забайкалья и внутренних гор Тувы: от холодных засушливых в горных районах с субаридным климатом до прохладных гумидных в районах с суббореальным климатом. Значительной сухостью и холодностью климата отличаются внутренние горы Тувы, где, по данным метеостанции “Тоора-Хем”, среднегодовая температура воздуха колеблется от –6.8 до –2.4°С, годовая сумма осадков 323 мм (217–486 мм), ТГКн варьирует от –6.4 до –0.5 и условия роста хвойных лесов близки к условиям среднетаежной зоны на широте 59–62° с. ш., но значительно суше. В районах с недостаточным увлажнением, в местах, как правило, островного произрастания хвойных лесов однолетние и 2–3-летние засушливые сезоны с суммой осадков 171–289 мм составляют от 10 до 54% от числа учтенных лет, а ТГКн в отдельные годы в некоторых пунктах опускается до –7…–12°С, т.е. до уровня северных популяций.
Для оценки степени внутривидовой дифференциации хвойных видов по тем же показателям снова были использованы дискриминантный и кластерный анализы характеристик групп популяций внутри видов из разных природно-климатических зон. В результате анализа установлена существенная (F = 2.89–497.9, p < 0.038–0.0000) дифференциация групп популяций всех лесообразующих хвойных видов по КНАР1 и КНАР2, за исключением 1–3 сочетаний популяций у Pinus sylvestris и P. sibirica. При этом были выявлены следующие общие географические закономерности дифференциации хвойных видов: большее сходство между популяциями, произрастающими в смежных подзоне южной тайги и в лесостепной зоне по КНАР1, КНАР2 и СМ значениям метеорологических показателей; то же в подзонах средней, северной тайги и горной бореальной зоне; то же между географически отдаленными горными образованиями с субаридным и бореальным типом поясности. Несмотря на сходство по СМ-характеристикам популяций из подзоны средней тайги и горных районов бореальной зоны с популяциями, произрастающими в горах с суббореальным типом поясности (на юге Сибири), они в целом существенно различаются по КНАР1 и КНАР2, исключение составляют местообитания популяций Pinus sibirica и Abies sibirica. Этими различиями, вероятно, объясняется гибель большинства происхождений сосны обыкновенной при испытании в предгорьях Западного Саяна (Ирошников, 1977) с суббореальным типом поясности, отличающимся по КНАР от всех других ПКЗ. Популяции Pinus sibirica, произрастающие в горах с субаридным климатом, обнаруживают большее сходство с популяциями суббореального климата, а популяции Pinus sylvestris, растущие в условиях субаридного климата – с популяциями таежной зоны, Larix sibirica перекрывает оба сочетания.
Таким образом, популяции всех хвойных видов из смежных природно-климатических зон, а также популяции, находящиеся на значительном расстоянии друг от друга, например в горах с бореальным и субаридным типами поясности, и некоторые популяции отдельных видов из других ПКЗ можно рассматривать в качестве близких (по комплексным КНАР) потенциально возможных источников семян для целей лесовосстановления в случае катастрофической гибели лесов на большой площади. Для уточнения этого вопроса конечно потребуется увеличение набора внутрисезонных климатических показателей.
ЗАКЛЮЧЕНИЕ
В результате проведенного исследования на основе анализа гидрометеорологических данных, использованных нами для изучения климатической нормы адаптивных реакций популяций, были установлены регулярные и нерегулярные (экстремальные) КНАР основных лесообразующих видов хвойных в Средней Сибири за период 1960–2013 гг. Выявлены достоверные различия между всеми видами по их отношению к межсезонной изменчивости климата, наибольшие – по среднемноголетним значениям и КНАР1. Отмечено значительное сужение обобщенных КНАР (и высокий эксцесс в распределении) у видов Abies sibirica и Picea obovata, что согласуется с приведенными в литературе данными по усыханию пихтовых и еловых лесов в России, объясняемому влиянием изменения климата и загрязнения воздуха.
Последовательный анализ сходства и различий между видами по показателям изменчивости климата на разных уровнях их обобщения позволил обнаружить сходство полученных классификаций с филогенетическими построениями для видов сем. Pinaceae, составленным на основе палеоботанических, анатомо-морфологических и генетических исследований. Благодаря этому был сделан вывод о возможности использования предлагаемого нами подхода в качестве дополнительного в филогенетических реконструкциях разных видов многолетних древесных растений, а также выдвинут ряд предположений о последовательности расхождения родов в сем. Pinaceae и о возможном повышении устойчивости бореальных лесов во времени при изменениях климата и в пространстве, обеспеченном перекрыванием светлохвойных и темнохвойных видов по КНАР. В частности, об этом свидетельствует последовательность расширения КНАР видов по комплексным гидротермическим характеристикам.
Необходимо отметить, что популяции внутри видов больше всего различаются по сопряженным показателям КНАР (в 1.7–4.0 раза по Т и Р, в 3–23 раза по ГТК и в 4–35 раз по ТГКн). Кроме того, установлена достоверная внутривидовая дифференциация всех пяти хвойных видов по КНАР между группами популяций из разных природно-климатических зон. Это свидетельствует о существовании видоспецифичной пространственной внутривидовой структуры (по комплексным характеристикам условий: СМ, КНАР1, КНАР2, а также другим, сопряженным с ними биоценотическим факторам). Поэтому рациональное ведение лесного хозяйства в хвойных лесах невозможно без сохранения внутривидового разнообразия основных лесообразующих хвойных видов, т.е. их сохранения не только на немногочисленных и относительно небольших по площади особо охраняемых природных территориях (ООПТ), но и на всем протяжении ареалов видов. Это будет способствовать более быстрому восстановлению лесов при любых негативных климатических изменениях. Особую ценность в этой связи представляют популяции с широкой КНАР по нескольким метеорологическим показателям, произрастающие в средней тайге и в горах с бореальным и субаридным типами поясности, в некоторых горных местообитаниях с суббореальным типом поясности, а также на границах ареалов видов. Некоторые из них, по-видимому, могут обладать потенциальной способностью в первом поколении адаптироваться к новым условиям произрастания на значительном расстоянии по широте (до 600–1000 км) и долготе (до 1000–1300 км).
Работа выполнена в рамках бюджетного проекта ФГБНУ ЗСО ИЛ СО РАН, ФИЦ КНЦ СО РАН (проект № 0346-2016-0301), при финансовой поддержке РФФИ и КГАУ ККФПНиНТД грант № 18-44-240002.
Список литературы
Авров Ф.Д., 1998. Эколого-генетические основы устойчивости популяций и плантационного выращивания лиственницы в Сибири. Автореф. дис. докт. с.-х. наук. Красноярск: СибГТУ. 36 с.
Алтухов Ю.П., 2004. Динамика генофондов при антропогенных воздействиях // Вестн. ВОГиС. Т. 8. № 2. С. 40–59.
Бажина Е.В., 2010. О факторах усыхания пихтовых лесов в горах Южной Сибири // Изв. Иркутского гос. ун-та. Сер. биол. Экол. № 3. С. 20–25.
База данных Всероссийского научно-исследовательского института гидрометеорологической информации (ВНИИГМИ-МЦД) http://meteo.ru.
Барталев С.А., Егоров В.А., Ершов Д.В. и др., 2011. Спутниковое картографирование растительного покрова России по данным спектрорадиометра MODIS // Современные проблемы дистанционного зондирования Земли из космоса. Т. 8. № 4. С. 285–302.
Бобров Е.Г., 1978. Лесообразующие хвойные СССР. Л.: Наука ЛО. 189 с.
Боровиков В.П., 2003. Statistica. Искусство анализа данных на компьютере. 2-е изд. СПб.: Питер. 688 с.
Будыко М.И., 1975. Экологические факторы эволюции // Журн. общ. биологии. Т. 3. № 1. С. 36–47.
Бузыкин А.И., Пшеничникова Л.С., 2008. Ресурсно-экологический потенциал лесов Красноярского края // Хвойные бореал. зоны. Т. 25. № 3–4. С. 327–332.
Вернадский В.И., 1940. Биогеохимические очерки. М.; Л.: Изд-во АН СССР. 250 с.
Глотов Н.В., Тараканов В.В., 1985. Норма реакции генотипа и взаимодействие генотип–среда в природной популяции // Журн. общ. биологии. Т. 46. № 6. С. 760–770.
Грант В., 1991. Эволюционный процесс. М.: Мир. 488 с.
Замолодчиков Д.Г., 2011. Оценка климатогенных изменений разнообразия древесных пород по данным учетов лесного фонда // Успехи соврем. биологии. Т. 131. № 4. С. 382–392.
Жильцова, Е.Л., Анисимов О.А., 2009. О точности воспроизведения температуры и осадков на территории России глобальными климатическими архивами // Метеорология и гидрология. № 10. С. 79–89.
Иогансен В., 1933. Элементы точного учения об изменчивости и наследственности. М.; Л.: Госиздат колхозн. и совхозн. лит. 409 с.
Ирошников А.И., 1977. Географические культуры хвойных в Южной Сибири // Географические культуры и плантации хвойных в Сибири. Новосибирск: Наука СО. С. 4–110.
Калашников Е.Н., 1998. Мониторинг нарушенности лесов Сибири // Сиб. эколог. журн. № 1. С. 49–57.
Каппер О.Г., 1946. Изучение экотипов древесных пород // Науч. зап. Воронежского ЛХИ. Т. IX. С. 56–64.
Карта “Леса СССР” М 1 : 2500000, 1990. М.: Федеральная служба лесного хозяйства. 16 л.
Козин Е.К., 2013. Массовые распады темнохвойных лесов как естественный этап возрастного развития // Бюл. бот. сада–института ДВО РАН. Т. 10. С. 4–14.
Колесников Б.П., 1961. Генетическая классификация типов леса и ее задачи на Урале // Вопросы классификации растительности. Тр. ин-та биологии. Вып. 27. С. 47–59.
Кулагин Ю.З., 1980. Лесообразующие виды, техногенез и прогнозирование. М.: Наука. 113 с.
Леса СССР. Леса Урала, Сибири и Дальнего Востока. Т. 4, 1969 / Под ред. Жукова А.Б. М.: Наука. 766 с.
Майр Э., 1974. Популяции, виды и эволюция: Пер. с англ. М.: Мир. 464 с.
Манько Ю.И., Гладкова Г.А., 1995. О факторах усыхания пихтово-еловых лесов на Дальнем Востоке // Лесоведение. № 2. С. 3–12.
Медников Б.М., 1986. Аксиомы биологии. М.: Знание. 154 с.
Мельник П.Г., Смирнов В.Н., Камышова Л.В., 2009. 90-летний опыт географических культур сосны обыкновенной в Бузулукском бору // Лесн. вестник. № 1. С. 88–94.
Молотков П.И., Патлай И.Н., Давыдова Н.И. и др., 1982. Селекция лесных пород. М.: Лесн. пром-сть. 224 с.
Мониторинг биологического разнообразия лесов России: методология и методы, 2008 / Под ред. Исаева А.С. М.: Наука. 453 с.
Молчанов А.А., 1978. Влияние леса на окружающую среду. М.: Наука. 358 с.
Павлов И.Н., 2015. Биотические и абиотические факторы усыхания хвойных лесов Сибири и Дальнего Востока // Сиб. экол. журн. № 4. С. 537–554.
Петрашов В.В., 2006. Глаза и мозг эволюции. Новая теория эволюции организмов. 3-е изд. М.: Прометей. 464 с.
Протопопов В.В., 1975. Средообразующая роль темнохвойного леса. Новосибирск: Наука. 327 с.
Разумовский С.М., 1981. Закономерности динамики биоценозов. М.: Наука. 229 с.
Рахманов В.В., 1984. Гидроклиматическая роль лесов. М.: Лесн. пром-сть. 240 с.
Семериков В.Л., Ирошников А.И., Ласкоук М., 2007. Структура изменчивости митохондриальной ДНК и послеледниковая история лиственницы сибирской (Larix sibirica Ledeb.) // Экология. № 3. С. 1–9.
Сочава В.Б., 1956. Темнохвойные леса // Растительный покров СССР. Т. 1. М.; Л.: Изд-во АН СССР. С. 139–216.
Тимофеев-Ресовский Н.В., Яблоков А.В., Глотов Н.В., 1973. Очерк учения о популяциях. М.: Наука. 217 с.
Тихонова И.В., Тараканов В.В., Кнорре А.А., 2012. Вклад генотипических и метеорологических факторов в изменчивость годичных приростов древесины на клоновых плантациях сосны // Экология. № 3. С. 163–169.
Усольцев В.А., Маленко А.А., Азарёнок В.А., 2016. Биологическая продуктивность елово-пихтовых насаждений в климатических градиентах Евразии // Вестн. Алтайского гос. аграрн. ун-та. № 6. Вып. 140. С. 65–71.
Хромов С.П., Мамонтов Л.И., 1974. Метеорологический словарь. М.: Гидрометеоиздат. 569 с.
Чавчавадзе Е.С., Яценко-Хмелевский А.А., 1978. Семейство сосновые (Pinaceae) // Жизнь растений. М.: Просвещение. Т. 4. С. 350–374.
Шварц С.С., 1980. Экологические закономерности эволюции. М.: Наука. 280 с.
Шишкин М.А., 1988. Эволюция как эпигенетический процесс // Современная палеонтология. М.: Недра. С. 142–169.
Шмальгаузен И.И., 1968. Кибернетические вопросы биологии. Новосибирск: Наука. 213 с.
Шутяев А.М., 2007. Изменчивость хвойных видов в испытательных культурах Центрального Черноземья. М.: ВНИИЛМ. 296 с.
Allen C., Macalady A.K., Chenchouni H. et al., 2010. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests // For. Ecol. Manage. V. 259. P. 660–684.
Davin E., De Noublet-Ducoudre N., 2010. Climatic Impact of Global-Scale Deforestation: Radiative versus Nonradiative Processes // J. Climate. V. 23. P. 97–112.
Eiche V., Andersson E., 1974. Survival and growth in Scots pine (Pinus silvestris L.): Provenance experiments in Northern Sweden // Theor. Appl. Genet. V. 44. № 2. P. 49–57.
Farjon A.A., 2010. Handbook of the World’s Conifers. Koninklijke Brill, Leiden; Boston: Brill Acad. Publishers. 1150 p.
Forest Decline in the Atlantic and Pacific Region, 1993 /Eds Huettl R.F., Mueller-Dombois D. Berlin; Heidelberg: Springer-Verlag. 366 p.
Gernandt D.S., Magallo’n S., Lo’pez G.G. et al., 2008. Use of simultaneous analyses to guide fossil-based calibrations of Pinaceae phylogeny // J. Plant Sci. V. 169. № 8. P. 1086–1099.
Giertych M., 1979. Summary of results on Scots pine (Pinus sylvestris L.) height growth in IUFRO provenance experiments // Silvae Genetica. V. 28. № 4. P. 136–152.
Hargreaves A.L., Samis K.E., Eckert C.G., 2014. Are species’ range limits simply niche limits writ large? A review of transplant experiments beyond the range // Am. Nat. V. 183. № 2. P. 157–173.
Jansons Ā., Baumanis I., 2005. Growth dynamics of Scots pine geographical provenances in Latvia // Baltic Forestry. V. 11. № 2. P. 29–37.
Kirkpatrick M., Barton N.H., 1997. Evolution of a species’ range // Am. Nat. V. 150. № 1. P. 1–23.
Leslie A.B., Beaulieu J.M., Rai H.S., Crane P.R., Donoghue M.J., Mathews S., 2012. Hemisphere-scale differences in conifer evolutionary dynamics // Proc. Natl Acad. Sci. V. 109. № 40. 16217 doi 10.1073/pnas.1213621109
Lewontin R.C., 1974. The analysis of variance and the analysis of causes // Amer. J. Hum. Genet. V. 26. № 3. P. 400–411.
Mitchell T.D., Jones P.D., 2005. An improved method of constructing a database of monthly climate observations and associated high-resolution grids // Int. J. Climatology. № 25. P. 693–712.
Polezhaeva M.A., Semerikov V.L., Lascoux M., 2010. Cytoplasmic DNA variation and biogeography of Larix Mill, in Northeast Asia // Molecular Ecology. V. 19. № 6. P. 1239–1252.
Schlichting C.D., Pigliucci M., 1998. Phenotypic evolution: A reaction norm perspectives. Sunderland, Massachusetts: Sinauer Associates, Inc., Publishers. 340 p.
Semerikov V.L., Semerikov L.F., Lascoux M., 1999. Intra- and interspecific allozyme variability in Eurasian Larix Mill. species // Heredity. V. 82. № 2. P. 193–204.
Sexton J.P., McIntyre P.J., Angert A.L., Rice K.J., 2009. Evolution and Ecology of Species Range Limits // Ann. Rev. Ecol., Evol. Systematics. V. 40. P. 415–436.
Van Mantgem P.J., Stephenson N.L., Byrne J.C. et al., 2009. Widespread increase of tree mortality rates in the western United States // Science. V. 323. P. 521–524.
Willyard A., Syring J., Gernandt D.S. et al., 2007. Fossil calibration of molecular divergence infers a moderate mutation rate and recent radiations for Pinus // Mol. Biol. Evolution. V. 24. P. 90–101.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии