

Журнал общей биологии, 2020, T. 81, № 1, стр. 47-53
Вариабельность величины отношения массы листа растения к его площади в условиях кратковременных ежесуточных понижений температуры
Е. Н. Икконен 1, *, Т. Г. Шибаева 1
1 Институт биологии КарНЦ РАН, ФИЦ “Карельский научный центр РАН”
185910 Петрозаводск, Пушкинская ул., 11, Россия
* E-mail: likkonen@gmail.com
Поступила в редакцию 12.07.2019
После доработки 19.09.2019
Принята к публикации 18.10.2019
Аннотация
Изучали влияние кратковременного (2 ч) ежесуточного понижения температуры (ДРОП-воздействие, от англ. drop – падение) до 12, 8, 4 или 1°С на величину отношения сухой массы листа к его площади (LMA, leaf mass per area) у теплолюбивых (огурец, томат, сладкий перец, баклажан) и холодостойких (озимая пшеница, цветная капуста) растений. На примере отдельных видов исследована роль светового фактора (интенсивность освещения растений в период действия ДРОП, световые условия роста растений и длительность фотопериода) в отклике параметра LMA на ДРОП-воздействия. Несмотря на высокую межвидовую вариабельность LMA (среди теплолюбивых видов величина LMA увеличивалась в ряду: огурец–томат–перец–баклажан), между видами не было выявлено существенных качественных различий в отклике LMA на ДРОП-воздействия. Внутривидовая вариабельность была обусловлена световыми и температурными условиями роста растений, при этом изменения температуры в диапазоне положительных значений влияли на вариабельность LMA у теплолюбивых, но не у холодостойких видов. Фактор света оказывал существенное влияние на изменчивость параметра LMA у теплолюбивых видов при ДРОП-воздействиях. Тогда как у растений, подвергавшихся ДРОП-воздействиям в темноте, величина LMA оставалась неизменной или повышалась, у растений, испытавших ДРОП на свету, она снижалась, причем степень изменения LMA увеличивалась с повышением интенсивности ДРОП. Более высокий уровень освещенности растений способствовал росту величины LMA, при этом ДРОП-воздействия в темноте повышали, а ДРОП-воздействия на свету снижали степень увеличения LMA с возрастанием интенсивности освещения растений. Увеличение фотопериода с 16 до 24 ч вызывало повышение величины LMA у растений, но при этом продолжительность фотопериода не влияла на отклик параметра LMA на ДРОП-воздействия. Таким образом, кратковременные периодические понижения температуры до низких положительных величин могут вызывать существенные изменения LMA у теплолюбивых, но не у холодостойких растений. При этом световые условия являются важным модулирующим фактором данных изменений. Высокая степень вариабельности параметра LMA при интенсивных ДРОП-воздействиях отражает способность растений к существенным структурным преобразованиям, необходимым для их роста и развития в изменяющихся температурных условиях.
Величина отношения сухой массы листа к его площади (LMA, leaf mass per area) считается одним их ключевых параметров, связанных с ростом растений (Lambers, Poorter, 1992). Показатель LMA, который в научной литературе может также называться SLW (specific leaf weight) или УППЛ (удельная поверхностная плотность листа), широко используется в экологических и сельскохозяйственных исследованиях, но в гораздо меньшей степени в исследованиях физиологии растений (Poorter et al., 2009). Последнее малооправданно, поскольку фотосинтетические способности растений во многом определяются величиной LMA, а вариабельность данной величины может рассматриваться как один из механизмов структурных преобразований, обеспечивающих рост устойчивости растений к изменениям условий среды.
Величина параметра LMA может существенно изменяться как на межвидовом, так и внутривидовом уровне (Poorter et al., 2009; Васфилов, 2011). Межвидовая вариабельность LMA может быть связана с различиями в величине доли апопласта в сухой массе листа вследствие особенностей структуры и функций листьев разных видов (Васфилов, 2011). Виды с низкими значениями LMA, как правило, лучше приспособлены к местообитаниям, богатым ресурсами, а виды с высокими значениями этого параметра чаще представительны в малопродуктивных местообитаниях, неблагоприятных для роста (Poorter et al., 2009). В пределах вида LMA варьирует в зависимости от возраста листа и фенологической фазы растений, а также от факторов окружающей среды, основными среди которых считаются свет, CO2, вода, минеральное питание и температура (Niinemets et al., 2004; Poorter et al., 2009; Васфилов, 2011). Значительное влияние на величину LMA может оказывать совместное действие холода и света с наложением эффектов данных факторов. Раздельное действие пониженной температуры и высокой освещенности способствуют увеличению LMA (Трунова, 2007; Poorter et al., 2009; Васфилов, 2011), но их совместное действие может вызывать повреждения фотосинтетического аппарата (Allen, Ort, 2001) и, как следствие, снижение поступления ассимилятов для формирования структуры листа и уменьшение LMA.
Несмотря на нестабильность температуры в природных условиях, отклик растений на кратковременные (часы) ежесуточные понижения температуры (ДРОП-воздействия, от англ. drop – падение) менее изучен, чем их отклик на длительное (недели и более) постоянное действие низкой температуры (ПНТ) (Марковская и др., 2000; Шибаева и др., 2019). При этом известно, что реакции растений на ДРОП и ПНТ-воздействия могут различаться не только количественно, но и качественно (Шибаева и др., 2018). Многократно подтверждалось, что длительное действие холода инициирует структурные преобразования в листе, вызывающие повышение LMA (Кислюк и др., 1983; Atkin et al., 2006; Трунова, 2007; Gorsuch et al., 2010), но о влиянии ДРОП-воздействий на данный параметр известно мало (Икконен и др., 2015). Кроме того, вопрос о совместном влиянии света и ДРОП-воздействий на LMA растений остается пока открытым, несмотря на то, что растения довольно часто подвергаются действию ДРОП в светлое время. В задачи данного исследования входила оценка влияния ДРОП-воздействий различной интенсивности на параметр LMA у теплолюбивых (огурец, томат, сладкий перец, баклажан) и холодостойких (озимая пшеница, цветная капуста) видов и выявление роли светового фактора (наличие/отсутствие света во время ДРОП-воздействий, световые условия роста растений и продолжительность фотопериода) в отклике LMA на ДРОП-воздействия.
МАТЕРИАЛЫ И МЕТОДЫ
Эксперимент 1. Влияние интенсивности ДРОП-воздействий на LMA
Растения огурца (Cucumis sativus L., гибрид Кураж F1), томата (Solanum lycopersicum L., гибрид Верлиока F1), сладкого перца (Capsicum annuum L., с. Нежность) и цветной капусты (Brassica oleracea var. botrytis L., с. Снежный шар) выращивали в камере искусственного климата (Vötsch, Германия) при поливе питательным раствором (рН 6.2–6.4), температуре воздуха 23°С днем и 20°С ночью, освещенности 300 мкмоль/(м2 с) ФАР, фотопериоде 16 ч, влажности воздуха 70%. В возрасте 7 сут, когда только начинал формироваться первый лист, растения огурца были разделены на пять групп, четыре из которых подвергали ДРОП-воздействиям. Первую группу ежедневно в течение 13 сут переносили на 2 ч в камеру искусственного климата (Snijders Microclima 1750, Нидерланды) с температурой 12°C (вариант ДРОП 12), вторую − 8°C (ДРОП 8), третью − 4°C (ДРОП 4), четвертую − 1°C (ДРОП 1), сохраняя прочие условия неизменными. Внутри каждой группы растения разделяли на две подгруппы, одну из которых переносили в низкотемпературные условия в конце ночного периода, а вторую − в начале светового периода. Для сравнительной оценки влияния ДРОП-воздействий и постоянного действия низкой температуры (ПНТ) на исследуемый параметр пятая группа растений огурца в возрасте 14 сут была перенесена на 6 сут в камеру с температурой 12°C, фотопериодом 16 ч и ФАР, равной 100 мкмоль/(м2 с) (вариант ПНТ). Шестая группа растений, которая не подвергалась низкотемпературным воздействиям, служила контролем. Растения томата и перца в возрасте 3–4 настоящих листьев в течение 13 сут подвергали ДРОП-воздействиям, снижая температуру на 2 ч до 12, 8 и 4°C (варианты ДРОП 12, ДРОП 8, ДРОП 4). Растения томата и перца вариантов ДРОП 8 и растения томата варианта ДРОП 4 разделяли на две подгруппы для переноса в условия низкой температуры в конце ночного или начале светового периода. Растения цветной капусты в фазе 3 листьев в течение 13 сут подвергали действию температуры 4 и 1°С (варианты ДРОП 4, ДРОП 1) в течение 2 ч в конце ночного периода. Растения, не подвергавшиеся ДРОП-воздействиям, служили контролем.
Проростки озимой пшеницы (Triticum aestivum L., с. Московская-39) выращивали в рулонах фильтровальной бумаги на питательном растворе в камере Vötsch при температуре 22°С, 150 мкмоль/(м2 с) ФАР, фотопериоде 12 ч, влажности воздуха 70%. Часть проростков пшеницы с возраста 7 дней ежедневно в течение 6 сут переносили в конце ночи на 3 ч в камеру с температурой 4°C (вариант ДРОП 4), вторую часть проростков выращивали в течение 6 сут при температуре 4°C, 100 мкмоль/(м2 с) ФАР и прочих сохраненных условиях (вариант ПНТ), а третья, не подвергавшаяся действию холода, служила контролем.
Эксперимент 2. Влияние уровня освещения на LMA огурца в условиях ДРОП-воздействий
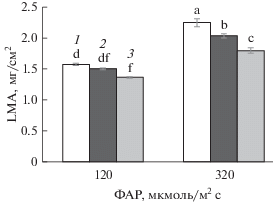
Растения огурца (Cucumis sativus L., гибрид Зозуля F1) выращивали в камере Vötsch при температуре 23°С, фотопериоде 12 ч, влажности воздуха 70% и освещенности 120 мкмоль/(м2 с) или 320 мкмоль/(м2 с) ФАР. Когда первый лист достигал 60–70% от своей максимальной площади, растения в течение 8 сут подвергали 2-часовому действию температуры 9°C в конце ночного периода или в начале дневного периода, сохраняя прочие условия неизменными. Растения, не подвергавшиеся ДРОП-воздействиям, были приняты за контроль.
Эксперимент 3. Влияние продолжительности фотопериода на LMA в условиях ДРОП-воздействий
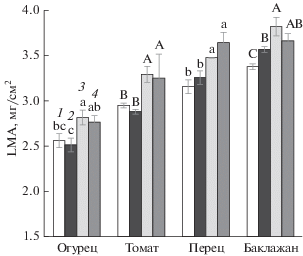
Растения огурца (Cucumis sativus L., гибрид Зозуля F1), сладкого перца (Capsicum annuum L., с. Нежность), томата (Solanum lycopersicum L., гибрид Верлиока F1) и баклажана (Solanum melongena L., с. Алмаз) выращивали в камерах искусственного климата при температуре 26°С, 150 мкмоль/(м2 с) ФАР, фотопериоде 16 или 24 ч. Часть растений, выращиваемых при 16- или 24-часовом фотопериоде, ежедневно в конце ночи переносили на 2 ч в камеру с температурой 10°С.
Отбор растительного материала для определения LMA был выполнен через сутки после окончания последнего ДРОП или ПНТ-воздействия. После окончания всех низкотемпературных воздействий растения находились в камере Vötsch при первоначально заданных оптимальных свето-температурных условиях. Из закончивших рост листовых пластинок огурца, томата, перца, баклажана и цветной капусты острозаточенным стальным цилиндром вырезали диски площадью 2.6 см2. Отобранные высечки листовых пластинок высушивали при 105°С до постоянного значения веса. Величину LMA растений огурца, томата, перца, баклажана и цветной капусты рассчитывали как отношение сухой массы высечки листовой пластинки к ее площади. У растений пшеницы определяли площадь листьев и их сухую массу путем высушивания при температуре 105°С. Величину LMA для пшеницы рассчитывали как отношение сухой массы листьев к их площади.
На рис. 1–4 представлены результаты в виде средних значений (6 и более повторностей в каждом варианте отдельного опыта) и их стандартных ошибок. Достоверность различий между средними значениями исследованных параметров определяли при P < 0.05 на основе дисперсионного анализа (LSD тест) с использованием программного обеспечения Statistica (v. 8.0.550.0, “StatSoft Inc.”).
Рис. 1.
Величина LMA у растений огурца (а), томата (б) и сладкого перца (в), не подвергавшихся (контроль) и подвергавшихся кратковременным (2 ч) ежесуточным (6 сут) понижениям температуры до 12°С (ДРОП 12), 8°С (ДРОП 8), 4°С (ДРОП 4) и 1°С (ДРОП 1) в конце темнового (1) или начале светового (2) периода и продолжительному (6 сут) действию низкой (12°С) температуры (ПНТ). Здесь и на рис. 2–4 разные буквы указывают на достоверность различий средних значений при Р < 0.05. * означает достоверность различий средних значений величины LMA у растений, подвергавшихся ДРОП-воздействиям в темноте (1) и на свету (2).
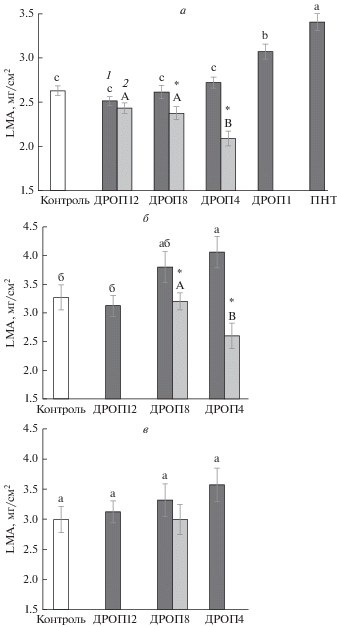
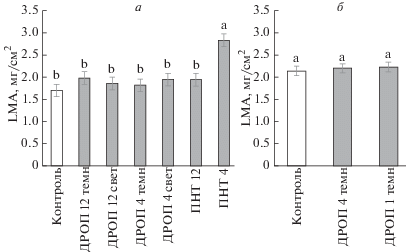
Рис. 2.
Величина LMA у растений пшеницы (а) и цветной капусты (б), не подвергавшихся (контроль) и подвергавшихся кратковременным (2 ч) ежесуточным (6 сут) понижениям температуры до 12°С (ДРОП 12), 4°С (ДРОП 4) или 1°С (ДРОП 1) в темноте (ДРОП темн) или на свету (ДРОП свет), а также продолжительному (6 сут) действию температуры 12°С (ПНТ 12) или 4°С (ПНТ 4).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Влияние кратковременных ежесуточных понижений температуры (ДРОП-воздействий) на величину LMA огурца, томата и сладкого перца зависело не только от значений, до которых опускается температура при ДРОП-воздействиях, но в большей степени от того, в темноте или на свету они происходили (рис. 1, 3). Тогда как растения, подвергавшиеся ДРОП-воздействиям в темноте, демонстрировали тенденцию к повышению LMA, у растений, испытавших ДРОП на свету, величина LMA существенно снижалась, т.е. наличие света во время понижений температуры у теплолюбивых растений качественно меняло реакцию LMA на ДРОП-воздействия. Следует отметить, что статистически достоверное влияние ДРОП в темноте на величину LMA выявлено у растений огурца и томата только при понижении температуры до значений из зоны повреждения (1 и 4°С). Так, величина LMA повышалась на 24% при ДРОП-воздействиях с использованием температуры 4°С (ДРОП 4) у томата и на 17% при снижении температуры до 1°С (ДРОП 1) у огурца (рис. 1а). У холодостойких видов (озимая пшеница и цветная капуста) чувствительность LMA к ДРОП отсутствовала независимо от воздействующей температуры (рис. 2).
Длительный рост растений в условиях пониженных температур, как правило, способствует утолщению листа и, соответственно, увеличению LMA (Кислюк и др., 1983; Трунова, 2007), что подтвердили результаты данной работы, показавшие максимальный среди опытных вариантов рост величины LMA у листьев огурца и пшеницы (рис. 2) в вариантах ПНТ. Утолщение листьев, выросших при пониженных температурах, часто связывают с увеличением размеров клеток палисадной и губчатой части мезофилла, а также с повышением количества слоев мезофильных клеток (Gorsuch et al., 2010). Данные структурные преобразования, увеличивающие толщину фотосинтезирующей ткани, могут частично способствовать поддержанию высокой интенсивности фотосинтеза растений в условиях холода (Campbell et al., 2007; Gorsuch et al., 2010). Выявленное в данной работе увеличение LMA у теплолюбивых видов может быть также связано с уменьшением площади листьев в условиях ДРОП (Шибаева и др., 2019), что сопровождается уплотнением упаковки клеток из-за снижения их размера и, следовательно, увеличением вклада апопласта в LMA. Также повышение значений LMA в условиях интенсивных темновых ДРОП-воздействий могло быть следствием накопления в листьях крахмала из-за ограничения потребностей зон роста в фотоассимилятах.
В общей тенденции повышения LMA при ДРОП-воздействиях в темноте исключение составили результаты, полученные для растений огурца в варианте ДРОП 12, у которых значения исследуемого параметра были на 6% ниже контрольных (рис. 1а). Это соответствует результатам более раннего исследования, показавшего усиление фотосинтетической активности растений огурца под влиянием ДРОП-воздействий (Икконен и др., 2015). Более низкие значения LMA могут способствовать росту интенсивности фотосинтеза посредством снижения затрат на формирование листа и потерь световой энергии, повышения доли фотосинтетически активных тканей по отношению к объему клеточных стенок в сухой массе (Васфилов, 2011) или увеличения проводимости СО2 к строме хлоропластов (Poorter et al., 2009).
При действии ДРОП на свету величина LMA уменьшалась пропорционально снижению значения действующей на растения температуры (рис. 1а, б). Причем степень изменения величины LMA в результате ДРОП-воздействий на свету была выше, чем при темновом ДРОП. Возможно, что снижение значений LMA при ДРОП-воздействиях на свету связано с ингибированием фотосинтеза, как это было показано ранее на примере растений огурца (Ikkonen et al., 2017), что может ограничивать поступление ассимилянтов, используемых для повышающих LMA структурных преобразований.
Уровень освещенности листа в период его роста является одним из самых значимых факторов, определяющих вариабельность LMA (Poorter et al., 2009). Повышение уровня освещенности, приводящее к увеличению суточного интеграла освещения, вызывает повышение величины LMA (Poorter et al., 2009; Васфилов, 2011), что подтвердили результаты данной работы, полученные для растений огурца, которые выращивали при двух уровнях ФАР − 120 и 320 мкмоль/(м2 с) (рис. 3). Значения LMA были значительно ниже у растений, выросших при 120 мкмоль/(м2 с) ФАР, независимо от того, подвергались они или нет действию низкой температуры, однако действия ДРОП модифицировали вариабельность LMA при изменении световых условий роста растений, при этом степень изменения параметра зависела от того, на свету или в темноте происходили ДРОП-воздействия. Так, ДРОП в темноте на 18% повышал, а ДРОП на свету на 14% снижал процент увеличения LMA с изменением интенсивности ФАР от 120 до 320 мкмоль/(м2 с).
Рост значений LMA с повышением освещенности связывают как с увеличением плотности листа вследствие более плотной упаковки клеток палисадной ткани и повышенного содержания неструктурных углеводов (Niinemets et al., 1998), так и с утолщением листа (Poorter et al., 2009). Уровень освещенности, при котором росли растения, влиял на отклик параметра LMA на ДРОП-воздействия в большей степени, когда растения подвергались им в темновой период (рис. 3). Если световой ДРОП вызывал снижение LMA независимо от освещенности растений, то в результате темновых ДРОП величина LMA снижалась в условиях низкой ФАР и проявляла тенденцию к повышению при более высокой ФАР. Следует отметить, что сходное влияние ДРОП-воздействия оказали и на интенсивность фотосинтеза растений огурца (Икконен и др., 2015), что выявляет тесную взаимосвязь между структурной организацией листа и активностью его фотосинтетического аппарата.
Увеличение фотопериода с 16 до 24 ч вызывало повышение значений LMA у растений огурца и перца на 10%, а у томата и баклажана на 13% (рис. 4), независимо от того, подвергались или нет растения ДРОП-воздействиям. Как и при росте растений в условиях высокой освещенности, влияние продолжительности светового периода на вариабельность параметра может быть связано с накоплением в листьях крахмала, повышением толщины и плотности палисадной ткани (Васфилов, 2011). В наших опытах с фотопериодами разной продолжительности освещенность растений была одинаковой, но значения LMA в условиях более продолжительного фотопериода были выше. Эти результаты еще раз подтверждают наблюдения, показавшие, что именно суточный интеграл освещения, а не уровень освещенности, оказывает наибольшее влияние на величину LMA (Chabot et al., 1979; Niinemets et al., 2004).
Исследованные теплолюбивые виды растений различались между собой по величине LMA. Этот показатель увеличивался в ряду: огурец–томат–перец–баклажан (рис. 4). Между видами не было, однако, выявлено существенных качественных различий в отклике LMA на изменение температурных и световых условий роста.
В отличие от теплолюбивых видов, которые в данном исследовании подвергались кратковременным воздействиям температуры как из зоны их закаливания (12 и 8°С), так и повреждения (4 и 1°С), холодостойкие виды испытывали понижения температуры только до значений из зоны их закаливания, но не повреждения (температуры ниже нуля). ДРОП-воздействия температурами из зоны закаливания не влияли на LMA у растений независимо от отношения исследованных видов к холоду. Ранее также было определено, что, несмотря на значительные качественные и количественные различия в реакциях холодостойких и теплолюбивых растений на постоянное продолжительное действие пониженных температур, их реакции на кратковременные периодические понижения температуры до низких положительных величин в целом качественно схожи (Шибаева и др., 2019). Как у теплолюбивых, так и у холодостойких растений ДРОП-воздействия температурой из зоны закаливания не приводят к значимым изменениям фотосинтетической активности и скорости накопления биомассы, однако вызывают морфогенетический эффект, способствующий увеличению компактности растений. Ранее было определено, что при температурах ДРОП из зоны повреждения у теплолюбивых растений происходят изменения, приводящие к увеличению соотношения дыхания и фотосинтеза, что сопровождается снижением скорости накопления растением надземной биомассы, уменьшением высоты растений и площади листьев (Икконен и др., 2018). В данной работе также изменения LMA у теплолюбивых растений наблюдались только при температурах ДРОП-воздействий, которые считаются повреждающими при их длительном действии.
ЗАКЛЮЧЕНИЕ
Результаты исследования величины отношения массы листа растения к его площади (LMA) на примере четырех теплолюбивых (огурец, томат, сладкий перец, баклажан) и двух холодоустойчивых (озимая пшеница, цветная капуста) видов показали ее межвидовую и внутривидовую вариабельность. В пределах положительных значений температурные условия роста определяли вариабельность LMA у теплолюбивых, но не у холодостойких видов. Тогда как продолжительное действие низкой температуры (ПНТ) стимулировало повышение величины LMA, эффект кратковременных ежесуточных понижений температуры (ДРОП-воздействий) зависел от световых условий, при которых растения подвергались действию ДРОП. Величина LMA снижалась, если действия ДРОП осуществлялись на свету, и увеличивалась, если растения подвергались ДРОП-воздействиям в темноте. Степень изменения LMA повышалась с усилением интенсивности ДРОП, т.е. со снижением действующей на растения низкой температуры, при этом значимые изменения величины LMA у теплолюбивых растений достигались только при температурах ДРОП, считающихся повреждающими при их длительном действии на растения (4 и 1°С), а максимальная степень изменения LMA достигалась в условиях ПНТ. Фактор света оказывал существенное влияние на вариабельность параметра LMA у теплолюбивых видов при действии ДРОП. Так, эффекты ДРОП на свету и в темноте качественно различались, и присутствие света во время ДРОП-воздействий усиливало степень изменения LMA. Кроме того, уровень освещенности растений в период их выращивания оказывал влияние на отклик LMA на ДРОП: эффект действия ДРОП в темноте был более выражен при низком уровне освещения, а эффект ДРОП на свету – при более высокой освещенности. Продолжительность фотопериода мало повлияла на отклик LMA на ДРОП-воздействия, однако значения LMA были выше в условиях более продолжительного фотопериода, видимо благодаря более высокому суточному интегралу освещения. Очевидно, что кратковременные ежесуточные понижения температуры в темноте до значений 8–12°С не оказывают существенного влияния на значения LMA, однако понижения температуры до значений, близких к нулевым, могут вызывать существенные изменения величины LMA у теплолюбивых растений. При этом световые условия являются важным модифицирующим фактором данных изменений.
Работа выполнена в рамках государственного задания Карельского научного центра РАН (№ 0218-2019-0074) и при частичной поддержке Российского фонда фундаментальных исследований (№ 19-29-05174). В работе использовано оборудование Центра коллективного пользования ФИЦ “Карельский научный центр РАН”.
Список литературы
Васфилов С.П., 2011. Анализ причин изменчивости отношения сухой массы листа к его площади у растений // Журн. общ. биологии. Т. 72. № 6. С. 436–454.
Икконен Е.Н., Шибаева Т.Г., Титов А.Ф., 2015. Реакция фотосинтетического аппарата листа у Cucumis sativus L. на кратковременное ежесуточное понижение температуры // Физиология растений. Т. 62. № 4. С. 528–532.
Икконен Е.Н., Шибаева Т.Г., Титов А.Ф., 2018. Влияние кратковременных ежесуточных понижений температуры на соотношение дыхания и фотосинтеза у теплолюбивых растений // Физиология растений. Т. 65. № 1. С. 63–69.
Кислюк И.М., Васьковский М.Д., Буболо Л.С., Палеева Т.В., 1983. Влияние температуры на строение листьев и фотосинтез Сarex lugens (Cyperaceae) и Arctagrostis arundinacea (Poaceae) // Ботан. журн. Т. 68. С. 1325–1332.
Марковская Е.Ф., Сысоева М.И., Харькина Т.Г., Шерудило Е.Г., 2000. Влияние кратковременного снижения ночной температуры на рост и холодостойкость растений огурца // Физиология растений. Т. 47. № 4. С. 511–515.
Трунова Т.И., 2007. Растение и низкотемпературный стресс. М.: Наука. 54 с.
Шибаева Т.Г., Шерудило Е.Г., Титов А.Ф., 2018. Реакция растений Cucumis sativus L. на длительное постоянное и кратковременные ежесуточные воздействия низких температур // Физиология растений. Т. 65. № 2. С. 68–77.
Шибаева Т.Г., Икконен Е.Н., Шерудило Е.Г., Титов А.Ф., 2019. Влияние ежесуточных кратковременных понижений температуры на теплолюбивые и холодостойкие растения // Физиология растений. Т. 66. № 4. С. 266–276.
Allen D.J., Ort D.R., 2001. Impact of chilling temperatures on photosynthesis in warm-climate plants // Trends Plant Sci. V. 6. № 1. P. 36–42.
Atkin O.K., Loveys B.R., Atkinson L.J., Pons. T.L., 2006. Phenotypic plasticity and growth temperature: Understanding interspecific variability // J. Exp. Bot. V. 57. № 2. P. 267–281.
Campbell C., Atkinson L., Zaragoza-Castells J., Lundmark M., Atkin O., Hurry V., 2007. Acclimation of photosynthesis and respiration is asynchronous in response to changes in temperature regardless of plant functional group // New Phytol. V. 176. № 2. P. 375–389.
Chabot B.F., Jurik T.W., Chabot J.F., 1979. Influence of instantaneous and integrated light-flux density on leaf anatomy and photosynthesis // Am. J. Bot. V. 66. № 8. P. 940–945.
Gorsuch P.A., Pandey S., Atkin O.K., 2010. Temporal heterogeneity of cold acclimation phenotypes in Arabidopsis leaves // Plant Cell Environ. V. 33. № 2. P. 244–258.
Ikkonen E.N., Shibaeva T.G., Titov A.F., 2017. The role of light in cucumber plant response to a diurnal short-term temperature drop // J. Stress Physiol. Biochem. V. 13. № 2. P. 35–44.
Lambers H., Poorter H., 1992. Inherent variation in growth rate between higher plants: A search for physiological causes and ecological consequences // Adv. Ecol. Res. V. 23. P. 187–261.
Niinemets Ü., Kull O., Tenhunen J.D., 1998. An analysis of light effects on foliar morphology, physiology, and light interception in temperate deciduous woody species of contrasting shade tolerance // Tree Physiol. V. 18. № 10. P. 681–696.
Niinemets Ü., Kull O., Tenhunen J.D., 2004. Within-canopy variation in the rate of development of photosynthetic capacity is proportional to integrated quantum flux density in temperate deciduous trees // Plant Cell Environ. V. 27. № 3. P. 293–313.
Poorter H., Niinemets U., Poorter L., Wright I.J., Villar R., 2009. Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis // New Phytol. V. 182. № 3. P. 565–588.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии