

Журнал общей биологии, 2019, T. 80, № 3, стр. 226-237
Злаки и разнотравье на степных пастбищах, их токсические свойства и сравнительная роль в питании растительноядных млекопитающих
Б. Д. Абатуров 1, *, А. Е. Скопин 2, **
1 Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский просп., 33, Россия
2 ВНИИ охотничьего хозяйства и звероводства им. проф. Б.М. Житкова
610000 Киров, ул. Преображенская, 79, Россия
* E-mail: abaturovbd@mail.ru
** E-mail: scopin@bk.ru
Поступила в редакцию 02.08.2018
После доработки 17.09.2018
Принята к публикации 28.11.2018
Аннотация
Две группы растений – злаки и разнотравье, образующие основу кормовой массы в пастбищных экосистемах, различаются питательной ценностью, степенью переваримости. К более переваримым относятся представители разнотравья, менее переваримы злаковые травы. Растительноядным млекопитающим – двугорбому верблюду (Camelus bactrianus), сайгаку (Saiga tatarica), американскому бизону (Bison bison), лошади Пржевальского (Equus przewalskii) – свойственна отрицательная связь между долей злаков в потребленном корме и его переваримостью. Низкие и относительно постоянные величины переваримости (45–59%) наблюдаются во все сезоны у лошадей Пржевальского и бизонов, кормовой рацион которых почти полностью состоит из злаков. Более высокая переваримость (до 73%) отмечается у верблюдов и сайгаков – потребителей разнотравья. Для всех видов характерен неодинаковый выбор злаков и разнотравья для питания. Сайгаки и верблюды явное предпочтение отдают более переваримому и питательному разнотравью. Бизоны и лошади Пржевальского целенаправленно выбирают более грубые и менее переваримые злаки. Различия в предпочтении злаков или разнотравья связаны не только с питательностью кормовых растений, но и с их токсичностью. Бизоны и лошади Пржевальского выбирают для питания преимущественно менее питательные, слабопереваримые, но лишенные токсичности злаковые растения и почти полностью избегают потребления обогащенного токсинами разнотравья. Сайгаки и верблюды потребляют токсичное, но более питательное разнотравье и избегают злаков из-за их пониженной переваримости. Однако в этом случае необходимы развитые механизмы устойчивости животных к токсичным метаболитам. В противном случае велик риск отрицательных последствий, который особенно широко распространен в популяциях сайгаков.
Отличительная особенность питания растительноядных млекопитающих – неоднородность и изменчивость питательной ценности (качества) кормовой растительности, что в итоге отражается на непостоянстве пищевой обеспеченности животных и неустойчивости их популяций. Кормовая растительность на природных пастбищах разделяется на две различающиеся по питательной ценности группы – злаки и разнотравье11, которые в зарубежной литературе обозначаются как Грассес и Форбс (Grasses, Forbs) или Грасс и Брауз (Grass, Browse) (Holechek, 1984; Hofmann, 1989, 1991; Gordon, Prins, 2008). Разнотравье превосходит злаки по питательности, в частности листья большинства видов пастбищного разнотравья богаче злаковых растений по содержанию и многообразию синтезированных в растении питательных веществ и вторичных метаболитов в том числе у многих видов – токсичных. Большинство злаков характеризуются повышенной механической прочностью тканей (склеренхимы) и клеточных стенок, что вызвано высоким содержанием лигнина и кремнезема, инкрустирующих клеточную стенку и механические ткани и снижающих их переваримость. По этой причине злаки в отличие от разнотравья относятся к грубым кормам с пониженной переваримостью (Van Soest, Jones, 1968; Holechek, 1984; McNаughton et al., 1985; Абатуров, 2005; Massey et al., 2007; Clauss et al., 2008).
Тем не менее реакция самих пастбищных млекопитающих на отмеченные особенности растительности остается неясной. Известно лишь, что млекопитающие с разной морфологией пищеварительного аппарата неодинаково реагируют на злаковые и разнотравные кормовые растения, что во многом определяет деление растительноядных млекопитающих по типам питания и пищевой специализации. Среди копытных выделяются высокоизбирательные потребители легкоусвояемого разнотравья, менее избирательные потребители преимущественно злаковых растений с более низкой питательной ценностью и большая группа животных с промежуточным питанием (Hofmann, 1991). Повышенная прочность механической ткани у злаков особенно заметно отражается на пищеварении жвачных, функционирование четырехкамерного желудка которых требует максимального измельчения потребленной растительности и дополнительных трат времени и энергии на ее пережевывание. Моногастричные млекопитающие, в частности лошади, для которых характерна заднекишечная ферментация потребленного корма, успешно поедают и даже предпочитают грубые кормовые растения, в том числе злаки (Hofmann, 1991; Duncan, 1992; Menard et al., 2002; Позднякова и др., 2011). Давно замечено, что, например, ковыли – одни из самых распространенных степных злаков – лучше поедаются лошадьми, хуже – овцами и крупным рогатым скотом и хуже всего – верблюдами (Ларин и др., 1950). К последней категории относятся и сайгаки, которые в опытах по кормлению набором разных видов растений, а также в полевых экспериментах со свободной пастьбой всегда избегали поедания не только ковылей, но и злаков вообще (Абатуров и др., 1982, 1998, 2005). Совершенно очевидно, что и требования к качеству кормовых ресурсов у животных с разной спецификой питания неодинаковы, что служит причиной различий в предпочтении определенных кормов и выраженной у большинства травоядных избирательности питания.
Важнейший показатель кормового качества пастбищных растений – переваримость. Не столь важно количество потребленной растительности, как ее усвоенная масса, зависящая от переваримости. Совершенно очевидно, от соотношения количества злаков и разнотравья в потребленной кормовой массе и от способности их усваивать зависит выбор животным злаков или разнотравья. Казалось бы, более грубые и менее переваримые злаки должны уступать разнотравью по участию в питании, однако многие виды животных как жвачные со сложным многокамерным желудком, нуждающиеся в легкопережевываемом (легкоизмельчаемом) корме, так и моногастричные с заднекишечным типом ферментации целенаправленно выбирают для питания именно менее питательные злаки. Возникает необходимость выяснить, насколько ощутима зависимость переваримости корма от состава потребляемой растительности, прежде всего злаков и разнотравья, у видов с разными типами пищеварения, и чем объясняется выбор для питания и предпочтение животными тех или иных по качеству кормов.
СООТНОШЕНИЕ ЗЛАКОВ И РАЗНОТРАВЬЯ В ПИТАНИИ ПАСТБИЩНЫХ МЛЕКОПИТАЮЩИХ
Сравним соотношение злаков и разнотравья в питании разных видов млекопитающих. Для сравнения используем материалы изучения кормовых ресурсов и питания сайгаков (S. tatarica) в северном (Волго-Уральское междуречье) и северо-западном (Калмыкия) Прикаспии, проводимого нами с 1980 года по настоящее время, а также материалы исследований в 2012–2017 годах полувольной группировки крупных копытных, включающей американского бизона (B. bison), лошадь Пржевальского (E. przewalskii), двугорбого верблюда (C. bactrianus) и сайгаков на огороженном степном разнотравно-злаковом пастбище (вольерный комплекс Ассоциации “Живая природа степи”), расположенном в Кумо-Манычской долине в подзоне сухих разнотравно-дерновиннозлаковых степей. Отметим, что во всех случаях оценка состояния (видового состава и надземной массы) пастбищной растительности, видового состава и переваримости потребленного животными корма (рациона) выполнялись сопряженно в одно и то же время (Абатуров и др., 1982, 1998, 2005, 2008, 2015, 2018; Ларионов и др., 2008; Казьмин и др., 2016, 2017). При этом состав и состояние растительности на пастбище оценивали путем учета надземной фитомассы на стандартных укосных площадках; состав потребляемых сайгаками растений и их долю в рационе определяли методом микрогистологического кутикулярно-копрологического анализа непереваренных остатков (экскрементов) животных, а также прямым визуальным подсчетом скусываемых растений ручными сайгаками при их свободной пастьбе; переваримость рациона определяли непрямым методом по соотношению в потребленном корме (в рационе) и фекалиях естественных маркеров – лигнина и органогенного кремния (Абатуров и др., 2003). Отметим, что видовой состав растительности и переваримость пищи определяли на одних и тех же образцах.
Северная часть Прикаспийской низменности – это область сухих степей и пустынь. Зональным типом растительности на ее большей части до конца 1990-х годов были ксерофитно-разнотравные сообщества (Kochia prostrata, Tanacetum achilleifolium, Artemisia pauciflora, A. lerchiana и др.) со значительным участием степных злаков: житняков, ковылей, типчака и др. (Stipa spp., Festuca valesiaca, Agropyron pectiniforme, A. fragile, A. desertorum и др.). В это время в составе фитоценозов неизменно преобладало разнотравье. Еще в середине 1990-х годов (1996 год) на равнинных территориях северного Прикаспия на долю разнотравья приходилось >60% надземной фитомассы, доля злаковых не превышала 40% (Абатуров, Джапова, 2015).
К концу 1990-х годов произошло преобразование растительности, проявившееся в экспансии и абсолютном доминировании в растительном покрове злаковых сообществ. С 1983 по 2012 год в сухих степях Калмыкии многочисленные разнотравные сообщества с преобладанием полыней (A. lerchiana, A. pauciflora), прутняка (K. prostrata) и других растений сменились злаковыми: ковыльными (Stipa spp.), житняковыми (А. fragile), эфемероидными (Poa bulbosa) и однолетниковыми (Anisantha tectorum, Eragrostis minor) (Джапова, 2008; Джапова и др., 2013). По нашим данным, доля массы злаков в растительности Черных земель Калмыкии в 2006 и 2007 годов превысила 90%, тогда как на долю разнотравья большую часть года приходилось <10%. Таким образом, смена растительности привела к резкому уменьшению в фитоценозе массы предпочитаемого сайгаками разнотравья и к увеличению участия злаков, в том числе ковылей, с более низкой питательной ценностью (Ларионов и др., 2008).
Изменение состава надземной фитомассы отразилось на питании пасущихся сайгаков. Рацион сайгаков на равнинных сухостепных территориях Прикаспийской низменности в прошлые годы (1996 год), т.е. до смены растительных сообществ, состоял преимущественно из разнотравья: маревые, сложноцветные, бобовые (K. prostrata, Salsola laricina, Ceratocarpus arenarius, Galatella tatarica, G. villosa, Artemisia spp., Medicago romanica). Доля злаков в рационе была предельно низка и не превышала 1–2% (Абатуров и др., 1982, 1998, 2005). Совсем иная картина наблюдалась после трансформации пастбищной растительности. Замена на равнинах Прикаспийской низменности ксерофильной разнотравно-кустарничковой растительности сообществами с доминированием злаков привела к резкому изменению состава поедаемых растений. Так, в 2006–2007 годах с господством злаков на пастбищах Черных земель Калмыкии в составе рациона сайгаков почти во все сезоны преобладали злаки. Их доля в рационе, как правило, была >50%, в отдельные сезоны достигала 72% (Ларионов и др., 2008). Отметим, что, несмотря на предельно низкое участие разнотравья в надземной фитомассе этих пастбищ (<10%), его доля в рационе лишь немного уступала злакам и колебалась в разные сезоны от 28 до 42%, что свидетельствует о высокой кормовой избирательности сайгаков. Сходное соотношение злаков и разнотравья было свойственно питанию сайгаков на степном пастбище в долине Маныча (2015–2016 годы) при относительно равном участии злаков и разнотравья в пастбищной растительности (доля злаков в фитоценозе в разные сезоны 57–66%). Доля злаков в питании в разные сезоны изменялась в пределах 18–38%, при этом животные заметно избегали их выбора. Таким образом, за весь период исследований (1996–2016 гг.) при всех сменах растительности доля злаков в составе рациона сайгаков изменялась в пределах 1–72%. Соответственно и переваримость рациона варьировала в диапазоне 55–73% (табл. 1).
Таблица 1.
Доля злаков в рационе пасущихся копытных и переваримость потребленного корма на степных пастбищах (по: Абатуров и др., 1998, 2018; Ларионов и др., 2008; Казьмин и др., 2016)
| Животное | Год | n | Доля злаков в рационе, % | Переваримость рациона, % | ||||
|---|---|---|---|---|---|---|---|---|
| min | max | cредняя | min | max | cредняя | |||
| Верблюд | 2015–2016 | 23 | 3 | 91 | 32.9 ± 6.3 | 54 | 72 | 65.2 ± 1.15 |
| Сайгак | 1996–2016 | 30 | 1 | 72 | 31.4 ± 3.8 | 55 | 73 | 61.8 ± 0.76 |
| Лошадь Пржевальского | 2015–2016 | 23 | 69 | 100 | 92.4 ± 1.77 | 53 | 59 | 55.7 ± 0.32 |
| Бизон | 2014–2015 | 7 | 79 | 95 | 86.1 ± 2.1 | 45 | 52 | 49.3 ± 1.08 |
Поскольку злаковые травы обладают более низкой переваримостью по сравнению с разнотравьем, можно ожидать изменения показателей переваримости потребленной растительности в зависимости от соотношения злаков и разнотравья, в частности, от обилия злаковых трав в составе рациона. Действительно, оценка зависимости переваримости корма от количества злаков в потребленном сайгаками корме показала тесную отрицательную связь между этими показателями. Эта связь описывается уравнением регрессии: y = –0.17x + 67.29 при высоком коэффициенте детерминации (R2 = 0.72) (рис. 1).
Рис. 1.
Связь переваримости рациона сайгаков (ось y) с долей злаков (ось x) в составе потребленного корма на степных пастбищах Северного и Северо-западного Прикаспия (Волго-Уральское междуречье, Кумо-Манычская долина) за период 1996–2016 гг. Для рис. 1 – 3 уравнения регрессии для сайгака: y = –0.17x + + 67.29; R2 = 0.72; p < 0.01; n = 30; для верблюда: y = = ‒0.16x + 70.57; R2 = 0.79; p < 0.01; n = 23; для бизона: y = –0.32x + 77.23; R2 = 0.41; p < 0.01; n = 7; для лошади Пржевальского: y = –0.09x + 64.24; R2 = 0.50; p < 0.05; n = 23. Источники данных для рис. 1– 3 в табл. 1.
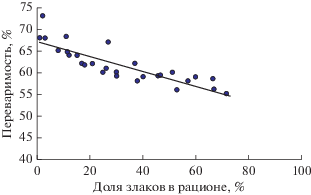
Из уравнения следует, что при увеличении доли злаков в составе рациона пропорционально снижается его переваримость. Следует напомнить, что сайгаки крайне остро реагируют на переваримость корма и, следовательно, на участие злаков в потребляемом корме. Нами было показано ранее, что полноценное функционирование популяции (поддержание жизни, рост, лактация) обеспечивается при переваримости не ниже 68%, а для физиологических нужд на уровне поддержания жизни требуется переваримость выше 59% (Абатуров, 1999; Abaturov, Subbotin, 2011).
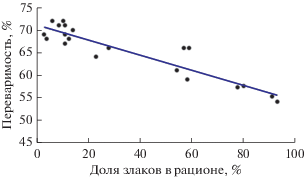
Сходное негативное отношение к злакам свойственно верблюдам. Как и сайгаки, верблюды явно избегают их потребления, хотя они более разнообразны в выборе корма (Абатуров и др., 2018). При относительно близком соотношении злаков и разнотравья в пастбищной растительности (доля злаков в фитоценозе в разные сезоны 30–40%, разнотравья – 60–80%) участие злаков в составе рациона верблюдов в летнее и осеннее время не превышало 50% (в среднем соответственно 11–19%), лишь в короткий ранневесенний период (апрель) было высоким и достигало 90% (в среднем 77%). За весь период исследований (2015–2016 гг.) доля злаков в рационе варьировала в широких пределах – от 3 до 91%, при этом переваримость колебалась от 54 до 72% (табл. 1). Регрессионный анализ показал, что переваримость корма у верблюдов, как и у сайгаков, отрицательно связана с потреблением злаков и меняется пропорционально доле злаков в рационе, при этом описывается уравнением регрессии с очень близкими к сайгаку параметрами: y = –0.16x + 70.57 при столь же высоком уровне связи (R2 = 0.79) (рис. 2).
Рис. 2.
Связь переваримости рациона верблюдов (ось y) с долей злаков (ось x) в составе потребленного корма на степных пастбищах Кумо-Манычской впадины (2015–2016 гг.).
Совсем иное отношение к составу кормовых растений у бизонов и лошадей Пржевальского, пасущихся на тех же пастбищных участках. Несмотря на кардинальные различия в морфологии пищеварительного аппарата и специфике питания, отношение к злакам и разнотравью обоих видов одинаково. Бизоны – типичные представители жвачных копытных – по нашим данным потребляли с высокой избирательностью практически исключительно злаки, на долю которых в составе их рациона приходилось во все сезоны в среднем 86% (Казьмин и др., 2016). При этом диапазон варьирования был узок – 79–95%, соответственно и коэффициент переваримости был низким и относительно неизменным: 45–52% (в среднем 49%) (табл. 1). Поскольку питание бизонов ограничивается преимущественно злаками и однородно по составу, уравнение регрессии y = –0.32x + 77.23 имеет низкий показатель связи переваримости с долей злаков (R2 = 0.41) и заметно отличается от показателей питания верблюдов и сайгаков. Однако весьма важно, что низкие показатели переваримости в данном случае сочетаются с высокой долей злаков в питании бизонов, подтверждая этим зависимость переваримости от обилия злаков в потребленном корме.
Нужно отметить, что абсолютное преобладание злаков в питании – характерное свойство бизонов. В прериях Северной Америки в районах естественного обитания бизонов злаки служат основой их питания. В разных типах прерий злаки всегда преобладали в корме, их доля в составе рациона превышала 90%, соответственно доля разнотравья была <10% (Peden, 1974; Coppedge et al., 1998; Steuter, Hidinger, 1999).
Лошади Пржевальского – нежвачные моногастричные копытные – как и бизоны, явное предпочтение отдают злакам и избегают разнотравья (Позднякова и др., 2011; Казьмин и др., 2017). В нашем случае их питание почти во все сезоны года было предельно однородно, ограничивалось преимущественно злаками, участие которых в рационе варьировало в пределах 69–100%, соответственно и переваримость корма снижена и практически неизменна – 53–59% (в среднем 55.7%) (табл. 1). Связь переваримости с долей злаков в питании описывается уравнением регрессии y = –0.09x + 64.24 с низким коэффициентом детерминации (R2 = 0.50). Как и в случае с бизонами, низкая переваримость потребленного корма у лошадей Пржевальского сочетается с высоким обилием злаков в их рационе, что опять же демонстрирует тесную зависимость переваримости от обилия злаков в потребленном корме. Таким образом, особенности питания как бизонов, так и лошадей Пржевальского, отличные от питания верблюдов и сайгаков, указывают на причинную связь низкой переваримости корма этих животных, вызванную полным доминированием злаков в их питании.
Как видим, каждый отдельный вид копытных по показателям переваримости отрицательно реагирует на обилие злаковых трав в потребляемом корме. Оценим эту связь для объединенной группы животных, образующую единую выборку, независимую от их видовой принадлежности и пищевой специализации. Оказалось, что связь для объединенной выборки имеет тот же линейный характер, общий для всех видов. Примечательно, что абсолютное большинство значений, включающих все отметки для сайгака, верблюда, лошади Пржевальского и основную часть точек для бизона, лежит в 95% области доверительного интервала (рис. 3). Линия регрессии отразила отрицательную зависимость переваримости от обилия злаков в питании и описывается общим уравнением, близким к предыдущим: y = –0.17x + 67.3. Точки, отражающие питание бизонов и лошади Пржевальского, здесь располагаются отдельной группой на общей линии, но в узкой области с наивысшей долей злаков и соответственно низкими величинами переваримости, что, очевидно, отражает видовую специфику питания сравниваемых животных. Распределение точек на диаграмме демонстрирует общий для травоядных характер зависимости величин переваримости от соотношения злаков и разнотравья в их питании и, возможно, независимый от структуры пищеварительного аппарата этих животных (многокамерный желудок животных со жвачным типом питания, моногастричный тип с заднекишечной ферментацией у нежвачных) и их пищевой специализации. Эти данные подтверждают одинаковую для исследованных животных пониженную питательную ценность (переваримость) злаков по сравнению с разнотравьем.
Рис. 3.
Связь переваримости корма с долей злаков в рационе сайгаков, верблюдов, бизонов и лошадей Пржевальского на степных пастбищах Северного и Северо-западного Прикаспия и Кумо-Манычской впадины в 1996–2016 гг. Непрерывная прямая – линия регрессии для всех видов: y = –0.17x + 67.3; R2 = = 0.73; p < 0.01; n = 83. Пунктиром обозначен 95% доверительный интервал.
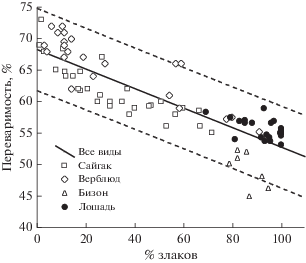
Вместе с тем из этих же данных следует, что многие виды травоядных, в данном случае лошади и бизоны, несмотря на пониженное кормовое качество злаков и более высокую переваримость разнотравья, тем не менее ограничивают питание исключительно злаками и избегают потребления разнотравья. Возникает вопрос о причине столь, казалось бы, нерационального выбора кормовых растений.
ТОКСИЧЕСКИЕ СВОЙСТВА КОРМОВЫХ РАСТЕНИЙ НА СТЕПНЫХ ПАСТБИЩАХ И ИХ РОЛЬ В ПИТАНИИ ЖИВОТНЫХ
Из изложенного выше следует, что две группы растений – злаки и разнотравье – существенно различаются по питательной ценности, в частности по степени переваримости. К более переваримым относятся представители разнотравья с менее прочными, легко измельчаемыми тканями. Менее переваримы злаковые травы с жесткими прочными тканями, требующими больших затрат времени и энергии на их пережевывание и переваривание, что особенно негативно отражается на питании жвачных. Ранее нами было показано, что такие представители жвачных, как сайгаки, именно по этой причине отдают явное предпочтение в питании разнотравью, а при вынужденном преимущественном питании злаками они нежизнеспособны (Абатуров и др., 2005; Абатуров, Джапова, 2015). Тем не менее, как следует из вышеизложенного, другие виды травоядных, в том числе жвачные, явное предпочтение в выборе растений отдают менее переваримым злакам и избегают более питательного разнотравья.
Современные знания о питании растительноядных дают основание полагать, что различия в предпочтении животными злаков и разнотравья связаны не только с обычными показателями питательности растений, но и с таким важным качеством кормовых растений, как их токсичность. В настоящее время хорошо известно, что различные растения в процессе вегетации синтезируют токсичные компоненты, уменьшающие их пригодность для питания (Owen-Smith, 1988). Совершенно очевидно, что животные по-разному реагируют на токсические свойства кормовых растений, с чем связаны их различия в выборе объектов питания (Freeland, Janzen, 1974; Lindroth, 1989; McArthur et al., 1991; Bryant et al., 1992; Скопин, 2003).
Токсичность растений определяется преимущественно химическими соединениями вторичного обмена и используется растениями для защиты от фитофагов и патогенов. Они крайне разнообразны и во многом специфичны для растений определенных систематических групп (Wink, Schimmer, 2009). Кроме того, даже в одних и тех же видах растений состав и концентрации вторичных метаболитов сильно варьируют в зависимости от условий среды, места произрастания, сезонности, жизненности самого растения, степени его дефолиации и прочих факторов (Bohm, 2009). Известно, что повышенная засоленность почв увеличивает токсичность многих растений, что особенно характерно для засушливых, в том числе степных экосистем. В частности, в степях Прикаспийской низменности с характерной для них повышенной почвенной засоленностью отмечена более высокая токсичность растений (Гусынин, 1955).
По химическому строению вторичные метаболиты обычно объединяются в несколько больших групп: алкалоиды, фенольные соединения (фенольные кислоты, фенилпропаноиды, танины и флавоноиды), терпеноиды, гликозиды, органические кислоты, кумарины и пр. (Rosenthal, Berenbaum, 1991). Количество выделяемых соединений в каждой группе необычайно велико и поэтому действие множества вторичных метаболитов в определенном кормовом растении всегда комплексно (Karasov, Martinez del Rio, 2007). Большинство вторичных метаболитов обладает токсичными свойствами. Некоторые соединения (алкалоиды, глюкозинолаты и гликозиды) обладают быстрым токсическим эффектом вследствие ускоренного проникновения в ткани, другие менее мобильные вещества (танины и флавоноиды) действуют медленно и не оказывают острого токсического эффекта. Как правило, высокотоксичные компоненты синтезируются в быстрорастущих тканях растений, а нерастворимые метаболиты – в долгоживущих тканях (Coley et al., 1985). Каждая группа вторичных соединений имеет свои особенности физиологического воздействия на организм животных (Van Soest, 1994). Клиническая картина при отравлениях растениями отличается у разных травоядных (Гусынин, 1955; Garland, Barr, 1998). Для одних видов животных одни и те же растения токсичны, для других нейтральны и пригодны для питания. Например, давно известно, что хвощ полевой (Equisetum arvense) и некоторые виды полыней токсичны для лошадей, но безвредны для овец (Суслов, 1935; Шохор, Будников, 1942). Ряд вторичных токсичных соединений обладает выраженными тератогенными, эстрогенными и галлюциногенными эффектами (Шиманов, 1972; Скопин, 2004; Matsuura, Fett-Neto, 2017).
Во многом выбор кормов для питания зависит от концентрации тех или иных токсичных соединений в растениях. В частности, пороговая для животных концентрация смолы в почках березы равна 50%, и ее превышение полностью ограничивает потребление корма млекопитающими (Sinclair, Smith, 1984). Если в эфедре концентрация эфедрина более 0.5%, то только тогда это растение может быть рассмотрено как токсичное (Vanselow, 2011). Для овец доза растительных танинов начинает проявлять токсичный эффект только при концентрации свыше 1.5 г на кг массы тела (Hervas et al., 2003). Полевки потребляют без последствий токсичные корма только в случае, если концентрация танинов не превышает 6% в рационе (Dietz et al., 1994). Существует важное свойство кормовых растений, в результате которого концентрация токсинов в растениях (танинов, эстрогенов и цианогенов) может увеличиваться в ответ на пресс со стороны травоядных (Levin, 1976).
Большую роль в ограничении токсического действия растительных метаболитов и их детоксикации играют микроорганизмы рубца жвачных. Однако в свою очередь большое количество танинов отрицательно влияет на концентрацию самих микроорганизмов в рубце. В частности, высокая концентрация токсичных соединений в кормовых растениях приводит к гибели рубцовых простейших (Скопин, 2007). Танины ингибируют целлюлозолитические микроорганизмы (Van Soest, 1994; McMahon et al., 2000). Микробиота рубца переводит некоторые безвредные субстанции в токсины. Детоксикация токсинов происходит не только в рубце, но также в печени и слизистой кишечника.
Для защиты от токсичных метаболитов, содержащихся во многих видах кормовых растений, травоядные активно используют литофагию в виде регулярного поедания глины. Содержащиеся в глине минералы группы смектитов, как известно, сильные сорбенты. Они связывают метаболиты и тем самым нейтрализуют их токсичное действие (Паничев, 1990; Barboza et al., 2009). По этой причине поедание глины широко распространено среди различных видов диких и домашних копытных, в частности, по нашим наблюдениям, среди сайгаков, горных баранов, овец. Именно на этом свойстве глины основано фармакологическое действие лекарственного препарата Смекта, широко применяемого в настоящее время в медицинской практике.
Разнотравье и злаки кардинально различаются по токсическим свойствам. Хорошо известно, что разнотравье богато токсичными метаболитами, тогда как в злаках они практически отсутствуют (Owen-Smith, 1988; Скопин, 2003). Поэтому затронутое нами выше объяснение причин различного отношения травоядных к выбору злаков или разнотравья для питания прямо связано с различием токсических свойств этих растений. Как показано выше, многие копытные, в нашем случае бизоны и лошади Пржевальского, выбирают для питания преимущественно менее питательные, слабопереваримые, но зато лишенные токсичности и безопасные злаковые растения и почти полностью избегают потребления обогащенного токсинами разнотравья. Подобные особенности питания характерны не только для копытных, но известны для многих видов грызунов и зайцеобразных. У двух видов полевок (Microtus ochrogaster, M. californicus) и сибирского лемминга (Lemmus sibiricus) потребление легкопереваримых видов разнотравья сопровождалось низким рационом и потерями веса тела, тогда как потребление слабопереваримых злаков компенсировалось повышенным рационом (Batzly, Cole, 1979). Полевки даже при очень высоких плотностях лишь незначительно увеличивали потребление токсичных растений (Batzli, Pitelka, 1975). Такая же картина прослежена при изучении питания полевки Брандта (Lasiopodomys brandti) в Монголии (Сухчулун, Абатуров, 2009). В данном случае оценка связи между количеством потребленного корма и переваримостью показала, что злаки и разнотравье по этим параметрам отчетливо различаются. Для злаков была характерна низкая переваримость и высокий уровень потребления. Если для разнотравья валовое суточное потребление, обеспечивающее усвоение корма на поддерживающем уровне (3.22 г/особь), составляло 5.42 ± 0.45 г/особь, то для злаков этот показатель значительно (в 1.5 раза) больше – 8.27 ± 0.80 г/особь. Тем не менее при питании разнотравьем, несмотря на более высокую переваримость, наблюдался отрицательный баланс массы тела. Полевки потребляли его меньше и не удовлетворяли потребности в корме. Было показано также, что фенольные соединения в растительном корме снижают выживаемость и приводят к задержке роста полевок (Lindroth, Batzli, 1984). Высокий уровень танинов уменьшает потребление корма и вызывает задержку в росте у домашних жвачных, таких как овцы (Min et al., 2003).
Вместе с тем многие дикие и домашние травоядные широко используют или даже предпочитают для питания более питательное, но токсичное разнотравье и, наоборот, избегают злаков из-за их пониженной питательности (переваримости). Однако в этом случае для нейтрализации токсинов часто используется литофагия или необходимо наличие развитых биохимических форм адаптации организма к токсическому действию растительных метаболитов. В противном случае велик риск отрицательных последствий, который особенно широко распространен в популяциях сайгаков в виде частых и многочисленных падежей этих животных. Негативное влияние ядовитых растений на домашних копытных чаще проявляется в степной и пустынной зонах, которым свойственно особенно большое разнообразие растений, обладающих высокотоксичными свойствами (Никитина, Выходцев, 1950; Гусынин, 1955). Характерно, что такие представители типичных степных жвачных, как сайгаки, в выборе кормовых растений абсолютно предпочитают разнотравье, а при питании преимущественно злаками нежизнеспособны (Абатуров и др., 2005). Однако по этой причине они используют литофагию, активно поедая глину с содержащимися в ней активными минеральными сорбентами.
Были высказаны предположения, что интенсивное поедание разнотравья животными в годы их высокой численности индуцирует синтез в растениях токсичных метаболитов, выполняющих защитные функции и ограничивающих потребление таких растений животными (Bryant, 1981; Batzly, 1983; Bucyanayandi, Bergeron, 1990, и др.). Однако эти соображения пока во многом гипотетичны. Несмотря на множество исследований, все еще невозможно сделать точных экологических заключений о роли токсичных метаболитов в выборе кормовых растений травоядными (Bryant et al., 1992; Karasov, Martinez del Rio, 2007). Практически отсутствуют прямые свидетельства зависимости выбора корма от уровня его токсичности у диких травоядных млекопитающих.
Все это отражает слабую изученность роли токсичных свойств пастбищной растительности в обеспеченности пищей и питании травоядных животных. Совершенно очевидно, что в настоящее время возникла необходимость проведения специальных исследований токсических функций метаболитов растений в питании и освоении растительных кормовых ресурсов травоядными животными в природных пастбищных экосистемах.
ЗАКЛЮЧЕНИЕ
Две группы растений – злаки и разнотравье, составляющие основу кормовой растительной массы в травяных пастбищных экосистемах, – существенно различаются по питательной ценности, в частности по степени переваримости. К более переваримым относятся представители разнотравья с непрочными легко измельчаемыми тканями. Менее переваримы злаковые травы с жесткими прочными тканями, требующими больших затрат времени и энергии на их пережевывание и переваривание, что особенно негативно отражается на питании жвачных. Очевидно, что от соотношения злаков и разнотравья в составе рациона зависит переваримость потребленного корма. Действительно, для питания дикой популяции сайгаков в разные временные отрезки с 1996 по 2016 год, отличавшиеся изменчивым соотношением злаков и разнотравья в пастбищной (кормовой) растительности, свойственна отчетливая отрицательная связь между долей злаков в потребленном корме и переваримостью пищи. Диапазон переваримости варьировал от 55 до 73% при изменении доли злаков в питании от 1 до 72%. Близкая по параметрам отрицательная связь свойственна верблюдам, у которых при изменении участия злаков в питании в диапазоне 3–91% переваримость пропорционально менялась в пределах 54–72%. У лошадей Пржевальского и бизонов, облигатных потребителей злаков, в питании которых абсолютно преобладали злаки (соответственно 69–100% и 79–95%), переваримость корма заметно снижена и однородна (соответственно 53–59% и 45–52%), что отражает отрицательное влияние злаков на переваримость потребленного корма.
Объединение всех исследованных видов в одну группу продемонстрировало общую отрицательную зависимость величин переваримости от обилия злаков в их питании, практически не зависящую от структуры пищеварительного аппарата этих животных и их пищевой специализации. Эти данные подтверждают одинаковую для травоядных животных пониженную питательную ценность (переваримость) злаков по сравнению с разнотравьем. Переваримость корма у четырех исследованных видов копытных (верблюды, сайгаки, бизоны и лошади Пржевальского), различающихся типами пищеварения и кормовой специализацией, практически в одинаковой мере отрицательно связана с долей злаков в их питании.
От соотношения количества злаков и разнотравья в потребленной кормовой массе и от способности их усваивать зависит не только переваримость пищи, но и выбор животными для питания злаков или разнотравья. Такие представители жвачных, как сайгаки и верблюды, явное предпочтение в питании отдают более переваримому и питательному разнотравью. В то же время бизоны, относящиеся к жвачным, и лошади Пржевальского – представители моногастричных с заднекишечным типом ферментации – целенаправленно и, казалось бы, нерационально выбирают более грубые и менее переваримые злаки.
Современные знания о питании растительноядных дают основание полагать, что различия в предпочтении теми или иными животными злаков или разнотравья связаны не только с обычными показателями питательности растений, но и с таким важным качеством кормовых растений, как их токсичность. Известно, что разнотравье богато токсичными метаболитами, тогда как в злаках они практически отсутствуют. Поэтому различное отношение травоядных к выбору злаков или разнотравья для питания непосредственно связано с неодинаковой реакцией этих животных на токсичность кормовых растений. Многие копытные, в нашем случае бизоны и лошади Пржевальского, выбирают для питания преимущественно менее питательные, слабопереваримые, но зато лишенные токсичности безопасные злаковые растения и почти полностью избегают потребления обогащенного токсичными метаболитами разнотравья. Вместе с тем многие дикие и домашние травоядные, в нашем случае сайгаки и верблюды, предпочитают для питания более питательное, но токсичное разнотравье и, наоборот, избегают лишенных токсинов злаков из-за их пониженной питательности (переваримости). Однако в этом случае для исключения токсического действия метаболитов необходимо наличие развитых форм адаптации организма к токсинам или их нейтрализации, в том числе широко распространенной среди растительноядных млекопиткающих литофагии. В противном случае велик риск отрицательных последствий, который особенно широко распространен в популяциях сайгаков в виде частых и многочисленных падежей.
Работа выполнена при поддержке РФФИ № 18-04-00172.
Список литературы
Абатуров Б.Д., 1999. Критические параметры качества растительных кормов для сайгаков (Saiga tatariса) на естественном пастбище в полупустыне // Зоол. журн. Т. 78. № 8. С. 999–1010.
Абатуров Б.Д., 2005. Кормовые ресурсы, обеспеченность пищей и жизнеспособность популяций растительноядных млекопитающих // Зоол. журн. Т. 84. № 10. С. 1251–1271.
Абатуров Б.Д., Джапова Р.Р., 2015. Кормовая обеспеченность и состояние сайгаков (Saiga tatarica) на степных пастбищах с разным соотношением злаков и разнотравья // Изв. РАН. Сер. биол. № 2. С. 207–214.
Абатуров Б.Д., Холодова М.В., Субботин А.Е., 1982. Интенсивность питания и переваримость кормов у сайгаков // Зоол. журн. Т. 61. № 12. С. 1870–1881.
Абатуров Б.Д., Петрищев Б.И., Колесников М.П., Субботин А.Е., 1998. Сезонная динамика кормовых ресурсов и питание сайгака на естественном пастбище в полупустыне // Успехи соврем. биологии. Т. 118. Вып. 5. С. 564–584.
Абатуров Б.Д., Колесников М.П., Никонова О.А., Позднякова М.К., 2003. Опыт количественной оценки питания свободнопасущихся млекопитающих в естественной среде обитания // Зоол. журн. Т. 82. № 1. С.104–114.
Абатуров Б.Д., Ларионов К.О., Колесников М.К., Никонова О.А., 2005. Состояние и обеспеченность сайгаков кормом на пастбищах с растительностью разных типов // Зоол. журн. Т. 84. № 3. С. 377–390.
Абатуров Б.Д., Ларионов К.О., Джапова Р.Р., Колесников М.П., 2008. Качество кормов и обеспеченность сайгаков (Saiga tatarica) пищей в условиях восстановительной смены растительности на Черных землях Калмыкии // Зоол. журн. Т. 87. № 12. С. 1524–1530.
Абатуров Б.Д., Казьмин В.Д., Колесников М.П., 2015. Питание бизонов (Bison bison), верблюдов (Camelus bactrianus) и лошадей (Equus caballus) при совместной пастьбе на изолированном степном пастбище // Зоол. журн. Т. 94. № 12. С. 1470–1478.
Абатуров Б.Д., Казьмин В.Д., Джапова Р.Р., Аюшева Е.Ч., Джапова В.В., Нохаева Д.В., Колесников М.П., Миноранский В.А., Кузнецов Ю.Е., 2018. Кормовые ресурсы, питание и обеспеченность пищей свободно пасущихся верблюдов (Camelus bactrianus) на пастбищах степной природной зоны // Зоол. журн. Т. 97. № 3. С. 348–361.
Гусынин И.А., 1955. Токсикология ядовитых растений. М.: Сельхозгиз. 330 с.
Джапова Р.Р., 2008. Динамика пастбищ и сенокосов Калмыкии. Элиста: Изд-во Калмыцк. ун-та. 176 с.
Джапова Р.Р., Васькина Н.А., Халгинова Б.В., 2013. Антропогенная динамика растительного покрова в пустынной зоне на территории Республики Калмыкия // Современная ботаника в России. Т. 2. Тольятти: Кассандра. С. 203–205.
Казьмин В.Д., Абатуров Б.Д., Демина О.Н., Колесников М.П., 2016. Кормовые ресурсы и питание полувольных бизонов (Bison bison) на степном пастбище долины западного Маныча // Зоол. журн. Т. 95. № 2. С. 234–244.
Казьмин В.Д., Абатуров Б.Д., Джапова Р.Р., Аюшева Е.Ч., 2017. Пищевая адаптация лошади Пржевальского к условиям обитания в степях долины Маныча // Пространственно-временная динамика биоты и экосистем Арало-Каспийского бассейна. Оренбург: ИПК “Университет”. С. 212–217.
Ларин И.В., Агагабян Ш.М., Работнов Т.А. и др., 1950. Кормовые растения сенокосов и пастбищ СССР. Т. I. / Под ред. Ларина И.В. М.; Л.: Гос. изд-во с.-х. лит-ры. 688 с.
Ларионов К.О., Джапова Р.Р., Розенфельд С.Л., Абатуров Б.Д., 2008. Питание сайгаков (Saiga tatarica) на пастбищах Черных земель Калмыкии в условиях восстановительной смены растительности и остепнения // Зоол. журн. Т. 87. № 10. С. 1259–1269.
Никитина Е., Выходцев И., 1950. Ядовитые и вредные растения пастбищ и сенокосов Киргизии. Фрунзе: Изд-во Кирг. АН СССР. 100 с.
Паничев А.М., 1990. Литофагия в мире животных и человека. М.: Наука. 224 с.
Позднякова М.К., Жарких Т.Л., Ясинецкая Н.И., Колесников М.П., 2011. Количественная оценка питания полувольной группировки лошади Пржевальского (Equus przewalskii) в степном местообитании (заповедник “Аскания-Нова”) // Зоол. журн. Т. 90. № 3. С. 368–376.
Скопин А.Е., 2003. Значение компонентов качественного состава кормовых растений диких животных. Киров: ВНИИОЗ. 203 с.
Скопин А.Е., 2004. Биологически активные соединения галлюциногенных макромицетов России // Селекция против наркотиков. Краснодар: КНИИСХ РАСХН. С. 104–110.
Скопин А.Е., 2007. Антипитательные свойства кормовых древесных растений // Вопросы физиологии, содержания, кормопроизводства и кормления, селекции с/х животных, биологии пушных зверей и птиц, охотоведения. Киров: ВГСХА. С. 209–213.
Суслов А.Ф., 1935. Отравление хвощем полевым // Коневодство. № 6. С. 52–53.
Сухчулуун Г., Абатуров Б.Д., 2007.Зависимость питания и обеспеченности кормом полевки Брандта (Lasiopodomys brandti) от особенностей растительности // Зоол. журн. Т. 86. № 8. С. 999–1007.
Шиманов В.Г., 1972. Гормональная активность пастбищных растений и влияние их на плодовитость каракульских овец. Ташкент: ФАН. 244 с.
Шохор А.Н., Будников П.И., 1942. Материалы по изучению ядовитости полыни // Тр. Вет. ин-та Красной Армии и Окружной военно-вет. лаб. № 4. С. 203–207.
Abaturov B.D., Subbotin A.E., 2011. Forage quality thresholds for Saiga antelope in a semideserts rangeland // Rus. J. Theriol. V. 10. № 2. P. 71–81.
Barboza P.S., Parker K.L., Hume I.D., 2009. Integrative Wildlife Nutrition. Berlin – Heidelberg: Springer-Verlag. 342 p.
Batzli G.O., 1983. Responses of arctic rodent populations to nutritional factors // Oikos. V. 40. P. 396–406.
Batzli G.O., Cole F.R., 1979. Nutritional ecology of microtine rodents: Digestibility of forage // J. Mammal. V. 60. P. 740–750.
Batzli G.O., Pitelka F.A., 1975. Vole cycles: test of another hypothesis // Am. Nat. V. 109. P. 482–487.
Bohm B.A., 2009. The Geography of Phytochemical Races. N.Y.: Springer Publ. 348 p.
Bryant J.P., 1981. Phytochemical deterrence of snowshoe hare browsing by adventitious shoots of four Alaskan trees // Science. V. 213. P. 889–890.
Bryant J.P., Reichardt P.B., Clausen T. P., Provenza F.D., Kuropat P.J., 1992. Woody plant–mammal interactions // Herbivores. Their interaction with secondary plant metabolites. San-Diego; N.Y.: Acad. Press. V. 2. P. 343–370.
Bucyanayandi J.-D., Bergeron J.-M., 1990. Effects of food quality on feeding patterns of meadow voles (Microtus pennsylvanicus) along a community gradient // J. Mammalogy. V. 71. № 3. P. 390–396.
Clauss M., Kaiser T., Hummel J., 2008. The morphophysiological adaptations of browsing and grazing mammals // Ecol. Browsing Grazing / Eds Gordon I.J., Prins H.H.T. B.: Springer-Verlag. P. 47–88.
Coley P.D., Bryant J.P., Chapin III F.S., 1985. Resource availability and plant antiherbivore defense // Science. V. 230. P. 895–899.
Coppedge B.R., Leslie D.M., Jr., Shaw J.H., 1998. Botanical composition of bison diets on tallgrass prairie in Oklahoma // J. Range Management. V. 51. № 4. P. 379–382.
Dietz B.A., Hagerman A.E., Barret G.W., 1994. Role of condensed tannin on salivary tanninbinding proteins, bioenergetics and nitrogen digestibility in Microtus pennsylvanicus // J. Mammalogy. V. 75. P. 880–889.
Duncan P., 1992. Horses and Grasses. The Nutritional Ecology of Equids and Their Impact on the Camargue. N.Y.: Springer-Verlag. 287 p.
Freeland W.J., Janzen D.H., 1974. Strategies in herbivory by mammals: The role of plant secondary compounds // Am. Nat. V. 108. P. 269–289.
Garland T., Barr A.C. (eds). 1998. Toxic Plants and Other Natural Toxicants. Oxon-New-York: CABI Publishing. 585 p.
Gordon I.J., Prins H.H.T. (eds). 2008. The Ecology of Browsing and Grazing. Berlin, Heidelberg: Springer. 330 p.
Hervas G., Perez V., Giraldez F.J., Mantecon A.R., Almar M.M., Frutos P., 2003. Intoxication of sheep with quebracho tannin extract // J. Comparative Pathology. V. 129. P. 44–54.
Hofmann R.R., 1989. Evolutionary steps of ecophysiological adaptation and diversification of ruminants: A comparative view of their digestive system // Oecologia. V. 78. P. 443–457.
Hofmann R.R., 1991. Die Wiederkauer // Biol. Unserer Zeit. V. 21. № 2. S. 73–80.
Holechek J.L., 1984. Comparative contribution of Grasses, Forbs, and Shrubs to the nutrition of range ungulates // Rangelands. V. 6. № 6. P. 261–263.
Karasov W.H., Martinez del Rio C., 2007. Physiological Ecology: How Animals Process Energy, Nutrients, and Toxins. Princeton; Oxford: Princeton Univ. Press. 741 p.
Levin D.A., 1976. Alkaloid-bearing plants: An ecogeographic perspective // Am. Nat. V. 110. № 972. P. 261–284.
Lindroth R.L., 1989. Mammalian herbivore-plant interactions // Plant-Animal Interactions / Ed. Abrahamson W.G. N.Y.: McGraw Hill B.C. P. 163–206.
Lindroth R.L., Batzli G.O., 1984. Plant phenolic as chemical defenses: Effects of natural phenolics on survival and growth of prairie voles (Microtus ochrogaster) // J. Chemical Ecol. V. 10. P. 229–244.
Massey F.P., Ennos A.R., Hartley S.E., 2007. Grasses and the resource availability hypothesis: The importance of silica-based defenses // J. Ecology. V. 95. № 3. P. 414–424.
Matsuura H.N., Fett-Neto A.G., 2017. Plant alkaloids: Main features, toxicity, and mechanisms of action // Plant Toxins / Eds Gopalakrishnakone P., Carlini C. et al. Dordrecht: Springer Science Business Media. P. 243–261.
McArthur C., Hagerman A.E., Robbins C.T., 1991. Physiological strategies of mammalian herbivores against plant defenses // Plant Defenses against Mammalian Herbivory / Eds Palo R.T., Robbins C.T. Boca Raton: CRC Press. P. 103–114.
McMahon L.R., McAllister T.A., Berg B.P., Majak W., Acharya S.N., Popp J.D., Coulman B.E., Wang Y., Cheng K.-J., 2000. A review of the effects of forage condensed tannins on ruminal fermentation and bloat in grazing cattle // Can. J. Plant Science. V. 80. P. 469–485.
McNaughton S.J., Tarrants J.L., McNaughton M.M., Davis R.H., 1985. Silica as a defense against herbivory and a growth promotor in African grasses // Ecology. V. 66. P. 528–535.
Menard C., Duncan P., Fleurance G. et al., 2002. Comparative foraging and nutrition of horses and cattle in European wetlands // J. Applied Ecol. V. 39. P. 120–133.
Min B.R., Barry T.N., Attwood G.T., McNabb W.C., 2003. The effect of condensed tannins on the nutrition and health of ruminants fed fresh temperate forages: a review // Animal Feed Science Technology. V. 106. P. 3–19.
Owen-Smith R.N., 1988. Megaherbivores: The Influence of Very Large Body Size on Ecology. Cambridge: Cambridge Univ. Press. 369 p.
Peden D.G., Van Dyne G.M., Rice R.W., Hansen R.M., 1974. The trophic ecology of Bison bison L. on the shortgrass plains // J. Applied Ecol. V. 11. P. 489–497.
Rosenthal G.A., Berenbaum M.R. (eds). 1991. Herbivores: Their Interaction with Secondary Plant Metabolites. San-Diego; N.Y.: Acad. Press. V. 1. 468 p.
Sinclair A.R.E., Smith J.N.M., 1984. Do plant secondary compounds determine feeding preferences of snowshoe hares? // Oecologia. V. 61. P. 403–410
Steuter A., Hidinger L., 1999. Comparative ecology of bison and cattle on mixed-grass prairie // Great Plains Research. V. 9. № 2. P. 329–342.
Van Soest P.J., 1994. Nutritional Ecology of the Ruminant. Ithaca; London: Cornell Univ. Press. 476 p.
Van Soest P.J., Jones L.H.P., 1968. Effect of silica in forages upon digestibility // J. Dairy Science. V. 51. № 10. P. 1644–1648.
Vanselow K.A., 2011. The high-mountain pastures of the Eastern Pamirs (Tajikistan). Dissertation. Nürnberg: Friedrich-Alexander-Universität. 234 p.
Wink M., Schimmer O., 2009. Molecular modes of action of defensive secondary metabolites // Annual Plant Reviews. V. 39. P. 21–161.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии