

Журнал общей биологии, 2019, T. 80, № 2, стр. 145-160
Растительные сообщества суровых и благоприятных местообитаний: особенности организации, структура доминирования и ее связь с видовым богатством
В. В. Акатов 1, *, Т. В. Акатова 2, С. Г. Чефранов 1
1 Майкопский государственный технологический университет
385000 Майкоп, Первомайская, 191, Россия
2 Кавказский государственный природный биосферный заповедник
385000 Майкоп, Советская, 187, Россия
* E-mail: akatovmgti@mail.ru
Поступила в редакцию 16.04.2018
После доработки 07.06.2018
Принята к публикации 03.07.2018
Аннотация
Cчитается, что растительные сообщества суровых и благоприятных местообитаний организованы разным способом. Первые – преимущественно абиотической средой и случайными процессами (S‑модель), вторые – преимущественно конкуренцией (C-S-R-модель). Можно ожидать, что особенности организации оказывают влияние на структуру доминирования ценозов и характер ее связи с видовым богатством. Цель нашего исследования – проверить это предположение на примере травяных сообществ относительно стабильных местообитаний различных районов и высотных поясов Западного Кавказа и Предкавказья (альпийских ковров, пустошей и лугов, субальпийских лугов и болот, нижнегорных лугов, степей, травяного яруса лесов). В качестве критерия степени суровости среды использовали надземную биомассу травостоя в период его максимального развития. Относительное участие видов в травостое оценивали как отношение их биомассы к общей биомассе травостоя на площадке (D) или к биомассе травостоя, оставшейся после удаления из нее биомассы видов более высокого ранга (K). Под структурой доминирования понимали относительное участие в травостое вида первого ранга (степень доминирования, индекс Бергера–Паркера), а также соотношение значений D и K для двух-трех видов с наибольшим участием в травостое. Результаты показали: 1) если структуру доминирования оценивать на основе величины D, то она варьирует в широких пределах и почти не связана с условиями среды; 2) в C-S-R-ценозах значение К для видов 1-го ранга (К1) в среднем выше, чем для менее значимых видов (K2 и K3), в S-ценозах они в среднем примерно одинаковы; при этом, если рассматривать C-S-R- и S-ценозы отдельно, то величина (K1 – K2) не связана с продуктивностью местообитаний; 3) пространственная изменчивость структуры доминирования в целом несколько выше на участках суровых местообитаний, чем благоприятных; 4) C‑S-R-ценозы характеризуются более тесной связью между степенью доминирования и видовым богатством, чем S-ценозы. Сделано предположение, что величину (K1 – K2) можно рассматривать в качестве ценотического показателя степени выраженности у доминантов черт С-стратегии, а значит, и в качестве критерия при отнесении растительных сообществ к С-R-S- или S-модели.
Под средой обитания растений понимают совокупность окружающих условий, прямо или косвенно влияющих на их произрастание. Если она характеризуется низким уровнем ресурсов (элементов минерального питания, света, воды), экстремальными температурами, коротким периодом вегетации, высокой или низкой кислотностью почвы или присутствием в ней токсинов (тяжелых металлов, солей и т.д.), то ее можно рассматривать как суровую; в противном случае – как благоприятную (Grime, 1977; Бигон и др., 1989; Boscaiu et al., 2008; Миркин, Наумова, 2012; Zefferman et al., 2015).
Считается, что растительные сообщества суровых и благоприятных местообитаний организованы разным способом (Grime, 1977; Работнов, 1983; Grace, 1991; Cornell, Lawton, 1992; Cornell, 1993; Olff, Bakker, 1998; Mangla et al., 2011; Paquette, Messier, 2011; Миркин, Наумова, 2012; Hejda et al., 2016). В первом случае они характеризуются низкой интенсивностью межвидовой конкуренции (Grime, 1977; Работнов, 1983; Peet, Christensen, 1988; Cornell, Lawton, 1992; Lamb, Cahill, 2008; Миркин, Наумова, 2012, и др.). Это предполагает, что плотность популяций в таких ценозах в значительной мере регулируется абиотическими условиями среды, популяционная динамика каждого вида не зависит от других видов, их колонизация новыми видами является случайным процессом (Cornell, Lawton, 1992; Cornell, 1993; Павлов и др., 1998; Миркин, Наумова, 2012). Решающее значение для формирования таких сообществ имеет размер пула толерантных видов, вероятность попадания тех или иных из них в то или иное конкретное место, а также их дальнейшего выживания вне зависимости от ближайших соседей (Yodzis, 1978; Гиляров, 2010; Онипченко, 2013). Доминируют в них виды, которые, как и другие, являются слабыми конкурентами, но более остальных толерантны к воздействию абиотических (стрессовых) факторов (S-стратеги, по: Grime, 1977; патиенты, по: Раменский, 1938). Но чем благоприятнее условия среды, тем выше биомасса и продукция сообществ, а соответственно плотность и высота травостоя, тем выше интенсивность межвидовой конкуренции и, как следствие, вероятность вытеснения (замещения) особей одних видов другими (Grime, 1977; Yodzis, 1978; Huston, 1979; Peet, Christensen, 1988; Bengtsson et al., 1994; Piper, 1995; Drobner et al., 1998; Sammul et al., 2000, 2006; Mulder et al., 2004; Poggio, Ghersa, 2011; Миркин, Наумова, 2012; Онипченко, 2013). Соответственно плотность популяций в сообществах благоприятных местообитаний в значительной мере регулируется межвидовыми взаимоотношениями (Миркин, Наумова, 2012). Доминируют в них конкурентно мощные виды, способные захватывать и удерживать пространство (С-стратеги, по: Grime, 1977; виоленты, по: Раменский, 1938).
В соответствии с полимодельной концепцией Б.М. Миркина (Mirkin, 1994; Миркин, Наумова, 2012), растительные сообщества суровых местообитаний относятся к абиотической S-модели, благоприятных – к C-R-S-модели (доминируют C-стратеги, но среди сопутствующих видов преобладают R- и S-стратеги). Сделано предположение, что особенности организации фитоценозов этих типов могут проявиться в их структуре и функциональных связях, в том числе в степени выраженности доминантов и зависимости от них второстепенных видов и видового богатства (Миркин, Наумова, 2012). Однако знания об этом остаются неопределенными.
В частности известно, что на участках сообществ благоприятных местообитаний, где конкуренция достигает высокой интенсивности, наиболее успешные С-стратеги могут почти полностью монополизировать пространство и ресурсы (Huston, 1979; Peet, Christensen, 1988; Bengtsson, 1994; Миркин, Наумова, 2012). Кроме того, конкуренция за минеральные ресурсы считается преимущественно симметричной, а за свет – асимметричной (Peet, Christensen, 1988; Lamb, Cahill, 2008). Все это предполагает, что степень доминирования в сообществах благоприятных местообитаний должна быть в среднем выше, чем экстремальных. Однако полидоминантность высокопродуктивных древостоев тропических лесов противоречит таким представлениям. На их примере было показано, что интенсивная конкуренция за свет не ведет к значительной выраженности доминантов, если они сформированы экологически эквивалентными (конкурентно симметричными) видами (Bell, 2000; Hubbell, 1979, 2005; Zhang et al., 2015). Сделано предположение, что отсутствие явных доминантов в некоторых типах лугов имеет ту же природу (Василевич, 2014).
С другой стороны, сообщества экстремальных местообитаний также могут характеризоваться высокой степенью доминирования. Это связано с разной устойчивостью видов к воздействию стрессовых факторов (Magurran, 1988; Василевич, 1991; Чернов, 2005; Миркин, Наумова, 2012). Кроме того, в суровых условиях произрастает весьма небольшой набор видов, что автоматически ограничивает число тех из них, которые способны доминировать (Василевич, 1991). Однако дифференциация S-стратегов по степени толерантности к среде может быть снижена в результате случайных процессов. Возможно поэтому низкий уровень доминирования нередко встречается и в S-ценозах, как, например, в сообществах альпийских ковров и пустошей Западного Кавказа (Onipchenko et al., 1998).
Сообщества разных моделей могут отличаться степенью сходства структуры доминирования на соседних участках однородных местообитаний. Высокое сходство может означать пространственную упорядоченность (высокую организацию) растительных сообществ и, по мнению В.И. Василевича (1983), она чаще возникает в жестких условиях среды. Однако в соответствии с другой точкой зрения, если в биологическом сообществе отсутствуют сильные взаимодействия между видами и его структура определяется в основном влиянием факторов среды на отдельные популяции, то в пределах биотопа оно, скорее всего, будет представлять собой мозаику из весьма отличающихся друг от друга участков (Бурковский, Мазей, 2016).
Чем выше участие доминирующих видов в формировании фитоценозов, тем, обычно, ниже их видовое богатство (Wright, 1983; Wright et al., 1993; Palmer, Maarel, 1995; Maarel et al., 1995; Ernest, Brown, 2001; McKane et al., 2002). Связь этих параметров является одной из общепризнанных экологических закономерностей, однако в одних случаях она является тесной, а в других – слабой или отсутствует (Stirling, Wilsey, 2001; Aksenova et al., 2004; Cherednichenko, 2004; Houlahan, Findlay, 2004; Ma, 2005; Wilsey, Stirling, 2007; Lamb, Cahill, 2008; Sasaki, Lauenroth, 2011; Csergö et al., 2013; Bartha et al., 2014, и др.). Высказываются предположения, что это может быть связано со способом организации сообществ, но как – мнения расходятся.
Так, в ряде работ был сделан вывод, что сила влияния доминантов на видовое богатство зависит от степени выраженности у них черт С-стратегиии (Bobbink, Willems 1987; Grime, 1977, 2001; Somodi et al., 2008; Csergö et al., 2013). Поэтому логично было бы предположить, что связь между степенью доминирования и числом видов в сообществах благоприятных местообитаний должна быть более тесной, чем суровых. Кроме того, вторые обычно характеризуются малым размером видового пула. Причем это может быть связано не только с ограниченным количеством доступных ресурсов, но и с необходимостью формирования в процессе эволюции механизмов устойчивости к экстремальным явлениям, а также с преимущественно небольшим размером и изолированностью таких ценозов (Бигон и др., 1989; Qian, Ricklefs, 2004; Longino, Colwell, 2011). В результате можно ожидать, что многие из сообществ суровых местообитаний являются флористически неполночленными. Поэтому снижение в них участия доминантов и соответственно высвобождение ресурсов не обязательно приведет к ощутимому росту числа видов. Напротив, в сообществах, сформированных на благоприятных местообитаниях, потенциально способны произрастать многие виды (значительный видовой пул). Отсюда следует, что доминанты могут быть наиболее значимым, если не единственным фактором, определяющим их число (Миркин, Наумова, 2012; Bartha et al., 2014).
Однако в соответствии с альтернативной гипотезой, связь между структурой значимости видов и видовым богатством может быть более тесной в сообществах, где конкуренция выражена слабо или отсутствует (Stirling, Wilsey, 2001; Ma, 2005; Wilsey, Stirling, 2007; Sasaki, Lauenroth, 2011). По мнению Стирлинга и Вилси, значения обеих характеристик в таких ценозах определяются преимущественно скоростью иммиграции видов, и тесная связь между ними предсказывается нейтральными моделями (neutral diversity models) (Caswell, 1976; Bell, 2000; Stirling, Wilsey, 2001; Wilsey, Stirling, 2007). Напротив, в конкурентных сообществах эти характеристики определяются разными процессами. Видовое богатство более чувствительно к скорости иммиграции видов, доминирование – к интенсивности биотических взаимодействий. Поэтому изменения в участии видов в таких сообществах не обязательно должны сопровождаться изменениями их числа, а соответственно связь между этими характеристиками может оказаться слабой (Wilsey, Stirling, 2007). В частности, снижение численности доминирующих видов может вести к росту численности субдоминантов (т.е. к формированию олигодоминантной структуры) без последствий для видового богатства (Wilsey, Stirling, 2007; Munson, Lauenroth, 2009; Souza et al., 2011; Sasaki, Lauenroth, 2011).
Наконец, обратим внимание, что утверждение об отсутствии или низкой интенсивности конкуренции в сообществах растений экстремальных местообитаний нередко оспаривается, по крайней мере применительно к тем из них, которые имеют сомкнутый травостой (Goldberg, Novoplansky, 1997; Sammul et al., 2006; Онипченко, 2013). Так, в соответствии с гипотезой Фретвелла и Оксанена, интенсивность конкуренции является минимальной в сообществах с низкой, но не с самой низкой, продуктивностью (Fretwell, 1977; Oksanen et al., 1981; Sammul et al., 2000). Подтверждение этой точки зрения было получено, в частности, в ходе полевых экспериментов на нескольких растительных сообществах альпийского пояса Западного Кавказа (Onipchenko et al., 2009). Другие попытки оценить характер связи между продуктивностью сообществ и интенсивностью конкуренции методом экспериментов дали разные результаты: во многих случаях она действительно оказывалась положительной, но в других отсутствовала или даже была отрицательной (Sammul et al., 2000, 2006). По мнению Саммул с соавторами, одной из причин контрастных результатов могло быть воздействие на растительные сообщества выпаса животных, интенсивность которого коррелирует с первичной продукцией (Oksanen et al., 1981; Sammul et al., 2006). Кроме того, известно, что в сообществах экстремальных местообитаний широко распространены положительные взаимоотношения между видами (Миркин, Наумова, 2012; Онипченко, 2013), но какую роль они играют в структурировании ценозов и как это влияет на связь доминирования с видовым богатством остается не ясным.
В настоящей работе мы попытались оценить, как особенности организации растительных сообществ суровых и благоприятных местообитаний проявляются в их структуре доминирования, ее пространственной изменчивости на однородных местообитаниях, а также в характере ее связи с видовым богатством. При этом акцент был сделан на выявлении преобладающих (усредненных) тенденций в растительном покрове, для чего в качестве объектов исследования были использованы травяные сообщества широкого спектра местообитаний.
МАТЕРИАЛ И МЕТОДИКА
Объекты исследований
Объектами исследований явились травяные фитоценозы относительно стабильных местообитаний различных районов и высотных поясов (от 30 до 2800 м над ур. м.) Западного Кавказа и Предкавказья: альпийские ковры, пустоши и луга, субальпийские луга и болота, нижнегорные луга, травяной ярус ольховых, буковых и дубовых лесов, разные варианты степей. В соответствии с существующими представлениями, сообщества лугов относятся к конкурентной (C-S-R) модели (Работнов, 1983; Mirkin, 1994; Onipchenko et al., 1998; Миркин, Наумова, 2012). В частности, доминанты альпийских и субальпийских лугов характеризуются относительно большой биомассой, крупными семенами и относительно низкой скоростью роста, что соответствует С-стратегии; сопутствующие виды этих сообществ имеют черты C-, S- и R-стратегий (Onipchenko et al., 1998). Сообщества остальных типов сформированы в условиях более или менее значительного воздействия стрессовых факторов. Так, ценозы альпийских ковров приурочены к долгоснежным местообитаниям, и поэтому имеют короткий период вегетации; альпийских пустошей – к малоснежным местообитаниям (вершинам и гребням хребтов), характеризующимся низкими температурами почвы в зимний период; сообщества субальпийских болот сформированы в условиях избытка воды; степей – ее недостатка; травяного яруса лесов – недостатка света или, если это сообщества лесных эфемероидов, то в условиях короткого периода вегетации. Все они могут быть отнесены к S-модели или переходному типу (Grime, 1977, 2001; Работнов, 1983; Onipchenko et al., 1998; Миркин, Наумова, 2012).
Методы сбора фактического материала
В основу работы положены 595 проб надземной биомассы, отобранных на площадках 0.25 м2 в течение полевых сезонов 2014–2017 гг. Пробы были взяты на типичных участках сообществ без признаков зоогенных и антропогенных нарушений в период максимального развития травостоя. Часть из них была отобрана регулярным способом в виде трансект, включающих 10 площадок по 0.25 м2; другие – сериями по 3–6 площадок на участок. Во втором случае выбирали варианты сообществ с наиболее высоким, наиболее низким и средним проективным покрытием доминирующих видов, которое оценивали визуально. Пробы разбирали по видам и взвешивали. Затем одну–три наиболее типичные пробы из серии высушивали и взвешивали. Сухую биомассу для остальных проб серии определяли на основе значений коэффициента усушки. Небольшой размер площадок для отбора проб (0.5 × 0.5 м) означает, что полученные нами результаты имеют отношение к локальному масштабу.
Методы анализа
В качестве показателя пригодности среды для произрастания растений рассматривали их надземную сухую биомассу на 0.25 м2 в период максимального развития травостоя (W). Для сообществ, не испытывающих существенного воздействия фитофагов, данную характеристику нередко используют как суррогат их годичной надземной продукции. Поскольку границы между суровыми и благоприятными местообитаниями являются условными, особенности структуры доминирования и ее связи с видовым богатством анализировали на градиенте их продуктивности (запаса надземной биомассы травостоя). Причем мы рассматривали только ту часть градиента, в пределах которой наблюдался рост видового богатства сообществ – до 200 г сухого вещества на 0.25 м2 (Акатов, Акатова, 2016). Можно ожидать, что местообитания с более высокой биомассой фитоценозов еще более благоприятны для произрастания растений. Однако, как показывают результаты многих исследований, ценозы с наиболее высокой биомассой чаще характеризуются относительно низким видовым богатством (Бигон и др., 1989; Adler et al., 2011; Šímová et al., 2013; Василевич, 2015, и др.). Причем это может быть результатом не только интенсивной конкуренции за свет, но также небольшого числа особей растений на площадках из-за их крупного размера (Бигон и др., 1989; Oksanen, 1996; Adler et al., 2011; Šímová et al., 2013; Василевич, 2015, и др.). Поэтому низкое видовое богатство в ценозах с высокой продукцией некоторыми авторами считается артефактом (Oksanen, 1996).
Под структурой доминирования можно понимать соотношение относительного участия (плотности, биомассы) видов первых двух–трех рангов, которая может быть оценена посредством как D1, D2, D3, так и K1 (=D1), K2 и K3. В нашем случае показатели D отражают соотношение между влажной биомассой вида того или иного ранга и общей влажной биомассой травостоя на 0.25 м2. Величина D1 рассматривается как степень доминирования (индекс Бергера–Паркера: Berger, Parker, 1970; Magurran, 1988), а разница между D1 и D2 может отражать общую структуру доминирования в сообществах. Строгих границ для определения ее типов не существует, однако условно, если эта разница превышает 50%, то доминанты называют абсолютными (Работнов, 1983), а сообщества можно рассматривать как сверхдоминантные, если она составляет 15–50% – монодоминантные, менее 15% – олиго- или полидоминантные.
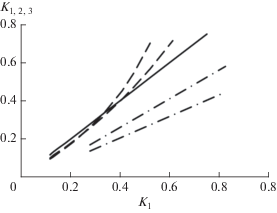
Величина К показывает, какую часть оставшихся ресурсов сообщества использует каждый вид в порядке снижения их участия в сообществе (увеличения ранга): K1 (=D1) = W1/WR, K2= W2/(WR – W1), K3 = W3/( WR – W1– W2), где W1, W2 и W3 влажная биомасса видов первого–третьего ранга на 0.25 м2. Теоретически возможны разные варианты соотношения между K1 (=D1), K2 и K3 (рис. 1). Так, на небольших участках ценозов или отдельных их компонентов (например, ярусов) последовательность оценок биомассы (численности) видов нередко хорошо соответствует геометрической прогрессии (модель Мотомуры) (Уиттекер, 1980; Magurran, 1988; Panchal, Pandey, 2004; Caruso et al., 2007; Кузнецова, 2009; Zhang et al., 2012). Это означает, что виды всех рангов, в том числе первых, характеризуются примерно равными значениями К. Другие типы (модели) структуры участия видов предполагают как более, так и менее выраженное, чем в геометрической модели, доминирование одного из видов (Уиттекер, 1980; Левич, 1980; Magurran, 1988; Лебедева, Криволуцкий, 2002; Ferreira, Petrere-Jr., 2008, и др.). К первым, например, относится гиперболическая модель, предложенная А.П. Левичем (1980). В соответствии с ней величина K убывает с увеличением ранга видов (рис. 1). Ко вторым – модель Макартура (Уиттекер, 1980; Magurran, 1988; Шитиков и др., 2011). В соответствии с этой моделью значения K1 (=D1) могут быть как выше, так и ниже, чем K2 и K3 (рис. 1).
Рис. 1.
Ожидаемые (расчетные) соотношения между значениями K для первого по рангу вида (K1) и двух видов последующих рангов (K2 и K3) в сообществах с ранговой структурой обилия видов, соответствующей разным моделям. Геометрическая модель – сплошная линия (K1, K2 и K3); гиперболическая модель – сплошная (K1) и пунктирные с точкой (K1, K2 и K3) линии; модель Макартура – сплошная (K1) и пунктирные (K1, K2 и K3) линии. Для расчетов использовались уравнения, представленные в работах: Уиттекер, 1980; Кузьминов, 2009. Значения параметров моделей задавались произвольно.
Пространственную изменчивость структуры доминирования оценивали на трансектах, состоящих из 10 площадок, путем расчета четырех показателей: DF ', а также коэффициента вариации (CV) значений D1, (D1 – D2) и (K1 – K2). Величина DF ' = DF – DFs, где DF – доля площадок, на которых наиболее значимый для трансекты в целом вид имел первый ранг (“частота доминирования” Де Фриза: Баканов, 1987), DFs – значение DF в случае, если ранг видов определяется только случайными процессами (вероятностная характеристика для площадок с определенным числом видов, полученная с использованием формулы Бернулли).
Анализ влияния запаса биомассы на тесноту связи между степенью доминирования и видовым богатством включал два этапа. На первом мы оценили тесноту такой связи для 23 групп сообществ с доминированием определенного вида(-ов). С этой целью был использован коэффициент корреляции рангов Спирмена (RSD). Он малочувствителен к сильно отличающимся значениям характеристик и позволяет измерять степень сопряженности между признаками независимо от закона распределения и формы связи. На втором этапе мы определили характер и тесноту связей между RSD, средней для групп продукцией сообществ (W) и другими характеристиками.
Тип связи между характеристиками определяли путем построения линейных и квадратичных моделей регрессии. Предполагалось, что статистическая значимость только линейных коэффициентов регрессии свидетельствует о линейном характере связи, линейных и квадратичных или только квадратичных – о криволинейном (проверяли на основе t-критерия Стьюдента). Если связь не являлась линейной, то при необходимости производили ее линеаризацию путем логарифмирования значений независимого параметра, а затем повторно тестировали на линейность. Тесноту связи между характеристиками определяли путем расчета нескорректированного коэффициента детерминации (для оценки его статистической значимости использовался F-критерий Фишера), парного коэффициента корреляции Пирсона и частного коэффициента корреляции. При сравнении регрессионных моделей с разным числом параметров использовали скорректированный коэффициент детерминации ($R_{{adj}}^{2}$). В некоторых случаях влияние факторов на изучаемую характеристику оценивали методом однофакторного дисперсионного анализа (ANOVA). Расчеты проводили с использованием программ Microsoft Excel 2007 и Statistica 6.0.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Надземная биомасса сообществ
Данные по запасам надземной биомассы сообществ предположительно S- и C-S-R-моделей, а также сообществ с доминированием определенных видов или их групп, представлены в табл. 1. Наибольшим ее запасом характеризуются субальпийские и низкогорные луга, а также степи с доминированием Stipa pulcherrima. Меньшим – сообщества сухих степей, субальпийских болот, альпийских пустошей и ковров; наименьшими значениями этой характеристики – сообщества травяного яруса лесов (S-модель). Среди предположительно конкурентных сообществ (C-S-R-модель) в среднем наиболее низкой биомассой характеризуются альпийские луга с доминированием Kobresia macrolepis. Как видно из табл. 1, участки с низкой биомассой (менее 50 г сухого вещества на 0.25 м2) встречаются также в пределах луговых сообществ с доминированием Alchemilla retinervis, Geranium gymnocaulon и Botriochloa ischaemum. Местообитания таких C-S-R-ценозов также можно рассматривать как суровые.
Таблица 1.
Характеристика изученных растительных сообществ
| Сообщества, доминанты | n | W | S | D1(=K1) | (D1 – D2) | (K1 – K2) | RSD |
|---|---|---|---|---|---|---|---|
| Альпийские луга (в целом) | 62 | 73.3/22.4/171.4 | 10.2/2.0/24.0 | 0.74/0.25/0.99 | 0.63/0.02/0.99 | 0.23/–0.16/0.70 | – |
| Alchemilla retinervis Buser | 14 | 101.5/63.7/39.4 | 7.4/4.0/11.0 | 0.83/0.59/0.97 | 0.72/0.31/0.95 | 0.21/–0.09/0.59 | –0.677 |
| Geranium gymnocaulon DC | 22 | 76.2/39.7/171.4 | 14.7/9.0/24.0 | 0.57/0.25/0.86 | 0.43/0.02/0.81 | 0.18/–0.16/0.50 | –0.766 |
| Kobresia macrolepis Meinsh. | 21 | 39.7/22.4/73.4 | 7.5/4.0/12.0 | 0.79/0.61/0.95 | 0.68/0.46/0.91 | 0.22/–0.08/0.53 | –0.695 |
| Субальпийские луга (в целом) | 59 | 107.7/46.4/173.2 | 17.2/6.0/35.0 | 0.32/0.19/0.96 | 0.36/0.00/0.95 | 0.16/–0.15/0.67 | – |
| Brachypodium rupestre (Host) Roem. et Schult. | 20 | 100.4/69.7/160.9 | 20.5/6.0/35.0 | 0.50/0.26/0.92 | 0.37/0.03/0.90 | 0.21/–0.06/0.67 | –0.687 |
| Calamagrostis arundinacea (L.) Roth | 16 | 107.8/46.4/163.6 | 14.4/6.0/27.0 | 0.61/0.31/0.90 | 0.44/0.04/0.87 | 0.17/–0.15/0.61 | –0.831 |
| Нижнегорные луга (в целом) | 106 | 97.7/22.2/186.2 | 12.1/3.0/23.0 | 0.59/0.17/0.94 | 0.43/0.01/0.92 | 0.18/–0.31/0.60 | – |
| Botriochloa ischaemum (L.) Keng | 23 | 88.7/38.3/149.4 | 12.8/6.0/19.0 | 0.60/0.26/0.93 | 0.42/0.06/0.90 | 0.12/–0.12/0.44 | –0.705 |
| Brachypodium pinnatum (L.) P.Beauv. | 20 | 100.7/56.8/142.7 | 13.2/8.0/21.0 | 0.69/0.24/0.88 | 0.54/0.02/0.81 | 0.26/–0.05/0.55 | –0.729 |
| Calamagrostis epigeios (L.) Roth | 22 | 120.3/54.9/186.8 | 7.5/3.0/15.0 | 0.71/0.34/0.94 | 0.57/0.02/0.92 | 0.17/–0.31/0.48 | –0.661 |
| Geranium sanguineum L. | 13 | 130.6/95.4/163.6 | 22.9/18.0/29.0 | 0.60/0.50/0.67 | 0.49/0.38/0.58 | 0.33/0.18/0.44 | –0.694 |
| Альпийские пустоши и ковры (в целом) | 38 | 38.0/8.2/96.8 | 14.2/6.0/28.0 | 0.41/0.19/0.73 | 0.21/0.00/0.64 | 0.06/–0.09/0.39 | – |
| Carex tristis M.Bieb. и др. | 18 | 47.3/20.8/96.8 | 14.8/9.0/23.0 | 0.38/0.22/0.63 | 0.18/0.01/0.43 | 0.03/–0.08/0.24 | –0.454 |
| Taraxacum stevenii DC. и др. | 20 | 31.1/8.2/69.6 | 12.9/6.0/22.0 | 0.47/0.24/0.88 | 0.23/0.01/0.64 | 0.07–0.09/0.39 | –0.568 |
| Субальпийские болота (в целом) | 40 | 43.5/18.2/95.6 | 8.1/2.0/6.0 | 0.50/0.24/0.89 | 0.24/0.01/0.83 | 0.00/–0.27/0.80 | – |
| Carex rostrata Stokes | 23 | 34.4/18.2/62.1 | 6.5/2.0/16.0 | 0.49/0.24/0.89 | 0.23/0.01/0.83 | –0.01/–0.21/0.43 | –0.482 |
| Степи (в целом) | 93 | 67.1/11.3/139.5 | 13.4/6.0/32.0 | 0.48/0.16/0.48 | 0.30/0.61/0.97 | 0.09/–0.24/0.47 | – |
| Stipa pulcherrima C.Koch | 16 | 103.0/70.4/168.8 | 20.5/14.0/32.0 | 0.41/0.16/0.72 | 0.33/0.02/0.64 | 0.16/–0.11/0.31 | –0.491 |
| Agropyron pinifolium Nevski | 16 | 77.1/54.9/105.3 | 11.2/8.0/14.0 | 0.44/0.20/0.71 | 0.28/0.01/0.61 | 0.12/–0.08/0.40 | –0.474 |
| Stipa lessingiana Trin. et Rupr. | 10 | 37.1/16.0/68.9 | 9.1/7.0/11.0 | 0.48/0.26/0.81 | 0.30/0.09/0.72 | 0.06/–0.15/0.34 | –0.3 |
| Artemisia lercheana Weber ex Stechm. | 23 | 50.2/14.3/75.3 | 8.0/6.0/9.0 | 0.66/0.28/0.98 | 0.47/0.03/0.97 | 0.08/–0.37/0.30 | 0 |
| Травяной ярус лесов (в целом) | 166 | 26.9/3.8/84.8 | 6.3/3.0/19.0 | 0.65/0.25/0.98 | 0.46/–0.02/0.98 | 0.05/–0.48/0.72 | – |
| Aegopodium podagraria L. | 18 | 26.5/7.9/40.1 | 5.2/3.0/7.0 | 0.66/0.44/0.88 | 0.41/–0.03/0.82 | 0.02/–0.46/0.41 | 0.289 |
| Allium ursinum L. | 19 | 29.0/11.8/42.6 | 6.1/4.0/8.0 | 0.68/0.34/0.89 | 0.50/0.02/0.82 | 0.09/–0.37/0.39 | –0.444 |
| Dryopteris filix-mas (L.) Schott | 22 | 26.8/8.2/54.3 | 4.2/3.0/6.0 | 0.73/0.43/0.98 | 0.53/0.00/0.97 | 0.00/–0.48/0.91 | 0.123 |
| Festuca montana M.Bieb. | 28 | 13.8/3.8/53.5 | 4.6/3.0/8.0 | 0.77/0.49/0.98 | 0.65/0.10/0.98 | 0.16/–0.26/0.72 | –0.347 |
| Symphytum grandiflorum DC. | 20 | 18.8/8.1/28.7 | 3.6/2.0/6.0 | 0.74/0.42/0.99 | 0.54/0.08/0.99 | 0.05/–0.34/0.99 | –0.423 |
| Trifolium medium L. | 15 | 26.3/15.5/34.6 | 13.1/10.0/18.0 | 0.46/0.31/0.67 | 0.25/0.02/0.50 | 0.08/–0.11/0.23 | –0.042 |
| Лесные эфемероиды (в целом) | 30 | 8.8/4.5/14.8 | 8.0/6.0/11.0 | 0.33/0.21/0.58 | 0.09/0.00/0.48 | –0.03/–0.17/0.31 | – |
| Corydalis marschalliana (Pallas exWilld.) Pers. | 20 | 7.1/2.8/11.5 | 8.0/6.0/12.0 | 0.34/0.21/0.59 | 0.10/0.00/0.48 | –0.03/–0.20/0.31 | 0.138 |
Примечание. n – число проб; W – сухая надземная биомасса (г/0.25 м2); S – число видов растений на 0.25 м2; D1– степень доминирования (индекс Бергера–Паркера); (D1 – D2) и (K1 – K2) – показатели структуры доминирования (цифры в поле таблицы – средние/минимальные/максимальные значения параметров). RSD – теснота связи между степенью доминирования и видовым богатством в растительных сообществах с доминированием определенных видов (коэффициент корреляции рангов Спирмена; полужирным выделены значения, статистически значимые на уровне Р < 0.05).
Структура доминирования на площадках
Соотношение между средним запасом надземной биомассы сообществ с доминированием определенных видов и средними значениями характеристик D1, D2, D3, а также (D1 – D2), показано на рис. 2а, б и в табл. 2. Из них следует, что только величина D2 статистически значимо связана с биомассой: чем выше средний для сообществ запас биомассы, тем ниже среднее относительное участие в их сложении видов второго ранга. Значения других параметров, характеризующих структуру доминирования сообществ, варьируют в широких пределах и практически не связаны с продуктивностью местообитаний. В частности, в среднем более высокой степенью доминирования (D1) характеризуются как относительно продуктивные луговые ценозы, так и низкопродуктивные сообщества травяного яруса лесов; менее высокой – низкопродуктивные нелесные сообщества, наиболее низкой – сообщества эфемероидов широколиственных лесов. При этом на отдельных участках биомасса доминирующих видов в сообществах лугов, субальпийских болот, степей и травяного яруса лесов может составлять более 70–90% от общей биомассы травостоя, в сообществах альпийских лугов и пустошей их доля не превышает 70%, а лесных эфемероидов – 50% (табл. 1).
Рис. 2.
Соотношения между средней надземной биомассой сообществ с доминированием определенного вида (W) и средними значениями параметров, характеризующих их структуру доминирования: а – D1, D2, D3; б – (D1– D2); в – K1 (=D1), K2, K3; г – (K1 – K2). На рис. 2а и в черные кружки – относительное участие видов первого ранга (D1 = K1), белые квадраты – второго (D2, K2), черные квадраты – третьего (D3, K3).
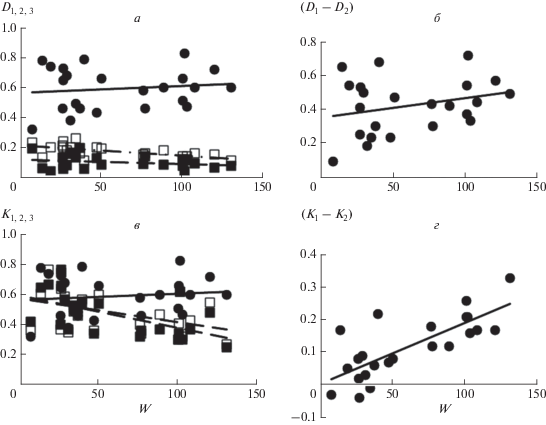
Таблица 2.
Соотношение между средними значениями параметров, характеризующих растительные сообщества с доминированием определенных видов
| Переменные | n | Модель | R2 | P | |
|---|---|---|---|---|---|
| y | x | ||||
| D1(=K1) | W | 23 | y = –0.0004x + 0.565 | 0.014 | NS |
| D2 | W | 23 | y = –0.00073x + 0.214 | 0.403 | < 0.05 |
| D3 | W | 23 | y = –0.0003x + 0.115 | 0.067 | NS |
| K2 | W | 23 | y = –0.0016x + 0.575 | 0.178 | < 0.05 |
| K3 | W | 23 | y = –0.0022x + 0.597 | 0.338 | < 0.01 |
| (D1 – D2) | W | 23 | y = 0.0011x + 0.351 | 0.072 | NS |
| (K1 – K2) | W | 23 | y = 0.0019x + 0.004 | 0.580 | < 0.01 |
| Smax | W | 23 | y = 0.0019x + 0.004 | 0.440 | < 0.01 |
| RSD | W | 23 | y = 0.1472x + 0.082 | 0.473 | < 0.01 |
| RSD | (K1 – K2) | 23 | y = 6.4191x2 – 4.083x – 0.094 | 0.565 | < 0.01 |
| RSD | Smax | 23 | y = 0.0012x2 – 0.0676x + 0.278 | 0.389 | < 0.01 |
Примечание. n – число сообществ; здесь и в табл. 3: R2– нескорректированный коэффициент детерминации (P – уровень статистической значимости, NS – значение не значимо); для табл. 2 и 3 –полужирным выделены значения линейного коэффициента регрессии, значимые на уровне Р < 0.05.
Величина (D1 – D2) несколько выше в ценозах с более высокой надземной биомассой, однако данная связь статистически не значима (рис. 2б, табл. 2). В среднем сверхдоминантная структура (разница между D1 и D2 превышает 0.5) чаще встречается в сообществах альпийских лугов с относительно низкой биомассой, чем субальпийских и нижнегорных – с относительно высокой. В травяном ярусе лесов часто встречается как доминантная, так и сверхдоминантная структура; в сообществах степей, субальпийских болот, альпийских пустошей и ковров – преимущественно доминантная; лесных эфемероидов – олигодоминантная.
На рис. 2в, г и в табл. 2 показано соотношение между средней надземной биомассой сообществ с доминированием определенных видов и средними значениями величин K1 (=D1), K2, K3 и (K1 – K2). Из них следует, что в сообществах с относительно высокой биомассой (луга и степи с доминированием Stipa pulcherrima и Agropyron pinifolium) значения K для видов 1-го ранга существенно выше, чем для менее значимых видов. По мере роста суровости местообитаний эта разница сокращается, и в сообществах с биомассой менее 50 г/0.25 м2 (суровые местообитания) значения K1, K2 и K3 сильно варьируют, но в среднем становятся близкими. При этом в низкопродуктивных луговых ценозах с доминированием Kobresia macrolepis (альпийский пояс) среднее значение (K1 – K2) остается высоким (табл. 1).
На рис. 3 и 4 и в табл. 3 показаны соотношения между надземной биомассой сообществ и значениями D1 (=K1), (D1 – D2) и (K1 – K2) для отдельных площадок. В основном они подтверждают выводы, сделанные выше на основе усредненных данных для сообществ с доминированием определенных видов. Так, значения параметров D1 (=K1) и (D1 – D2) варьируют в широких пределах и не связаны с биомассой (рис. 3а, б), а параметра (K1 – K2) – растут с увеличением запаса биомассы (рис. 4а). Кроме того, в сообществах лугов и степей с относительно высоким запасом биомассы (с доминированием Stipa pulcherrima, Agropyron pinifolium и др.) значения (K1 – K2) в среднем выше, чем в остальных сообществах (рис. 4б, в), и это различие является статистически значимым: ANOVA, n1 = 281, n2 = 314, F = 64.1, Р < 0.01. Причем в пределах сообществ этих групп значения (K1 – K2) не связаны с биомассой (табл. 3).
Рис. 3.
Соотношения между надземной биомассой сообществ на отдельных площадках (W) и значениями параметров: а – D1(=K1), б – (D1– D2).
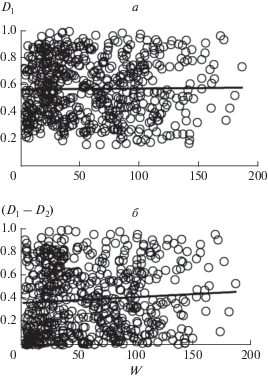
Рис. 4.
Соотношения между надземной биомассой сообществ на отдельных площадках (W) и значениями (K1– K2). а – построены на основе всех данных, б – на основе данных по сообществам лугов и наиболее продуктивных участков степей, в – на основе данных по другим типам сообществ.
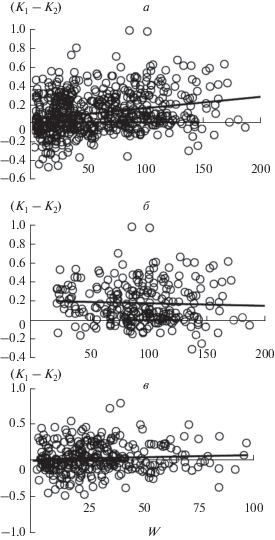
Таблица 3.
Соотношение между запасом надземной биомассы и показателями структуры доминирования в изученных растительных сообществах
| Зависимая переменная | № | n | Модель | R2 | P |
|---|---|---|---|---|---|
| D1 (=K1) | 1 | 595 | y = 0.000x + 0.567 | 0.000 | NS |
| (D1 – D2) | 2 | 595 | y = 0.0005x + 0.359 | 0.006 | NS |
| (K1 – K2) | 3 | 595 | y = 0.0012x + 0.032 | 0.055 | < 0.01 |
| (K1 – K2) | 4 | 228 | y = –0.0003x + 0.203 | 0.002 | NS |
| (K1 – K2) | 5 | 367 | y = 0.0006x + 0.023 | 0.003 | < 0.05 |
Отсутствие существенной связи между продуктивностью, а соответственно способом организации сообществ, и значениями D1 и (D1 – D2) не является неожиданностью. Похожие выводы были сделаны и другими авторами (Vermeer, Verhoeven, 1987; Chalcraft et al., 2009; Василевич, 2015). Можно предположить, что это связано с большим числом факторов, разнонаправленно действующих на сообщества суровых и благоприятных местообитаний (см. выше). Более интересным нам представляется значительное превышение в среднем величины K1 (=D1) над K2 и K3 в сообществах некоторых типов. Оно предсказывается гиперболической моделью (рис. 1), в основе которой лежат положительные обратные связи второго порядка. Это может означать, например, что доминант оказывается более многочисленным по сравнению с другими видами не только из-за его более высокой конкурентоспособности, но и потому, что его высокая численность (биомасса, покрытие) сама по себе способствует росту численности (Василевич, 1991), а соответственно и более сильному его воздействию на среду обитания (например, путем накопления слоя ветоши, изменения светового режима, физико-химических свойств почвы, аллелопатии и т.д.: Работнов, 1983; Grime, 2001; Callaway, Ridenour, 2004; Reinhart et al., 2005; Hulme, Bremner, 2006; Somodi et al., 2008; Csergő et al., 2013; Bartha et al., 2014), что в свою очередь еще более усиливает его конкурентную мощность.
Поскольку способность воздействовать на экотоп имеется преимущественно у С-стратегов, величину (K1 – K2) предположительно можно рассматривать в качестве ценотического показателя степени выраженности у доминантов этой стратегии, а значит, и в качестве критерия при отнесении растительных сообществ к С-R-S- или S-модели. В пользу этого предположения свидетельствует также отсутствие связи между значениями этой величины и запасом биомассы в пределах групп сообществ, организованных предположительно сходным способом (рис. 4б, в). Это подчеркивает, что величина (K1 – K2) определяется преимущественно биоэкологическими особенностями доминирующих видов, в то время как надземная биомасса травостоев сама по себе, а соответственно их высота и густота не оказывают на нее существенного влияния. При этом подчеркнем, что величина (K1 – K2) является статистической, т.е. может быть определена только на основе выборочных данных.
Если использование величины (K1 – K2) в качестве критерия при определении модели организации сообществ является правомерным, то наши данные позволяют уточнить представление об экологических ареалах S- и C-S-R-ценозов. Так, в наиболее суровых местообитаниях (сухая надземная биомасса менее 20 г на 0.25 м2) нами были обнаружены преимущественно S-ценозы, на градиенте биомассы от 20 до 100 г (субэкстремальные и субблагоприятные местообитания) – как S-, так и C-S-R-сообщества, более 100 г – только C-S-R-ценозы (рис. 4б, в). Результатом такого перекрытия ареалов является положительная корреляция между запасом надземной биомассы сообществ и значениями (K1 – K2) (рис. 2г, 4а, табл. 2, 3).
Структура доминирования: пространственная изменчивость
На рис. 5а и в табл. 4 показано соотношение между средней надземной биомассой сообществ (W) и долей площадок с доминированием наиболее значимых на трансектах видов (использовали скорректированные значения данной характеристики – DF '). Как следует из табл. 4, формально статистически значимая связь между W и DF ' отсутствует. При этом на рис. 5а хорошо видно, что в сообществах со средней продукцией более 100 г/0.25 м2 значения этой характеристики высокие, а менее – варьируют в широких пределах. Из рис. 5б–г видно, что участки сообществ с низким запасом биомассы характеризуются несколько более высоким уровнем варьирования величин D1, (D1 – D2) и (K1 – K2), чем ценозы с относительно высоким ее запасом, однако статистически значимая связь наблюдается только между W и варьированием (K1 – K2) (табл. 4). Из табл. 4 также следует, что величина DF ' в сообществах лугов и наиболее продуктивных степей (преимущественно C-S-R- и переходные модели) статистически значимо выше, а уровень варьирования величин (D1 – D2) и (K1 – K2) ниже, чем в сообществах других типов (S-модель). Это может означать, что доминанты С-стратеги лучше удерживают лидирующую позицию на каждом участке однородного местообитания, чем S-стратеги. А также, что дифференциация видов по степени толерантности к среде обуславливает формирование менее упорядоченной (предсказуемой) пространственной структуры доминирования, чем по степени их конкурентоспособности за ресурсы.
Рис. 5.
Соотношение между средними значениями надземной биомассы (W) и показателями степени варьирования структуры доминирования для участков сообществ, заложенных в виде трансект в пределах однородных местообитаний. Показатели, характеризующие структуру доминирования сообществ: а – D1(=K1); б – (D1– D2); в – (K1– K2); г – скорректированное значение DF ' (доля площадок на трансекте с доминированием наиболее значимого вида). В качестве показателя степени варьирования значений D1(=K1), (D1 – D2) и (K1 – K2) использовался коэффициент вариации (CV).
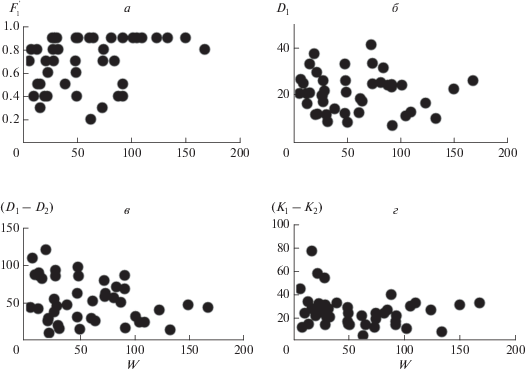
Таблица 4.
Результаты анализа пространственной изменчивости структуры доминирования на участках (трансектах) однородных местообитаний методом однофакторного дисперсионного анализа (ANOVA)
| Параметры | W, г/0.25 м2 | Модель организации | |||||
|---|---|---|---|---|---|---|---|
| < 29 | 29–73.5 | > 73.5 | F | 1 | 2 | F | |
| n =15 | n =14 | n =14 | n =23 | n = 20 | |||
| CV/D1(=K1) | 22.3 | 19.3 | 20.8 | 0.48 | 21.1 | 21.5 | 0.07 |
| CV/(D1 – D2) | 66.9 | 47.9 | 46.0 | 2.50 | 62 | 44.5 | 4.13 |
| CV/(K1 – K2) | 33.1 | 19.6 | 24.4 | 4.22 | 30.3 | 20.7 | 5.92 |
| $F_{1}^{'}$ | 0.64 | 0.74 | 0.71 | 1.14 | 0.63 | 0.77 | 5.14 |
Примечание. n – число трансект; W – сухая надземная биомасса (г/0.25 м2); F – фактические значения критерия Фишера (полужирным выделены значения, статистически значимые на уровне Р < 0.05); модели организации: 1 – S-ценозы, 2 – преимущественно C-S-R- и переходные ценозы; в качестве показателя степени варьирования значений D1(=K1), (D1 – D2) и (K1 – K2) использовался коэффициент вариации (CV); $F_{1}^{'}$ – доля площадок на трансекте с доминированием наиболее значимого вида (скорректированное значение).
Связь между степенью доминирования и видовым богатством
Как следует из рис. 6а и табл. 2 и 3, теснота связи между степенью доминирования (D1) и видовым богатством (S) не одинакова в сообществах с доминированием разных видов и положительно скоррелирована с величиной их биомассы (W). Кроме того, обратим внимание, что сообщества с биомассой более 50 г сухого вещества на 0.25 м2 характеризуются преимущественно высокой теснотой этой связи, с менее 50 г/0.25 м2 – значительным варьированием RSD. Такой характер соотношения между W и RSD может быть результатом разных процессов. Так, можно ожидать, что чем выше продуктивность сообществ, тем 1) выше интенсивность конкуренции в целом и соответственно вероятность конкурентного исключения видов (так, по данным Саммул с соавторами, именно 50 г/0.25 м2 является пороговым значением данного параметра, ниже которого интенсивность конкуренции близка к нулю: Sammul et al., 2006); 2) больше размер видового пула сообществ, максимально возможное видовое богатство (Smax) и соответственно выше потенциальная роль доминантов в ограничении числа видов; 3) сильнее выражены черты С-стратегии у доминантов. В случае важности второго и третьего механизмов значения Smax и (K1 – K2) должны коррелировать как с W, так и с RSD, причем связь между ними и RSD должна быть более тесной, чем между W и RSD. Ситуация, когда, напротив, более тесной является связь между W и RSD, может свидетельствовать в пользу важности первого механизма.
Рис. 6.
Соотношения между средними значениями параметров W, Smax и (K1 – K2) в сообществах с доминированием определенного вида и теснотой связей в них между степенью доминирования и видовым богатством (оценивалась на основе коэффициента корреляции рангов Спирмена, RSD). а – между W и RSD; б – Smax и RSD; в – (K1– K2) и RSD.
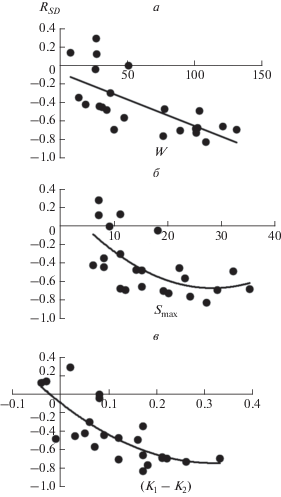
Из табл. 2 видно, что между средней надземной биомассой (W) и максимальным видовым богатством (Smax) растительных сообществ наблюдается линейная положительная статистически значимая связь. Из рис. 6б и табл. 2 следует, что между Smax и RSD имеется статистически значимая криволинейная монотонно убывающая связь, которая, однако, является менее тесной, чем между W и RSD (в первом случае $R_{{adj}}^{2}$ = 0.328, во втором $R_{{adj}}^{2}$ = 0.448). После логарифмирования значений Smax мы повторно проверили наличие в ней нелинейной составляющей путем включения в линейное уравнение регрессии квадратичного компонента, однако он оказался статистически незначимым. Частный коэффициент корреляции между lnW и RSD (т.е. при постоянных значениях Smax) является статистически значимым (r = −0.508, n = 23, P < 0.05), а между lnSmax и RSD (при постоянных значениях W) – не значимым (r = −0.280, n = 23).
Как было показано выше, между средней биомассой (W) и величиной (K1 – K2) наблюдается линейная положительная статистически значимая связь (рис. 2г, табл. 2). Из рис. 6в и табл. 2 следует, что между (K1 – K2) и RSD наблюдается статистически значимая криволинейная монотонно убывающая связь, которая при этом является более тесной, чем между W и RSD (в первом случае $R_{{adj}}^{2}$ = 0.522, во втором $R_{{adj}}^{2}$ = 0.448). Частный коэффициент корреляции между ln(K1 – K2) и RSD (при постоянных значениях W) является статистически значимым (r = −0.492, n = 23, P < 0.05) и более высоким, чем между lnW и RSD (при постоянных значениях разницы между K1 и K2): r = −0.366, n = 23, P < 0.1.
Таким образом, мы не обнаружили значительного вклада Smax в объяснение вариации RSD, независимого от вклада W. Однако влияние (K1 – K2) на RSD можно рассматривать как существенное и в определенной степени не зависимое от W. Это может означать, что связь между W и RSD определяется как общей интенсивностью межвидовых взаимодействий, так и степенью выраженности черт С-стратегии у доминирующих видов.
ЗАКЛЮЧЕНИЕ
Итак, считается, что растительные сообщества суровых и благоприятных местообитаний организованы разным способом. Первые – преимущественно абиотической средой и случайными процессами (S-модель организации), вторые – преимущественно конкуренцией (C-S-R-модель). Можно ожидать, что структурирующие возможности этих факторов не одинаковы и это каким-то образом проявляется в структуре ценозов и характере функциональных связей. Целью нашего исследования было проверить это предположение на примере структуры доминирования и характера ее связи с видовым богатством. При этом акцент был сделан на выявлении преобладающих (усредненных) тенденций в растительном покрове, для чего были изучены травяные сообщества широкого спектра местообитаний. В качестве критерия степени благоприятности (суровости) среды использовали надземную биомассу травостоя в период его максимального развития. Относительное участие видов в травостое оценивали как отношение их биомассы к общей биомассе травостоя на площадке (D) или к биомассе травостоя, оставшейся после удаления из нее биомассы видов более высокого ранга (K). Под структурой доминирования понимали относительное участие в травостое вида первого ранга (степень доминирования), а также соотношение значений D и K для двух-трех видов с наибольшим участием в травостое.
Полученные нами результаты можно свести к следующему:
1. Если структуру доминирования оценивать на основе величины D, то она варьирует в широких пределах и почти не связана с продуктивностью местообитаний. Более определенным является варьирование соотношения между значениями величины K для видов первого и второго рангов. В C-S-R-ценозах, независимо от запаса их биомассы, значения K1 в среднем существенно выше, чем K2; в S-ценозах независимо от их биомассы – они в среднем примерно одинаковы.
2. Сделано предположение, что величину (K1 – K2) можно рассматривать в качестве ценотического показателя выраженности у доминантов черт С-стратегии, а значит, и в качестве критерия при определении модели организации растительных сообществ. Использование данного подхода позволило уточнить ареалы S- и C-S-R-ценозов на градиенте продуктивности местообитаний и сделать вывод, что они существенно перекрываются.
3. Сообщества суровых местообитаний (преимущественно S-модель) характеризуются более высокой пространственной изменчивостью структуры доминирования, чем благоприятных (преимущественно C-S-R- и переходные модели).
4. Сообщества C-S-R-модели организации характеризуются более тесной связью между степенью доминирования и видовым богатством, чем S-ценозы. Наши результаты позволяют предположить, что это связано с разной интенсивностью межвидовой конкуренцией и жизненной стратегией доминирующих видов.
Таким образом, результаты показали, что особенности организации растительных сообществ суровых и благоприятных местообитаний проявляются в ряде аспектов структуры доминирования и характере ее связи с видовым богатством. При этом, если в качестве критерия высокой организованности ценозов рассматривать наличие устойчивых отношений между их элементами, а также их низкую вариабельность в относительно стабильных условиях среды (Василевич, 1983; Azovsky, 2009; Бурковский, Мазей, 2016), то сообщества благоприятных местообитаний можно считать более организованными, чем суровых.
Авторы благодарны В.Г. Онипченко за ценные замечания по работе. В статье приведены результаты исследований, выполненных при финансовой поддержке Российского фонда фундаментальных исследований (грант № 16-04-00228).
Список литературы
Акатов В.В., Акатова Т.В., 2016. Уровень доминирования в травяных сообществах с разными моделями организации // Разнообразие и классификация растительности. Сб. науч. тр. Гос. Никитского ботан. сада / Под ред. Корженевского В.В., Ермакова Н.Б. Ялта. Т. 143. С. 16–24.
Баканов А.И., 1987. Количественная оценка доминирования в экологических сообществах. Деп. в ВИНИТИ 08.12.1987. № 8593-В87. 63 с.
Бигон М., Харпер Дж., Таунсенд К., 1989. Экология. Особи, популяции и сообщества. В 2-х томах. Т. 2. М.: Мир. 477 с.
Бурковский И.В., Мазей Ю.А., 2016. О сложности экологических систем (на примере сообщества морских инфузорий) // Russian J. Ecosystem Ecology. V. 1. № 1. С. 1–14. doi 10.21685/2500-0578-2016-1-4
Василевич В.И., 1983. Очерки теоретической фитоценологии. Л.: Наука. 248 с.
Василевич В.И., 1991. Доминанты в растительном покрове // Ботан. журн. Т. 76. № 12. С. 1674–1681.
Василевич В.И., 2014. Видовое разнообразие суходольных лугов северо-запада европейской России // Ботан. журн. Т. 99. № 2. С. 226–236.
Василевич В.И., 2015. Видовое разнообразие влажных лугов Европейской России // Ботан. журн. Т. 100. № 4. С. 372–381.
Гиляров А.М., 2010. В поисках универсальных закономерностей организации сообществ: прогресс на пути нейтрализма // Журн. общ. биологии. Т. 71. № 5. С. 386–401.
Кузнецова Н.А., 2009. Сообщества в экстремальных и антропогенных условиях (на примере таксоценозов коллембол) // Виды и сообщества в экстремальных условиях. Сб., посвященный 75-летию акад. Юрия Ивановича Чернова / Под ред. Бабенко А.Б., Матвеевой Н.В., Макарова О.Л., Головач С.И. Москва; София: Т-во науч. изд. КМК – PENSOFT Рb1. С. 412–429.
Кузьминов А.Н., 2009. Концептуальная модель ценологического управления в социально-экономических системах // Terra economicus (Экономический вестник Ростовского гос. ун-та). Т. 7. С. 28–32.
Лебедева Н.В., Криволуцкий Д.А., 2002. Биологическое разнообразие и методы его оценки // География и мониторинг биоразнообразия. М: Изд-во Научного и учебно-методического центра. С. 8–76.
Левич А.П., 1980. Структура экологических сообществ. М.: Изд-во МГУ. 181 с.
Миркин Б.М., Наумова Л.Г., 2012. Современное состояние основных концепций науки о растительности. Уфа: Гилем. 488 с.
Онипченко В.Г., 2013. Функциональная фитоценология: синэкология растений. M.: Красанд. 640 с.
Павлов В.Н., Онипченко В.Г., Аксенова А.А., Волкова Е.В., Зуева О.И., Макаров М.И., 1998. Роль конкуренции в организации альпийских фитоценозов Северо-Западного Кавказа: экспериментальный подход // Журн. общ. биологии. Т. 59. № 5. С. 453–476.
Работнов Т.А., 1983. Фитоценология. М.: Изд-во МГУ. 296 с.
Раменский Л.Г., 1938. Введение в комплексное геоботаническое использование земель. М.: Сельхозгиз. 620 с.
Уиттекер Р., 1980. Сообщества и экосистемы. М.: Прогресс. 327 с.
Чернов Ю.И., 2005. Видовое разнообразие и компенсационные явления в сообществах и биотических системах // Зоол. журн. Т. 84. № 10. С. 1221–1238.
Шитиков В.К., Зинченко Т.Д., Розенберг Г.С., 2011. Макроэкология речных сообществ: концепции, методы, модели. Тольятти: Кассандра. 255 с.
Adler P.B., Seabloom E.W., Borer E.T. et al., 2011. Productivity is a poor predictor of plant species richness // Science. V. 333. P. 1750–1753.
Aksenova A.A., Onipchenko V.G., Blinnikov M.S., 2004. Experimental study of plant relationships. Dominant removals. Alpine lichen heats // Alpine ecosystems in the Northwest Caucasus / Ed. Onipchenko V.G. Dordrecht: Kluver Acad. Publ. P. 236–244.
Azovsky A.I., 2009. Structural complexity of species assemblages and spatial scale of community organization: A case study of marine benthos // Ecological Complexity. V. 6. P. 308–315.
Bartha S., Szentes Sz., Horváth A., Házi J., Zimmermann Z., Molnár Cs. et al., 2014. Impact of mid-successional dominant species on the diversity and progress of succession in regenerating temperate grasslands // Applied Vegetation Sci. V. 17. № 2. P. 201–213.
Bell G., 2000. The distribution of abundance in neutral communities // Am. Nat. V. 155. № 5. P. 606–617.
Bengtsson J., Fagerstram T., Rydin H., 1994. Competition and coexistence in plant communities // TREE. V. 9. № 7. P. 246–250.
Berger W.H., Parker F.L., 1970. Diversity of planktonic foraminifera in deep-sea sediments // Science. V. 168. P. 1345–1347.
Bobbink R., Willems J.H., 1987. Increasing dominance of Brachypodium pinnatum (L.) Beauv. in Chalk Grasslands: a threat to a species-rich ecosystem // Biol. Conserv. V. 40. № 4. P. 301–314.
Boscaiu M., Lull C., Lidon A., Bautista I., Donat P., Mayoral O., Vicente O., 2008. Plant responses to abiotic stress in their natural habitats // Horticulture. V. 65. № 1. P. 53–58.
Callaway R.M., Ridenour W.M., 2004. Novel weapons: a biochemically based hypothesis for invasive success and the evolution of increased competitive ability // Frontiers Ecol. Environ. № 2. P. 433–436.
Caruso T., Pigino G., Bernini F., Bargagli R., Migliorin M., 2007.The Berger–Parker index as an effective tool for monitoring the biodiversity of disturbed soils: a case study on Mediterranean oribatid (Acari: Oribatida) assemblages // Biodivers. Conserv. V. 16. P. 3277–3285.
Caswell H., 1976. Community structure: a neutral model analysis // Ecol. Mon. V. 46. P. 327–354.
Chalcraft D.R., Wilsey B.J., Bowles C., Willig M.R., 2009. The relationship between productivity and multiple aspects of biodiversity in six grassland communities // Biodiversity Conserv. V. 18. P. 91–104.
Cherednichenko O.V., 2004. Experimental study of plant relationships. Dominant removals. Removals in the Geranium gymnocaulon – Hedysarum caucasicum meadow // Alpine ecosystems in the Northwest Caucasus / Ed. Onipchenko V.G. Dordrecht: Kluver Acad. Publ. P. 244–250.
Cornell H.V., 1993. Unsaturated patterns in species assemblage: the role of regional processes in setting local species richness // Species diversity in ecological communities: historical and geographical perspectives / Eds Ricklefs R.E., Schluter D. Chicago: Chicago Univ. Press. P. 243–253.
Cornell H.V., Lawton J.H., 1992. Species interactions, local and regional processes, and limits to the richness of ecological communities: a theoretical perspective // J. Anim. Ecol. V. 61. P. 1–12.
Csergő A.M., Demeter L., Turkington R., 2013. Declining diversity in abandoned grasslands of the Carpathian Mountains: do dominant species matter? // PloS One. V. 8. № 8: e73533. doi:10.1371
Drobner U., Bibby J., Smith B., Wilson J.B., 1998.The relation between community biomass and evenness: what does community theory predict, and can these predictions be tested? // Oikos. V. 82. P. 295–302.
Ernest S.K.M., Brown J.H., 2001. Homeostasis and compensation: the role of species and resources in ecosystem stability // Ecology. V. 82. № 8. P. 2118–2132.
Ferreira F.C., Petrere-Jr. M., 2008. Comments about some species abundance patterns: classic, neutral, and niche partitioning models // Braz. J. Biol. V. 68. № 4. P. 1003–1012.
Fretwell D., 1977. The regulation of plant communities by food chains exploiting them // Perspectives in Biology and Medicine. V. 20. P. 169–185.
Goldberg D., Novoplansky A., 1997. On the relative importance of competition in unproductive environments // J. Ecology. V. 85. P. 409–418.
Grace J.B., 1991. A clarification of the debate between Grime and Tilman // Functional Ecol. V. 5. P. 583–587.
Grime J.P., 1977. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory // Am. Nat. V. 111. № 982. P. 1169–1194.
Grime J.P., 2001. Plant strategies, vegetation processes, and ecosystem properties. 2nd Edition. Chichester: John Wiley and Sons. 417 p.
Hejda M., Stajerova K., Pyšek P., 2016. Dominance has a biogeographical component: do plants tend to exert stronger impacts in their invaded rather than an native range? // J. Biogeogr. V. 44. № 1. P. 18–27.
Houlahan J.E., Findlay C.S., 2004. Effect of invasive plant species on temperate wetland plant diversity // Conservation Biology. V. 18. № 4. P. 1132–1138.
Hubbell S.P., 1979. Tree dispersion, abundance, and diversity in a tropical dry forest // Science. V. 203. P. 1299–1309.
Hubbell S.P., 2005. Neutral theory in community ecology and the hypothesis of functional equivalence // Functional Ecol. V. 19. P. 166–172.
Hulme P.E., Bremner E.T., 2006. Assessing the impact of Impatiens glandulifera on riparian habitats: partitioning diversity components following species removal // J. Applied Ecol. V. 43. P. 43–50.
Huston M., 1979. A general hypothesis of species diversity // Am. Nat. V. 113. № 1. P. 81–101.
Lamb E.G., Cahill J.F., 2008. When competition does not matter: grassland diversity and community composition // Am. Nat. V. 171. P. 777–787.
Longino J.T., Colwell R.K., 2011. Density compensation, species composition, and richness of ants on a neotropical elevational gradient // Ecosphere. V. 2. № 3. P. 1–20.
Ma M., 2005. Species richness vs. evenness: independent relationship and different responses to edaphic factors // Oikos. V. 111. P. 192–198.
Maarel E., van der, Noest V., Palmer M.W., 1995.Variation in species richness on small grassland quadrats: niche structure or small-scale plant mobility? // J. Vegetation Sci. V. 6. P. 741–752.
Magguran A., 1988. Ecological diversity and its measurement. Princeton, NJ: Princeton Univ. Press. 181 p.
Mangla S., Sheley R.L., James J.J., Radosevich S.R., 2011. Role of competition in restoring resource poor arid systems dominated by invasive grasses // J. Arid Environments. V. 75. P. 487–493.
McKane R.B., Johnson L.C., Shaver G.R., Nadelhoffer K.J., Rastetter E.B. et al., 2002. Resource-based niches provide a basis for plant species diversity and dominance in arctic tundra // Nature. V. 415. P. 68–71.
Mirkin B.M., 1994. Which plant communities do exist? // J. Veget. Sci. V. 5. № 2. P. 283–284.
Mulder C.P.H., Bazeley-White E., Dimitrakopoulos P.G., Hector A., Scherer-Lorenzen M., Schmid B., 2004. Species evenness and productivity in experimental plant communities // Oikos. V. 107. P. 50–63.
Munson S.M., Lauenroth W.K., 2009. Plant population and community responses to removal of dominant species in the shortgrass steppe // J. Veg. Sci. V. 20. P. 224–232.
Oksanen J., 1996. Is the humped relationship between species richness and biomass an artifact due to plot size? // J. Ecology. V. 84. P. 293–295.
Oksanen L., Fretwell S.D., Arrud J., Niemala P., 1981. Exploitation ecosystems in gradients of primary productivity // Am. Nat. V. 118. P. 240–261.
Olff H., Bakker J.P., 1998. Do intrinsically dominant and subordinate species exist? A test statistic for field data // Applied Veg. Sci. V. 1. P. 15–20.
Onipchenko V.G., Semenova G.V., Maarel E., van der, 1998. Population strategies in severe environments: alpine plants in the northwestern Caucasus // J. Veg. Sci. V. 9. P. 27–40.
Onipchenko V.G., Blinnikov M.S., Gerasimova M.A., Volkova E.V., Cornelissen J.H.C., 2009. Experimental comparison of competition and facilitation in alpine communities varying in productivity // J. Veg. Sci. V. 20. № 4. P. 718–727.
Palmer M.W., Maarel E., van der, 1995. Variance in species richness, species association and niche limitation // Oikos. V. 73. P. 203–213.
Panchal N.S., Pandey A.N., 2004. Analysis of vegetation of rampara forest in Saurashtra Region of Gujarat State of India // Trop. Ecol. V. 45. № 2. P. 223–231.
Paquette A., Messier C., 2011. The effect of biodiversity on tree productivity: from temperate to boreal forests // Global Ecol. Biogeogr. V. 20. P. 170–180.
Peet R.K., Christensen N.L., 1988. Changes in species diversity during secondary forest succession on the North Carolina piedmont // Diversity and pattern in plant communities / Eds During H.I., Werge M.I.A., Willems J.H. Hague. Netherlands: SPB Academic Publishing. P. 233–245.
Piper J.K., 1995. Composition of prairie plant communities on productive versus unproductive sites in wet and dry years // Can. J. Bot. V. 73. P. 1635–1644.
Poggio S.L.,Ghersa C.M., 2011. Species richnes and evenness as a function of biomass in arable plant communities // Weed Research. V. 51. P. 241–249.
Qian H., Ricklefs R.E., 2004. Taxon richness and climate in Angiosperms: is there a globally consistent relationship that precludes region effects? // Am. Nat. V. 163. № 5. P. 773–779.
Reinhart K.O., Greene E., Callaway R.M., 2005. Effects of Acer platanoides invasion on understory plant communities and tree regeneration in the Rocky Mountains // Ecography. V. 28. P. 573–582.
Sammul M., Kull K., Oksanen L., Veromann P., 2000. Competition intensity and its importance: results of field experiments with Anthoxanthum odoratum /// Oecologia V. 125. P. 18–25.
Sammul M., Oksanen L., Mӓgi M., 2006. Regional effects on competition–productivity relationship: a set of field experiments in two distant regions // / Oikos. V. 112. P. 138–148.
Sasaki T., Lauenroth W.K., 2011. Dominant species, rather than diversity, regulates temporal stability of plant communities // Oecologia. V. 166. № 3. P. 761–768.
Šímová I., Li Y.M., Storch D., 2013. Relationship between species richness and productivity in plants: the role of sampling effect, heterogeneity and species pool // J. Ecology. V. 101. P. 161–170.
Somodi I., Virágh K., Podani J., 2008. The effect of the expansion of the clonal grass Calamagrostis epigejos on the species turnover of a semi-arid grassland // Applied Vegetation Sci. V. 11. P. 187–194.
Souza L., Weltzin J.F., Sanders N.J., 2011. Differential effects of two dominant plant species on community structure and invisibility in an old-field ecosystem // J. Plant. Ecol. V. 4. P. 123–131.
Stirling G., Wilsey B., 2001. Empirical relationships between species richness, evenness, and proportional diversity // Am. Nat. V. 158. P. 286–300.
Vermeer J.G., Verhoeven J.T.A., 1987. Species composition and biomass production of mesotrophic fens in relation to the nutrient status of the organic soil // Acta Oecol-Oec Plant. V. 8. P. 321–330.
Wilsey B., Stirling G., 2007. Species richness and evenness respond in a different manner to propagule density in developing prairie microcosm communities // Plant. Ecol. V. 190. P. 259–273.
Wright D.H., 1983. Species–energy theory – an extension of species–area theory // Oikos. V. 41. P. 496–506.
Wright D.H., Currie D.J., Maurer B.A., 1993. Energy supply and patterns of species richness on local and regional scales // Species diversity in ecological communities: historical and geographical perspectives / Eds Ricklefs R.E., Schluter D. Chicago: Chicago Univ. Press. P. 66–75.
Yodzis P., 1978. Competition for space and the structure of ecological communities // Lecture Notes in Biomathematics. V. 25. P. 1–191.
Zefferman E., Stevens J.T., Charles G.K., Dunbar-Irwin M., Emam T., Fick S., Morales L.V., Wolf K. M., Young D.J.N., Young T.P., 2015. Plant communities in harsh sites are less invaded: a summary of observations and proposed explanations // AoB Plants. V. 7: plv056.
Zhang H., John R., Peng Z., Yuan J., Chu C., Du G., Zhou S., 2012. The relationship between species richness and evenness in plant communities along a successional gradient: a study from sub-alpine meadows of the Eastern Qinghai-Tibetan Plateau, China // PloS One. V. 7. № 11: e49024.
Zhang J., Qiao X., Liu Y., Lu J., Jiang M., Tang Z., Fang J., 2015. Species-abundance distributions of tree species varies along climatic gradients in China’s forests // J. Plant Ecol. V. 23. № 9. P. 1–7.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии