

Журнал общей биологии, 2019, T. 80, № 4, стр. 251-259
Имитационная компьютерная модель гибридной зоны: ассортативность спаривания в роли отбора против гибридов
А. С. Рубцов *
Государственный Дарвиновский музей
117292 Москва, ул. Вавилова, 57, Россия
* E-mail: alexrub@darwinmuseum.ru
Поступила в редакцию 06.09.2018
После доработки 26.03.2019
Принята к публикации 26.04.2019
Аннотация
Представление о пониженной приспособленности гибридов служит краеугольным камнем логического обоснования дискретности видов с точки зрения биологической концепции. Большинство природных гибридных зон между формами околовидового ранга узки и стабильны во времени, что хорошо согласуется с математической моделью “напряженной” (tension) зоны, предполагающей наличие отбора против гибридов. Полагают, что у гибридов разрушаются коадаптированные генные комплексы родительских форм, что неизбежно должно выражаться в их пониженной жизнеспособности и/или плодовитости. Однако до сих пор это утверждение остается не более чем гипотезой. Пониженный репродуктивный успех гибридов может быть связан и с другими причинами: например, они могут испытывать сложности при формировании брачных пар в силу редкости и необычности своего фенотипа. Данное исследование посвящено проверке этого предположения, для чего была написана имитационная компьютерная модель гибридной зоны. Основная цель работы заключалась в получении ответов на следующие вопросы. 1. Возможно ли формирование узкой и стабильной гибридной зоны при наличии ассортативности спаривания и отсутствии снижения жизнеспособности и плодовитости гибридов? Ассортативность спаривания в модели обеспечивается импринтингом: потомки запечатлевают облик одного из родителей и ищут соответствующего полового партнера. 2. Можно ли в этих условиях ожидать завершения процесса видообразования, и что для этого необходимо? Под видообразованием в данном случае понимается процесс установления симпатрии между исходно аллопатричными формами при низком уровне гибридизации и сохранении их внешнего облика. Исследование модели дало положительные ответы на оба вопроса. При отсутствии биотопической сегрегации между родительскими формами и наличии ассортативности спаривания формируется узкая и стабильная во времени гибридная зона. В этом случае гибриды несколько чаще особей с фенотипом родительских форм оставались холостыми (5–8% и 1–3% соответственно), что и обеспечивало наличие отбора, направленного на поддержание узости гибридной зоны. Для формирования симпатрии, помимо ассортативности спаривания, необходимо наличие биотопической сегрегации, причем гибриды должны быть менее успешны в выборе индивидуальных участков, чем родительские формы. В противном случае, когда биотопы обеих родительских форм для гибридов являются оптимальными, зона контакта сначала расширяется, а затем, после того как у родительских форм происходит нивелирование биотопической сегрегации, начинает медленно сужаться вплоть до формирования узкой гибридной зоны. При этом по селективно нейтральным локусам (анализ ДНК-маркеров) исходные формы становились неразличимыми на всем пространстве модели.
В рамках биологической концепции вида, пользующейся по настоящее время наибольшей популярностью (Arnold, 1997; Coyne, Orr, 2004; Price, 2008; Лавренченко, 2013), ключевым моментом видообразования является отказ особей дивергирующих таксонов от гетероспецифического спаривания (Майр, 1968). В связи с этим изучение зон вторичного контакта и гибридизации является наиболее информативным способом исследования процессов видообразования: анализ ситуаций, когда изолирующие механизмы оказываются неэффективными, позволяет ответить на вопрос об их возможном генезисе и особенностях функционирования. Теоретическое обоснование дискретности видов в рамках биологической концепции (подробнее см. Рубцов, 2015) базируется на гипотезе “усиления” (reinforcement), основные положения которой сводятся к следующим утверждениям (Butlin, 1987). В зоне вторичного контакта гибриды должны обладать пониженной жизнеспособностью и/или плодовитостью вследствие разрушения у них коадапированных генных комплексов родительских форм. Вследствие этого селективное преимущество должны иметь те особи родительских форм, которые обладают наследственной предрасположенностью к конспецифичному спариванию. Таким образом, отбор против гибридов способствует совершенствованию механизмов прекопуляционной изоляции, в первую очередь этологической, и завершению процесса видообразования (прекращению гибридизации). Формирование же коадаптированных генных комплексов является неизбежным следствием дивергенции на стадии географической изоляции, что описывается моделью Добржанского–Мёллера (Orr, 1995; Price, 2008; Добржанский, 2010).
Действительно, большинство природных гибридных зон узки и стабильны во времени, их структура соответствует модели “напряженной” (tension) зоны, предполагающей наличие отбора против гибридов (Barton, Hewitt, 1985). Тем не менее в целом ряде случаев природной гибридизации в узких гибридных зонах не было обнаружено сколь-либо заметного снижения жизнеспособности и плодовитости гибридов (Рубцов, 2015). В самом деле, существование отбора против гибридов не означает их пониженную приспособленность. В роли отбора против гибридов может выступать, например, ассортативность спаривания (половой отбор против гибридов), которая показана для многих случаев природной гибридизации (Randler, 2008). Гибриды, обладая нормальной жизнеспособностью и плодовитостью, могут, тем не менее, иметь пониженный репродуктивный успех вследствие затрудненного формирования брачных пар из-за редкости и необычности своего фенотипа. Особенно отчетливо это должно проявляться по краям гибридной зоны, где доля фенотипических гибридов невелика. Эта гипотеза высказывалась достаточно давно (Крюков, Блинов, 1989), но так и не была подтверждена эмпирически из-за очевидных сложностей методологического характера (Price, 2008).
Едва ли не единственным решением этой проблемы представляется моделирование подобных ситуаций в искусственных условиях. Традиционные математические модели зачастую чрезмерно упрощают реальную ситуацию, которую призваны отразить, что сильно ограничивает сферу их применения. Ситуация изменилась около 20 лет назад, когда с развитием компьютерной техники стало возможным моделирование на базе клеточных автоматов. Под последними понимают класс имитационных моделей, в которых время и пространство представлены дискретными единицами, а поведение самой модели полностью задается начальными условиями и правилами поведения ее элементов (Грабовский, 1995а). Поскольку многие виды птиц и млекопитающих в сезон размножения занимают индивидуальные участки, а стадии жизненного цикла (занятие участков, размножение, осенние миграции) обладают ярко выраженной сезонностью, подобные ограничения вполне допустимы. Следовательно, клеточные автоматы с успехом могут быть использованы для моделирования популяционных процессов при решении широкого спектра экологических и эволюционных задач. В данной работе представлены результаты исследования имитационной компьютерной модели гибридной зоны. В первую очередь интерес представляли ответы на следующие вопросы. Может ли ассортативность спаривания обеспечивать формирование узкой и стабильной во времени гибридной зоны? Можем ли мы ожидать завершения процесса видообразования (установление симпатрии между исходно аллопатричными формами при сохранении их первоначального фенотипического облика) при отсутствии снижения жизнеспособности и плодовитости гибридов?
Из опубликованных ранее работ наиболее близка к решению поставленных задач модель гибридной зоны, предложенная В.И. Грабовским (1995б). В цитированной работе рассматривается вариант одномерной модели, результаты анализа двумерного варианта модели опубликованы в интернете11. В данной модели особи описываются гибридным индексом, гибридный индекс потомства наследуется как среднее арифметическое гибридных индексов родителей. Исходные формы характеризуются значением индекса –5 и +5 соответственно, гибриды имеют промежуточные значения. Выбор партнера в случае ассортативного спаривания (чем более близки индексы потенциальных партнеров, тем больше вероятность формирования брачной пары), биотопическая сегрегация (две особи могут занять один и тот же участок, если их гибридные индексы различаются знаком – больше и меньше 0 соответственно) и жизнеспособность гибридов (гибриды умирают, если модуль их гибридного индекса меньше порогового значения) определяются гибридным индексом.
Таким образом, модель В.И. Грабовского соответствует полигенному варианту наследования признаков, причем внешний облик и биотопические предпочтения особей оказываются генетически сцепленными признаками. Очевидно, что это лишь частный случай, далеко не всегда реализующийся в природе. Действительно, недавние исследования природной гибридизации черной (Corvus corone) и серой (C. cornix) ворон с помощью полногеномного секвенирования показали, что аллопатрические популяции родительских видов статистически значимо отличаются всего лишь по пяти локусам, локализованным в разных хромосомах, а в гибридной зоне фиксированные различия наблюдаются только по одному из них (Poelstra et al., 2014). Поэтому в модели, описываемой в данной статье, был реализован другой механизм наследования признаков: внешний облик и биотопические предпочтения особей определяются небольшим числом менделирующих генов, и сами признаки наследуются независимо друг от друга.
В модели В.И. Грабовского особи ищут полового партнера с таким же, как и у них, гибридным индексом, а гибридизация возникает вследствие ошибок при распознавании, точность которого задается экспериментатором перед запуском модели. В природе, по всей видимости, реализуется иной механизм. Во-первых, образ полового партнера задается не генетически, а с помощью полового импринтинга, по крайней мере у птиц: птенцы, находясь в гнезде, запечатлевают облик своих родителей и впоследствии ищут полового партнера, соответствующего запечатленному образу (Ten Cate, 1991). Во-вторых, показано, что интенсивность гибридизации возрастает в условиях редкости одного из контактирующих видов (Randler, 2002): в условиях дефицита конспецифичных партнеров птицы “смягчают” критерии поиска (принцип “вакуумной активности”; Панов, 1989). Такой механизм выбора полового партнера может существенно снижать уровень гибридизации за счет увеличения доли возвратных скрещиваний.
И, наконец, биотопическая сегрегация в модели В.И. Грабовского реализуется по принципу “все или ничего”. В нашей модели введен отдельный параметр, который определяет вероятность занятия особью субоптимального биотопа (свободный участок с оптимальным биотопом занимается всегда со 100% вероятностью).
ОПИСАНИЕ МОДЕЛИ
Модель гибридной зоны написана на языке Free Pascal в свободно распространяемой графической среде Lazarus (Алексеев и др., 2010). Действие происходит на территории размером 120 × 200 индивидуальных участков. Каждый участок может быть свободен или занят одной особью и представляет собой биотоп, который характеризуется одним из двух возможных значений. Биотопы распределены по территории в случайном порядке. Каждая особь обладает фенотипом, памятью и генотипом. Фенотип слагается из трех признаков: 1) внешнего облика, по которому происходит выбор полового партнера (брачная окраска), 2) биотопических предпочтений и 3) нейтральных признаков (аналог ДНК-маркеров). Память хранит внешний облик одного из родителей особи, по которому она выбирает себе полового партнера (половой импринтинг), а также используется для отражения текущего состояния особи (ее готовность к размножению, заняла ли она свой индивидуальный участок, спаривается ассортативно или случайным образом). Генотип состоит из десяти генов, каждый из которых представлен в популяции двумя аллелями. Генотип передается от родителей к потомству в соответствии с правилами классической генетики (свободная комбинаторика). Фенотип полностью определяется генотипом в соответствии с матрицей, задаваемой экспериментатором в каждом новом прогоне модели. В эксперименте, результаты которого изложены в данной статье, внешний облик особей задавался двумя комплементарными генами с неполным доминированием (каждое новое сочетание аллелей разных генов определяет новый вариант признака), а биотопические предпочтения – одним геном с неполным доминированием. Семь оставшихся генов определяли нейтральный признак по принципу полимерии (каждый ген имеет два аллеля со значениями “0” и “1”, значение признака определяется суммой значений аллелей всех генов), который не влиял на фенотип, но экспериментатор мог следить за его распределением в пространстве модели. Каждый цикл модели состоит из трех стадий: поиск партнера, размножение и расселение.
Поиск партнера. Каждая особь просматривает участки в радиусе двух участков от своего местонахождения (всего 24 участка) и ищет свободного партнера с внешним обликом, соответствующим облику одного из своих родителей (половой импринтинг). Если партнер найден и тоже выбрал данную особь в качестве партнера, то формируется брачная пара. Если после просмотра всех 24 участков партнер не найден, то данная особь осуществляет поиск еще раз, выбирая в качестве партнера любую особь без пары (принцип “вакуумной активности”; Панов, 1989). Особи, не сумевшие сформировать брачную пару, умирают. Особи, сформировавшие брачную пару, остаются каждый на своем участке и приступают к размножению.
Размножение. Брачная пара оставляет четырех потомков (по паре на каждом из участков родителей), после чего умирает. Каждая пара потомков запоминает облик родителя, на участке которого они находятся. У потомков формируется генотип в результате свободного комбинирования аллелей их родителей и фенотип согласно заданным экспериментатором правилам.
Расселение. Каждая новая особь перемещается в случайном направлении в пределах десяти участков от места своего рождения, после чего ищет участок в радиусе двух участков от своего нового местоположения (всего 25 участков). Если участок свободен и его биотоп оптимален для данной особи, она занимает его. Если биотоп не оптимален, особь выбирает его с вероятностью, задаваемой экспериментатором перед запуском модели. Если выбранный участок занят, особь вытесняет резидента с вероятностью 20%. Резидент, вытесненный с занятого им участка, умирает. Особи, не сумевшие занять свой индивидуальный участок, умирают.
Для каждого набора заданных условий модель реализовалась в пяти повторностях, чтобы оценить влияние стохастических факторов. Для каждого имитационного эксперимента фиксировались: ширина гибридной зоны, состав пар, число холостых особей в каждом фенотипическом классе в центре и по краям гибридной зоны. Эти данные экспортировались в файл Excel и подвергались статистической обработке в пакете программ Statistica 10.0.
РЕЗУЛЬТАТЫ
Эксперимент 1. Ассортативность спаривания и биотопическая сегрегация отсутствуют.
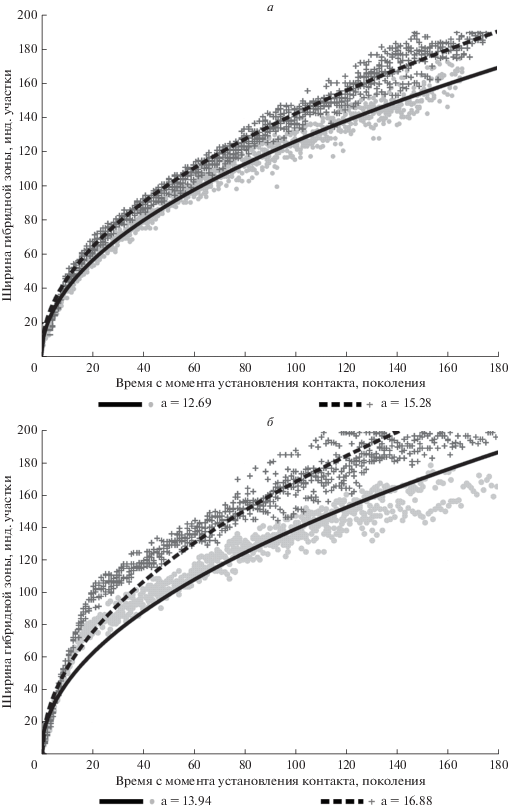
Гибридная зона расширяется пропорционально корню квадратному от времени вступления форм во вторичный географический контакт. При увеличении количества генов, определяющих анализируемый признак, скорость расширения гибридной зоны несколько увеличивается (рис. 1а). В данном эксперименте внешний облик особей является селективно нейтральным признаком, поскольку не влияет на выбор партнера (ассортативность спаривания отсутствует). Это дает возможность рассматривать признак внешнего облика, задаваемый двумя генами, и нейтральный признак, задаваемый семью генами, как эквивалентные.
Рис. 1.
Зависимость ширины гибридной зоны (количество индивидуальных участков) от времени с момента установления географического контакта (поколения) в условиях отсутствия ассортативности спаривания. а – отсутствие начальной биотопической сегрегации, б – наличие начальной биотопической сегрегации. Сплошная линия – фенотип задан двумя генами, пунктирная линия – фенотип задан семью генами. Уравнение регрессии: $y = a\surd x.$
Эксперимент 2. Ассортативности спаривания нет, биотопическая сегрегация есть. Для гетерозигот по гену биотопической сегрегации оба биотопа эквивалентны оптимальному.
Модель ведет себя сходным образом с экспериментом 1, но расширение зоны контакта происходит быстрее (рис. 1б). На начальных стадиях формирования зоны гибридизации она расширяется прямо пропорционально времени своего существования вследствие наличия биотопической сегрегации между родительскими формами. Поскольку гетерозиготы по гену биотопической сегрегации обладают селективным преимуществом перед гомозиготами (в отличие от последних, они могут занимать любой участок), зона контакта по этому признаку расширяется быстрее, чем по признаку брачной окраски. Как только на краях гибридной зоны биотопическая сегрегация между родительскими формами нивелируется, скорость расширения гибридной зоны замедляется. Под родительскими формами здесь понимается совокупность особей с внешним обликом, соответствующим внешнему облику двух исходных форм в начальный момент времени. По признаку биотопических предпочтений и нейтральному признаку они могут отличаться от исходных.
Эксперимент 3. Ассортативность спаривания есть, биотопической сегрегации нет.
Примерно к 30-му поколению после вступления гибридизирующих форм во вторичный контакт формируется узкая и стабильная во времени и пространстве гибридная зона шириной около 35 индивидуальных участков и долей фенотипических гибридов в центральной ее части около 20%. Доля особей, не сумевших сформировать брачную пару, составляет для родительских фенотипов 1–2% в центре гибридной зоны и 3–5% по краям; для гибридов – соответственно 5 и 8%. По нейтральному признаку формируется широкая клина на всем пространстве модели: при удалении от гибридной зоны доля аллелей, специфичных для данной формы, увеличивается.
Эксперимент 4. Есть ассортативность спаривания и биотопическая сегрегация. Для гетерозигот по гену биотопической сегрегации оба биотопа эквивалентны оптимальному.
Зона контакта сначала расширяется (максимальная ширина зоны контакта в различных имитационных экспериментах равна 110 индивидуальным участкам), но после примерно 60-го поколения начинает сужаться вплоть до формирования к 200-му поколению узкой и стабильной гибридной зоны шириной около 35 индивидуальных участков, как в эксперименте 3. Ключевым моментом в данной ситуации является то, что, как и в эксперименте 2, гибридная зона по признаку биотопической сегрегации расширяется быстрее, чем по признаку брачной окраски, поскольку гетерозиготы обладают преимуществом при выборе индивидуальных участков, имея возможность занимать любые местообитания. Как только по краям гибридной зоны появляются особи с внешним обликом родительских форм, но гетерозиготные по гену биотопических предпочтений, они начинают быстро расселяться, и биотопическая сегрегация между родительскими формами нивелируется. Таким образом, условия модели меняются и становятся похожими на условия в эксперименте 3. Соответственным образом меняется и поведение модели, приводя к формированию узкой и стабильной гибридной зоны по признаку брачной окраски. По нейтральному же признаку формы становятся неразличимыми на всей территории модели.
Эксперимент 5. Есть ассортативность спаривания и биотопическая сегрегация. Для гетерозигот по гену биотопической сегрегации оба биотопа являются субоптимальными.
Между родительскими формами образуется симпатрия на всей территории модели со случайной гибридизацией, доля фенотипических гибридов не превышает 10%.
В экспериментах 4 и 5 поведение модели зависит от того, с какой вероятностью особи занимают субоптимальный биотоп. Результаты исследования модели с различным значением этого параметра представлены на рис. 2.
ОБСУЖДЕНИЕ
По результатам анализа математических моделей можно выделить три категории генов в соответствии с характером их поведения в зоне вторичного контакта (Barton, Hewitt, 1985; Hewitt, 1988): 1. Гены, аллели которых имеют селективное преимущество в новом для них генетическом окружении, распространяются по всему ареалу гибридизирующих форм с постоянной скоростью (“адаптивная волна”); 2. Гены, аллели которых находятся под давлением отрицательного отбора в чужеродной генетической среде, формируют узкую и стабильную во времени гибридную зону (“напряженная” зона); 3. Клина, формируемая селективно нейтральными аллелями, расширяется пропорционально корню квадратному со времени формирования вторичного географического контакта (“нейтральная диффузия”).
Поведение нашей имитационной модели полностью определяется этими особенностями, однако полученные результаты позволяют прийти к нескольким не вполне очевидным выводам. Во-первых, узкая гибридная зона может формироваться в условиях нормальной, по сравнению с родительскими формами, приспособленности гибридов. Отбор против гибридов в данном случае обеспечивается ассортативностью спаривания: гибридам сложнее сформировать брачную пару из-за редкости своего фенотипического облика. Сила отбора невелика: в экспериментах с моделью особи с обликом родительских форм оставались холостыми в 1–3% случаев, а гибриды – в 5–8%. Однако этого оказалось достаточно не только для поддержания узости гибридной зоны, но и для того, чтобы отбор против гибридов приводил к постепенному сужению широкой зоны симпатрии после нивелирования биотопической сегрегации между родительскими формами. По всей видимости, сила отбора не имеет существенного значения: она влияет только на скорость процесса, но не на конечный результат.
Во-вторых, для завершения процесса видообразования (т.е. установления симпатрии между родительскими формами при сохранении их фенотипического облика и при низком уровне гибридизации) снижение приспособленности гибридов не является необходимым условием. Процесс видообразования можно считать завершенным при наличии существенных различий между дивергирующими формами, как во внешнем облике, так и в биотопических предпочтениях: гибриды должны быть менее успешны и при формировании брачных пар, и при занятии индивидуальных участков.
В большинстве экспериментов наша модель показала сходные результаты с моделью В.И. Грабовского. Когда биотопическая сегрегация и ассортативность спаривания отсутствуют, то в обеих моделях гибридная зона постепенно расширяется, причем скорость ее расширения падает со временем. А при отсутствии биотопической сегрегации и наличии ассортативности спаривания формируется узкая и стабильная во времени зона контакта, соответствующая математической модели “напряженной” зоны. Как уже отмечалось выше, стабильность такой зоны во времени поддерживается отбором против гибридов. Существенным отличием модели В.И. Грабовского является то, что экспериментатор может задать такие параметры ассортативности, которые будут полностью исключать гибридизацию. Тем не менее в этом предельном случае механизм поддержания узости зоны контакта принципиально такой же: на границе ареала репродуктивный успех снижается из-за того, что особям труднее найти конспецифичного партнера.
Принципиально различные результаты обе модели показывают лишь в том случае, когда между исходными формами наблюдаются как биотопическая сегрегация, так и ассортативность спаривания. В нашей модели происходит либо установление симпатрии между исходными формами при сохранении их первоначального внешнего облика и поддержании гибридизации на низком уровне (в случае, когда гибриды при занятии индивидуальных участков менее успешны, чем родительские формы). Либо зона контакта сначала расширяется, а потом начинает постепенно сужаться вплоть до формирования “напряженной” зоны. В модели же В.И. Грабовского происходит сначала расширение зоны контакта на все пространство модели, а потом постепенное нарастание уровня гибридизации вплоть до нивелирования генетических различий между гибридизирующими формами. В целом по результатам анализа этой модели получается парадоксальный вывод: формы, находящиеся на начальных стадиях дивергенции, вступая во вторичный географический контакт, сохраняют генетическую уникальность, а на поздних стадиях, практически достигнув видового уровня, наиболее подвержены риску генетического слияния через гибридизацию (Грабовский, 2002). Судя по характеру поведения моделей, столь явные различия в конечных результатах объясняются различными механизмами наследования признаков. В модели В.И. Грабовского как внешний облик, так и биотопические предпочтения определяются гибридным индексом, т.е. оба признака оказываются генетически сцепленными. В нашей же модели внешний облик особей и их биотопические предпочтения определяются различными генами, поэтому в пространстве модели оба признака формируют независимые клины. Из сопоставления моделей можно сделать вывод, что в случае генетического сцепления признаков брачной окраски и биотопических предпочтений при экологическом превосходстве гибридов от близкородственных форм, вступивших в географический контакт, мы вправе ожидать их слияния посредством гибридизации.
Отдельно следует обсудить понятия “узкой” и “широкой” зоны контакта и гибридизации. В имитационных компьютерных моделях ширину зоны удобно измерять в индивидуальных участках. В то же время анализ математических моделей показывает, что ширина, например, “напряженной” зоны определяется не размерами индивидуальных участков особей, а величиной натальной дисперсии (Hewitt, 1988): w ~ σ2/s, где σ – различия в дисперсии между взрослыми и молодыми особями, s – сила отбора против гибридов. Как показывают исследования, у мухоловки-пеструшки (Ficedula hypoleuca) на Куршской косе (Chernetsov et al., 2006) величина натальной дисперсии для самцов составляет 4.3 ± 0.6 км (n = 83), для самок – 6.6 ± 0.8 км (n = 43), т.е. приблизительно ее можно принять за 5 км, что, вероятно, справедливо для большинства мелких воробьиных птиц. В исследованиях нашей модели ширина “напряженной” стабильной во времени гибридной зоны составляла 35 участков при величине натальной дисперсии в 10 участков, а максимальный размер “широкой” зоны равнялся 110 участкам. Соответственно, если принять величину натальной дисперсии в 5 км, то получим для “узкой” зоны ширину в 20 км, а для “широкой” – 55 км. Иными словами, “широкая” гибридная зона является таковой только в сравнении с “напряженной” зоной, в масштабах же ареалов гибридизирующих форм она может выглядеть узкой.
Результаты данного исследования можно трактовать как модель видообразования (табл. 1). При всей ее схожести с тем, что предлагает биологическая концепция вида, между ними есть существенное различие: здесь критерием для выделения видов является не степень репродуктивной изоляции между анализируемыми таксонами (оцениваемая, например, по доле фенотипических гибридов в зоне контакта), а уровень морфологической дивергенции между ними, создающий основу для ассортативности спаривания. Кроме того, для гипотезы реинфорсмента, которая в рамках биологической концепции вида служит обоснованием дискретности видов (Рубцов, 2015), в нашей модели просто нет места: способность к гетероспецифичному спариванию не снижает репродуктивный успех особи, а, наоборот, повышает его, давая возможность оставить потомство в условиях недостатка конспецифичных партнеров.
Таблица 1.
Ситуация в зоне вторичного контакта в зависимости от уровня дивергенции гибридизирующих форм
| Стадия | Различия | Таксономический статус | Поведение гибридной зоны | |
|---|---|---|---|---|
| ВО | БС | |||
| 1 | + | – | Подвиды | Скорость расширения уменьшается со временем |
| 1а | + | + | Подвиды | Скорость расширения уменьшается со временем |
| 2 | ++ | – | Мегаподвиды | Узкая и стабильная во времени |
| 3 | ++ | + | Полувиды | Сначала расширяется, потом сужается до стадии 2 |
| 4 | ++ | ++ | Виды | Симпатрия, случайная гибридизация |
В представленной схеме видообразования (табл. 1) наибольший интерес представляет стадия 3. По формальным критериям – ширине гибридной зоны и доле фенотипических гибридов – она может быть неотличима от двух предыдущих стадий, но по уровню дивергенции она близка к завершению процесса видообразования. Наиболее же существенно то, что традиционный молекулярно-генетический способ оценки уровня дивергенции таксонов, основанный на количестве замен нуклеотидов в селективно нейтральных локусах, адекватно работающий для остальных стадий, в данном случае может дать некорректные результаты. Именно на этой стадии по результатам исследования модели уровень смешения геномов родительских форм по нейтральным локусам максимален. В результате с высокой долей вероятности мы не сможем выделить видоспецифичные аллели и ошибочно примем наблюдаемую картину изменчивости за проявление предкового полиморфизма.
Подводя итог вышесказанному, можно сказать, что результаты исследования имитационной модели гибридной зоны могут дать повод для ревизии теоретических основ биологической концепции вида и послужить основой для ее модернизации.
Автор выражает глубокую признательность В.И. Грабовскому за глубокий анализ работы и ценные замечания в ходе подготовки рукописи. Работа выполнена при финансовой поддержке РФФИ (гранты № 17-04-00903 и № 18-04-00770).
Список литературы
Алексеев Е.Р., Чеснокова О.В., Кучер Т.В., 2010. Free Pascal и Lazarus: Учебник по программированию. М.: ALT Linux; Издательский дом ДМК-пресс. 440 с.
Грабовский В.И., 1995а. Клеточные автоматы как простые модели сложных систем // Успехи соврем. биологии. Т. 115. № 4. С. 412–418.
Грабовский В.И., 1995б. Зоны вторичного контакта: эффекты отбора против гибридов, ассортативности спариваний и конкуренции // Журн. общ. биологии. Т. 56. № 3. С. 291–309.
Грабовский В.И., 2002. Зоны вторичного контакта: эффекты отбора против гибридов, ассортативности спариваний и конкуренции. http://nature.air.ru/models/contact.htm.
Добржанский Ф.Г., 2010. Генетика и происхождение видов. М.; Ижевск: Ин-т компьютерных исследований, НИЦ “Регулярная и хаотическая динамика”. 384 с.
Крюков А.П., Блинов В.Н., 1989. Взаимодействие серой и черной ворон (Corvus cornix L., C. corone L.) в зоне симпатрии и гибридизации: есть ли отбор против гибридов? // Журн. общ. биологии. Т. 50. № 1. С. 128–135.
Лавренченко Л.А., 2013. Гибридогенное видообразование у млекопитающих: иллюзия или реальность? // Журн. общ. биологии. Т. 74. № 4. С. 253–267.
Майр Э., 1968. Зоологический вид и эволюция. М.: Мир. 597 с.
Панов Е.Н., 1989. Гибридизация и этологическая изоляция у птиц. М.: Наука. 512 с.
Рубцов А.С., 2015. Репродуктивная изоляция и понятие вида у птиц // Зоол. журн. Т. 94. № 7. С. 816–831.
Arnold M.L., 1997. Natural Hybridization and Evolution. N.Y.: Oxford Univ. Press. 215 p.
Barton N.H., Hewitt G.M., 1985. Analysis of hybrid zones // Annu. Rev. Ecol. Syst. V. 16. P. 113–148.
Butlin R., 1987. Speciation by reinforcement // Trends Ecol. Evol. V. 2. № 1. P. 8–13.
Chernetsov N., Sokolov L.V., Kosarev V., Leoke D., Markovets M. et al., 2006. Sex-related natal dispersal of pied flycatchers: How far away from home? // Condor. V. 108. № 3. P. 711–717.
Coyne J.A., Orr H.A., 2004. Speciation. Sunderland, MA: Sinauer Associates. 545 p.
Hewitt G.M., 1988. Hybrid zones – natural laboratories for evolutionary studies // Trends Ecol. Evol. V. 3. № 7. P. 158–167.
Orr H.A., 1995. The population genetics of speciation: The evolution of hybrid incompatibilities // Genetics. V. 139. № 4. P. 1805–1813.
Poelstra J.W., Vijay N., Bossu C.M., Lantz H., Ryll B. et al., 2014. The genomic landscape underlying phenotypic integrity in the face of gene flow in crows // Science. V. 344. № 6190. P. 1410–1414.
Price T.D., 2008. Speciation in Birds. Greenwood Village, CO: Roberts and Company. 470 p.
Randler C., 2002. Avian hybridization, mixed pairing and female choice // Anim. Behav. V. 63. № 1. P. 103–119.
Randler C., 2008. Mating patterns in avian hybrid zones – a meta-analysis and review // Ardea. V. 96. № 1. P. 73–80.
Ten Cate C., 1991. Sexual imprinting, mate preferences and plumage evolution // Acta XX Congr. Intern. Ornithol. Wellington, New Zealand. P. 1081–1088.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии