

Журнал общей биологии, 2019, T. 80, № 4, стр. 286-296
Факультативное охотничье поведение грызунов как возможная эволюционно стабильная стратегия
С. Н. Пантелеева 1, 2, *, Я. В. Левенец 1, Ж. И. Резникова 1, 2
1 Институт систематики и экологии животных СО РАН
630091 Новосибирск, ул. Фрунзе, 11, Россия
2 Новосибирский национальный исследовательский государственный
630090 Новосибирск, ул. Пирогова, 1, Россия
* E-mail: psofia@mail.ru
Поступила в редакцию 06.09.2018
После доработки 04.02.2019
Принята к публикации 11.04.2019
Аннотация
У трех видов грызунов с разными типами питания впервые выявлено и детально описано целенаправленное взаимодействие с подвижной добычей по типу “хищник–жертва”. Зерноядная полевая мышь, зеленоядная узкочерепная полевка и всеядный хомячок Кэмпбелла обладают одинаково эффективным стереотипным охотничьим поведением, во многом сходным с поведением специализированного насекомоядного вида – обыкновенной бурозубки. При этом скорость охоты у грызунов ниже, чем у насекомоядных. У грызунов, в отличие от насекомоядного вида, стереотип охотничьего поведения проявляется факультативно – полностью, но не у всех особей. Доля “охотников” у узкочерепных полевок вдвое меньше, чем у полевых мышей. У разных видов различаются тактики умерщвления добычи: полевые мыши, узкочерепные полевки и бурозубки обездвиживают насекомое серией быстрых укусов; хомячки Кэмпбелла откусывают у добычи конечности, что, по-видимому, является проявлением более специализированного охотничьего поведения. Характер охотничьих атак различен: грызуны, вначале схватывая добычу зубами, переходят к захвату лапами, а бурозубки используют только зубы, что говорит об относительной примитивности их хищнического поведения. Хомячки Кэмпбелла могут начать атаку с захвата как зубами, так и лапами, что характеризует их охотничье поведение как наиболее эволюционно продвинутое среди исследованных видов. Стереотипы охотничьего поведения у всех трех видов грызунов проявляются по принципу “все и сразу” и не подвержены влиянию опыта. Охотничье поведение грызунов можно рассматривать как эволюционно стабильную стратегию, поддерживающую способность популяций использовать охоту на подвижных насекомых для расширения спектра пищевых ресурсов.
Грызуны – разнообразная и процветающая группа, составляющая около 40% млекопитающих по числу видов и обладающая, помимо морфологических и физиологических особенностей, целым рядом поведенческих адаптаций. Способности грызунов к выбору оптимальной диеты и переключению на новые источники пищи в меняющейся среде до сих пор исследовались главным образом для зеленоядных видов по отношению к разным видам растений (Soininen et al., 2013), а также для всеядных видов с широкими преференциями. Так, у серых крыс Rattus norvegicus Berkenhout, обитающих в высоко изменчивой среде, проявления неофобии по отношению к новым запахам пищи минимальны (Modlinska, Stryjek, 2016). Почти все виды грызунов являются всеядными в той или иной мере (Landry, 1970), и для многих из них отмечено включение в рацион беспозвоночных и даже мелких позвоночных (Левенец и др., 2016). Однако сведения о животной пище грызунов получены на основе анализа желудков и фекалий (Левенец и др., 2016). Наблюдения за поимкой добычи производились у грызунов, отловленных в естественных условиях и помещенных в лабораторию (Rowe A., Rowe M., 2006), а отдельные наблюдения за охотой на беспозвоночных (саранчовые и бабочки) в природе известны лишь для хомячка Кэмпбелла (Левенец и др., 2019). Процесс охоты на подвижную добычу до недавнего времени исследовался лишь на примере нескольких видов: хищного кузнечикового хомячка Onychomys torridus Coues, всеядных белоногого и оленьего хомячков Peromyscus leucopus Rafinesque и P. maniculatus Wagner (Kreiter, Timberlake, 1988; Timberlake, Washburne, 1989), сирийского хомячка Mesocricetus auratus Waterhouse (Langley, 1986; Polsky, 1977), а также серой крысы R. norvegicus (Haug, Johnson, 1991). На примере хомячков была выявлена роль индивидуальных навыков: специализированные кузнечиковые хомячки с первой встречи с потенциальной жертвой проявляют во всех деталях врожденное охотничье поведение, тогда как у белоногого и оленьего хомячков эффективное взаимодействие с добычей требует накопления опыта (Kreiter, Timberlake, 1988). Это значит, что у разных видов грызунов координация моторных актов при охотничьем поведении управляется генетическими программами в различной степени. Эти работы положили начало исследованию различных адаптаций у грызунов при охоте на подвижную добычу. Было выяснено, что представители рода кузнечиковых хомячков Onychomys являются специализированными охотниками, которые убивают беспозвоночных и мелких позвоночных животных как в естественных, так и в лабораторных условиях и обладают морфологическими и физиологическими адаптациями к взаимодействию с подвижной добычей, в том числе такой опасной, как скорпионы (Sarko et al., 2011). Охотничье поведение лабораторных линий домовой мыши по отношению к сверчкам в последние годы используется как модель для изучения перцептивных (Hoy et al., 2016) и нейрологических (Han et al., 2017) механизмов взаимодействия грызуна и добычи. Недавно обнаруженное в лабораторных экспериментах охотничье поведение по отношению к насекомым у полевых мышей Apodemus agrarius Pallas (Panteleeva et al., 2013) и рыжих полевок Myodes glareolus Schreber (Konczal et al., 2016), не имеющих, так же как и домовые мыши, морфологических или физиологических адаптаций к охоте, позволяет предположить наличие у грызунов с разными типами питания поведенческих адаптаций к добыванию подвижных беспозвоночных. Возникает вопрос о выявлении у грызунов специфических стереотипов поведения, использование которых может расширять адаптивный потенциал вида за счет возможности переключения на подвижную добычу. Под стереотипом мы понимаем поведенческую последовательность, состоящую из устойчиво повторяющихся элементов (Пантелеева и др., 2010).
В наших исследованиях впервые было выявлено постоянное взаимодействие мелких млекопитающих с рыжими лесными муравьями в естественных условиях (Пантелеева и др., 2016). Это дало толчок к лабораторным экспериментам, в которых была впервые выявлена активная охота полевых мышей A. agrarius на подвижных насекомых, в том числе таких многочисленных, агрессивных и опасных, как муравьи (Panteleeva et al., 2013; Reznikova et al., 2017). Показано, что охотничье поведение полевых мышей организовано в стереотип, включающий в себя обнаружение, преследование, атаку и обработку добычи. Атака, как часть охотничьего стереотипа, включает бросок на добычу и ее схватывание и может рассматриваться как фиксированный комплекс действий (ФКД) (Дьюсбери, 1981; Зорина и др., 2013), т.е. является видоспецифическим, врожденным и шаблонным комплексом двигательных актов. В этологии поведенческий акт рассматривается как врожденный, если он проявляется с достаточной полнотой с первого раза (Дьюсбери, 1981). Наши эксперименты (Reznikova et al., 2017) показали, что целостный охотничий стереотип проявляется у молодых “наивных” (т.е. выращенных вне контакта с соответствующими стимулами) животных полностью и без предварительного опыта, иными словами, по принципу “все и сразу” (Резникова, 2005) и по эффективности сравним с соответствующими стереотипами специализированных плотоядных видов. Важно отметить, что целостный врожденный стереотип охотничьего поведения у полевой мыши носит факультативный характер, т.е. проявляется полностью, но не у всех животных.
Мы предположили, что наличие носителей охотничьего стереотипа в популяциях нехищных видов грызунов является эволюционно-стабильной стратегией (Maynard Smith, Price, 1973), позволяющей популяциям расширять пищевой спектр за счет подвижных насекомых. Теория эволюционно стабильных стратегий (ЭСС) была предложена Мэйнардом Смитом и Прайсом (Maynard Smith, Price, 1973) для объяснения сосуществования в популяциях групп индивидуумов с альтернативными стратегиями разрешения конфликтов (классическим примером являются стратегии “голубя” и “ястреба”). ЭСС – это такая стратегия, которая, будучи принята достаточно большим количеством особей в популяции, не может быть вытеснена другой стратегией, т.е. никакая другая стратегия не может обеспечить более высокой приспособленности. При изменении внешних условий соотношение представителей альтернативных стратегий в популяции может меняться, но стратегии остаются эволюционно стабильными. Под стратегией понимается свод правил, определяющих, какой из альтернативных поведенческих паттернов будет принят индивидуумом в любой ситуации в течение жизни. Каждый индивидуум может принять только одну стратегию, и разные стратегии предполагают соответствующие различия в генотипах. В отличие от стратегии, тактика – это использование одного из существующих фенотипических вариантов поведенческих паттернов. Теория ЭСС, по свидетельству ее авторов, была частично основана на идеях теории игр и частично – на работах МакАртура (MacArthur, 1965) и Гамильтона (Hamilton, 1967) об эволюции соотношения полов. Это предопределило сравнительно широкое использование модели ЭСС, в частности для описания стратегий спаривания (Dominey, 1984) и фуражировочного поведения в условиях внутривидовой конкуренции (Sirot, 2000). Сходные с ЭСС идеи, основанные на теории игр, лежат в основе исследования взаимодействия хищника и жертвы на примере ястреба-перепелятника и травника (Quinn, Cresswell, 2004). Лабильное соотношение в популяциях групп индивидуумов с различными стратегиями питания с позиции ЭСС до сих пор не обсуждалось.
Для проверки гипотезы о наличии “охотников” в популяциях грызунов с различными типами питания как проявления эволюционно стабильной стратегии, мы в качестве первого шага исследовали стереотипы охотничьего поведения у трех видов грызунов в сравнении с насекомоядным видом (обыкновенной бурозубкой) как “эталоном хищника”. Нас интересовали такие показатели, как стабильность стереотипа охотничьего поведения, его видовая специфичность и доля “охотников” в исследуемых группах грызунов, отражающая ситуацию в природных популяциях. Стабильность и специфичность стереотипа позволяют считать его проявлением именно стратегии, а не тактики индивидуумов.
МАТЕРИАЛЫ И МЕТОДЫ
Лабораторные исследования проводили в 2014–2018 гг. на трех видах грызунов с различными типами питания и структуры популяций: зерноядной полевой мыши A. agrarius (n = 26; 13 самцов и 13 самок), зеленоядной узкочерепной полевке Lasiopodomys gregalis Pallas (n = 46; 23 самца и 23 самки) и эврифаге – хомячке Кэмпбелла Phodopus campbelli Thomas (n = 19; 8 самцов и 11 самок). В родстве друг с другом животные каждого вида не состояли. Согласно классификации В.С. Громова (2008), полевые мыши относятся к видам с системой агрегаций индивидуальных участков обитания, узкочерепные полевки живут слабо консолидированными семейными группами или колониями, а хомячки Кэмпбелла занимают обособленные индивидуальные участки. Из 26 полевых мышей 17 особей были отловлены на территории Приобской лесостепной провинции в 30 км от г. Новосибирска в естественных условиях, 9 являлись потомками второго поколения ранее отловленных животных. Все узкочерепные полевки были отловлены на территории Карасукского стационара ИСиЭЖ СО РАН (Новосибирская обл.). Аутбредные хомячки Кэмпбелла (n = 19) предоставлены виварием ИЦиГ СО РАН, где они содержались на протяжении 20 поколений. Все особи хомячка Кэмпбелла и 9 полевых мышей, рожденные в виварии, ранее не контактировали с насекомыми, т.е. являлись “наивными” по отношению к потенциальной добыче. Для сравнения исследовано поведение обыкновенной бурозубки Sorex araneus Linnaeus, взятой из естественных условий (n = 11). В качестве подвижной безопасной добычи использовали мраморных тараканов Nauphoeta cinerea Olivier (средняя длина тела равна 27.93 ± 0.4 мм). В 2018 г. провели дополнительные эксперименты с хомячками Кэмпбелла (n = 10), предлагая им в разных сериях тестов не только безопасных насекомых, но и опасных – рыжих лесных муравьев Formica aquilonia Yarr., предъявляемых на аренах группами по 10 особей (по аналогии с: Panteleeva et al., 2013). Все животные содержались в индивидуальных клетках, имели постоянный доступ к воде и пище без ограничений. Рацион включал зерновые смеси, фрукты и овощи, а также белковые компоненты (творог, вареные яйца).
Для исследования охотничьего поведения животным в прозрачных аренах 30 × 30 × 35 см предлагалась добыча и производилась видеозапись их реакций (всего около 100 ч наблюдений). Тем особям, которые охотились при первом предъявлении насекомого, в пределах одного теста последовательно предлагали еще до двух единиц добычи. Тестирование длилось до завершения поедания жертвы либо, если животное не проявляло интерес к добыче, прекращалось через 10 мин. Таким образом, в зависимости от реакций зверька, тест мог закончиться на первой, второй или третьей единице добычи. Первую реакцию на добычу можно было наблюдать для каждого животного только однажды: при первом предъявлении в первом тесте. Для получения сопоставимого количества поведенческих последовательностей полевые мыши и обыкновенные бурозубки тестировались по 2 раза, узкочерепные полевки – по 3, а хомячки Кэмпбелла – по 7 раз. С помощью программы Noldus Observer XT 10.1 были выявлены и проанализированы последовательности элементов поведения, полученные во всех тестах (подробно о методе анализа см.: Reznikova et al., 2017). Это позволило сопоставить характер и тактику охотничьего поведения у разных видов. Результативность охоты оценивалась как соотношение количества успешных атак, заканчивающихся поимкой добычи, и неудачных атак, когда добыча была упущена, и животное прекращало активный поиск. Скорость охоты оценивалась как количество элементов поведения, совершенных в секунду. Детально анализировались только акты успешной охоты, закончившиеся поимкой и поеданием добычи, но фиксировались также и все случаи неудачных атак. В каждом тесте животное могло продемонстрировать до трех успешных атак и неограниченное количество неудачных. Сравнения долей охотившихся и не охотившихся особей у разных видов, а также успешных и неудачных атак у разных особей, проводились с помощью точного теста Фишера с поправкой Бонферрони. Сравнение количества неудачных атак, предшествовавших успешным, у разных видов проведено с помощью критерия Манна–Уитни. Для построения схемы стереотипов охотничьего поведения рассчитывали матрицы вероятностей перехода от одного поведенческого элемента к другому (Марковский процесс первого порядка) (Casarrubea et al., 2008). Сравнение скорости охоты и количества элементов в стереотипах проведено с помощью H-критерия Краскела–Уоллиса с поправкой Бонферрони. При сравнении представлены медиана, первый и третий квартили: Me (Q1–Q3).
РЕЗУЛЬТАТЫ
Все исследованные виды продемонстрировали целенаправленное взаимодействие с подвижной добычей по типу “хищник–жертва” (Caro, 1980), включающее обнаружение, сближение (преследование), атаку, обработку добычи, обездвиживание и поедание. В отличие от насекомоядного вида с облигатным проявлением охотничьего поведения (охотились все 11 бурозубок), у грызунов стереотип проявлялся факультативно, т.е. не у всех зверьков. При первом предъявлении добычи активное преследование и атаку продемонстрировали 65.4% (17 из 26) полевых мышей, 36.8% (7 из 19) хомячков Кэмпбелла и 18.5% (9 из 46) узкочерепных полевок. Отметим, что среди охотившихся полевых мышей 6 зверьков (2 самца и 4 самки) были рождены в виварии, т.е. 6 из 9 “наивных” животных проявили полную реакцию на добычу при первом предъявлении, а 3 не проявили. Среди зверьков, отловленных в естественных условиях, соотношение примерно такое же: 11 из 17.
После всех повторных тестов количество особей, проявивших охотничье поведение, несколько увеличилось: у полевых мышей – до 80.8%, у хомячков Кэмпбелла – до 63.2%, и до 39.1% у узкочерепных полевок. Рост числа “охотников” не достоверен: для хомячков p = 0.1939, для полевок p = 0.06595, для полевых мышей p = 0.3487 (точный тест Фишера).
Оценку результативности охоты у разных видов проводили только для охотившихся особей. По результатам первого тестирования (с последовательным предъявлением до трех насекомых) наиболее успешными “охотниками” оказались полевые мыши и обыкновенные бурозубки: соответственно 69.2% (45 из 65) и 62.3% (33 из 53) атак закончились поимкой и поеданием добычи. Это достоверно выше, чем у хомячков Кэмпбелла (20%; 7 из 20 атак) и узкочерепных полевок (25.4%; 16 из 63 атак) (точный тест Фишера, p < 0.0017 для обоих случаев). В первом тесте все охотившиеся мыши и бурозубки ловили и поедали добычу, иногда после нескольких неудачных атак. У 22% (2 из 9 особей) охотившихся узкочерепных полевок и у 57% (4 из 7 особей) хомячков Кэмпбелла наблюдались только неудачные атаки, т.е. охотничье поведение проявлялось, но добыча так и не была поймана. У 9 полевых мышей и 2 узкочерепных полевок успешными были все атаки, т.е. они ни разу не упускали добычу. У хомячков Кэмпбелла во всех охотах успешным атакам предшествовала хотя бы одна неудачная. У полевок успешным атакам предшествовало больше неудачных атак – 5.0 (2.8–6.3), чем у полевых мышей – 1.0 (1.0–2.0), хомячков Кэмпбелла – 1.0 (1.0–1.3) и обыкновенных бурозубок – 2.0 (1.0–2.8) (H = 11.1, p < 0.05).
По результатам всех повторных тестов результативность охоты, т.е. соотношение успешных и неудачных атак у полевых мышей (83 и 39), хомячков Кэмпбелла (43 и 42) и обыкновенных бурозубок (61 и 34), достоверно не различалась как между видами, так и с результатами первого тестирования для каждого из видов. Результативность охоты узкочерепных полевок осталась самой низкой (34 и 92) (p < 0.0024 для всех случаев). Количество неудачных атак, предшествующих успешной поимке добычи, после повторных тестов (без учета первого) у полевых мышей – 1.0 (1.0–1.0), хомячков Кэмпбелла – 1.0 (1.0–1.0) и обыкновенных бурозубок – 1.0 (1.0–1.0) изменилось незначительно, а у узкочерепных полевок достоверно снизилось до 2.0 (1.0–3.0) (H = 11.2, p < 0.05) и стало сходным с тем же показателем у полевых мышей и обыкновенных бурозубок. Изменение этого показателя у полевок не отразилось на результативности охоты.
Наиболее быстро охотились обыкновенные бурозубки, совершая 2.9 (1.8–4.2) элемента поведения в секунду. Их скорость охоты была достоверно выше, чем у грызунов: полевых мышей – 2.1 (1.5–2.8) (H = 11.2, p < 0.001), узкочерепных полевок – 1.6 (1.2–2.2) (H = 20.5, p < 0.001) и хомячков Кэмпбелла – 1.4 (1.0–1.9) (H = 35.2, p < 0.001). Скорость охоты у полевых мышей была выше, чем у хомячков Кэмпбелла (H = 20.0, p < 0.001), а узкочерепные полевки по этому показателю достоверно не отличались ни от полевых мышей, ни от хомячков.
В охотничьем поведении исследованных видов выделено 19 элементов, по два из них наблюдались только у бурозубок или только у грызунов. Элементы поведения разделены на три типа:
1) ключевые элементы, без которых реализация стереотипа невозможна: преследование добычи бегом (Q) или спокойным шагом (S), укус (W), захват добычи лапами (E) (только у грызунов);
2) дополнительные элементы (“приготовления” к охоте и “обработка” добычи) присутствовали не во всех стереотипах: принюхивание (D), перенос добычи в зубах (G), перехват добычи лапами (R) (только у грызунов), откусывание конечностей добычи (H), придерживание добычи одной (N) или двумя лапами (M) (только у бурозубок);
3) “шумовые” элементы, не влияющие на реализацию стереотипа: замирание (C), поворот корпуса на 90° (V), разворот корпуса на 180° (B), поворот головы (F), вертикальная стойка (I), стойка с опорой на арену (Y), движения назад (U), прыжок (J) и чистка (X).
Полученные значения количества отдельных элементов поведения, приходящихся на один охотничий стереотип у разных видов, представлены в табл. 1.
Таблица 1.
Количество элементов поведения, приходящихся на один охотничий стереотип, у исследованных видов
| Элементы поведения и переходы между ними | Вид | |||
|---|---|---|---|---|
| полевая мышь | узкочерепная полевка | хомячок Кэмпбелла | обыкновенная бурозубка | |
| Укус (W) | 20 (10–30) | 6 (2–20.5) | 8 (4.5–8) | 15 (8–22) |
| Захват лапами (E) | 7 (4–11) | 2 (1–8.5) | 10 (2.5–17.5) | – |
| Перехват (R) | 1 (0–2) | 0 (0–0) | 1 (0–2) | – |
| Откусывание конечностей (H) | 1 (0–2) | 1 (0–2.8) | 3 (2–6) | 1 (0–3) |
| Одиночный укус (W1) | 4 (3–7) | 2 (1–7) | 6 (2.5–10) | 1 (0–3) |
| Двойной укус (W2) | 1 (0–3) | 1 (0–3) | 1 (0–1.5) | 1 (0–1) |
| Множественный укус (Wn) | 3 (1–4) | 0 (0–1.8) | 0 (0–0.5) | 2 (1–3) |
Чтобы составить представление о том, как разные исследованные виды атакуют добычу и как манипулируют с ней, мы сравнили количество элементов поведения, связанных с атакой и обработкой добычи, приходящихся на один стереотип (табл. 1). Количество укусов у узкочерепных полевок и хомячков Кэмпбелла меньше, чем у полевых мышей (H = 87.5, p < 0.001; H = 17.7, p < 0.001 соответственно) и обыкновенных бурозубок (H = 7.8, p < 0.005; H = 11.7, p < 0.005). Узкочерепные полевки совершали захваты лапами и перехваты добычи очень редко, значительно реже, чем хомячки Кэмпбелла (H = 14.6, p < 0.0016; H = 14.9, p < 0.001 соответственно) и полевые мыши (H = 9.9, p < 0.0083; H = 14.9, p < 0.001). Хомячки Кэмпбелла откусывали конечности добычи чаще, чем особи всех остальных видов, что отражает специфику паттерна: полевые мыши (H = 33.8, p < 0.001), узкочерепные полевки (H = 14.4, p < 0.001) и обыкновенные бурозубки (H = 21.2, p < 0.001).
Чтобы выяснить, каким образом “охотник” повреждает добычу, исследовали повторяемость элемента поведения “укус”. Вся совокупность этих элементов была разделена на три группы: одиночные, двойные (два последовательно совершаемых укуса) и множественные (три и более укуса подряд). Результаты представлены в табл. 1. Количество одиночных укусов в охотничьих стереотипах обыкновенных бурозубок достоверно меньше, чем в стереотипах полевых мышей (H = 36.4, p < 0.001) и хомячков Кэмпбелла (H = 32.2, p < 0.001). У узкочерепных полевок и обыкновенных бурозубок количество одиночных укусов не различалось. По количеству одиночных и двойных укусов стереотипы грызунов не различались между собой. Полевые мыши и обыкновенные бурозубки совершали множественные укусы чаще, чем узкочерепные полевки (H = 16.6, p < 0.001; H = 12.5, p < 0.001) и хомячки Кэмпбелла (H = 43.3, p < 0.001; H = 37.6, p < 0.001). Несмотря на меньшее количество множественных укусов в стереотипах узкочерепных полевок, можно сказать, что по характеру атаки они сходны с полевыми мышами и бурозубками, т.е. схватывают насекомое зубами и повреждают серией укусов. Таким образом, полевые мыши, узкочерепные полевки и обыкновенные бурозубки завершают атаку на насекомое сходным образом – искусывают насмерть.
На основании матриц вероятностей переходов между элементами поведения построены схемы охотничьих стереотипов (рис. 1). У всех четырех видов охотничий стереотип начинался со сближения с добычей бегом (Q) или спокойным шагом (S), затем могло следовать принюхивание (D). Далее, у полевых мышей и узкочерепных полевок захват добычи происходил зубами – укус (один, но чаще несколько подряд) (W). У хомячков Кэмпбелла за сближением с добычей с большей вероятностью следовал захват зубами – укус (W), с меньшей вероятностью – захват добычи лапами (E). Отметим, что у хомячков вероятность первичного захвата добычи лапами составляет более 25%, тогда как у мышей и полевок подобные действия наблюдались в единичных случаях и заканчивались неуспешно. Если хомячки после первичного захвата добычи лапами способны ее удержать и перейти к повреждению и поеданию, то мыши и полевки, захватив насекомое лапами, сразу его теряют, и успешные атаки для них возможны только при начале атаки с захвата зубами. С момента захвата добычи схемы стереотипов охотничьего поведения грызунов и обыкновенных бурозубок расходятся. После укуса (W) грызуны схватывают добычу обеими лапами (Е) для поедания. У бурозубок этот элемент поведения отсутствует, они реже одной (N), чаще двумя (M) передними конечностями прижимают добычу ко дну арены, продолжая наносить укусы, и через некоторое время приступают к поеданию. После поимки все зверьки могли переносить добычу в зубах (G). Если атака начиналась с укуса (W), то в дальнейшем грызуны схватывали добычу лапами (Е), и после этого могли наблюдаться перехваты (R), которые включали манипуляции с удерживаемой в лапах добычей (повороты, перевороты). У бурозубок элемент поведения “перехват”, естественно, отсутствовал. В дальнейшем все зверьки могли откусывать конечности добычи (H), наиболее часто такое поведение отмечалось у хомячков Кэмпбелла. В поведенческой последовательности последний элемент поведения, предшествующий поеданию, выступает в качестве окончания успешного охотничьего стереотипа. У грызунов такими финальными элементами могли быть: укус (W), захват лапами (E), перехват (R) или откусывание конечностей добычи (H); у бурозубок: укус (W), придерживание добычи одной лапой (N), откусывание конечностей добычи (H), а также принюхивание (D) и перенос добычи в зубах (G), встречавшиеся единично.
Рис. 1.
Схемы охотничьих стереотипов полевой мыши (а), узкочерепной полевки (б), хомячка Кэмпбелла (в) и обыкновенной бурозубки (г). Обозначения элементов поведения см. в разделе “Результаты”. В квадратных скобках указаны дополнительные элементы. Тонкой пунктирной линией обозначены некоторые неустойчивые связи между элементами (p < 0.2). Простой линией обозначены устойчивые связи (0.2 ≤ p < 0.5). Жирной линией обозначены высоко устойчивые связи между элементами (p ≥ 0.5).
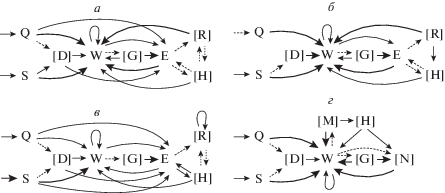
Стоит напомнить, что все исследованные хомячки и часть полевых мышей родились в лаборатории и были “наивными” по отношению к потенциальной добыче. Стереотип охотничьего поведения у этих животных либо проявлялся полностью при первой же или одной из последующих встреч с добычей, по принципу “все и сразу”, либо не проявлялся совсем. Сравнение стереотипов охоты полевых мышей, родившихся в лаборатории (n = 6), и полевых мышей, отловленных в естественных условиях (n = 11), не выявило никаких различий в их охотничьем поведении. Как и при межвидовых сравнениях, анализировали количество поведенческих элементов, приходящихся на один стереотип: “укус” (H = 0.8, NS), “захват лапами” (H = 0.2, NS), “перехват” (H = 1.2, NS), “откусывание конечностей добычи” (H = 0.3, NS), а также количество одиночных (H = 0.2, NS), двойных (H = 2.9, NS) и множественных укусов (H = 0.3, NS). Схемы стереотипа у диких и лабораторных полевых мышей также не отличались и полностью идентичны схеме, представленной на рис. 1.
Полученные данные позволили составить представление о тактике охоты у разных видов, т.е. совокупности приемов для повреждения или умерщвления добычи. Характер охотничьих атак у грызунов и насекомоядных различен: грызуны после захвата добычи зубами осуществляют захват лапами, а бурозубки используют только зубы. Тактики охоты оказались сходными у полевых мышей, узкочерепных полевок и обыкновенных бурозубок: быстрое повреждение добычи продолжительной серией укусов. Тактика охоты хомячков Кэмпбелла весьма специфична: повреждение и обездвиживание добычи у них всегда происходит путем откусывания конечностей. Это выражается в устойчивости связей между элементами поведения “захват лапами” и “откусывание конечностей” (0.2 ≤ p < 0.5). Этот прием стабильно использовался всеми особями (n = 19). Поскольку такая тактика охоты выявлена у грызунов впервые, мы решили проверить, универсальна ли она для данного вида, и используют ли хомячки Кэмпбелла тот же прием для обездвиживания не только безопасной добычи, но и опасных насекомых. Для этого отдельной группе из 10 хомячков предлагали рыжих лесных муравьев. Однако если для полевых мышей муравьи обладают высокой гедонистической ценностью и съедаются без остатка (Panteleeva et al., 2013), то хомячки Кэмпбелла, как оказалось, их умерщвляют, но не едят. Если охотничье поведение полевых мышей по отношению к тараканам и муравьям в целом сходно (как к съедобному объекту), то у хомячков Кэмпбелла прием для повреждения опасных насекомых совсем иной. Они совершают захват только зубами (никогда не захватывают муравьев лапами), затем берут насекомое в лапы и сразу, или после нескольких перехватов, повернув муравья к себе брюшком, разгрызают его, бросая затем поврежденное насекомое на дно арены. Отметим, что если охотничье поведение по отношению к тараканам проявлялось не у всех хомячков и не всегда было успешным, то повреждение агрессивных муравьев наблюдалось у всех особей с одинаково высокой эффективностью. Поведение хомячков по отношению к муравьям можно отнести не к охотничьему, а к оборонительному, и проявляется оно не факультативно, как в случае охоты, а облигатно.
ОБСУЖДЕНИЕ
В наших экспериментах у трех видов грызунов с разными типами питания впервые выявлено и детально описано целенаправленное взаимодействие с подвижной добычей по типу “хищник–жертва”. Зерноядная полевая мышь, зеленоядная узкочерепная полевка и всеядный хомячок Кэмпбелла обладают одинаково эффективным стереотипным охотничьим поведением, во многом сходным с поведением специализированного насекомоядного вида – обыкновенной бурозубки. При этом скорость охоты у грызунов ниже, чем у насекомоядных. Структура охотничьего стереотипа оказалась универсальной у всех исследованных видов грызунов, независимо от их пищевой специализации. При одинаковом наборе элементов охотничьего поведения, сходном порядке действий и вероятностей перехода между ними у разных видов, стоит отметить, что у узкочерепных полевок, по сравнению с остальными видами, достоверно меньше повторяются такие элементы поведения, как “захват лапами” (после поимки зубами) и “перехват” (поиск более удобного положения); количество множественных укусов при умерщвлении насекомого у них также меньше, чем у других видов. Это придает им своеобразную неуверенность в обработке добычи и, возможно, является причиной несколько меньшей результативности охоты по сравнению с другими видами. У разных видов различается тактика повреждения добычи: полевые мыши, узкочерепные полевки и обыкновенные бурозубки обездвиживают насекомое серией быстрых укусов (искусывают насмерть), тогда как хомячки Кэмпбелла откусывают у добычи конечности, держа ее в лапах. Этот элемент поведения описан для грызунов впервые и, по-видимому, является проявлением более специализированного охотничьего поведения. Интересно отметить, что в естественных условиях найденные в норах хомячков Кэмпбелла саранчовые были лишены голов и конечностей, и можно полагать, что к ним был применен тот же прием (Левенец и др., 2019).
Для формирования представлений о возможной эволюции охотничьего поведения у исследованных видов стоит обратиться к имеющейся картине эволюции охотничьего поведения у грызунов. Предположительно, эволюционные истоки охотничьего поведения наземных млекопитающих исходят от общего всеядно-насекомоядного предка (Ewer, 1973), и эволюционный переход от растительноядного образа жизни к хищническому менее вероятен, чем от всеядного (Shipman, Walker, 1989). Синтез анатомических, генетических и палеонтологических данных позволил установить, что общим предком плацентарных млекопитающих было небольшое насекомоядное древесное животное (Meredith et al., 2011; O’Leary et al., 2013). Происхождение охотничьего поведения у грызунов остается неясным. Грызуны могли унаследовать уже имевшийся у насекомоядных предков стереотип охоты на мелкую подвижную добычу, либо такое поведение сформировалось повторно. Некоторые представления об эволюционном пути охотничьего поведения у грызунов можно получить, применив сравнительно-этологический подход. Поведенческие признаки, являясь продуктом отбора, могут быть использованы для филогенетических построений. Так, Лоренц, основываясь на том, что инстинктивные движения имеют систематическое значение, впервые использовал признаки поведения и вокализации для рассмотрения филогении системы утиных (Lorenz, 1941). Впоследствии его ученик Лейхаузен использовал сходный подход, реконструируя эволюционное древо охотничьего поведения наземных млекопитающих (Leyhausen, 1965; Eisenberg, Leyhausen, 1972). Вместе с Эйзенбергом они фиксировали последовательности действий, совершаемых хищником во время охоты, представив их в виде этограмм. Исследователи охватили обширное число видов от насекомоядных до приматов, но грызуны не вошли в этот список. В результате обобщения сравнительно-этологических данных они пришли к ряду заключений. Атака добычи серией укусов (захват и умерщвление добычи только с помощью челюстей) является примитивной формой охотничьего поведения. Появление в репертуаре захвата добычи передними лапами – более прогрессивная в эволюционном плане черта. Смещение функции захвата добычи к передним конечностям стало основой для дальнейшей дифференциации типов укуса: либо точно направленный одиночный смертельный укус, либо серия укусов. Смертельный укус является “недавним достижением”, а серии укусов характеризуются как более архаичный признак. В эволюционном плане такой прием, когда для захвата и удержания используются передние конечности, а челюсти смыкаются в смертельном укусе, увеличивает эффективность хищника. Лэнгли (Langley, 1987, 1994) детально исследовал стереотипы охоты на беспозвоночных у специализированных хищных грызунов и грызунов-эврифагов. Опираясь на картину эволюции охотничьего поведения (Eisenberg, Leyhausen, 1972), он впервые классифицировал охотничье поведение грызунов и показал, что неспециализированные грызуны-эврифаги в большинстве случаев начинают атаку на добычу с захвата зубами (укуса), а затем, удерживая ее в зубах, захватывают ее в передние лапы (Langley, 1987, 1994). Принципиально иным образом действуют специализированные хищные кузнечиковые хомячки, которые чаще всего в начале атаки захватывают добычу в передние лапы и только потом умерщвляют ее одним или несколькими укусами. Различные представители отряда насекомоядных, в частности землеройковые Soricidae, ежовые Erinaceidae, тенрековые Tenrecidae, атакуют добычу только сериями укусов, демонстрируя наиболее примитивный тип атаки (Eisenberg, Leyhausen, 1972). Таким образом, в охотничьем стереотипе грызунов по сравнению с насекомоядными наблюдаются прогрессивные черты (захват добычи лапами), а наиболее продвинутый стереотип охоты наблюдается у специализированных хищных кузнечиковых хомячков (Langley, 1994).
Для того чтобы найти место исследованных нами видов в этой картине, важно проанализировать начало атаки на добычу. Среди трех исследованных видов характер охотничьих атак у грызунов и насекомоядных различен: грызуны после захвата добычи зубами осуществляют захват лапами, а бурозубки используют только зубы. Поскольку у полевых мышей и узкочерепных полевок захват лапами следует после захвата зубами, охотничий стереотип у них можно считать более примитивным, чем у специализированных хищных кузнечиковых хомячков. Хомячки Кэмпбелла могут начать атаку с захвата как зубами, так и лапами (более чем в 25% случаев), что характеризует их охотничье поведение как наиболее продвинутое среди исследованных видов.
Видовой сценарий формирования стереотипа охотничьего поведения в онтогенезе у всех исследованных видов грызунов оказался сходен с таковым у специализированных хищных кузнечиковых хомячков: стереотип проявлялся у “наивных” животных по принципу “все и сразу”, являясь целостным и не меняясь в ходе повторных предъявлений добычи. Некоторым животным для “пробуждения” целостного стереотипа требовалось, по-видимому, кумулятивное действие стимулов, исходящих от добычи. Роль аккумуляции стимулов в запуске “врожденных высвобождающих механизмов” (Дьюсбери, 1981) была показана в исследованиях охотничьего поведения обыкновенной жабы (Ewert, 1987); позднее необходимость аккумуляции сенсорных сигналов в коре головного мозга для принятия решений у животных была продемонстрирована неоднократно (Watson, Platt, 2008). Доля особей с отсрочкой проявления охотничьего стереотипа в наших выборках составляет у трех видов грызунов от 15 до 26%. Заметим, что в наших экспериментах не наблюдалось совершенствование приемов охоты по мере накопления опыта, как это было продемонстрировано ранее у неспециализированных хомячков р. Peromyscus (Kreiter, Timberlake, 1988). Среди исследованных нами видов можно отметить лишь некоторое повышение числа успешных охот при повторных встречах с добычей у узкочерепных полевок. Скорость охоты, характер и результативность атак и, главное, порядок и соотношение элементов поведения в стереотипах у всех трех видов грызунов, как и у бурозубок, не были подвержены влиянию опыта.
Наши лабораторные эксперименты, по-видимому, отражают соотношение “охотников” и особей, равнодушных к насекомым, в естественных популяциях исследованных видов. Примечательно, что, хотя “охотников” среди зеленоядных полевок в наших экспериментах было вдвое меньше, чем среди полевых мышей, по характеру стереотипа, скорости и эффективности охоты все виды грызунов были сходны. Это позволяет предположить, что в популяциях грызунов с разными типами питания (по крайней мере, у некоторых видов) постоянно присутствует та или иная доля особей-носителей стереотипа охотничьего поведения, а сам стереотип может рассматриваться как поведенческая адаптация, позволяющая расширить спектр пищевых ресурсов путем активной охоты на насекомых. Стереотипы охотничьего поведения у исследованных видов грызунов носят характер универсального сценария с проявлением черт видовой специфичности, которые выражаются в характере атак и в способах обездвиживания добычи, а также в частоте проявления отдельных элементов поведения. Отмеченные видоспецифичные особенности не варьируют у представителей каждого вида, носят врожденный характер и либо проявляются у определенной доли особей, либо не проявляются совсем. Эти ключевые характеристики позволяют нам рассматривать наличие охотничьих стереотипов у грызунов как стратегию в рамках теории ЭСС.
В целом охотничье поведение грызунов, по-видимому, можно рассматривать как эволюционно стабильную стратегию, когда доля носителей определенного типа поведения может возрастать при определенных условиях, не захватывая популяцию полностью (Maynard Smith, Price, 1973). О подобном потенциале охотничьего поведения грызунов говорят эксперименты с целенаправленным отбором “охотников” у зерно-зеленоядного вида – рыжей полевки M. glareolus (Konczal et al., 2016). После 13 поколений такого отбора доля полевок, демонстрирующих охотничье поведение по отношению к сверчкам, в отдельных линиях зверьков была в 5 раз выше, чем в контрольных группах.
Ранее подобные эксперименты проводились на различных генетических линиях домовых мышей (Butler, 1973) и на сирийских хомячках (Polsky, 1978). У хомячков за 8 поколений были получены линии “ловцов” и “тормозов”: задержка перед атакой на добычу различалась у этих групп в 5 раз. Исследования генетических механизмов, отвечающих за проявление охотничьего поведения у инбредных линий мышей, показали, что высокий уровень хищнической агрессии (как ранее называли охотничье поведение грызунов физиологи и генетики) является доминантным признаком (Никулина, Попова, 1983).
Характер охотничьего поведения грызунов до нашей работы в деталях не исследовался, и для изучения сравнительной “конкурентоспособности” различающихся стереотипов в условиях целенаправленного отбора нужны специальные эксперименты. На основании наших предварительных данных можно сделать осторожное предположение о том, что количество носителей стереотипа охотничьего поведения может варьировать между популяциями в разные годы и, по-видимому, связано с колебаниями внешних условий. Следует отметить, что, хотя стереотип охотничьего поведения проявляется у определенной доли особей исследованных видов по принципу “все и сразу”, зверьки, равнодушные к насекомым, могут являться носителями “спящих” фрагментов целостного стереотипа (dormant behavioural patterns; Reznikova, Panteleeva, 2008). Феномен фрагментации стереотипа охотничьего поведения был недавно продемонстрирован нами на примере некоторых видов грызунов (Резникова и др., 2017). Можно предположить, что поведенческим механизмом, позволяющим при необходимости быстро нарастить количество “охотников” в популяциях, может являться распределенное социальное обучение (Резникова, Пантелеева, 2015): если в популяции имеются носители полного стереотипа охотничьего поведения, то возможные носители фрагментов стереотипа могут быстро их “достроить” за счет простых форм социального обучения. Эти вопросы требуют дальнейших исследований.
ЗАКЛЮЧЕНИЕ
Детальные экспериментальные исследования взаимодействия мелких млекопитающих с подвижными насекомыми показали, что зерноядная полевая мышь, зеленоядная узкочерепная полевка и всеядный хомячок Кэмпбелла обладают охотничьим поведением столь же эффективным, что и у насекомоядного вида – обыкновенной бурозубки. Структура стереотипа охотничьего поведения у исследованных видов грызунов универсальна и не зависит от пищевой специализации. У разных видов различаются тактики повреждения добычи: полевые мыши и узкочерепные полевки обездвиживают насекомое серией быстрых укусов так же, как это делают бурозубки; хомячки Кэмпбелла откусывают у добычи конечности, что, по-видимому, является проявлением более специализированного охотничьего поведения. Характер охотничьих атак у грызунов и насекомоядных различен: грызуны, вначале захватывая добычу зубами, переходят к захвату лапами, а бурозубки используют только зубы, что говорит об относительной примитивности их хищнического поведения. Хомячки Кэмпбелла могут начать атаку с захвата как зубами, так и лапами, что характеризует их охотничье поведение как наиболее эволюционно продвинутое среди исследованных видов. Характер и соотношение элементов поведения у всех трех видов грызунов проявляются по принципу “все и сразу” и не подвержены влиянию опыта. Охотничье поведение грызунов можно рассматривать как эволюционно стабильную стратегию, поддерживающую способность популяций использовать охоту на подвижных насекомых для расширения спектра пищевых ресурсов.
Исследования поддержаны грантами РФФИ (№17-04-00702 и №18-34-00119) и программы ФНИ государственных академий наук на 2013–2020 гг., проект № VI.51.1.10. (АААА-А16-116121410120-0).
Список литературы
Громов В.С., 2008. Пространственно-этологическая структура популяций грызунов. М.: Т-во науч. изд. КМК. 581 с.
Дьюсбери Д.М., 1981. Поведение животных: Сравнительные аспекты. М.: Мир. 480 с.
Зорина З.А., Полетаева И.И., Резникова Ж.И., 2013. Основы этологии и генетики поведения: Учебник. 3-е изд., испр. и доп. М.: Изд-во МГУ. 384 с.
Левенец Я.В., Пантелеева С.Н., Резникова Ж.И., 2016. Экспериментальное исследование питания насекомыми у грызунов // Евразиатский энтомол. журн. Т. 6. № 6. С. 550–554.
Левенец Я.В., Пантелеева С.Н., Резникова Ж.И., Гуреева А.В., Феоктистова Н.Ю., Суров А.В., 2019. Экспериментальный сравнительный анализ охотничьего поведения четырех видов хомячков подсемейства Сricetinae // Зоол. журн. Т. 98. № 6. С. 673–683.
Никулина Э.М., Попова Н.К., 1983. Генетический анализ агрессивности хищника у мышей // Генетика. Т. 16. № 7. С.105–110.
Пантелеева С.Н., Данзанов Ж.А., Резникова Ж.И., 2010. Оценка сложности поведенческих стереотипов у муравьев на примере анализа охотничьего поведения Myrmica rubra (Hymenoptera, Formicidae) // Зоол. журн. Т. 89. № 12. С. 500–509.
Пантелеева С.Н., Резникова Ж.И., Синькова О.Б., 2016. Пространственно-этологические аспекты взаимодействия мелких млекопитающих с рыжими лесными муравьями // Журн. общ. биологии. Т. 77. № 5. С. 329–341.
Резникова Ж.И., Пантелеева С.Н., 2005. Экспериментальное исследование формирования охотничьего поведения в онтогенезе муравьев // Докл. АН. Т. 401. № 1. С. 139–141.
Резникова Ж.И., Пантелеева С.Н., 2015. Возможные эволюционные механизмы “культуры” у животных: гипотеза распределенного социального обучения // Журн. общ. биологии. Т. 76. № 4. С. 309–323.
Резникова Ж.И., Пантелеева С.Н., Новиковская А.А., 2017. Фрагментарность поведенческих стереотипов как возможная основа распределенного социального обучения в популяциях и сообществах // Современные проблемы биологической эволюции: мат-лы III Междунар. конф., к 130-летию со дня рождения Н.И. Вавилова и 110-летию со дня основания Государственного Дарвиновского музея. М.: ГДМ. С. 57–60.
Butler K., 1973. Predatory behavior in laboratory mice: Strain and sex comparisons // J. Comp. Physiol. Psychol. V. 85. № 2. P. 243–249.
Caro T.M., 1980. The effects of experience on the predatory patterns of cats // Behav. Neural Biol. V. 29. № 1. P. 1–28.
Casarrubea M., Sorbera F., Crescimanno G., 2008. Multivariate analysis of the modifications induced by an environmental acoustic cue on rat exploratory behavior // Physiol. Behav. V. 93. № 4. P. 687–696.
Dominey W.J., 1984. Alternative mating tactics and evolutionarily stable strategies // Am. Zool. V. 24. № 2. P. 385–396.
Eisenberg J.F., Leyhausen P., 1972. The phylogenesis of predatory behavior in mammals // Z. Tierpsychol. V. 30. № 1. P. 59–93.
Ewer R.F., 1973. The Carnivores. Ithaca: Cornell Univ. Press. 494 p.
Ewert J.P., 1987. Neuroethology of releasing mechanisms: Prey-catching in toads // Behav. Brain Sci. V. 10. № 3. P. 337–368.
Hamilton W.D., 1967. Extraordinary sex ratios // Science. V. 156. № 3774. P. 477–488.
Han W., Tellez L.A., Rangel M.J., Motta S.C. et al., 2017. Integrated control of predatory hunting by the central nucleus of the amygdala // Cell. V. 168. № 1. P. 311–324.
Haug M., Johnson F., 1991. Environmental and biological incidences over predatory aggression in rodents // Comp. Biochem. Physiol. V. 99. № 3. P. 205–293.
Hoy J.L., Yavorska I., Wehr M., Niell C.M., 2016. Vision drives accurate approach behavior during prey capture in laboratory mice // Curr. Biol. V. 26. № 22. P. 3046–3052.
Konczal M., Koteja P., Orlowska-Feuer P., Radwan J., Sadowska E.T., Babik W., 2016. Genomic response to selection for predatory behavior in a mammalian model of adaptive radiation // Mol. Biol. Evol. V. 33. № 9. P. 2429–2440.
Kreiter N., Timberlake W., 1988. Form and development of predation on crickets in adults of Peromyscus maniculatus bairdi and P. leucopus noveboracensis // J. Comp. Psychol. V. 102. № 3. P. 269–278.
Landry S.O., Jr., 1970. The Rodentia as omnivores // Q. Rev. Biol. V. 45. № 4. P. 351–372.
Langley W.M., 1986. Development of predatory behavioral in the southern grasshopper mouse (Onychomys torridus) // Behaviour. V. 99. № 3. P. 275–295.
Langley W.M., 1987. Specializations in the predatory behavior of grasshopper mice (Onychomys leucogaster and O. torridus): A comparison with the golden hamster (Mesocricetus auratus) // J. Comp. Physiol. Psychol. V. 101. № 4. P. 322–327.
Langley W.M., 1994. Comparison of predatory behaviors of deer mice (Peromyscus maniculatus) and grasshopper mice (Onychomys leucogaster) // J. Comp. Physiol. Psychol. V. 108. № 4. P. 394–400.
Leyhausen P., 1965. Über die Funktion der Relativen Stimmungshierarchie (Dargestellt am Beispiel der phylogenetischen und ontogenetischen Entwicklung des Beutefangs von Raubtieren) // Z. Tierpsychol. V. 22. № 4. P. 412–494.
Lorenz K., 1941. Vergleichende Bewegungsstudien an Anatinen // J. Ornithol. V. 89. P. 19–29.
MacArthur R.H., 1965. Patterns of species diversity // Biol. Reviews. V. 40. № 4. P. 510–533.
Maynard Smith J., Price G.R., 1973. The logic of animal conflict // Nature. V. 246. № 5427. P. 15–18.
Meredith R.W., Janečka J.E., Gatesy J., Ryder O.A., Fisher C.A. et al., 2011. Impacts of the Cretaceous terrestrial revolution and KPg extinction on mammal diversification // Science. V. 334. № 6055. P. 521–524.
Modlinska K., Stryjek R., 2016. Food neophobia in wild rats (Rattus norvegicus) inhabiting a changeable environment – a field study // PloS One. V. 11. № 6. P. e0156741.
O’Leary M.A., Bloch J.I., Flynn J.J., Gaudin T.J., Giallombardo A. et al., 2013. The placental mammal ancestor and the post–K-Pg radiation of placentals // Science. V. 339. № 6120. P. 662–667.
Panteleeva S., Reznikova Z., Vygonyailova O., 2013. Quantity judgments in the context of risk/reward decision making in striped field mice: first “count,” then hunt // Front. Psychol. V. 4. P. 45–53.
Polsky R.H., 1977. The ontogeny of predatory behaviour in the golden hamster (Mesocricetus a. auratus): I. The influence of age and experience // Behaviour. V. 61. № 1–2. P. 26–57.
Polsky R.H., 1978. The ontogeny of predatory behavior in the golden hamster (Mesocricetus a. auratus): IV. Effects of prolonged exposure, ITI, size of prey and selective breeding // Behaviour. V. 65. № 1–2. P. 27–39.
Quinn J.L., Cresswell W., 2004. Predator hunting behaviour and prey vulnerability // J. Anim. Ecol. V. 73. № 1. P. 143–154.
Reznikova Zh., Panteleeva S., 2008. An ant’s eye view of culture: Propagation of new traditions through triggering dormant behavioural patterns // Acta Ethol. V. 11. № 2. P. 73–80.
Reznikova Z., Levenets J., Panteleeva S., Ryabko B., 2017. Studying hunting behaviour in the striped field mouse using data compression // Acta Ethol. V. 20. № 2. P. 165–173.
Rowe A.H., Rowe M.P., 2006. Risk assessment by grasshopper mice (Onychomys spp.) feeding on neurotoxic prey (Centruroides spp.) // Anim. Behav. V. 71. № 3. P. 725–734.
Sarko D.K., Leitch D.B., Girard I., Sikes R.S., Catania K.C., 2011. Organization of somatosensory cortex in the Northern grasshopper mouse (Onychomys leucogaster), a predatory rodent // J. Comp. Neurol. V. 519. № 1. P. 64–74.
Shipman P., Walker A., 1989. The costs of becoming a predator // J. Hum. Evol. V. 18. № 4. P. 373–392.
Sirot E., 2000. An evolutionarily stable strategy for aggressiveness in feeding groups // Behav. Ecol. V. 11. № 4. P. 351–356.
Soininen E.M., Ravolainen V.T., Bråthen K.A., Yoccoz N.G., Gielly L., Ims R.A., 2013. Arctic small rodents have diverse diets and flexible food selection // PLoS One. V. 8. № 6. P. e68128.
Timberlake W., Washburne D.L., 1989. Feeding ecology and laboratory predatory behavior toward live and artificial moving prey in seven rodent species // Anim. Learn. Behav. V. 17. № 1. P. 2–11.
Watson K.K., Platt M.L., 2008. Neuroethology of reward and decision making // Phil. Trans. R. Soc. B: Biol. Sci. V. 363. № 1511. P. 3825–3835.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии