

Журнал общей биологии, 2019, T. 80, № 1, стр. 43-56
Гибридизация как фактор инвазионной активности чужеродных видов золотарника (Solidago)
Ю. К. Виноградова 1, *, М. А. Галкина 1
1 Главный ботанический сад им. Н.В. Цицина РАН,
127276 Москва, ул. Ботаническая, 4, Россия
* E-mail: gbsad@mail.ru
Поступила в редакцию 30.05.2017
После доработки 01.03.2018
Принята к публикации 27.04.2018
Аннотация
Согласно одной из гипотез успешность инвазионных видов обусловлена усилением гибридизационных процессов во вторичном ареале. Цель исследования – прогнозирование скорости дальнейшего внедрения в естественные фитоценозы сформировавшихся во вторичном ареале гибридогенных таксонов золотарника (Solidago L.) с проверкой гибридогенного характера этих таксонов экспериментальными и молекулярно-генетическими методами, а также тестирование гипотезы о более высокой конкурентоспособности и инвазионной активности гибридов по сравнению с родительскими видами. Для сравнения приводится анализ гибридогенной активности некоторых других представителей сем. Asteraceae североамериканского происхождения (Bidens spp., Erigeron sect. Conyza spp.) на основании данных, полученных авторами ранее. Показано, что не все таксоны с промежуточными морфологическими признаками являются гибридами. Так, ни экспериментальным, ни молекулярно-генетическим методами не подтверждено гибридное происхождение “Bidens × garumnae” и “Solidago сanadensis × S. gigantea”. Возникшие в Европе гибриды Solidago × niederederi, S. × snarskisii и Conyza × rouyana встречаются пока во вторичном ареале родительских видов единично и менее конкурентоспособны, чем их предковые формы. Тенденция к натурализации наблюдается только у S. × niederederi. Таким образом, гипотеза о более высокой конкурентоспособности и инвазионной активности гибридов по сравнению с родительскими видами не всеобъемлюща, и подтверждающие ее примеры являются скорее исключением, нежели правилом.
Инвазии агрессивных чужеродных видов – важнейшая проблема, стоящая перед всеми странами мира. Внедрение инвазионных видов приводит к обеднению аборигенной флоры значительных территорий и часто наносит существенный экономический ущерб. Затраты, связанные с контролем инвазионных видов, превышают в Европе 12 млрд евро в год (McGeoch et al., 2010). В настоящее время большое внимание уделяется воздействию инвазионных видов на окружающие фитоценозы, расширению их вторичных ареалов и, как следствие, неизбежному взаимодействию и гибридизации с представителями аборигенных флор. Изучение процессов микроэволюции и механизмов, контролирующих устойчивость инвазионных видов при внедрении в естественные сообщества, особенно актуально в связи с возрастанием антропогенного воздействия и прогрессирующим увеличением доли нарушенных и квазиприродных местообитаний (Tokhtar et al., 2011).
Одной из гипотез, объясняющих успешность произрастания растений на новой родине, служит предположение об усилении гибридизационных процессов во вторичном ареале (Elton, 1958; Ellstrand, Schierenbeck, 2000). В непривычных условиях чужеродные виды могут формировать гибриды как с близкородственными аборигенными видами, так и с другими чужеродными растениями, расселяющимися на данной территории. Часто гибриды лучше приспособлены к условиям вторичного ареала, чем родительские таксоны (Abbott et al., 2003; Bleeker et al., 2007; Zalapa et al., 2009), в результате чего они преобразуют ландшафты новой родины. Удачная рекомбинация генетических признаков родительских видов сокращает lag-фазу (период адаптации чужеродного таксона к новым условиям, в ходе которого еще не происходит активное внедрение в естественные фитоценозы и расширение вторичного ареала) и приводит к образованию новых активных “видов-трансформеров”. В наибольшей степени к гибридизации предрасположены перекрестноопыляемые виды растений. Симпатрические виды менее склонны к межвидовой гибридизации, чем аллопатрические виды или популяции (Albuquerque et al.,1996).
Продуктами спонтанной гибридизации с вовлечением одного или обоих чужеродных родителей являются 2% чужеродных для Европы видов (41 вид). Доля гибридогенных таксонов среди инвазионных видов Средней России достигает 10% (Виноградова, Майоров, 2015). В результате гибридизации сформировались, например, активно расселяющиеся Reynoutria × bohemica, Symphytum × uplandicum, Aster × salignus, Amelanchier × spicata и др.
Долгое время важнейшим фактором ограничения гибридизации была географическая изоляция, однако в настоящее время близкородственные таксоны вступают в контакт друг с другом с помощью множества антропогенных “коридоров” (Williamson, 1996). Так, межрегиональная иммиграция происходит при интродукции растений, которая может быть частой и повторяющейся, а потому значительно увеличивающей иммиграционный поток (Heywood, 1989). Если рассмотреть ситуацию, когда условия для гибридизации близкородственных таксонов уже существуют, возможно несколько вариантов развития событий: а) чужеродный вид может исчезнуть; б) оба таксона могут сосуществовать; в) может появиться новый гибридогенный таксон; г) исчезает аборигенный таксон.
При гибридизации происходит генетическая ассимиляция, и новые гены вливаются в один или в оба родительских вида. Гибриды, даже будучи фертильными, могут быть, однако, репродуктивно изолированы от родительских растений из-за действия отбора по репродуктивным признакам (аллополиплопдия, гетерозиготные транслокации, рекомбинация, видоспецифичные различия митохондриальной ДНК) и/или факторов, предопределяющих скрещивание (фенология цветения, разделение экологических ниш). Межвидовая гибридизация может также облегчить натурализацию редких генотипов, а затем и привести к увеличению их доли путем обратного скрещивания с чужеродными родительскими таксонами или же гибридизации между самими гибридами. Большие преимущества в отборе для чужеродных аллелей должны привести к более быстрому замещению естественных аллелей при наличии гибридизации и более медленному – без гибридизации (Tokhtar et al., 2011).
Период вытеснения (замещения) значительно сокращается с увеличением иммиграционного потока и селективной дифференциации. Иммиграция и отбор действуют различными способами: увеличение уровня иммиграции приводит к замещению аборигенных видов путем их подавления, тогда как увеличение селективной дифференциации в пользу чужеродного вида ведет к замещению аборигенного путем генетической ассимиляции, не оставляя “чистых” аборигенных видов. При умеренной и высокой скорости иммиграции потеря аборигенных видов может протекать быстро как при гибридизации, так и при ее отсутствии. Учитывая высокую численность видов, интродуцируемых человеком, исчезновение аборигенных видов может возрастать в результате одной только гибридизации (Huxel, 1999).
Хорошо изучена гибридизация в р. Oenothera, представители которого имеют особый генетический механизм поддержания адаптивных генотипов. Существование чистых линий сравнительно редко нарушается гибридизацией, в результате которой возникают новые генотипы, которые могут стабилизироваться и дать начало новым микровидам (Грант, 1984). Видообразование в этом случае чрезвычайно активно, а lag-фаза коротка (Tokhtar et al., 2011). Подтверждением тому служат описанные К. Ростаньским 32 вида энотеры, четыре из которых являются инвазионными.
Центральноамериканские Bidens pilosa и B. alba стали инвазионными видами на Гавайских островах. Они вытесняют аборигенные виды череды и могут скрещиваться как с некоторыми из них (в частности, с B. menziesii), так и между собой, образуя фертильные гибриды B. pilosa × B. alba. Фертильность их пыльцы, однако, сравнительно невелика и составляет от 31 до 49% (Knope et al., 2013).
Высокую гибридогенную активность проявляют и виды р. Aster s.l. A.× salignus Willd. – гибрид североамериканских A. lanceolatus и A. novi-belgii – внедряется в естественные ценозы на территории Европы (Brouillet et al., 2006).
Потенциальным последствием гибридизации оказывается способность гибрида внедриться на территорию, на которой не произрастали родительские виды. При этом общий размер популяции может быть увеличен в связи с появлением новых ресурсных возможностей. Однако случаев относительной устойчивости, а тем более – сверхдоминирования гибридов зарегистрировано не так уж много. Классический пример относится к восточно-американскому гексаплоиду Spartina alterniflora (материнский вид), который при скрещивании с евразиатско-африканским гексаплоидом S. maritima (отцовский вид) в Англии и во Франции образовал стерильный S. × townsendii. Этот гибрид был отмечен в середине XIX в. в Европе только в двух точках и не проявлял способности к натурализации до появления аллотетраплоидной формы S. × anglica C.E. Hubb., возникшей в 1890-х. Эта форма морфологически отличается от S. × townsendii и способна расти в более широком диапазоне условий. Сейчас S. × anglica распространилась очень широко, захватив колоссальные пространства в приливной зоне на побережье Англии, Франции, Северной и Южной Америки, Австралии и Новой Зеландии (Guénégou, Levasseur, 1993; Anouche et al., 2016; Salmon et al., 2016). Одновременно с S. × anglica во Франции от тех же родительских видов независимо возник другой гибрид – S. × neyrautii. С помощью ISSR-метода выявлено, что эти гибриды имеют различные генотипы (Baumel et al., 2003) и существенно отличаются по биосинтезу DMSP (диметилсульфониопропионата) – вещества, которое обычно синтезируют водоросли и крайне редко – покрытосеменные. Тетраплоидные виды р. Spartina не имеют способности к синтезу DMSP. Листья S. alterniflora производят больше DMSP, чем листья S. maritima, а что касается их гибридов, то S. × townsendii по данному показателю близок к S. maritima, а S. × neyrautii занимает промежуточное положение между родительскими видами. Полиплоид S. × anglica по синтезу DMSP не имеет различий с S. × townsendii (Rousseau et al., 2016).
На основании этого примера была выдвинута гипотеза (Levin et al., 1996), что гибриды могут лучше всего существовать в условиях, не освоенных ни одним из родителей. Еще одним подобным примером служит гибридизация в Германии заносной Rorippa austriaca с аборигенной R. sylvestris с образованием комплекса гибридных форм с уровнями плоидности 3x–5x, размножающихся половым путем и распространяющихся в регионах, где родительские виды не встречаются (Bleeker, 2003).
Гибридизация увеличивает угрозу исчезновения многих видов в связи с интрогрессией (Levin и др., 1996; Rhymer, Simberloff, 1996). Высокую степень интрогрессии часто проявляют ветроопыляемые виды, например, дубы. Гибридизация и интрогрессия могут привести к образованию гибридного комплекса, состоящего из множества гибридов благодаря большому числу локусов. Таким образом, мультилокусы, по-видимому, увеличивают число гибридных типов и генетических комплексов и ускоряют сокращение “чистых” природных типов. Кроме того, большое число локусов по существу уменьшает вероятность наличия “чистого” индивидуума любого родительского происхождения (Huxel, 1999).
Без интрогрессии гибриды, будучи репродуктивно изолированными, могут быстро сформировать новый вид. При наличии интрогрессии процесс видообразования замедляется, поскольку происходят обратные скрещивания с родительскими линиями. Вопрос о влиянии гибридизации и интрогрессии на скорость замещения аборигенных видов близкородственными заносными решался с помощью математической модели, предполагающей однолокусную двуаллельную схему наследования с различным уровнем межвидовой гибридизации (Huxel, 1999). Хотя модель и не учитывала вегетативно размножающиеся гибриды, результаты показали, что замещение природных таксонов чужеродными может произойти очень быстро (меньше, чем за пять поколений). Согласно полученным результатам, гибридизация и интрогрессия могут усилить степень замещения аборигенных видов чужеродными. Интрогрессия увеличивает замещение видов при низкой степени иммиграции, но препятствует ему, когда у аборигенного вида есть значительное преимущество в отборе, а также при более высоком уровне иммиграции. Однако, поскольку интрогрессия связана с увеличением частоты гибридов, воздействие на аборигенный таксон продолжает оставаться высоким и вероятность его исчезновения значительно возрастает (Huxel, 1999).
Обсуждаемая математическая модель хорошо описывает поведение видов шалфеев (Salvia) на территории Хорватии. Здесь идет спонтанная гибридизация между аборигенным видом S. officinalis, встречающимся лишь в ограниченном числе местообитаний, и чужеродным S. fruticosa, активно приспосабливающимся к новым условиям среды. Отмечен бэккроссинг гибридов с родительским видом S. fruticosa, причем бэккроссы морфологически могут быть практически неотличимы от S. fruticosa (Radosavljević et al., 2016).
Интрогрессивная гибридизация ставит под угрозу и существование таксона Prunus fruticosa. В результате спонтанной гибридизации в Европе сформировались гибриды P.× eminens – результат скрещивания P. fruticosa и сбегающей из культуры (и по сути являющейся чужеродным видом) P. cerasus, а также P. × mohacsyana (P. fruticosa × P. avium). В отличие от дичающей P. cerasus, P. × mohacsyana не представляет угрозы для биоразнообразия, поскольку является стерильным таксоном. Во многих популяциях P. × eminens, однако, наблюдается бэккроссинг с родительским видом P. fruticosa и идет генетическая эрозия последнего (Macková et al., 2016).
В последнее время все большее внимание уделяется изучению гибридизации с точки зрения переноса определенных участков ДНК, экспрессии конкретных генов и влияния этих процессов на микроэволюцию образующихся новых таксонов (Mardis, 2008; Salmon et al., 2016; Rousseau et al., 2016). Так, североамериканский Helianthus annuus существенно расширил свой ареал к югу и встречается теперь в штате Техас, перекрывая естественный ареал другого североамериканского подсолнечника – H. debilis. Установлено, что в геноме H. annuus, произрастающего поблизости от местообитаний H. debilis, найдены аллели H. debilis (что показал анализ фрагментов и ядерной, и хлоропластной ДНК), – эти виды образуют гибриды, которые многократно скрещиваются с родительским видом H. annuus, и бэккроссы активно захватывают новые территории (Rieseberg et al., 2007).
Согласно математической модели Хакселя (Huxel, 1999), гибридизация с бесплодным потомством имеет небольшое влияние на замещение видов. Однако наблюдения за вегетативно размножающимся культигенным гибридом Reynoutria × bohemica Chrtek & Chrtková опровергают это положение. Гибридизация чужеродных для Европы видов R. sachalinensis и R. japonica привела в этом случае к генотипическим и геномным изменениям, повышению генетического разнообразия и стимулировала эволюцию новых таксонов (гибрида и бэккроссов) с высоким инвазионным потенциалом (Buhk, Thielsch, 2015). Вновь образованные генотипы закрепляются путем активного вегетативного размножения, и это существенно повысило способность комплекса Reynoutria к адаптации в условиях новой родины.
Таким образом, анализ научной литературы по проблеме влияния гибридизации на инвазионную активность чужеродных видов выявил как сведения в поддержку гипотезы Ч. Элтона об усилении гибридизационных процессов во вторичном ареале, так и информацию, противоречащую этой гипотезе. Более того, в большинстве случаев данные о гибридогенном характере того или иного таксона приведены только на основе сходства его морфологических признаков с характеристиками предполагаемых родительских видов. Вероятно, эти таксоны действительно возникли путем спонтанной гибридизации, но с уверенностью говорить об этом нельзя. Ведь морфологические изменения могли появиться как в результате микроэволюции чужеродных видов в новых условиях, так и вследствие влияния неблагоприятных экологических факторов (Галкина, Виноградова, 2016).
Диагностика чужеродных видов часто затруднена из-за дрейфа генов и возможной гибридизации с аборигенными или даже с другими близкородственными чужеродными видами. По традиционным диагностическим признакам не всегда возможно заключить, видим ли мы перед собой новую расу вида, возникшую в ходе микроэволюции, или же это новый гибрид, обладающий, возможно, более высоким инвазионным потенциалом, чем родительские таксоны, и нуждающийся в контроле его расселения и иных мерах борьбы.
В связи с этим целью наших многолетних исследований является прогнозирование скорости дальнейшего внедрения в естественные фитоценозы сформировавшихся во вторичном ареале таксонов гибридного характера экспериментальными и молекулярно-генетическими методами, а также тестирование гипотезы о более высокой конкурентоспособности и инвазионной активности гибридов по сравнению с родительскими видами. Параллельно мы проводим ревизию таксономически сложных гибридных комплексов с применением макро- и микроморфологических признаков, что позволяет оценить степень отличия предполагаемых гибридов от парентальных (родительских) видов.
Известно, что наиболее высокую инвазионную активность проявляют виды семейства Asteraceae (Виноградова и др., 2010), поэтому мы сосредоточили свое внимание на представителях этой группы чужеродных видов. Способность к гибридизации североамериканской Bidens frondosa с аборигенными B. tripartita и B. cernua мы уже подробно освещали в ряде работ (Галкина, Виноградова, 2016; Галкина и др., 2015). Гибридогенное происхождение так называемой Bidens × garumnae, якобы натурализующейся в среднем течении Волги (Васильева, Папченков, 2007, 2011), не подтверждено ни экспериментальным, ни молекулярно-генетическим методами. При выращивании на экспериментальном участке признаки, по которым ранее диагностировали этот таксон (“распластанная” форма роста и цельные листья), не наследовались. Более того, растения не имеют сходства с типовым гербарным образцом Bidens × garumnae Jeanj. & Debray [P]. Данный таксон следует, по-видимому, рассматривать как комплекс микровидов B. tripartita (Галкина и др., 2015).
Установлено, что череда, отмечаемая в последние десятилетия на территории Восточной Европы как “Bidens connata”, представляет собой не проникший из Северной Америки чужеродный вид (собственно B. connata Muhl. ex Willd.), а сформировавшийся во вторичном ареале комплекс гибридов и возвратных гибридов инвазионного вида североамериканского происхождения – B. frondosa – с аборигенной B. cernua. Этот таксон описан как B. decipiens Warnst. еще в 1895 г., типовой гербарий хранится в г. Эдинбурге [Е]. Действительно, семянки B. × decipiens покрыты волосками двух типов – дуплексными, из двух клеток (как у B. frondosa), и простыми многоклеточными (как у B. cernua). Кроме того, семянки B. × decipiens четырехгранные и имеют четыре ости (как у B. cernua) и покрыты бородавочками (как у B. frondosa). Корзинки у B. × decipiens по размерам и форме сходны с корзинками B. frondosa, а листья цельные, как у B. cernua. При выращивании на экспериментальном участке все эти признаки наследовались, молекулярно-генетический анализ подтвердил гипотезу о гибридогенном происхождении B. × decipiens (Виноградова, Галкина, 2015; Галкина, Виноградова, 2017).
Во вторичном ареале местонахождения B. × decipiens малочисленны и расположены на значительном расстоянии друг от друга. Через 50 лет после его описания этот таксон был собран во Франции и Чехии, в 1980-х – на Украине и в Беларусии. В России отмечен только в XXI в.: Московская обл. (2001), Калужская обл. (2012), Владимирская обл. (2013), Калининградская обл. (2013). Произрастает B. × decipiens в довольно труднодоступных районах, вдали от транспортных магистралей. Популяции немногочисленные и малой площади. На экспериментальном участке растения B. × decipiens росли медленнее, чем B. frondosa, формировали меньше корзинок и образовывали меньшее число семянок, т.е. были менее конкурентоспособны. Все это позволяет утверждать, что инвазионная активность B. × decipiens низка, и этот гибрид в ближайшем будущем не будет представлять угрозы естественным сообществам.
Еще одним объектом наших исследований были представители секции Conyza р. Erigeron. Растения с признаками, промежуточными между родительскими видами (североамериканским Erigeron canadensis и южноамериканскими E. bonariensis и E. sumatrensis), описал Ф. Сеннен в начале ХХ в. (Виноградова, Майоров, 2014). Разработанная авторами схема гибридизации таксонов, произрастающих во вторичном ареале родительских видов, представлена на рис. 1. В просмотренных нами гербарных хранилищах имеются лишь сборы начала ХХ в., а другие, более поздние находки, отсутствуют. Предпринятый нами в 2016 г. целенаправленный поиск растений с промежуточными признаками позволил обнаружить в Италии (Рим и Помпеи) и в Японии (Токусима, Киото, Токио) особи, схожие по комплексу признаков с Conyza × rouyana Sennen. С одной стороны, растения имели многорядную обертку корзинки с опушенными листочками обертки, и их побеги в целом были сильно опушены, как у E. sumatrensis. С другой стороны, растения имели мелкие, не вздутые в основании, корзинки с двурядной оберткой и зеленые линейно-ланцетные суженные к основанию листья с зубчато-пильчатым краем, что характерно для E. сanadensis. Однако предварительный молекулярно-генетический анализ с некоторыми праймерами не подтвердил гибридогенную природу этих образцов.
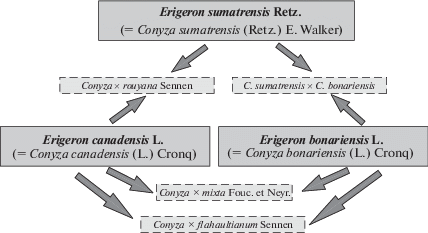
С уверенностью можно утверждать, что гибридные таксоны Erigeron sect. Conyza если и существуют, то в природе встречаются лишь единично, причем в пределах популяций родительских видов, и не проявляют тенденции к расселению. Семена растений с промежуточными признаками мы высеяли в 2017 г. на экспериментальный участок для более детального изучения с использованием методики, ранее примененной к другим таксонам.
В данной работе мы представляем результаты исследования гибридизационной активности представителей р. Solidago L.
ОБЪЕКТЫ И МЕТОДЫ
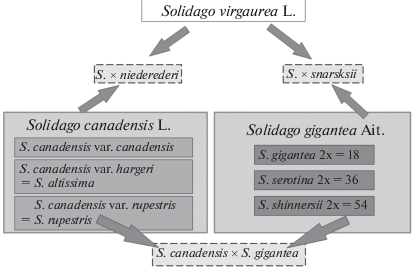
Североамериканские Solidago canadensis и S. gigantea из секции Solidago subsect. Triplinerviae входят в первую сотню (“Черную сотню”, Виноградова и др., 2015) наиболее агрессивных инвазионных видов России. Во вторичном ареале оба таксона представлены несколькими кариотипами с различным уровнем плоидности (Melville, Morton, 2011), нередко выделяемыми в качестве самостоятельных видов, и оба таксона способны гибридизировать с аборигенным золотарником S. virgaurea. Описаны S. × snarskisii Gudžinskas & Žalneravičius (= S. gigantea × × S. virgaurea) (Gudžinskas, Žalneravičius, 2016) и S. × niederederi Khek (= S. canadensis × S. virgaurea). В гербарии ГБС РАН [MHA] имеется образец, определенный А.А. Нотовым как гибрид S. gigantea × × S. canadensis, что нуждалось в подтверждении молекулярными методами. Разработанная авторами схема гибридизации представителей комплекса Solidago представлена на рис. 2.
Рис. 2.
Схема гибридизации в комплексе чужеродных и аборигенных видов рода Solidago (оригинальный рисунок). На белом фоне аборигенный вид S. virgaurea, на сером – чужеродные таксоны.
В ряде гербарных хранилищ России и Европы (H, С, DBN, DUB, S, UPS, PRC, PRA, LE, MW, MHA, KW, MSK, MSKU) просмотрены сборы таксонов из подсекции Triplinervae и проанализированы их макроморфологические диагностические признаки.
Исследование популяций в природе со сбором семян и гербария проведено в естественном ареале в США в штатах Пенсильвания и Миннесота (2005), а также во вторичном ареале S. gigantea, S. altissima и S.canadensis в Абхазии (2006, 2007), Венгрии (2011), Дании (2010), Словакии (2011), Белоруссии (2014) и Японии (2016). Предположительно гибридные особи, имеющие промежуточные признаки, были собраны нами в Московской обл. в парке Лосиный остров (2012), в Калужской обл. на окраине г. Калуги (2012), в Калининградской обл. (2013) и в Швеции в окрестностях г. Упсала (2014).
Использован классический морфологический метод с выявлением на собранном в природе массовом материале вариабельности макроморфологических признаков, позволяющих маркировать таксон. Наибольшее внимание уделяли строению соцветий и форме листовой пластинки (рис. 3, 4).
Рис. 3.
Розеточные листья различных таксонов рода Solidago L. а – S. canadensis, б – S. altissima, в – S. gigantea, г – S. gigantea c сизыми листьями, д – S. × niederederi, е – S. × snarskisii, ж – S. virgaurea.

Рис. 4.
Листья срединной формации различных таксонов рода Solidago L. а – S. canadensis, б – S. altissima, в – S. gigantea, г – S. gigantea c сизыми листьями, д – S. × niederederi, е – S. × snarskisii, ж – S. virgaurea.

В природе в спонтанных инвазионных популяциях исследовали по пять клонов, в каждом из которых отбирали по три растения. Выявляли среднее значение и амплитуду изменчивости 19 признаков вегетативных органов (форма листьев нижней формации, длина и ширина листьев нижней, средней и верхней формации, число зубчиков по краю листовой пластинки, общая длина побега, степень опушения стебля, степень опушения нижней и верхней стороны листьев нижней, средней и верхней формации, соотношение генеративной и вегетативной зон главной оси побегов, длина корневищ, число почек на единицу длины корневища) и 10 признаков генеративных органов (число веточек метелки, общее число корзинок в метелке, распределение корзинок по веточкам метелки в пределах соцветия, длина веточек метелки, длина “цветоножек” корзинок, длина корзинки, диаметр корзинки, соотношение длины обертки и длины корзинки, степень опушения оси соцветия (рахиса) и опушения обертки). Результаты исследования позволили четко различать таксоны в природных условиях (Виноградова, Квитка, 2006) и обнаружить впоследствии таксоны с промежуточными морфологическими признаками.
Существенным дополнением к этому методу стало использование микроморфологических признаков, выявленных с помощью цифрового микроскопа Keyence VHX-1000 и сканирующего электронного микроскопа LEO 1430 VP. Изучены 17 признаков вегетативных органов и 22 признака генеративной сферы растений, в частности, особенности кроющих трихом и спермодермы семянок, строение устьичного аппарата, размеры пыльцевых зерен (Виноградова, 2012; Vinogradova et al., 2017).
Проведено тестирование устойчивости макро- и микроморфологических признаков методом выращивания растений из семян различного происхождения на унифицированном агрофоне на экспериментальном участке в окрестностях г. Звенигород. Весной 2012 г. высеяны семена родительских таксонов: S. gigantea, S. altissima, S. canadensis и S.virgaurea, собранные в Московской обл. и в Венгрии. Осенью 2012 г. высеяны семена предполагаемых гибридов и их совместно произраставших родительских таксонов, собранные в окрестностях г. Калуги. В течение последующих лет у выросшего из семян потомства F1 определяли темпы роста, фенологический ритм развития и степень наследуемости морфологических признаков.
Гибридогенный характер образцов, имеющих промежуточные признаки, подтверждался молекулярно-генетическим методом. Для этого выделено 26 образцов ДНК из сухих листьев изучаемых таксонов в различных местообитаниях на территории Европы (табл. 1). Также мы провели дополнительный анализ с участием еще 28 образцов, преимущественно потомства F1 этих предполагаемых гибридов (табл. 2), выросших на экспериментальном участке, включив в него образцы 1 (а, б, в), 2 (а, б, в), 3, 10 и 11 из предыдущего анализа (это те же образцы, что указаны под данными номерами в табл. 1).
Таблица 1.
Перечень выделенных образцов ДНК – предполагаемые гибриды рода Solidago L. и родительские виды
| № образца |
Вид | Местонахождение и год сбора |
|---|---|---|
| 1 (а, б, в) | S. virgaurea | Московская обл., национальный парк Лосиный остров, 2014 г. |
| 2 (а, б, в) | S. gigantea | |
| 3 | S.×snarskisii | |
| 4 (а, б, в) | Калининградская обл., Зеленоградский р-н, окрестности пос. Цветное, залежь, 2013 г. | |
| 5 (а, б) | S. gigantea | |
| 6 | S. gigantea (сизая форма) |
Калининградская обл., Куршская коса, пос. Рыбачий, опушка леса, 2013 г. |
| 7 | S. canadensis | Беларусь, Ганцевичи, 2014 г. |
| 8 | S. canadensis (слабоопушенная) |
Беларусь, Ганцевичи, плантация клюквы, полученной из США, 2014 г. |
| 9 | S. canadensis × S. gigantea | Тверская обл., пос. Турчиново, 2009 г. |
| 10 | S. × snarskisii (опушенная форма, морфологическое сходство с S. niederederi) | Потомство растений из Калужской обл. (берег р. Ока), выращенное на экспериментальном участке в Московской обл. (окрестности г. Звенигород) |
| 11 | S. canadensis s.stricta (потомство) | Потомство растений из Венгрии, выращенное на экспериментальном участке в Московской обл. (окрестности г. Звенигород) |
| 12 (а, б) | S. canadensis s.stricta | Венгрия, в степи вдоль железной дороги, 2011 г. |
| 13 (а, б) | S. canadensis | |
| 14 | Solidago sp. (c колосовидным соцветием) | Абхазия, Сухумский р-н, Гульрипш, 2009 г. |
| 15 | S. rupestris | Абхазия, Каман, 2009 г. |
| 16 (а, б) | Абхазия, Сухум, 2007 г. |
Примечание. Буквами здесь и в табл. 2 обозначены разные экземпляры одного таксона из одной и той же локальной популяции.
Таблица 2.
Перечень выделенных образцов ДНК – потомство предполагаемых гибридов и некоторые родительские таксоны
| № образца |
Вид | Местонахождение |
|---|---|---|
| 1 (а, б, в, г) | S. virgaurea | Национальный парк Лосиный остров |
| 2 (а, б, в) | S. gigantea | |
| 3 | S. × snarskisii | |
| 4 (а, б, в, г) | S. gigantea | Потомство растений из Московской обл. (Хлебниково), выращенное на экспериментальном участке в Московской обл. (окрестности г. Звенигород) |
| 5 (а, б, в, г) | S. × niederederi | Калужская обл., берег р. Ока |
| 6 | S. virgaurea | Потомство растений из Московской обл. (Хлебниково), выращенное на экспериментальном участке в Московской обл. (окрестности г. Звенигород) |
| 7 (а, б, в, г) | Потомство растений из Калужской обл.(берег р. Ока), выращенное на экспериментальном участке в Московской обл. (окрестности г. Звенигород) | |
| 8 | S. graminifolia | Клон из ботанического сада Брно, выращенный на экспериментальном участке в окрестностях г. Звенигород, происхождение растения – Канада |
| 9 (а, б, в, г, д) | S. × snarskisii | Потомство растений из Калужской обл. (берег р. Ока), выращенное на экспериментальном участке в Московской обл. (окрестности г. Звенигород) |
| 10 | S. × snarskisii (опушенная форма, морфологическое сходство с S. niederederi) | |
| 11 | S. canadensis s. stricta | Потомство растений из Венгрии, выращенное на экспериментальном участке в Московской обл. (окрестности г. Звенигород) |
| 12 (а, б, в, г, д) | S. canadensis | |
| 13 | S. canadensis | Швеция, Ботанический сад Линнея |
| 14 | S. niederederi | Швеция, Упсала, Ботанический сад университета |
| 15 | S. × snarskisii | Швеция, Упсала |
| 16 | S. flexicaulis | Швеция, Упсала, Ботанический сад университета |
| 17 | S. sempervirens | Швеция, Ботанический сад Линнея |
Выделение ДНК проводилось CTAB-методом (Rogers, Bendich, 1985). Оценку гибридной природы образцов проводили с использованием фрагментного анализа межмикросателлитных участков ДНК (Inter Simple Sequence Repeats (ISSR)). Анализ проводили дважды – для природных популяций (28 образцов) и потомства F1 с экспериментального участка, при этом несколько образцов совпадали (табл. 1, 2). Полимеразную цепную реакцию (ПЦР) проводили в амплификаторе DNA Engine Dyad Peltier Thermal Cycler (Biorad, США). Для каждого праймера использовали разную температуру отжига. Предварительная денатурация проходила в течение 3 мин при 94°С. Элонгация длилась 30 с при 94°С, потом 30 с при 55°С с (САG)5 [М7], при 51°С с (AG)8YA [UBC 836], при 45°С с (GA)8 YG [UBC 841], при 50°С в случае с праймером (AC)8 (C/T)G [M2] и при 60° с праймером DBD(AC)7 [UBC 889]. Затем элонгация проходила в течение 1 мин при 72°С. Описанный процесс повторялся 35 раз, причем в каждый последующий раз элонгация длилась на 2 с дольше при 72°С. Завершающей стадией была элонгация 3 мин при 72°С. Продукты амплификации разделяли путем электрофореза в 1.7%-ном агарозном геле с добавлением бромида этидия в 1×ТВЕ-буфере в течение 1 ч. Затем фрагменты ДНК были сфотографированы в ультрафиолетовом свете. Для определения их длин использовался маркер молекулярной массы 100bp+. Обработку полученных данных производили в программе Cross Checker (Buntjer, 2000). Полученные данные были представлены в виде матрицы бинарных признаков, в которых наличие или отсутствие определенного фрагмента рассматривалось соответственно как 1 или 0. Полученные матрицы из 72 фрагментов ДНК для гибридов и 56 фрагментов для их потомства были обработана в программах Structure 2.2 и PAST 2.0 методом кластерного анализа (UPGMA) с использованием дистанции Жаккара. Дополнительно был проведен анализ данных методом Байеса (MCMC – Markov Chain Monte Carlo) в программе NewHybrids. Программа оценивает гибридный статус анализируемых растений по соответствию распределения фрагментов ДНК у предполагаемых родительских видов, гибридов первого и второго поколений и возвратных гибридов (бэккроссов).
В качестве внешней группы для анализа потомства, выросшего на экспериментальном участке, использованы следующие виды р. Solidago: S. graminifolia (L.) Salisb., S. sempervirens Michx. и S. flexicaulis L.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Хотя в природе определение S. gigantea, S. altissima и S. canadensis нередко затруднено, культивирование этих видов в однородных условиях экспериментального участка позволяет четче выявить их диагностические признаки. Виды различаются по темпу и продолжительности периода роста, фенологическому ритму развития, размерам и форме листьев, характеру опушения побегов, строению побеговых систем, по числу, размерам и форме корзинок, а также по обертке корзинок. Обнаружены и различия по микроморфологическим признакам (Виноградова, 2012; Vinogradova et al., 2017).
У гибридов диагностические признаки также стабильны: S. × niederederi имеет метельчатое соцветие, опушенные побеги и мелкие корзинки, как S. canadensis, и крупные розеточные листья в основании побега, как S. virgaurea. S. × snarskisii отличается от S. × niederederi неопушенными побегами и более крупными корзинками.
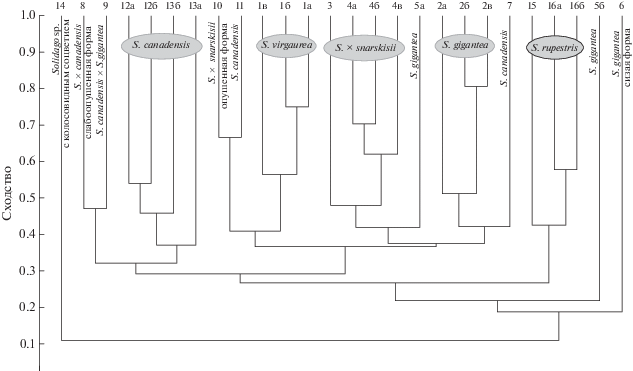
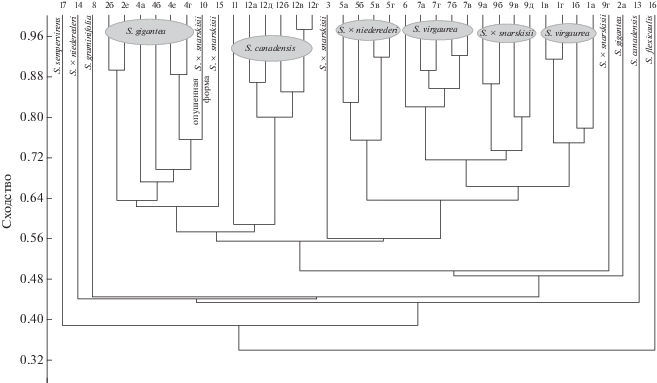
По результатам кластерного анализа, проведенного в программе PAST (рис. 5), растения разделились на два крупных кластера, также четко отделились образцы 10 и 11 – соответственно, опушенная форма гибрида S. × snarskisii и потомство S. canadensis s. stricta, выращенное в Московской обл. из семян, собранных с растений в Венгрии. И хотя дальнейший анализ в программе NewHybrids подтвердил гибридогенное происхождение образца 10, морфологически он сильно отличался от других особей S. × snarskisii, поэтому, вероятно, является представителем особого экотипа. В одном кластере оказались и их предполагаемые гибриды. В свою очередь он разделился на три субкластера – в один из них вошли растения S. virgaurea, в другой S. gigantea, собранные в Лосином острове, а в третий –гибриды из Лосиного острова и Калининградской обл. и одна из особей S. gigantea из Калининградской обл. В другой крупный кластер вошли все остальные растения – все таксоны S. canadensis (включая собранную в Венгрии S. canadensis s. stricta), предполагаемый гибрид S. canadensis × S. gigantea (собранный в Тверской обл. А.А. Нотовым), собранные в Абхазии S. rupestris и Solidago sp. с колосовидным соцветием, а также собранные в Калининградской обл. S. gigantea за исключением образца 5а. Таким образом, кластерный анализ четко разделил родительские виды, а практически все образцы S. × snarskisii оказались расположены между ветвями родительских видов, за исключением образца 10, который отличается от остальных особей S. × snarskisii и морфологически, поэтому его таксономическая принадлежность изначально вызывала сомнение.
Рис. 5.
Кластерный анализ фрагментов ДНК для предполагаемых гибридов Solidago gigantea × S. canadensis, родительских видов и некоторых других представителей р. Solidago. Номера образцов см. в табл. 1.
Полученную матрицу бинарных признаков по всем образцам, помимо PAST, проанализировали в программе Structure (рис. 6). С максимальной апостериорной вероятностью число популяций k = 3 (при k = 3 значение lnP(D) достигает максимума и составляет –1464.6, тогда как при k = 2 этот логарифм равен –1479.4 и –1466.5 при k = 4). Таким образом, наблюдаются значимые генетические различия между тремя группами растений. В первой группе оказались растения S. rupestris, собранные в Абхазии, за исключением одной особи. Выявлены достаточно большие внутрипопуляционные различия между особями S. virgaurea, программа отнесла их к разным группам (тогда как они расположены в непосредственной близости на диаграмме кластерного анализа в PAST). Образец 1в оказался в одной группе с S. gigantea и гибридами S. × snarskisii, он же программой NewHybrids (рис. 7) отнесен не к чистым видам, а к бэккроссам. Все предполагаемые гибриды S. × snarskisii отнесены в одну группу с растениями S. gigantea, что указывает на их близкое родство и вероятность того, что они все-таки являются не гибридами, а бэккроссами, т.е. возвратными гибридами S. × snarskisii на родительскую североамериканскую S. gigantea. Это совпадает с результатами обработки в NewHybrids для данных видов, в которой они отнесены к возвратным гибридам с вероятностью более 50% (рис. 7).
Рис. 6.
Результаты анализа в Structure. Номера образцов см. в табл. 1. Solidago sp. 1 – собранный в Абхазии экземпляр с колосовидным соцветием, Solidago sp. 2 – предполагаемый гибрид S. canadensis × S. gigantea.
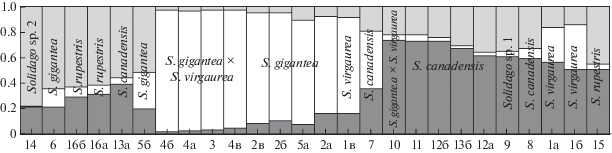
Рис. 7.
Результаты анализа в NewHybrids. Номера образцов см. в табл. 1. а – Bx2 – бэккросс S. × snarskisii на родительский вид S. virgaurea, Bx1 – бэккросс S. × snarskisii на родительский вид S. gigantea, F1 и F2 – гибриды первого поколения (один из них S. × snarskisii), Sp2 – S. virgaurea, Sp1 – S. gigantea. б – Bx2 – бэккросс гибрида S. canadensis × S. gigantea и родительского вида S. canadensis, Bx1 – бэккросс гибрида S. canadensis × × S. gigantea и родительского вида S. gigantea, F1 и F2 – гибриды первого поколения (один из них – S. canadensis × × S. gigantea), Sp2 – S. canadensis, Sp1 – S. gigantea.
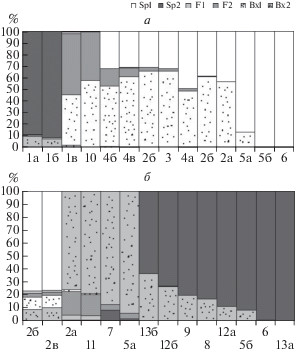
Обработка данных в NewHybrids показала (рис. 7а), что один из образцов S.virgaurea из Лосиного острова – гибрид (остальные чистые виды), а S. gigantea c одинаковой степенью вероятности можно отнести и к бэккроссам (т.е. возвратным гибридам), и к чистым видам. Образец 1в (S. virgaurea) оказался отнесен к гибридам, что, по всей видимости, является скорее не погрешностью работы программы, а наличием значительной межпопуляционной изменчивости этого полиморфного таксона. Все калининградские растения S. gigantea – чистые виды. Все растения, предположительно определенные нами как гибриды S. × snarskisii, являются возвратными гибридами (т.е. результатом скрещивания гибридов S. × snarskisii с родительской S. gigantea).
Образец 9, собранный А.А. Нотовым, по нашим данным не является гибридом S. canadensis и S. gigantea: по результатам обработки в NewHybrids это S. canadensis (рис. 7б).
Мы рассмотрели также другую группу образцов, преимущественно потомство F1 предполагаемых гибридов и родительских видов, выращенное в однородных условиях, а также некоторые другие виды, собранные в ботанических садах Швеции. Кластерный анализ показал (рис. 8), что выделились два небольших кластера, куда вошли несколько видов, собранных в ботанических садах Швеции, и один гибрид (S. flexicaulis, S. graminifolia, S. sempervirens, S. × niederederi) и третий с большинством образцов, внутри которого субкластеры выделились преимущественно по видовой принадлежности, за исключением двух образцов предполагаемых гибридов S. × snarskisii: 10 (опушенная форма, потомство собранных в г.Калуге растений) и 15 (образец, собранный в Швеции). В данном случае образцы S. × snarskisii разбросаны по разным кластерам, что не удивительно, поскольку для гибридов характерна значительная изменчивость, а последующий анализ в NewHybrids подтверждает гибридную природу этой группы.
Рис. 8.
Кластерный анализ фрагментов ДНК для потомства F1 некоторых представителей рода Solidago. Номера образцов см. в табл. 2.
Дальнейший анализ потомства в программе NewHybrids (рис. 9а) показал, что образцы 3 и 10 являются гибридами, они были проанализированы вместе с родительскими таксонами (рис. 7а), и этот анализ выявил их уже как бэккроссы, а образцы, являющиеся потомством S. gigantea × S. virgaurea, с большей вероятностью отнесены к родительским видам и с меньшей вероятностью, составляющей менее 20% (а для образцов 9а, б, в, г, д – и менее 10%), – к бэккроссам. При этом вероятнее всего, образец 15 действительно может быть бэккроссом – результатом скрещивания S. × snarskisii и S. gigantea (хотя вероятность составляет всего 15%), поскольку анализ предыдущей группы образцов, не включая потомство, показал, что, вероятно, имеет место обратный процесс – скрещивание гибридов S. × snarskisii с родительским видом S. gigantea.
Рис. 9.
Результаты анализа в NewHybrids. Номера образцов см. в табл. 2. а – Bx2 – бэккросс S. × snarskisii × S. virgaurea, Bx1 – бэккросс S. × snarskisii × S. gigantea, F1 и F2 – гибриды первого поколения (один из них – S. × snarskisii), Sp2 – S. virgaurea, Sp1 – S. gigantea. б – Bx2 – бэккросс S. × niederederi × S.canadensis, Bx1 – бэккросс S. × niederederi × S. virgaurea, F1 и F2 – гибриды первого поколения (один из них – S. × niederederi), Sp2 – S. canadensis, Sp1 – S. virgaurea.
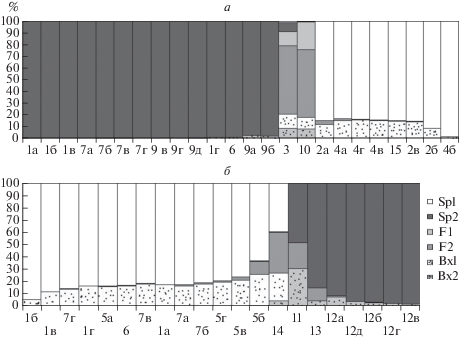
Что касается доказательства гибридогенного происхождения собранных нами растений S. niederederi (рис. 9б), то было выявлено, что образец из г. Упсалы действительно с вероятностью 33% является гибридом S. canadensis × S. virgaurea (но с вероятностью 22% отнесен программой к бэккроссу и с вероятностью 39% – к родительскому виду S. virgaurea). Гибридным оказалось потомство S. canadensis s. stricta, выращенное в окрестностях г. Звенигорода из семян, собранных в Венгрии, но, вероятно, это является неточностью в работе программы. Гибридный статус S. × niederederi подтвердился, но большинство особей было отнесено программой к бэккроссам S. niederederi × S. virgaurea.
Таким образом, гибридное происхождение S. × snarskisii и S. × niederederi подтверждено как экспериментальным, так и молекулярно-генетическим методами.
За первые 100 лет, прошедших с первоописания S. × niederederi из Верхних Альп в 1905 г., этот таксон расширил свой ареал незначительно. Его немногочисленные популяции были отмечены в других регионах Австрии, в Великобритании, Литве и Польше (Pagitz, 2016). Однако в последнее десятилетие картина несколько изменилась: в Польше площадь ареала S. × niederederi составляет уже 3 тыс. км2 (Pliszko, Zając, 2016). В Альпах она продвинулась в Западный Тироль и Каринтию (Австрия) и в Южный Тироль (Италия), и численность ее популяций значительно возросла (Pagitz, 2016). В Литве этот вид зафиксирован в 27 местообитаниях (Karpavičienė, Radušienė, 2016). В России, однако, S. × niederederi собрана лишь в двух местообитаниях (Тверская и Калужская области), да и там она встречается единично.
Второй гибрид, S. × snarskisii, встречается еще реже. Он описан на основании сборов в Литве (Gudžinskas, Žalneravičius, 2016), но ранее одна особь этого таксона была обнаружена С.Р. Майоровым в Московской обл. в национальном парке Лосиный остров (2011 г.) и два экземпляра – на окраине г. Калуги (2012 г.). Две немногочисленные популяции (не более десятка особей каждая) отмечены в пос. Цветное и пос. Логвино Калининградской обл. в 2013 г. и две особи – в г. Упсала (Швеция) в 2014 г. (MHA, MW). Интересно, что особи, имеющие промежуточные признаки (в частности, разветвленное соцветие), наши коллеги находили еще раньше, однако при выращивании двух поколений сеянцев из их семян на экспериментальном участке выяснялось, что мы имеем дело не с гибридом, а с родительской S. virgaurea.
Всхожесть семянок и S. × niederederi, и S. × snarskisii крайне низка: из всех семянок, собранных с одного соцветия (более 10 тыс.), проросло для первого таксона 8 штук, а для второго – 6! На экспериментальном участке особи F1 формируют более короткие корневища, чем родительские таксоны, и разрастаются медленнее. Угрозы их массового внедрения в естественные фитоценозы вторичного ареала пока нет.
ЗАКЛЮЧЕНИЕ
Не все таксоны с промежуточными морфологическими признаками являются гибридами. Ни экспериментальным, ни молекулярно-генетическим методами нами не подтверждено гибридное происхождение “S. сanadensis × S. gigantea”, как ранее не подтвердилось и гибридогенное происхождение другого предполагаемого гибрида “В. garumnae”.
В отличие от классических примеров активно натурализующихся Reynoutria × bohemica и Spartina × anglica, все изученные нами гибриды (Bidens × decipiens, Solidago × niederederi, S. × snarskisii и Conyza × rouyana) встречаются пока во вторичном ареале родительских видов единично и менее конкурентоспособны, чем их предковые формы. Тенденцию к натурализации начала проявлять в Центральной Европе лишь S. × niederederi, которая за последние 5 лет несколько расширила ареал в Польше, Литве и в Австрии. Таким образом, гибриды внутри р. Solidago не подтверждают гипотезу о большей конкурентоспособности и инвазионной активности гибридов по сравнению с родительскими видами, но, с другой стороны, и не опровергают ее полностью (по крайней мере, расширение ареала S. × niederederi требует изучения в ближайшее десятилетие).
Авторы выражают признательность анонимному рецензенту данной статьи, замечания которого помогли нам существенно улучшить текст. Исследования выполнены в рамках госзадания ГБС РАН “Биологическое разнообразие природной и культурной флоры: фундаментальные и прикладные вопросы изучения и сохранения” (№118 021 490 111-5) при частичной финансовой поддержке Российского фонда фундаментальных исследований, проект № 18-04-00411.
Список литературы
Васильева Н.В., Папченков В.Г., 2007. Распространение в бассейне Волги и биологические особенности Bidens frondosa L. // Естественные и инвазийные процессы формирования водных и наземных экосистем. Ростов-на-Дону: Юж. науч. центр РАН. С. 64–65.
Васильева Н.В., Папченков В.Г., 2011. Механизмы воздействия инвазионной Bidens frondosa L. на аборигенные виды череды // Рос. журн. биол. инвазий. № 1. С. 15–22.
Виноградова Ю.К., 2012. Биоразнообразие таксонов рода Solidago L. по микроморфологическим признакам // Биоразнообразие: проблемы изучения и сохранения. Тверь: Твер. гос. ун-т. С. 346–350.
Виноградова Ю.К., Квитка А.Ю., 2006. Инвазионные виды рода Solidago во флоре Средней России // Флористические исследования в Средней России. М.: Т-во науч. изд. КМК. С. 74–78.
Виноградова Ю.К., Майоров С.Р., 2014. История интродукции, микроэволюция и видообразование в роде Conyza Less. во вторичном ареале // Современная ботаника в России. Т. 4. Тольятти: Кассандра. С. 61–63.
Виноградова Ю.К., Галкина М.А., 2015. О возможности гибридогенного происхождения Bidens connata // XIII Московское совещание по филогении растений. 50 лет без К.И. Мейера. М.: Макс Пресс. С. 64–69.
Виноградова Ю.К., Майоров С.Р., 2015. Длительность lag–фазы как отражение микроэволюции растений во вторичном ареале // XIII Московское совещание по филогении растений. 50 лет без К.И. Мейера. М.: Макс Пресс. С. 70–74.
Виноградова Ю.К., Галкина М.А., 2016. Таксоны рода Bidens L. в Средней России (изменчивость и наследуемость биоморфологических признаков) // Вестн. Удм. ун-та. Сер. биол. Т. 26. Вып. 1. С. 53–62.
Виноградова Ю.К., Майоров С.Р., Хорун Л.В., 2010. Черная книга флоры Средней России. М.: “Геос”. 512 с.
Виноградова Ю.К., Абрамова Л.М., Акатова Т.В. и др., 2015. “Черная сотня” инвазионных растений России // Инф. бюл. совета бот. садов стран СНГ. Вып. 4 (27). С. 85–89.
Галкина М.А., Виноградова Ю.К., 2017. Гибридизация некоторых чужеродных таксонов рода Bidens L. в Восточной Европе // Изв. РАН. Сер. биол. № 4. С. 1–7.
Галкина М.А., Виноградова Ю.К., Шанцер И.А., 2015. Биологические особенности и микроэволюция инвазионных видов рода Bidens L. // Изв. РАН. Сер. биол. № 4. С. 382–392.
Грант В., 1984. Видообразование у растений. М.: Мир. 528 с.
Abbott R.J., James J.K., Milne R.I. et al., 2003. Plant introduction, hybridization and gene flow // Phil. Trans. R. Soc. Lond. V. 358. P. 1123–1132.
Ainouche M., Chelaifa H., Rousseau-Guetin M. et al., 2016. Genome merger as evolutionary springboard: Insights from recurrent hybridization and polyploidy in Spartina// International Conference on Polyploidy, Hybridization and Biodiversity. Rovinj: Croatian Biol. Society. P. 13.
Albuquerque G.S., Tauber C.A., Tauber M.J., 1996. Postmating reproductive isolation between Chrysopa quadripunctata and Chrysopa slossonae: mechanisms and geographic variation // Evolution. V. 50. P. 1598–1606.
Baumel A., Ainouche M.L., Misset M.T. et al., 2003. Genetic evidence for hybridization between the native Spartina maritima and the introduced Spartina alterniflora (Poaceae) in South-West France: Spartina × neyrautii re-examined // Plant. Syst. Evol. V. 237. P. 87 –97.
Bleeker W., 2003. Hybridization and Rorippa austriaca invasion in Germany // Mol. Ecol. V. 12. I. 7. P. 1831–1841.
Bleeker W., Schmitz U., Ristow M., 2007. Interspecific hybridization between alien and native plant species in Germany and its consequences for native biodiversity // Biol. Conserv. V. 137. I. 2. P. 248–253.
Brouillet L., Semple J.C., Allen G.A. et al., 2006. Symphyotrichium Nees // Flora North America. V. 20. Asteraceae. Pt 2. P. 465–539.
Buhk C., Thielsch A., 2015. Hybridisation boosts the invasion of an alien species complex: insights into future invasiveness // Persp. Plant Ecology, Evolution, Systematics. V. 17. № 4. P. 274–283.
Buntjer J.B., 2000. Cross Checker: computer assisted scoring of genetic AFLP data // Plant, Animal Genome VIIIth Conf. S. Diego, CA, January, 9–12, 2000. http://wheat.pw.usda.gov/jag/papers99/paper599/indexp599.html.
Gudžinskas Z., Žalneravičius E., 2016. Solidago × snarskisii nothosp. nov. (Asteraceae) from Lithuania and its position in the infrageneric classification of the genus // Phytotaxa. V. 253 (2). P. 147–155.
Guénégou M.C., Levasseur J.E., 1993. La nouvelle espèce amphiloïde Spartina anglica C.E. Hubbard: son origine, argumentation et implications // Biogeographica. V. 69. P. 125–133.
Ellstrand N.C., Shierenbeck K.A., 2000. Hybridization as a stimulus for the evolution of invasiveness in plants? // Proc. Nat. Amer. Soc. V. 97. № 13. P. 7043–7050.
Elton C.S., 1958. The ecology of invasions by animals and plants. L.: Methuen. 212 p.
Heywood V.H., 1989. Patterns, extents and modes of invasions by terrestrial plants. Biological Invasions: a Global Perspective / Ed. Drake J.A. Chichester: John Wiley & Sons Ltd. P. 31–60.
Huxel G.R., 1999. Rapid displacement of native species by invasive species: effects of hybridization // Biol. Conservation. V. 89. P. 143–152.
Karpavičienė B., Radušienė J., 2016. Morphological and anatomical characterization of Solidago × niederederi and other sympatric Solidago species // Weed Science. V. 64. I. 1. P. 61–70.
Knope M.L., Pender R.J., Crawford D.J. et al., 2013. Invasive congeners are unlikely to hybridize with native Hawaiian Bidens (Asteraceae) // Amer. J. Botany. V. 100. I. 6. P. 1221–1226.
Levin D.A., Francisco-Ortega J., Jansen R.K., 1996. Hybridization and the extinction of rate plant species // Cons. Biol. V. 10. P. 10–16.
Macková L., Vit P., Ďurišová L., P. Eliáš et al., 2016. Hybridization success is largely limited to homoploid Prunus hybrids – multidisciplinary approach // International Conference on Polyploidy, Hybridization and Biodiversity. Rovinj: Croatian Biol. Society. P. 113.
Mardis E.R., 2008. Next-generation DNA sequencing methods // Annual review of genomics and human genetics. V. 9. P. 397–402.
Melville R.M., Morton J.K., 2011. A biosystematic study of the Solidago canadensis (Compositae) complex. I. The Ontario populations // Can. J. Botany. V. 60. № 6. P. 976–997.
McGeoch M.A., Butchart C.H.M., Spear D. et al., 2010. Global indicators of biological invasion: species numbers, biodiversity impact and policy responces // Diversity Distributions. № 16. P. 95–108.
Pagitz K., 2016. Solidago x niederederi (S. canadensis x S. virgaurea ssp. virgaurea) in the Eastern Alps // Neobiota 2016. Vianden: Fond faune-flore. P. 194.
Pliszko A., Zając M., 2016. Current and potential distribution of Solidago ×niederederi (Asteraceae) in Poland // Neobiota 2016. Vianden: Fond faune-flore. P. 163.
Radosavliević I., Bogdanović S., Filipović M. et al., 2016. Contrasting patterns of phenotypic and genetic variation in hybrid offspring: causes and consequences // International Conference on Polyploidy, Hybridization and Biodiversity. Rovinj: Croatian Biol. Society. P. 97.
Rogers S.O., Bendich A.J., 1985. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues // Plant Mol. Biol. № 5. P. 69–76.
Rhymer J.M., Simberloff D., 1996. Extinction by hybridization and introgression // Ann. Rev. Ecol. and Syst. V. 27. P. 83–109.
Rieseberg L.H., Kim S-C., Randell R.A. et al., 2007. Hybridization and the colonization of novel habitats by annual sunflowers // Genetica. V. 129. P. 149–165.
Rousseau H., Dauvergne X., Boutte J. et al., 2016. Polyploidy and phenotypic novelty: Phylogenetic context of DMSP (dimethylsulfoniopropionate) biosynthesis in Spartina (Poaceae, Chloridoideae) // International Conference on Polyploidy, Hybridization and Biodiversity. Rovinj: Croatian Biol. Society. P. 59.
Salmon A., Boutte J., Rousseau H. et al., 2016. How to detect duplicated sequences within highly polyploid species without any reference? An introduction to the use of the Pyro- and Illu-haplotyper pipelines for Spartina genomics // International Conference on Polyploidy, Hybridization and Biodiversity. Rovinj: Croatian Biol. Society. P. 29.
Tokhtar V.K., Vinogradova Yu.K., Groshenko A.S., 2011. Microevolution and invasiveness of Oenothera L. species (Subsect. Oenothera, Onagraceae) in Europe // Russian J. Biol. Invasions. V. 2. № 4. P. 273–280.
Vinogradova Yu.K., Ryabchenko A.S., Galkina M.A., 2017. Achenial trichomes in invasive european Asteraceae: the taxonomic relevance // Annals Botany. V. 102. № 4. P. 730–741.
Williamson M., 1996. Biological Invasions. L.: Chapman & Hall. P. 33.
Zalapa J.E., Brunet J., Guries R.P., 2009. Patterns of hybridization and introgression between invasive Ulmus pumila (Ulmaceae) and native U. rubra // Amer. J. Botany. V. 96. P. 1116–1128.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии