

Журнал общей биологии, 2019, T. 80, № 2, стр. 135-144
Особенности фенологии и температурных норм развития двух видов Clematis L. (Ranunculaceae) в условиях заповедника “Галичья гора” (Липецкая область)
М. В. Ушаков 1, *, Т. В. Недосекина 1
1 Воронежский государственный университет, заповедник “Галичья гора”
399240 Липецкая область, Задонский район, п/о Донское, Россия
* E-mail: ushakov@dev-reserve.vsu.ru
Поступила в редакцию 15.01.2018
После доработки 20.08.2018
Принята к публикации 27.09.2018
Аннотация
Проведено сравнение фенодат, температурных норм и их региональных параметров для шести стадий сезонного развития у двух экологически различающихся видов ломоноса: Clematis integrifolia и C. recta. Сравниваемые температурные нормы получены по оригинальной методике на основе данных фенологических наблюдений, проводившихся в 1990–2014 гг. в заповеднике “Галичья гора” (Липецкая область). Все наблюдаемые стадии, за исключением стадии массового созревания семян, у C. recta происходили в более поздние сроки, чем у C. integrifolia. Установлено, что для лесного вида C. recta свойственна более высокая потребность в количестве необходимого для развития тепла, чем для C. integrifolia, растущего в травяных сообществах. На стадиях начала вегетации, цветения и начала созревания семян это выражалось в более высоких значениях и тепловых констант, и региональных параметров развития. Также предполагается, что в условиях заповедника у C. recta в зимний период развитие не останавливалось, и в это же время формировались цветочные почки. Исследования показали, что в ходе эволюции при адаптации к разным условиям существования у двух видов растений происходили изменения в физиологии, выражающиеся в том числе и в изменениях температурных потребностей для развития.
Фенологические события в жизни живых организмов, с одной стороны, являются физиологическим ответом на периодические изменения климатических факторов окружающей среды. С другой стороны, физиологический ответ имеет наследственную природу и проявляется в форме, специфичной для того или иного таксона. Порядок наступления разных фаз развития, как и энергетические затраты на их прохождение задаются наследственным аппаратом организма и являются результатом действия естественного отбора. Очевидно, что такой отбор, который должен приводить к различиям в прохождении фаз развития у родственных видов, носит адаптивный характер к условиям обитания.
Одним из факторов, определяющих развитие пойкилотермных организмов, выступает температура. Она активирует протекающие физиологические процессы и регулирует их скорость. В свое время было установлено, что для прохождения стадии развития организму требуется определенное количество тепла (Sanderson, Pears, 1913), для расчета которого Блунком предложена формула (Blunk, 1923)
где $\sum {{{T}_{{{\text{eff}}}}}} $ – тепловая константа или сумма эффективных температур (т.е. то тепло, которое необходимо набрать организму для достижения определенной стадии развития); $C$ – пороговая температура (т.е. температура, ниже которой развитие останавливается); $D$ – число дней развития при температуре среды ${{T}_{{\text{a}}}}$ или эффективной температуре ${{T}_{{{\text{eff}}}}}$ (т.е. число дней, когда ${{T}_{{\text{a}}}}$ равна или превышает C). Формула содержит две константы $\sum {{{T}_{{{\text{eff}}}}}} $ и C, которые непосредственно не определяются, а устанавливаются путем интерполяции при известных D и ${{T}_{{{\text{eff}}}}}.$В природных условиях в сильно переменной среде установление констант представляет большую трудность. Главные усилия исследователей направлены на определение пороговой температуры (Arnold, 1959; Bassett et al., 1961; Campbell et al., 1974). Авторами настоящей статьи также был предложен метод определения этой константы (Ушаков, Недосекина, 2017; Ushakov, Nedosekina, 2017), который, по нашему мнению, представляется более эффективным. В отличие от остальных подходов, он опирается на свойства уравнения Блунка и, можно сказать, непосредственно указывает на пороговую температуру стадии развития.
Мы применили наш подход для установления констант у двух видов растений из р. Clematis L., 1753 сем. Ranunculaceae Juss., 1789 – ломоноса цельнолистного C. integrifolia L., 1753 и ломоноса прямого C. recta L., 1753, произрастающих на территории заповедника “Галичья гора” в Липецкой области. Можно ожидать, что различия в экологии этих родственных видов должны вести к различиям в температурных потребностях их развития. Задачa исследования – выявление этих различий путем сравнения фенологии и температурных норм сезонного развития ломоноса цельнолистного и ломоноса прямого.
МАТЕРИАЛ И МЕТОДЫ
Заповедник “Галичья гора” расположен в центре Липецкой области в окружении сельскохозяйственного ландшафта. Он представлен склоновыми поверхностями, плакорами и поймами, имеет кластерную структуру, состоящую из шести небольших участков общей площадью в 231 га. Растительность состоит из нагорных дубрав, березняков, небольших сосняков, скальных и прибрежно-водных сообществ, а также сообществ пойменных и ксерофитных лугов.
Р. Clematis в заповеднике представлен двумя видами, которые и стали объектами исследования.
Ломонос цельнолистный С. integrifolia. Его ареал охватывает Кавказ, юг Западной и Восточной Сибири, Среднюю и Восточную Европу, Средиземноморье (Балканы), Среднюю и Малую Азию (Флора…, 2001). В России встречается на юге европейской части, в Сибири, Предкавказье. В средней полосе изредка встречается во всех областях Центрального Черноземья. В заповеднике “Галичья гора” произрастает в разнотравно-злаковых и кустарниковых сообществах по известняковым склонам и придолинным плато, по опушкам дубрав.
Ломонос прямой C. recta распространен на Кавказе, в Средней Европе, Средиземноморье и Малой Азии. В России встречается в Европейской части и в Предкавказье. Растет в широколиственных лесах, преимущественно в разреженных дубравах, на опушках, среди кустарников в долинах рек (Флора…, 2001). В заповеднике “Галичья гора” встречается в нагорных дубравах, по опушкам широколиственных лесов с преобладанием дуба и липы, сравнительно редко в кустарниково-злаково-разнотравных сообществах.
Рассматриваемые виды входят в разные секции. В наиболее проработанной на настоящий момент системе р. Clematis, предложенной Джонсоном (Johnson, 2001), С. integrifolia отнесен к секции Viorna (non Reichb.) Prantl., имеющей евразийское распространение, а C. recta – к секции Flammula DC из Восточной Азии. По данным молекулярно-генетических исследований (Miikeda et al., 2006; Xie et al., 2011), виды двух секций разошлись совсем недавно, приблизительно чуть больше 5 млн лет назад. Это совпадает с общим характером “взрывного” видообразования р. Clematis, вызванного, как считается, глобальными геологическими и эколого-климатическими изменениями в конце третичного периода (Xie et al., 2011).
Оба вида имеют сходную жизненную форму. Они принадлежат к гемикриптофитам по Раункиеру (Raunkiaer, 1907) и к кистекорневым травянистым многолетникам по И.Г. Серебрякову (1962). По феноритмотипу или характеру фенологического развития в годичном цикле (Борисова, 1972) изученные виды относятся к длительновегетирующим весеннее-летне-осеннезеленым растениям с периодом зимнего покоя. Осенью прямостоячие побеги у обоих видов отмирают до уровня почвы. Пазушные почки в верхней части корневища становятся почками возобновления, из которых в условиях заповедника “Галичья гора” еще осенью начинают отрастать новые побеги, остающиеся на протяжении всей зимы в подземном положении. Весной с приходом тепла из этих побегов развивается надземная часть растения.
Фенологические наблюдения проводили по общепринятым методикам (Бейдеман, 1954, 1974; Шульц, 1981) на постоянных маршрутах в пределах однородных экотопов для каждого вида. “Видимое” наступление фенологической фазы устанавливалось в тот день, когда в фазу вступало не менее 10% растений на определенной площадке. За начало вегетации принимали дату обнаружения побегов из почек возобновления, заложенных в основании сохранившихся генеративных побегов; за начало бутонизации – дату, когда бутоны едва заметны и еще зеленого цвета; начало цветения – когда в фазу вступает не менее 10% растений, хорошо заметны вполне раскрывшиеся венчики цветков, массовое цветение – при зацветании не менее половины наблюдаемых растений; начало созревания плодов наступает при побурении зеленых плодов (они приобретают светло-коричневую окраску), массовое созревание – когда плоды становились бурыми (окраска зрелых плодов). В исследовании использовался материал фенологических наблюдений в заповеднике “Галичья гора” с 1990 по 2014 год, а также полученные с метеопоста заповедника среднесуточные температуры воздуха за этот период. Подготовка к анализу включала в себя создание таблиц, столбцы которых отражали фазы развития, а строки содержали представленные в виде юлианского дня (номер дня от начала года) даты для каждого года наблюдений. Также создавалась таблица со среднесуточными температурами, в которой столбцы представляли годы наблюдений, а в пронумерованных строках – последовательность температур за каждый день года.
Оценка температурных норм выполнялась с помощью разработанного нами способа (Ушаков, Недосекина, 2017; Ushakov, Nedosekina, 2017). Из таблицы со среднесуточными температурами воздуха ${{T}_{{\text{m}}}}$ выбирались ряды чисел, ограниченные началом года и юлианским днем рассматриваемой стадии развития. Поиск пороговой температуры $C$ для стадии развития для каждого значения экзаменуемой температуры связан с вычислением линейной регрессии
где11 $\sum T $ – сумма среднесуточных температур воздуха, равных или превышающих экзаменуемую температуру, D – число дней развития при температуре, равной или превышающей экзаменуемую температуру, a и b – коэффициенты регрессии. Оценивали диапазон температур от –5 до 20°C с шагом в 1°C. Таким образом, рассчитывалось 26 регрессий, набор которых позволил получить две функции. Одну функцию образовывала последовательность значений коэффициента регрессии b в порядке перечисления тестируемой температуры, другую – последовательность значимости p критерия Стьюдента для этого коэффициента или последовательность значимости p для критерия Фишера качества подгонки регрессии под наблюдаемые данные (оба значения p совпадают). На графике двух функций на пороговую температуру указывало наименьшее значение b в области значений p, превышающих 5%-ный порог вероятности принятия нулевой гипотезы (т.е. когда b статистически не отличается от нуля и утрачивается линейная зависимость между $\sum T $ и D).После установления пороговой температуры C температура $T = {{T}_{{\text{m}}}} - C,$ по которой производилась оценка, принималась за эффективную температуру ${{T}_{{{\text{eff}}}}},$ т.е. ${{T}_{{{\text{eff}}}}} = {{\sum T } \mathord{\left/ {\vphantom {{\sum T } D}} \right. \kern-0em} D}$ ($\sum T $ выполнялась только для тех дней, для которых ${{T}_{{\text{m}}}} \geqslant C$). Сумму эффективных температур $\sum {{{T}_{{{\text{eff}}}}}} $ вычисляли путем аппроксимации зависимости между D и ${{T}_{{{\text{eff}}}}}$ по гиперболической формуле Блунка (Blunk, 1923)
Линеаризация формулы Блунка приводит к распространенному уравнению скорости развития G (Campbell et al., 1974):
Здесь b – коэффициент термолабильности, характеризующий зависимость скорости развития от температуры; $a = - bC.$ При использовании ${{T}_{{{\text{eff}}}}}$ линия регрессии проходит через начало координат, при использовании ${{T}_{{\text{m}}}}$ – через точку на оси абсцисс, соответствующую C.
Средние фенодаты, а также региональные показатели ($\sum T $, D, ${{T}_{{{\text{eff}}}}}$ и G) одной и той же стадии развития сравнивались между видами с помощью критерия Стьюдента, их дисперсии – с помощью критерия Фишера.
Все вычисления производили в программах MS Office Excel 2003, StatSoft STATISTICA 6.0 и MathCAD 2001.
РЕЗУЛЬТАТЫ
Все фенофазы (за исключением стадий созревания плодов), отмечаемые у ломоноса прямого в заповеднике “Галичья гора”, приходились на более поздние даты в сравнении с ломоносом цельнолистным (табл. 1). Эти различия статистически значимы (табл. 1, значения критерия Стьюдента). И если начало вегетации и начало бутонизации у лесного вида отставали от тех же стадий другого вида примерно на неделю, то на стадии цветения отставание уже составляло 2–3 недели. На стадиях созревания плодов статистических различий между видами не обнаружилось, возможно из-за сильной изменчивости фенодат, на которую указывают значения дисперсии.
Таблица 1.
Статистические данные по фенологическим наблюдениям стадий развития двух видов Clematis в заповеднике “Галичья гора” за период с 1990 по 2014 год
| Стадия развития | Вид Clematis | Число лет наблюдений | Средняя дата и ее ошибка | Критерий Стьюдента | Самая ранняя дата | Самая поздняя дата | Дисперсия | Критерий Фишера | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| $df$ | ${{t}_{{{\text{st}}}}}$ | $p$ | $F$ | $p$ | |||||||
| Начало вегетации | integrifolia | 23 | 20.IV ± 2 | 46 | 3.10 | 0.003 | 4.IV | 5.V | 84.66 | 2.14 | 0.072 |
| recta | 25 | 27.IV ± 1 | 12.IV | 7.V | 39.53 | ||||||
| Начало бутонизации | integrifolia | 24 | 13.V ± 1 | 47 | 2.74 | 0.009 | 5.V | 24.V | 28.80 | 2.61 | 0.025 |
| recta | 25 | 19.V ± 2 | 2.V | 7.VI | 75.16 | ||||||
| Начало цветения | integrifolia | 24 | 26.V ± 1 | 47 | 7.59 | 0.000 | 17.V | 10.VI | 44.55 | 1.65 | 0.234 |
| recta | 25 | 12.VI ± 2 | 25.V | 22.VI | 73.56 | ||||||
| Массовое цветение | integrifolia | 24 | 3.VI ± 2 | 47 | 6.85 | 0.000 | 22.V | 15.VI | 57.04 | 1.35 | 0.471 |
| recta | 25 | 19.VI ± 2 | 31.V | 1.VII | 77.21 | ||||||
| Начало созревания плодов | integrifolia | 24 | 9.VII ± 4 | 43 | 1.79 | 0.080 | 12.VI | 15.VIII | 470.98 | 1.85 | 0.171 |
| recta | 21 | 19.VII ± 3 | 26.VI | 27.VIII | 255.46 | ||||||
| Массовое созревание плодов | integrifolia | 23 | 7.VIII ± 4 | 42 | 0.57 | 0.569 | 30.VI | 30.VIII | 284.00 | 1.90 | 0.153 |
| recta | 21 | 4.VIII ± 3 | 20.VII | 31.VIII | 149.15 | ||||||
Вариативность фенологических дат является отражением физиологического ответа растений на межгодовые изменения климатических условий, при которых происходит развитие. Изменчивость в сроках начала вегетации, начала бутонизации, начала и массового цветения у растений существенно ниже в сравнении с более высокой изменчивостью сроков начала и массового созревания плодов. При межвидовых сравнениях в целом не было обнаружено значимых различий в изменчивости сроков наступления рассматриваемых фенологических явлений (табл. 1, критерий Фишера). Исключение составляла более высокая изменчивость в сроках бутонизации у ломоноса прямого.
Поиск пороговых температур C для каждой стадии развития у двух видов ломоноса (рис. 1, 2) показал, что эта константа принимает одинаковое значение для фенофаз начала и массового цветения, а также для фенофаз начала и массового созревания плодов.
Рис. 1.
Определение пороговой температуры для C. integrifolia. Графики функций коэффициента регрессии b и значимости p в диапазоне температур от –5 до 20°C. Пунктирная линия обозначает 5%-ный порог значимости. Стадии: а – начала вегетации; б – начала бутонизации; в – начала цветения; г – массового цветения; д – начала созревания плодов; е – массового созревания плодов.
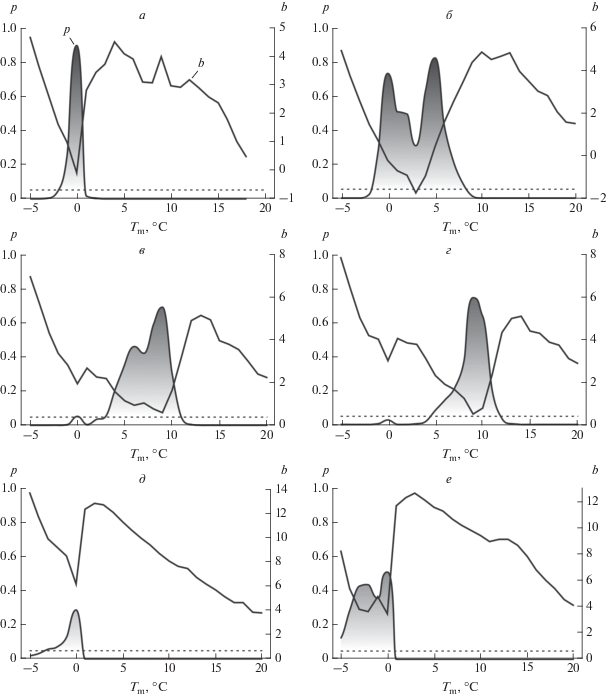
Рис. 2.
Определение пороговой температуры для C. recta. Обозначения и стадии развития те же, что и на рис. 1.
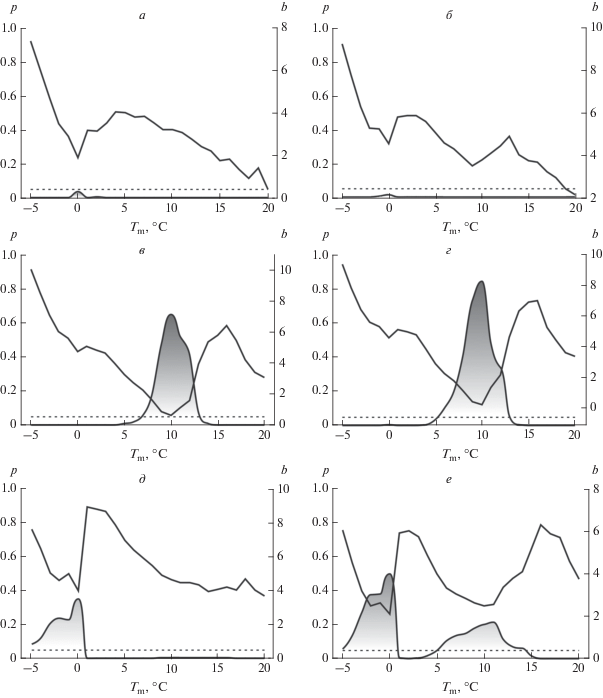
Совпадение пороговых температур для фенофаз начала и массового цветения говорит о том, что они являются лишь фазами одной стадии – цветения. Их рассмотрение позволяет судить о фенологической неоднородности популяции. Ее причинами могут быть как возрастной характер хронологических сдвигов у растений в сроках наступления фенофаз (Шнелле, 1961), так и особенности индивидуального развития вследствие взаимодействия между генотипом и внешней средой. Одну стадию развития также представляют начало и массовое созревание плодов, но на разных своих этапах. Поэтому различия между ними в значительной степени отражают продолжительность формирования плодов.
У ломоноса цельнолистного никаких неожиданностей при поиске пороговой температуры не обнаружилось (рис. 1). Однако они возникли при установлении этой константы для первых двух стадий сезонного развития у ломоноса прямого. На рис. 2а и 2б функция b имеет хорошо выраженные “зубцы”, указывающие на равное значение $C$ и для начала вегетации, и для начала бутонизации. Однако в обоих случаях функция p не “поднимается” выше 5%-ной границы, что говорит о сохранении линейности зависимости между $\sum T $ и D. Наличие выраженного “зубца” у функции b указывает на существование пороговой температуры. Низкие значения функции p свидетельствуют, однако, о том, что в зимний период развитие растения в условиях заповедника “Галичья гора” не останавливается, или такие задержки непродолжительны.
Нахождение пороговой температуры C позволило оценить тепловую константу для двух видов ломоноса (табл. 2). Ломоносу прямому для достижения большинства сезонных стадий развития требуется потребить больше тепла, чем ломоносу цельнолистному. Так, сумма эффективных температур $\sum {{{T}_{{{\text{eff}}}}}} $ для начала вегетации у этих двух видов различается почти на 100°C. Для достижения цветения ломоносу прямому также требуется “усвоить” примерно на 100°C больше, чем ломоносу цельнолистному, даже при том, что и пороговая температура для этой стадии у лесного вида выше на 1°C. Скорее всего эти особенности развития у растений сохраняются и на стадии бутонизации. Однако это утверждение осложнено более высокой пороговой температурой у ломоноса цельнолистного. В то же время предположение о бóльших температурных потребностях для достижения стадии бутонизации у ломоноса прямого может подтверждаться более поздними сроками наступления этой фенофазы (табл. 1).
Таблица 2.
Температурные нормы для шести стадий развития двух видов Clematis, вычисленные по данным фенологических наблюдений в заповеднике “Галичья гора”
| Стадия развития | C. integrifolia | C. recta | ||||
|---|---|---|---|---|---|---|
| $C$, $^\circ {\text{C}}$ | $\sum {{{T}_{{{\text{eff}}}}}} ,$$^\circ {\text{C}}$ | $b$ | $C$, $^\circ {\text{C}}$ | $\sum {{{T}_{{{\text{eff}}}}}} ,$$^\circ {\text{C}}$ | $b$ | |
| Начало вегетации | 0 | 149.1 | 0.0067 | 0 | 253.3 | 0.0039 |
| Начало бутонизации | 3 | 304.0 | 0.0033 | 0 | 539.1 | 0.0019 |
| Начало цветения | 9 | 184.9 | 0.0054 | 10 | 284.7 | 0.0035 |
| Массовое цветение | 9 | 258.5 | 0.0039 | 10 | 350.3 | 0.0029 |
| Начало созревания плодов | 0 | 1391.5 | 0.0007 | 0 | 1685.5 | 0.0006 |
| Массовое созревание плодов | 0 | 1995.4 | 0.0005 | 0 | 2046.3 | 0.0005 |
Очевидно, что сроки начала вегетации, бутонизации и цветения отражают особенности более быстрого развития маргинальной части особей. Массовое же цветение, а также начало и массовое созревание плодов в значительной степени выражают физиологическую “моду”. Разница в сумме эффективных температур между маргинальными и “модальными” фазами цветения у двух видов ломоноса практически одинакова и составила меньше 80°C.
Межвидовые различия в температурных потребностях утрачиваются на стадии массового созреванием семян, когда для ее достижения обоим видам требуется “освоить” практически одинаковое количество тепла (около 2000°C).
Температурные нормы в условиях местной среды находят свое выражение в региональных особенностях разных параметров развития, главным образом проявляющиеся через число дней развития D. Скорость развития G является просто его обратной величиной. Линейно коррелирующие друг с другом сумма температур $\sum T ,$ которая в российских исследованиях часто неверно принимается за тепловую константу, и эффективная температура ${{T}_{{{\text{eff}}}}}$ также обратно связаны с числом дней развития через сумму эффективных температур $\sum {{{T}_{{{\text{eff}}}}}} $ (Ушаков, Недосекина, 2017; Ushakov, Nedosekina, 2017). Их значения для двух видов ломоноса в условиях заповедника “Галичья гора” за период исследований представлены в табл. 3. Здесь прослеживается та же закономерность, что и при рассмотрении фенологических дат и температурных норм развития. Для межвидового сравнения по приведенным выше причинам использовано число дней развития D (табл. 4). Значимые различия в продолжительности прохождения стадии бутонизации у двух видов вполне ожидаемы в силу межвидовых различий по двум температурным константам. Стадии цветения у ломоноса прямого также значимо отличались от ломоноса цельнолистного в сторону большей своей продолжительности (примерно на две недели). Различия в продолжительности развития между двумя видами пропадали на стадии массового созревания семян. Отсутствие различий в продолжительности стадии вегетации вызвано особенностями вычисления числа дней развития у ломоноса прямого. Поскольку развитие этого вида предположительно происходит и в зимнее время, этот период оказался не учтенным в расчетах, из-за чего величина D для ломоноса прямого получилась явно заниженной даже при близких пороговых температурах у двух видов.
Таблица 3.
Параметры стадий развития двух видов Clematis в условиях заповедника “Галичья гора”, вычисленные по данным фенологических наблюдений за период с 1990 по 2014 год
| Стадия развития | Вид Clematis | $\sum T $, °C | ${{T}_{{{\text{eff}}}}}$, °C | $D$ | $G$ | ||||
|---|---|---|---|---|---|---|---|---|---|
| среднее и ошибка | мин. – макс. | среднее и ошибка | мин. – макс. | среднее и ошибка | мин. – макс. | среднее и ошибка | мин. – макс. | ||
| Начало вегетации | integrifolia | 180.7 ± 11.11 | 85.0–279.4 | 3.4 ± 0.29 | 1.4–6.5 | 57.4 ± 2.94 | 36–88 | 0.018 ± 0.0009 | 0.011–0.028 |
| recta | 270.4 ± 12.39 | 183.0–431.4 | 4.5 ± 0.19 | 2.3–6.5 | 61.2 ± 2.80 | 39–90 | 0.017 ± 0.0008 | 0.011–0.026 | |
| Начало бутонизации | integrifolia | 317.9 ± 9.92 | 248.5–448.9 | 7.3 ± 0.32 | 4.1–11.5 | 44.7 ± 1.27 | 36–63 | 0.023 ± 0.0006 | 0.016–0.028 |
| recta | 573.9 ± 26.01 | 366.3–826.4 | 6.3 ± 0.25 | 4.4–8.3 | 92.3 ± 2.92 | 70–121 | 0.011 ± 0.0003 | 0.008–0.014 | |
| Начало цветения | integrifolia | 202.4 ± 8.67 | 122.0–289.1 | 5.7 ± 0.30 | 3.2–8.8 | 36.2 ± 1.23 | 26–50 | 0.028 ± 0.0009 | 0.020–0.039 |
| recta | 303.5 ± 10.38 | 183.7–394.4 | 6.2 ± 0.29 | 3.5–9.4 | 50.0 ± 1.61 | 37–69 | 0.021 ± 0.0007 | 0.015–0.027 | |
| Массовое цветение | integrifolia | 275.5 ± 10.47 | 188.1–379.7 | 6.4 ± 0.31 | 3.7–8.9 | 44.1 ± 1.56 | 32–62 | 0.023 ± 0.0008 | 0.016–0.031 |
| recta | 374.9 ± 12.83 | 220.2–476.2 | 6.7 ± 0.29 | 3.7–10.0 | 57.3 ± 1.56 | 43–74 | 0.018 ± 0.0005 | 0.014–0.023 | |
| Начало созревания плодов | integrifolia | 1545.4 ± 80.78 | 1056.0–2052.0 | 9.7 ± 0.49 | 6.4–13.2 | 160.3 ± 3.05 | 138–189 | 0.006 ± 0.0001 | 0.005–0.007 |
| recta | 1739.7 ± 56.76 | 1436.0–2374.0 | 10.2 ± 0.33 | 8.0–12.6 | 172.0 ± 3.15 | 151–202 | 0.006 ± 0.0001 | 0.005–0.007 | |
| Массовое созревание плодов | integrifolia | 2132.4 ± 73.13 | 1345.0–2567.0 | 12.2 ± 0.44 | 7.5–15.9 | 175.3 ± 3.05 | 153–204 | 0.006 ± 0.0001 | 0.005–0.007 |
| recta | 2071.2 ± 40.75 | 1793.0–2424.0 | 11.8 ± 0.29 | 9.5–15.0 | 176.0 ± 3.15 | 155–206 | 0.006 ± 0.0001 | 0.005–0.006 | |
Таблица 4.
Статистические различия в числе дней развития $D$ между двумя видами Clematis в заповеднике “Галичья гора"
| Стадия развития | Критерий Стьюдента | Критерий Фишера | |||||
|---|---|---|---|---|---|---|---|
| $df$ | ${{t}_{{{\text{st}}}}}$ | $p$ | $\sigma _{1}^{2}$ | $\sigma _{2}^{2}$ | $F$ | $p$ | |
| Начало вегетации | 47 | 1.49 | 0.144 | 14.5 | 14.0 | 1.07 | 0.862 |
| Начало бутонизации | 45 | 4.91 | 0.000 | 10.9 | 14.3 | 1.73 | 0.204 |
| Начало цветения | 45 | 6.73 | 0.000 | 34.9 | 62.3 | 1.78 | 0.179 |
| Массовое цветение | 45 | 5.96 | 0.000 | 55.8 | 58.6 | 1.05 | 0.911 |
| Начало созревания плодов | 41 | 2.67 | 0.011 | 213.2 | 198.2 | 1.08 | 0.880 |
| Массовое созревание плодов | 41 | 0.17 | 0.867 | 213.2 | 198.2 | 1.08 | 0.880 |
ОБСУЖДЕНИЕ
Полученные результаты показывают, что два вида ломоноса, произрастающие в заповеднике “Галичья гора” в разных экологических условиях, имеют различия в температурных потребностях для своего развития. Между их температурными нормами, продолжительностью стадии развития и отмечаемыми фенологическими датами существует определенная связь. Увеличение потребности в необходимом количестве тепла для достижения конкретной стадии ведет к ее большей продолжительности и к более поздним датам наступления фенофазы.
Общей чертой в развитии двух видов ломоноса явилось практически одинаковое количество энергетических ресурсов, необходимых для достижения и прохождения стадии массового созревания семян. Это выразилось также и в близких фенологических датах. На то, что здесь играет роль общий объем соматических ресурсов, указывает пороговая температура, равная 0°C (Ушаков, Недосекина, 2017; Ushakov, Nedosekina, 2017).
На предшествующих стадиях, однако, ломонос прямой имел бóльшую потребность в количестве необходимого для развития тепла в сравнении с ломоносом цельнолистным. При этом у лесного вида предположительно отсутствует остановка в развитии в зимнее время на стадиях начала вегетации и начала бутонизации. Это может быть вызвано тем, что температура, при которой растение находилось зимой, была либо близка к пороговой температуре, либо была ниже нее, но не так сильно и продолжительно, чтобы развитие остановилось. Вероятно, этому способствовали более мягкие и стабильные зимние условия в лесных сообществах, чем на открытых пространствах. В определенной степени это могло быть обусловлено более мощным и постоянным снеговым покровом в лесу. Низкая теплопроводность снега уменьшает суточные колебания температуры поверхности почвы; снег эффективно действует как изолятор земли в самую холодную часть года (Bartlett et al., 2004). Обнаруженное явление для стадии начала бутонизации предполагает, что цветочные почки у ломоноса прямого, по всей видимости, закладывались зимой.
Потребность в бóльшем объеме тепла для развития на ранних стадиях у ломоноса прямого по сравнению с ломоносом цельнолистным скорее всего вызвана необходимостью конкурировать с другими растениями в условиях леса и, следовательно, с увеличением размера. Действительно, по литературным данным (Губанов и др., 2003) ломонос прямой в лесных сообществах достигает высоты 60–150 см, а ломонос цельнолистный, приуроченный к травяным сообществам, 20–70 см.
Проведенные исследования показали, что адаптация к разным условиям существования двух видов растений в ходе эволюции привела к изменениям в физиологии, выражающимся, в том числе, и в изменениях температурных потребностей для развития. Развитие в условиях конкуренции со стороны других видов требует от организма бóльших затрат ресурсов и, следовательно, бóльшего потребления энергии.
Список литературы
Бейдеман И.Н., 1954. Методика фенологических наблюдений при геоботанических исследованиях. М.; Л.: Изд–во АН СССР. 130 с.
Бейдеман И.Н., 1974. Методика изучения фенологии растений и растительных сообществ. Новосибирск: Наука. 155 с.
Борисова И.В.,1972. Сезонная динамика растительного сообщества. Полевая геоботаника. Т. IV. Л.: Наука. С. 5–94.
Губанов И.А., Киселeв К.В., Новиков В.С., Тихомиров В.Н., 2003. Иллюстрированный определитель растений Средней России. Т. 2: Покрытосеменные (двудольные: раздельнолепестные). М.: Т-во науч. Изд. КМК. 665 с.
Серебряков И.Г., 1962. Экологическая морфология растений: жизненные формы покрытосеменных и хвойных. М.: Высш. шк. 378 с.
Ушаков М.В., Недосекина Т.В., 2017. Метод оценки температурных норм развития по данным фенологических наблюдений в природе // Экология. № 5. С. 331–338.
Флора Восточной Европы, 2001. Т. 10. СПб.: Изд-во СПХФА. 670 с.
Шнелле Ф., 1961. Фенология растений. Л.: Гидрометеоиздат. 259 с.
Шульц Г.Э., 1981. Общая фенология. Л.: Наука. 188 с.
Arnold C.Y., 1959. The determination and significance of the base temperature in a linear heat unit system // Proc. American Soc. Horticultural Sci. V. 74. № 4. P. 430–445.
Bartlett M.G., Chapman D.S., Harris R.N., 2004. Snow and the ground temperature record of climate change // J. Geophysical Res. Earth Surface. V. 109. № F04008. P. 1–14.
Bassett I.J., Holmes R.M., MacKay K.H., 1961. Phenology of several plant species at Ottawa, Ontario, and an examination of the influence of air temperatures // Can. J. Plant Sci. V. 41. № 3. P. 643–652.
Blunk H., 1923. Die entwicklung des Dytiscus marginalis L. vom ei bis zur imago. 2. Die Metamorphose // Zeitschrift wissenschaftliche Zool. Bd. 121. S. 121–171.
Campbell A., Fraser B.D., Gilbert N., Gutierrez F.P., Mackauer M., 1974. Temperature requirements of some aphids and their parasites // J. Applied Ecol. V. 11. № 2. P. 431–438.
Johnson M., 2001. The Genus Clematis L. Sweden: Plantskola AB. Södertälje. 896 p.
Miikeda O., Kita K., Handa T., Yukawa T., 2006. Phylogenetic relationships of Clematis (Ranunculaceae) based on chloroplast and nuclear DNA sequences // Botanical J. Linnean Soc. V. 152. № 2. P. 153–168.
Raunkiaer C., 1907. Planterigets Livsformer: og deres Betydning for Geografien. Gyldendalske boghandel. Nordisk forlag. Kjøbenhavn og Kristiania. 132 p.
Sanderson E.D., Pears M.L., 1913. The relation of temperature to insect life // New Hampshire College Agricultural Experiment Station. Technical Bulletin. № 7. 123 p.
Ushakov M.V., Nedosekina T.V., 2017. Method for the evaluation of thermal requirements for development based on phenological observations // Rus. J. Ecol. V. 48. № 5. P. 409–416.
Xie L., Wen J., Li L.-Q., 2011. Phylogenetic analyses of Clematis (Ranunculaceae) based on sequences of nuclear ribosomal ITS and three plastid regions // Systematic Botany. V. 36. № 4. P. 907–921.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии