

Журнал общей биологии, 2020, T. 81, № 3, стр. 194-207
Единство популяции как способ выживания в нестабильной среде
А. К. Крылов 1, *, А. В. Марков 2, Ю. И. Александров 1, 3
1 Институт психологии РАН
129366 Москва, Ярославская ул., 13, Россия
2 Московский государственный университет им. М.В. Ломоносова, биологический факультет,
кафедра биологической эволюции
119234 Москва, Ленинские горы, 1, Россия
3 Московский Государственный психолого-педагогический университет
127051 Москва, Сретенка, 29, Россия
* E-mail: neuru@mail.ru
Поступила в редакцию 08.10.2019
После доработки 16.02.2020
Принята к публикации 25.02.2020
Аннотация
В настоящей работе при помощи компьютерного моделирования изучается влияние полиморфизма, специализации, кооперации и единства популяции на ее динамику в нестабильной среде. Рассмотрены мономорфные и полиморфные популяции, специализация и генерализация особей по отношению к ресурсным нишам, возможность перераспределения ресурсов между особями. Изучалась динамика численности популяций в стабильных и нестабильных условиях с разной амплитудой колебаний количества доступного ресурса. Показано, что в нестабильной среде единая генерализованная (полиморфная) популяция, занимающая все ресурсные ниши, получает преимущество по сравнению с набором независимых популяций, специализированных каждая под свою нишу. Обсуждаются социальные и экономические аналоги полученных результатов, свидетельствующие в пользу эффективности единства и кооперации членов сообщества в нестабильных средах.
Биоразнообразие можно рассматривать на разных уровнях, в том числе на уровне видов (видовое разнообразие, количество видов в сообществе), популяций и внутрипопуляционных группировок (фенотипическое разнообразие, количество фенотипических классов). Предполагается, что обе величины зависят от степени нестабильности среды и меняются разнонаправленно: в нестабильной среде оптимальным является более высокое фенотипическое разнообразие и меньшее видовое, сравнительно с ситуацией в более стабильной среде (Алещенко, Букварева, 1991; Букварева, Алещенко, 2010). Известна гипотеза о выгоде видового разнообразия в нестабильной среде (biodiversity insurance hypothesis, “страховая гипотеза”) (Yachi, Loreau, 1999), согласно которой в нестабильных условиях из-за колебаний параметров среды разные виды доминируют по очереди и ни один из них не может победить остальные в конкурентной борьбе. Нестабильность среды ослабляет межвидовую конкуренцию, стабилизирует или повышает суммарную продуктивность многовидового сообщества, а также способствует фенотипическому разнообразию (полиморфизму в популяции) (Алещенко, Букварева, 1991; Norberg et al., 2001; D’Odorico et al., 2008; Букварева, Алещенко, 2010; Eisenhauer et al., 2012; Isbell et al., 2015; см. также обзор: Букварева, Алещенко, 2013). Видовое разнообразие может быть связано с внутривидовым (фенотипическим) обратной зависимостью (Букварева, Алещенко, 2010), возможно, в связи с конкурентными отношениями между видами и кооперативными отношениями внутри полиморфного вида. К кооперативным отношениям внутри полиморфного вида (если их критерием считать взаимную пользу от наличия разных фенотипических классов) можно отнести распределение потомков каждого фенотипа по разным фенотипическим классам и перераспределение добытого одним фенотипом ресурса по всей популяции (см. ниже): в обоих случаях идет перераспределение ресурса или биомассы вида (популяции) по фенотипическим классам. Такой взгляд позволяет проводить аналогию и с человеческим сообществом: например, в более стабильном климате обнаруживаются страны с превалированием конкуренции в обществе, а в нестабильном – страны с превалированием кооперации (Кирдина и др., 2015).
В настоящей работе рассматривается гипотеза о том, что нестабильность среды является важным фактором эволюции биоразнообразия. Если рассматривать вопрос на уровне одного вида, то его можно сформулировать так: какие условия среды (стабильные или нестабильные) будут благоприятствовать поддержанию высокого уровня внутривидового полиморфизма, а какие – распаду вида на независимые мономорфные популяции, которые в дальнейшем смогут эволюционировать самостоятельно?
При такой постановке вопроса мы имеем в виду, что условия среды включают в себя и влияние других видов (хищников, жертв, паразитов). Нестабильность среды может выражаться в колебаниях климата или численностей хищников и жертв. “Кризисную ситуацию” в среде можно представить себе как снижение количества доступного ресурса (вызванное ухудшением климата или снижением численности жертв) или как повышение численности хищников. Степень нестабильности среды можно оценивать по силе и частоте таких кризисов. Конкретизируя вопрос о влиянии стабильности среды на биоразнообразие, рассмотрим гипотезу о том, что в нестабильной среде оптимальным является более высокое фенотипическое разнообразие популяции, чем в стабильной среде. Такое предположение сходно с гипотезой о повышении внутрипопуляционного фенотипического разнообразия в нестабильной среде (Алещенко, Букварева, 1991). Под фенотипическим разнообразием упомянутые авторы понимают не разную морфологию особей, а разнообразие ресурсных ниш и, соответственно, оптимальных для разных особей условий среды. В нашей модели используется такое же понимание фенотипических классов в полиморфной популяции. Для проверки гипотезы мы в ходе моделирования сравним численность такой полиморфной популяции с численностью мономорфной популяции, варьируя параметр нестабильности среды. Если в нестабильной среде численность полиморфной популяции будет значительно выше, чем численность мономорфной, это будет свидетельствовать о том, что полиморфизм дает преимущество в нестабильной среде.
Задача исследования, таким образом, состоит в том, чтобы выяснить, при каких условиях среды более эффективной (эффективность будем оценивать по численности) оказывается единая (фенотипически разнообразная, полиморфная) популяция, покрывающая все ресурсные ниши, а когда эффективнее набор независимых специализированных (фенотипически однообразных, мономорфных) популяций, каждая из которых адаптирована к своей ресурсной нише.
МЕТОДИКА (ОПИСАНИЕ МОДЕЛИ)
Отметим, что “реализованная ниша (вида) структурирована. Она состоит из субниш популяций, различающихся условиями их существования на ареале вида” (Северцов, 2013, с. 246). Разобьем всю совокупность особей одного вида на “функциональные фенотипические классы” по их ресурсным нишам, как это предложено в работе Г.М. Алещенко и Е.Н. Букваревой (1991), или, что аналогично, по локациям с разными ресурсно-климатическими условиями, к которым те или иные группы особей могут быть адаптированы (Ranta et al., 1997; Stenseth et al., 1999; Davis, Shaw, 2001). Рассмотрим ситуации с одной и с четырьмя ресурсными нишами (число четыре выбрано для простоты иллюстраций, можно выбрать другое число больше единицы, результаты не изменятся качественно). Ресурсные ниши можно представить себе как разные локации с одним видом ресурса или как разные виды ресурсов, важно лишь, чтобы колебания количества ресурса в каждой нише были независимыми от других (ниши с синхронно колеблющимся количеством ресурса влияют на результаты моделирования как единая ниша). Амплитуда колебаний количества ресурса в нише является мерой нестабильности среды: в стабильной среде колебания отсутствуют, в нестабильной они велики по амплитуде. Подобная нестабильность в природе может быть вызвана, например, климатическими колебаниями (Stenseth et al., 1999), при которых чем ближе локации, тем синхроннее колебания численности популяций вида в этих локациях (Ranta et al., 1997); колебаниями численности хищников или паразитов для данной популяции (или данного фенотипического класса), что также приводит к колебаниям численности как отдельных внутривидовых группировок, так и вида в целом (Krebs et al., 2001); ростом численности самой популяции, что приводит к исчерпанию ресурсов и последующему резкому спаду численности (Коротаев и др., 2015). В нашей модели колебания количества ресурса в одной нише, вызванные нестабильностью среды, приводят к колебаниям численности особей соответствующего фенотипического класса, “привязанного” к этой нише. Если в популяции нет внутреннего фенотипического разнообразия (ситуация “одна ниша”), то все особи и их потомки относятся к одному фенотипическому классу и находятся в одной нише. Если фенотипическое разнообразие есть (ситуация “четыре ниши”), то считаем, что потомки особей из каждой ниши равномерно распределяются по всем четырем нишам и там фиксируются. Такое равномерное распределение потомков по всем нишам можно считать результатом их миграции из ниши своих родителей в другие ресурсные ниши, не рассматривая ее динамику.
Нестабильность среды рассмотрим в двух вариантах: во-первых, как снижение количества ресурса в нише (аналог снижения численности жертв); во-вторых, как гибель определенной доли популяции этого фенотипического класса (аналог увеличения количества хищников). В обоих случаях будем в явном виде учитывать количество доступного ресурса в нише. Вопрос о законе размножения популяции является дискуссионным, есть несколько известных моделей, при этом классическая модель Ферхюльста (Ризниченко, Рубин, 1993) считается актуальной и при современном критическом анализе (Недорезов, 2015). Эта модель подходит для решения наших задач, поскольку в ней явно задается ресурсная емкость среды, что позволяет изучать динамику популяции при ограниченном ресурсе (Gonzalez, Holt, 2002; Andras et al., 2007; D’Odorico et al., 2008; Нефедов, 2012; Коротаев и др., 2015; Pereda et al., 2017).
В настоящей работе с помощью математического моделирования оценим, какова эффективность популяции в стабильных и нестабильных условиях, учитывая следующие факторы: нишевое разнообразие особей в популяции (аналог фенотипического разнообразия), степень специализации/генерализации особей, возможность перераспределения ресурсов между особями. Эффективность популяции оценим как среднюю за период времени численность популяции в определенной среде.
Нишевое разнообразие популяции представим следующим образом. Рассмотрим единую полиморфную популяцию, покрывающую все четыре ресурсные ниши и состоящую из четырех фенотипических классов, потомки которой равномерно заполняют все четыре класса (обозначим эту ситуацию как С4). Эту популяцию мы сопоставим по численности с четырьмя независимыми мономорфными популяциями, каждая из которых специализирована под свою нишу; потомки остаются в той же нише, что и родители (С1) (рис. 1). Такое сопоставление полиморфной популяции с несколькими мономорфными сделано для возможности прямого количественного сравнения численности и для выявления условий, благоприятствующих полиморфизму.
Рис. 1.
Рассмотренные стратегии жизнедеятельности популяции. Потоки ресурса показаны пунктиром, распределение потомков сплошными линиями. Прямоугольники обозначают ресурсные ниши четырех фенотипических классов. С1 – четыре независимых популяции специалистов, каждая занимающая свою ресурсную нишу; С4 – единая популяция специалистов, состоящая из четырех фенотипических классов (аналог фенотипического разнообразия), их потомки распределяются по всем классам равномерно; Г1 – четыре независимых популяции генералистов, каждая популяция черпает ресурс из двух ниш, но делает это в 2 раза менее эффективно, чем специалисты; Г4 – единая популяция генералистов, состоящая из четырех фенотипических классов, потомки которых распределяются по всем классам равномерно; К4 – единая популяция “коллективистов”, состоящая из четырех фенотипических классов, перераспределяющих добытый ресурс между всеми классами, пропорционально их численности.
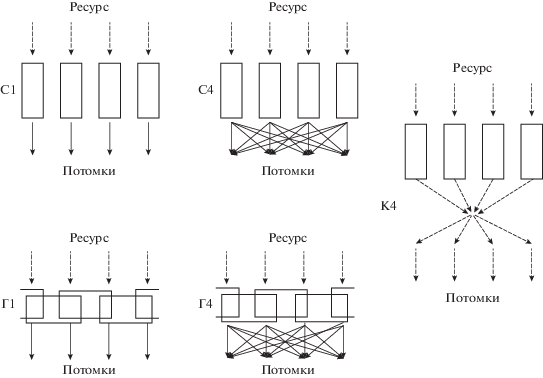
Степень толерантности фенотипов к вариабельности условий среды может быть высокой (генерализация) или низкой (специализация) (Алещенко, Букварева, 1991). В ходе моделирования мы будем сравнивать “специалистов” и “генералистов” (D’Odorico et al., 2008; Eisenhauer et al., 2012; Matias et al., 2013). Специалисты в нашей модели способны получать ресурс только из одной ниши, генералисты получают ресурс из двух соседних ниш, из каждой с коэффициентом 0.5 (нормировка для сопоставления со специалистами). Независимые мономорфные популяции генералистов Г1 будут сопоставлены с независимыми мономорфными популяциями специалистов С1 (рис. 1). Единая полиморфная популяция генералистов Г4, состоящая из четырех фенотипических классов, будет сопоставлена по численности с единой полиморфной популяцией специалистов С4, также состоящей из четырех фенотипических классов (рис. 1). Кроме того, рассмотрим так называемую плату за генерализацию (Matias et al., 2013) – за возможность получать ресурсы из двух ниш нужно платить снижением эффективности их использования, по сравнению со специалистами. В нашей модели плата за генерализацию состоит в том, что генералист осваивает лишь 95% добытого им ресурса (обозначим как Г95%), а генералист без такой платы осваивает все 100% (Г100%).
Перераспределение ресурсов – механизм кооперации внутри популяции, эффект которого мы тоже рассмотрим при моделировании полиморфной популяции, использующей разные ниши. У некоторых животных встречается распределение общей добычи между членами группы, добровольная передача добытого ресурса (food sharing) родственным или неродственным особям, взаимопомощь между определенными знакомыми индивидами (reciprocity), выпрашивание (tolerated scrounging) и терпимость к насильственному отъему ресурса другими особями (tolerated theft). Эти механизмы характерны, в частности, для кооперативных отношений людей и некоторых приматов (Jaeggi, Gurven, 2013а). Одной из возможных причин добровольной передачи ресурса считается механизм реципрокной взаимопомощи (reciprocity) (Реутов и др., 2019), при котором индивид делится добытым ресурсом с теми индивидами, от которых он сам ранее получал поддержку (Jaeggi, Gurven, 2013b). Поскольку мы не моделируем взаимодействия отдельных индивидов (как например, Pereda et al., 2017) и всех индивидов, принадлежащих одному фенотипическому классу, считаем одинаковыми относительно решаемой задачи, механизм обмена между индивидами можно масштабировать в модели на всю популяцию как обмен между фенотипическими классами. Мы не рассматриваем психологические мотивации участников обмена (Sznycer et al., 2017), а только ресурсный эффект обмена в рамках целой популяции. Возможность перераспределения ресурсов рассмотрим у единой популяции, состоящей из четырех фенотипических классов специалистов (как в ситуации С4), но у которых добытый ресурс равномерно перераспределяется между фенотипическими классами – стратегия “коллективизма” (обозначим эту ситуацию как К4; рис. 1, К4).
Имитируемый в модели (в ситуациях С4, Г4 и К4) внутрипопуляционный полиморфизм, при котором особи случайным образом распределяются по фенотипическим классам, различающимся по своей способности к эксплуатации тех или иных ресурсных ниш, можно рассматривать как вариант так называемого “биологического хеджирования ставок” (biological bet hedging). Быстро развивающаяся в последние годы концепция “биологического хеджирования ставок” основана на том, что эволюционирующие популяции (как и игроки на бирже) могут уменьшить риск, связанный с непредсказуемыми колебаниями среды (будь то цены на акции или количество осадков), путем оптимального распределения “ставок”, под которыми в случае биологических объектов понимаются инвестиции в различные адаптивные стратегии. За это, как правило, приходится платить снижением выигрыша (приспособленности, количества потомков) в благоприятных условиях (которым в нашей модели соответствует отсутствие кризиса), но в долгосрочной перспективе такая стратегия может оказаться наиболее успешной в непредсказуемо меняющейся среде. Классический пример биологического хеджирования — это растения, у которых часть семян всходит сразу, а часть — только спустя какое-то время (например, на следующий год). В стабильных благоприятных условиях это снижает репродуктивный успех, а в нестабильных — повышает (например, если случилась засуха, погубившая большую часть всходов этого года) (Childs et al., 2010; Starrfelt, Kokko, 2012).
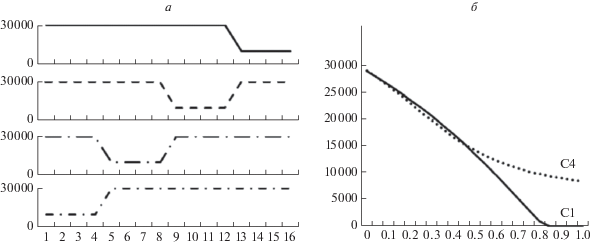
В нашей модели колебания количества ресурса в каждой нише моделируются как переходы между двумя дискретными состояниями: “нормальным” и “кризисным” (рис. 2а) (Gonzalez, Holt, 2002; D’Odorico et al., 2008; Wienand et al., 2017). Кризисная ситуация возникает по очереди в каждой из четырех ресурсных ниш (таким образом моделируется независимость колебаний в нишах). В каждый момент времени три ниши из четырех находятся в нормальной ситуации (рис. 2в); в следующий момент кризисная ситуация “перемещается” в соседнюю нишу. В нормальной ситуации в ресурсной нише есть максимальное количество ресурса (Rmax = 30 000 единиц ресурса), который может быть получен соответствующим фенотипом, адаптированным к данной нише. В кризисной ситуации ресурса меньше: в нише присутствует лишь определенный процент от Rmax (аналогично модели Винанда с соавт. (Wienand et al., 2017)). Аналогичные изменения ресурсной базы в природе могут быть связаны, например, с колебаниями погодных условий (Stenseth et al., 1999; Krebs et al., 2001; Бабурин, Чистяков, 2007). Количество ресурса во время кризиса равно Rmin = Rmax(1 – Н), где H – “сила кризиса”, являющаяся в модели мерой нестабильности среды. Например, если нестабильность среды Н = 0.9 (очень высокая), то Rmin = Rmax × 10% = 3000 (рис. 2а, в).
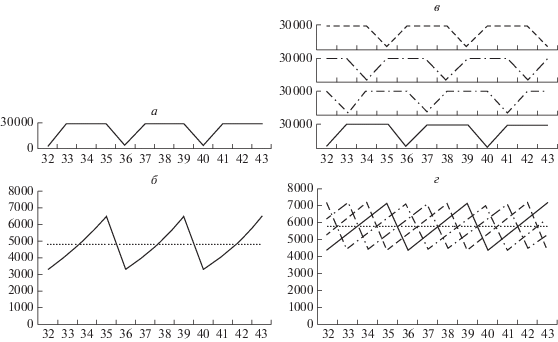
Рис. 2.
Пример динамики численности популяций в модели с короткими кризисами (длительность кризисов 1 такт). По горизонтали – время (показаны моменты от 32 до 43). а – колебания среды. По вертикали – количество ресурса в среде (3000 в кризисные такты, 30 000 в некризисные). Кризисная ситуация происходит каждые четыре такта. б – численность одного фенотипического класса при стратегии С1. По вертикали – численность. Пунктир – средняя численность всей популяции (равна 4817). в – колебания среды для четырех ниш. Обозначения как на а. Колебания ресурса в каждой нише сдвинуты по фазе на один такт для описания нестабильной среды с четырьмя различными нишами. г – численность всех четырех фенотипических классов для стратегии С4. Обозначения как на б. Пунктир – средняя численность всей популяции (равна 5786, что на 20% выше, чем на б). Стратегия С4 повышает относительную численность популяции в данной среде на 20% по сравнению со стратегией С1.
В модели одна единица ресурса может прокормить одну особь в единицу времени. Соответственно, количество доступного ресурса ограничивает численность популяции. В качестве закона размножения используется классический логистический закон размножения Ферхюльста (Ризниченко, Рубин, 1993):
Здесь N(t) – численность населения данной ниши в момент времени t, R – количество ресурса (“емкость среды”, равная Rmin либо Rmax), ra – коэффициент размножения (равен 0.3 для рис. 2, 3, 5; равен 0.6 для рис. 4), d – коэффициент смертности (равен 0.01). В некризисной ситуации количество ресурса R = Rmax = 30 000, и если, например, численность населения ниши N(t) = 3000, то N(t + 1) = 3000 + 0.3 × 3000 × 27 000/30 000 – 0.01 × × 3000 = 3781, т.е. прирост с учетом смертности +781. В кризисной ситуации количество доступного ресурса, как правило, оказывается меньше, чем численность населения ниши (т.е. чем численность соответствующего фенотипа). В этом случае 90% “избыточной” части населения погибает от голода. Например, если численность фенотипа в данный кризисный такт равна 4000 особей, а Rmin = 3000, то из 4000 – 3000 = 1000 особей 90% погибает и на следующем такте остается 3100 особей данного фенотипа (обитателей этой ниши). Однако в ситуациях С4 и Г4 часть потомков других фенотипов восполнят снизившуюся численность данного фенотипа на следующем такте (рис. 1).
Рис. 3.
Динамика численности популяции, занимающей одну нишу, в стабильной среде (Н = 0; величина ресурса в среде постоянна и равна 30 000; стартовая численность популяции 3000 особей). Кривая роста численности имеет S-образную форму.
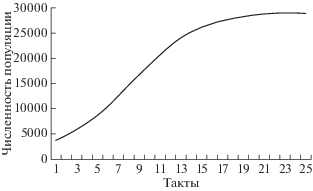
Рис. 4.
Результаты моделирования при длительности кризисов 1 такт. По горизонтальной оси – степень нестабильности среды (величина Н от 0 до 1), нестабильность увеличивается слева направо. а – зависимость численности популяции (вертикальная ось) от степени нестабильности среды при разных популяционных стратегиях. С ростом нестабильности среды численность снижается. В нестабильных условиях наиболее выгодной является стратегия кооперативного обмена ресурса (К4). Фенотипический полиморфизм с занятием разных ресурсных ниш (С4) выгодней мономорфизма и узкой нишевой специализации (С1), особенно в нестабильной среде. Генерализация особей (Г1) проигрывает специализации (С1) в стабильной среде (Н < 0.5), но выгоднее в нестабильной (Н > 0.7). б – графики эффективности продукции биомассы относительно величины потока ресурса (по вертикали). Обозначения те же. Видно еще более сильное превосходство стратегий К4 и Г1 над С1 по сравнению с а. в – аналогично а, но показаны стратегии генерализации без платы (Г100%) и с платой в 5% за генерализацию (Г95%). Генерализация без платы (Г100%) всегда выгоднее специализации (С1). При наличии небольшой платы (Г95%), генерализация выгодней только в нестабильной среде (Н > 0.7). г – отношение кривых на б к кривой С1. Показывает, насколько и при каких условиях другие стратегии выгоднее, чем С1. Видно, что в нестабильной среде (Н > 0.7) разнообразие ресурсных ниш (С4), генерализация (Г1) и кооперативное перераспределение ресурса (К4) выгоднее узкой нишевой специализации в 1.5 раза и более, зависимость нелинейная.
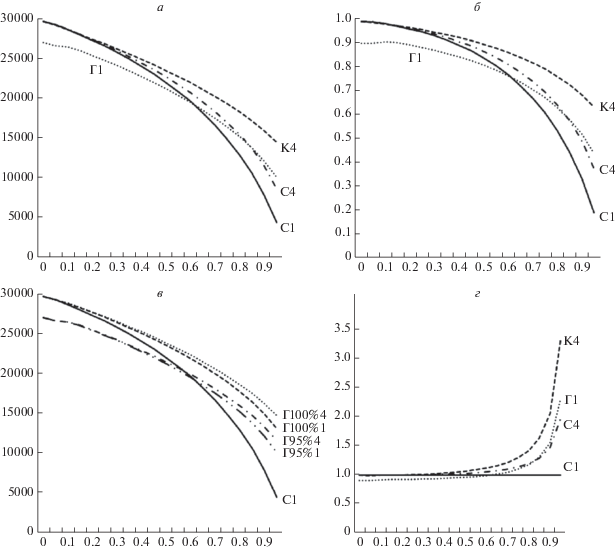
Рис. 5.
Результаты моделирования при длительности кризисов 4 такта. а – колебания ресурса в среде для четырех ниш. б – численность популяции (по вертикали) в зависимости от нестабильности среды (Н, по горизонтали). Стратегия С4 позволяет популяции выживать в нестабильной среде (при Н > 0.75).
Длительные кризисы такого типа не приводят к качественным изменениям по сравнению с короткими, поскольку на весь период кризиса фиксируются соответствующие низкие значения количества ресурса и численности. Поэтому дополнительно изучалось влияние длительных кризисов не путем уменьшения количества ресурса, а изъятием доли популяции (Менджел, Кларк, 1992). Для этого каждая ниша по очереди оказывалась в кризисной ситуации на четыре такта подряд (рис. 5а), причем количество ресурса не уменьшалось, но после применения формулы Ферхюльста снижалась численность популяции путем ее умножения на (1 – Н). Только в этом случае длительный кризис оказывает иное влияние, чем короткий.
Специалисты получают ресурс из одной ниши. Генералисты получают ресурс из двух соседних ниш, с коэффициентом 0.5. Если вводится плата за генерализацию, то из полученного ресурса вычитается 5%.
Начальная численность каждой из четырех популяций (или четырех фенотипов одной популяции) в каждой нише составляла по 3000 особей. Итоговая численность популяции оценивалась как среднее N(t) с момента t = 1000 до момента t = 2000. В модели рассматривались популяции, занимающие одну (С1, Г1) и четыре ниши (С4, Г4, К4). При этом для корректного сопоставления итоговых численностей для этих стратегий проводилась нормировка численности (величина N для С4, Г4, К4 делилась на 4). Оценивалась также эффективность продукции биомассы как отношение численности N к среднему количеству ресурса R в среде, что можно трактовать как КПД данной стратегии.
РЕЗУЛЬТАТЫ
В стабильной среде (Н = 0%) численность популяции в модели растет по логистическому закону (S-образная кривая) и далее остается на постоянном уровне (рис. 3), как и следует из модели размножения Ферхюльста, на которой основана наша модель. В нестабильной среде (Н = 90%) численность особей в нише колеблется в соответствии с колебаниями ресурса в этой нише (рис. 2). Соответственно, численность популяции, занимающей одну нишу (С1), колеблется в соответствии с колебаниями среды (рис. 2б). Численность популяции, занимающей несколько ниш (С4), оказывается более стабильна, так как колебания численности ее фенотипов, занимающих разные ниши, идут в противофазе (рис. 2г). Средняя численность полиморфной популяции, обладающей фенотипическим разнообразием (С4), оказывается в данном случае на 20% выше (сравните пунктирные линии на рис. 2б и г). Приведенный пример показывает, что увеличение фенотипического (нишевого) разнообразия в популяции может снижать колебания численности популяции (т.е. увеличивать ее стабильность) и повышать среднюю численность популяции в нестабильной среде по сравнению с популяцией без фенотипического (нишевого) разнообразия.
Рост нестабильности среды моделируется как увеличение силы кризисов, т.е. снижается средняя величина ресурса в среде, поэтому по мере роста нестабильности численность популяции ожидаемо снижается для всех типов стратегий (рис. 4а). Оказывается, что в нестабильной среде генерализация (Г1) выгоднее специализации особей (С1), фенотипическое разнообразие и полиморфизм популяции (С4) выгоднее мономорфизма и узкой специализации к одной нише (С1), а наиболее выгодно кооперативное перераспределение ресурса (К4) (рис. 4а, при Н > 0.6). В стабильной среде генерализация (Г1) менее эффективна, чем специализация (С1) (рис. 4а, Н < 0.5).
В нестабильной среде средний уровень поступающего ресурса ниже, чем в более стабильной, поэтому для изучения роли нестабильности среды имеет смысл сравнивать не только численность популяции при разных уровнях нестабильности (рис. 4а), но и отношение численности к величине поступающего из среды ресурса – аналог КПД продукции биомассы на единицу ресурса (рис. 4б). Форма кривых эффективности продукции биомассы (рис. 4б) аналогична форме кривых численности (рис. 4а), и порядок приоритета стратегий сохраняется, но относительная эффективность стратегий количественно меняется. Например, видно, что стратегия К4 еще более эффективна относительно стратегии С1, если оценивать эффективность продукции биомассы (рис. 4б) вместо численности популяции (рис. 4а). Также видно, что график К4 очень медленно спадает с ростом нестабильности среды, если оценивать эффективность продукции биомассы (рис. 4б). Это означает, что стратегия кооперативного перераспределения ресурса (К4) наиболее эффективно справляется с нестабильностью среды, а стратегия узкой нишевой специализации (С1) наименее эффективна. Для оценки этих различий представлены графики отношения эффективности разных стратегий относительно эффективности С1 (рис. 4г). Видно, что с ростом нестабильности среды эффективность продукции биомассы на единицу ресурса у всех прочих стратегий растет по сравнению с С1, т.е. с ростом нестабильности среды стратегия С1 становится всё менее выгодной по сравнению с другими. При этом фенотипическое разнообразие полиморфной популяции (С4) и генерализация особей (Г1) имеют сходный положительный эффект, а перераспределение добытого ресурса (К4) опережает их по эффективности. Полученные оценки позволяют ранжировать стратегии: наименее выгодной является С1, средней эффективностью обладают Г1 и С4, а самая выгодная К4 (рис. 4а, б, г).
Увеличение фенотипического разнообразия не дает существенного выигрыша в численности популяции генералистов (рис. 4в, сравните кривые Г100%1 и Г100%4, Г95%1 и Г95%4). Поэтому для оценки стратегии генерализации использовалась Г95%1 (Г1 на рис. 4а, б, г). При введении платы за генерализацию кривая численности опускается равномерно вниз (рис. 4в, кривая Г95%1 по сравнению с Г100%1, кривая Г95%4 по сравнению с Г100%4). Без платы за генерализацию (Г100%1 и Г100%4) в стабильной среде стратегия генерализации так же выгодна, как и стратегия специализации С1, а в нестабильной среде выигрывает у последней (рис. 4в). Введение даже небольшой платы за генерализацию в 5% уже приводит к тому, что в стабильной среде выгоднее специализация (рис. 4в, кривая С1), а в нестабильной – генерализация (рис. 4в, кривая Г95%1). Таким образом, соотношение эффективности стратегий специализации и генерализации зависит от наличия платы за генерализацию и ее величины, но в нестабильной среде генерализация в любом случае выгоднее.
При увеличении длительности кризисов до четырех тактов подряд (рис. 5а), с моделированием кризиса как изъятия доли особей на каждом такте, стратегия С1 оказывается неэффективной и популяция вымирает в нестабильной среде (рис. 5, кривая С1 при Н > 0.75). Однако увеличение фенотипического разнообразия (С4) спасает популяцию в этих условиях (рис. 5, кривая С4). Поэтому можно утверждать, что повышение нишевого разнообразия позволяет популяции выживать в нестабильной среде при сильных и долгих кризисах.
ОБСУЖДЕНИЕ
Поскольку фокусом работы является изучение влияния среды на популяцию, полностью учитывался поток ресурса и биомассы, что не всегда делается в моделях (Axelrod, 1984; Алещенко, Букварева, 1991; Norberg et al., 2001; Andras et al., 2007). В наиболее распространенном типе моделей кооперации (Axelrod, 1984; Andras et al., 2007; Kaznatcheev, Shultz, 2011) рассматриваются кооперативное и конкурентное взаимодействия двух индивидов, что описывается платежной матрицей 2 × 2 (2 индивида и 2 стратегии), в которой априорно задается, что при взаимной кооперации оба индивида получают больше ресурса, чем в других случаях. Выгода кооперативных отношений закладывается в таких моделях априорно, и механизмы кооперации не изучаются. Количество ресурса, получаемого из среды, определяется не средой, а типом социального взаимодействия. Такие модели не учитывают ограниченность объема доступного ресурса и не позволяют вести учет зависимого от среды потока ресурса из среды в популяцию. Поэтому для учета зависимости популяции от состояния среды в настоящей работе использовался закон размножения Ферхюльста, в котором явно учитывается ресурсная емкость, управляя которой можно моделировать колебания среды (Gonzalez, Holt, 2002; Wienand et al., 2017). В предложенной нами модели соблюдается закон сохранения материи (трансформации ресурса в биомассу популяции): в каждый момент времени задается количество ресурса в каждой ресурсной нише; далее регистрируется, сколько его добыто из ниши соответствующим этой нише фенотипическим классом, сколько из добытого затрачивается на питание имеющихся особей этого класса и сколько остается на размножение (одна новая особь на одну единицу ресурса). По результатам модельного эксперимента для каждой стратегии и каждого набора параметров вычислялось отношение количества особей N в популяции к имевшемуся в среде объему R ресурса. Эта величина N/R трансформации ресурса среды в биомассу популяции рассматривается как КПД используемой популяцией стратегии, и на этой основе проведено сравнение разных стратегий.
При увеличении фенотипического разнообразия популяции, что позволяет особям занимать дополнительные ресурсные ниши (рис. 1, С4), поток ресурса увеличивается, поэтому будет кратно увеличиваться численность популяции. Но такого же суммарного результата по численности могут достичь четыре независимые популяции разных видов, каждая занимающая свою нишу (рис. 1, С1). Показано, что увеличение фенотипического разнообразия (С4) выгодно в нестабильной среде, поскольку позволяет сглаживать колебания численности популяции за счет использования ресурсных ниш, колеблющихся в противофазе, и, например, при сильной нестабильности среды (Н = 0.9) дает выигрыш в численности популяции от +20% (рис. 2, при коэффициенте размножения ra = 0.3) до +100% (рис. 4, при коэффициенте размножения ra = 0.6).
Эти результаты согласуются с гипотезой о росте численности популяции при повышении ее фенотипического разнообразия в нестабильной среде (Алещенко, Букварева, 1991; Kussell, Leibler, 2005; Carja, Feldman, 2012), а также с результатами моделирования, показавшими, что повышение фенотипического разнообразия сообщества способствует выживанию в нестабильной среде (Алещенко, Букварева, 1991; Norberg et al., 2001).
В отличие от других работ, посвященных изучению биологических стратегий выживания в нестабильной среде, здесь нами в одной модели были сопоставлены и увеличение фенотипического разнообразия популяции (D’Odorico et al., 2008; Букварева, Алещенко, 2010), и генерализация особей (Алещенко, Букварева, 1991; D’Odorico et al., 2008; Букварева, Алещенко, 2010; Eisenhauer et al., 2012; Matias et al., 2013), и кооперативное перераспределение ресурсов (Кирдина, 2014; Pereda et al., 2017). Оказалось, что генерализация (Г1) и увеличение фенотипического (нишевого) разнообразия (С4) дают количественно сходный эффект (рис. 4, кривые Г1 и С4). Иными словами, стратегия увеличения фенотипического (нишевого) разнообразия популяции (рис. 1, С4) и стратегия генерализации особей (нишевого разнообразия особей; рис. 1, Г1) сопоставимы по своей эффективности.
Однако самой эффективной в нестабильной среде оказалась стратегия кооперативного перераспределения ресурсов (рис. 1, К4), которая по эффективности показала примерно двукратный выигрыш по сравнению с повышением фенотипического разнообразия популяции (С4) и генерализацией особей (Г1). Этот результат согласуется с нашей гипотезой о том, что нестабильная среда должна приводить к редистрибутивной экономике и кооперативным отношениям (рис. 6) (Кирдина и др., 2015). Аналогичный результат получен в модели, не учитывающей общий объем ресурса, в которой показано, что неопределенность среды способствует кооперации (Andras et al., 2007). Наш результат также аналогичен модельным результатам, показавшим, что вероятность выживания популяции в нестабильной среде (c невысокой вероятностью добычи ресурса) повышается, если индивиды делятся долей добытого ресурса с другими членами популяции (Pereda et al., 2017). У людей добровольная передача ресурса (food sharing) характерна для нестабильной среды (где ресурс может быть добыт в большом количестве, но с низкой вероятностью), но не для стабильной среды с гарантированным получением небольшого ресурса (Kaplan et al., 2012), что можно сопоставить с охотой и собирательством соответственно. Результаты этих работ согласуются с гипотезой настоящей работы. Интересно сопоставить механизм перераспределения добытого ресурса между особями популяции (food sharing) с механизмами межклеточного обмена метаболитами, которые позволяют, на основе принципов кооперации и синхронизации ритмов работы обменивающихся клеток, объединяться в многоклеточный организм (Швырков, 1995; Riera et al., 2007; Александров, 2008; Бродский, 2009; Magistretti, 2009; Греченко и др., 2015; Крылов, 2017).
Рис. 6.
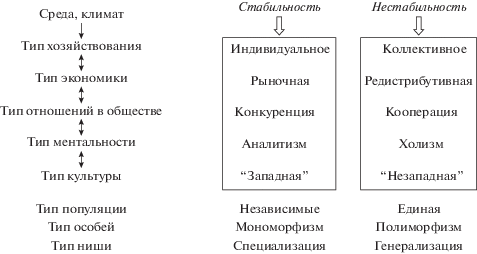
Взаимозависимости характеристик общества, определяемые средой. Гипотеза исследования (обозначена широкими стрелками) заключается в предположении о ключевой роли степени стабильности среды в формировании типа общества.
В настоящей работе не изучались условия, при которых независимые конкурирующие популяции будут иметь преимущество, эта задача не ставилась. Поэтому не удивительно, что во всех смоделированных ситуациях увеличение нишевого разнообразия оказалось выгодным (С4, Г1 и К4 выгоднее С1). В работе изучался вопрос о преимуществах единой популяции, обладающей функциональным разнообразием (рис. 1, С4), в сравнении с набором независимых узко специализированных популяций (рис. 1, С1). Показано, что с ростом нестабильности среды у набора независимых узко специализированных популяций (С1) быстро снижается численность (рис. 4а, кривая С1), а единая популяция, обладающая функциональным разнообразием (С4), оказывается более устойчивой (рис. 4a, кривая С4), особенно если она использует стратегию перераспределения ресурсов (рис. 4a, кривая К4).
При длительных сильных кризисах независимые узко специализированные популяции (С1) вообще не выживают (рис. 5, С1, при Н > 0.75). В этих же обстоятельствах единая популяция, обладающая функциональным разнообразием (С4), оказывается более устойчивой (рис. 5, С4). Длительные кризисы могут быть вызваны снижением количества ресурса в среде вследствие временного ухудшения погодных условий (Менджел, Кларк, 1992) или вследствие роста численности популяции до предела ресурсной емкости среды (Нефедов, 2012; Коротаев и др., 2015), антикорреляциями последовательных состояний среды (Gonzalez, Holt, 2002) или увеличением численности хищников или паразитов для данной популяции, что вызывает долговременное стрессовое воздействие (Krebs et al., 2001).
Независимые узко специализированные популяции (рис. 1, С1) в соответствии с моделью Ферхюльста хорошо выживают, по крайней мере, в стабильной среде (рис. 4, Н < 0.5). А с учетом платы за генерализацию (многонишевость) (Matias et al., 2013) популяция специалистов (С1) эффективнее в стабильных средах, чем популяция генералистов (Г1) (рис. 4, Н < 0.5). При этом генералисты эффективнее специалистов в нестабильных средах (рис. 4, Н > 0.5).
В настоящей работе методом математического моделирования изучалось, существуют ли условия среды, в которых стратегия кооперации и единства полиморфной популяции оказывается более эффективной, чем разделение на независимые мономорфные популяции. Проверялось предположение, что ресурсная стабильность среды является определяющим фактором: преимущества стратегии кооперации и единства популяции обнаруживаются в нестабильных средах (рис. 6). Логично также предположить, что в стабильных средах будет выгодна изоляция и конкуренция.
Кооперация и конкуренция рассматриваются как основополагающие стратегии существования сообществ не только в биологии, но и в экономике, психологии, культурологии, социальной нейронауке (см., например, Александров, Александрова, 2009; Peng, Hsieh, 2012; Alexandrov, Kirdina, 2013; Plass et al., 2013; Кирдина, 2014; Апанович и др., 2016). Выбор между кооперацией и конкуренцией связан с типом хозяйствования, экономическим укладом, типом мышления, ментальными моделями и типом культуры в данном обществе. При этом выделяются два альтернативных варианта (Nisbett et al., 2001; Александров, Кирдина, 2012; Нейсбит, 2012; Кирдина, 2014; Talhelm et al., 2014) (рис. 6):
– конкуренция характерна для обществ, типичными чертами которых являются индивидуалистская культура, частная собственность, рыночная экономика, ориентация на максимизацию прибыли, федеративное устройство, примат прав и свобод личности по отношению к ценностям сообщества, западная ментальность, аналитизм восприятия мира как набора обособленных объектов (можно обозначить как “западный” тип);
– кооперация характерна для обществ, типичными чертами которых являются коллективистская культура, редистрибутивная экономика (добытые ресурсы перераспределяются между членами коллектива), ориентация на снижение расходов, вертикальная иерархия централизованной власти, приоритет общих ценностей над личными, восточная ментальность, холистическое восприятие мира как набора взаимосвязей (можно обозначить как “незападный” тип).
Перераспределение ресурса (стратегия К4) между всеми особями популяции (жителями страны) через единый центр является специфичным механизмом “незападной” редистрибутивной экономики (Кирдина, 2014). На уровне страны это перераспределение является центральным регулятором финансов или прочих ресурсов от регионов-доноров к жителям регионов, нуждающихся в поддержке в данный период времени (как и в нашей модели в ситуации К4).
Представляется, что нишевое разнообразие особей в популяции более характерно для “незападной” культуры и экономики, поскольку единая популяция снижает вероятность конкуренции между фенотипическими классами, учитывая распределение потомков. Когда нишевое разнообразие популяции отсутствует, каждая популяция специализирована под свою нишу, подобно популяциям разных видов. Здесь можно усмотреть аналогии с “западной” культурой и экономикой. Независимые популяции концентрируют добытые ресурсы в росте численности только своего класса (или вида) и, в принципе, могут вступить в конкуренцию друг с другом (в настоящей работе не рассматривается).
В более стабильных климатических условиях обнаруживаются страны с “западным” стилем экономики, индивидуализмом и конкурентными отношениями, а в менее стабильных – страны с “незападным” стилем экономики, коллективизмом и кооперативными отношениями (Кирдина и др., 2015). Это согласуется с нашей гипотезой, которую можно переформулировать в этом контексте так: в нестабильной среде будет формироваться “незападная” экономика с кооперативными отношениями в сообществе и перераспределением ресурсов. Несмотря на долгую историю обсуждения и значимость роли отношений кооперации и конкуренции в сообществе, в настоящее время остаются слабо изученными причины и механизмы возникновения этих стратегий и их зависимость от условий среды. Можно предположить, что среда с ее климатическими и ресурсными особенностями определяет наиболее эффективный тип хозяйствования (например, в данной среде окажется предпочтительным оседлый образ жизни независимыми мелкими хозяйствами или, напротив, кочевой образ жизни единым племенем) (Гумилев, 1972; Нейсбит, 2012). Тип хозяйствования включает в себя индивидуалистский или коллективистский способ добычи ресурсов, а значит, формирует преимущественно отношения конкуренции или кооперации в данном обществе (Кирдина, 2014). Сформированный тип отношений хозяйствования связан с типом ментальности и закрепляется в виде культурных норм в данном обществе, поэтому внутри данной культуры или субкультуры (территориальные, профессиональные группы) наблюдается связь между типом хозяйствования и типом ментальности (Henrich et al., 2010; Александров, Кирдина, 2012; Нейсбит, 2012; Talhelm et al., 2014) (рис. 6).
В связи с изучением наличия и значения культуры и морали у животных (Laland, Galef, 2009) интересно сопоставить обсуждаемую гипотезу о нестабильности среды как причине формирования кооперативного поведения и коллективизма с данными о наличии таких отношений у животных в зависимости от нестабильности их ресурсной среды и с данными о закреплении таких отношений в “культуре” у животных (в частности, распределение пищи в стае) (Brosnan, 2006; Tan, Hare, 2013; см. обзор: Александров, Александрова, 2009). Кооперативные отношения могут быть вызваны и синергией, обусловленной коллективным поведением, например, при охоте стаей или коллективной защите. В целом можно говорить о том, что объединение функционально специализированных анатомически распределенных клеток в функциональную систему (Анохин, 1978; Швырков, 1995), органов в организм (Загускин, 2010), особей в семью или стаю, фенотипических классов в популяцию происходит за счет кооперативных отношений и позволяет созданному новому уровню целостности решать задачи в более сложной среде. Такое объединение создает новый уровень биологической иерархии (Букварева, Алещенко, 1997, 2010; Загускин, 2010), что соответствует рассматриваемому принципу: существование в определенном типе среды связано с формированием определенных отношений на биологическом уровне и, далее, на социальном. Отметим, что также имеется и обратное влияние – биота и антропогенная хозяйственная деятельность влияют на климат и среду (Букварева, 2010), что может закреплять формирование одного из двух типов отношений.
ЗАКЛЮЧЕНИЕ
Моделирование показало, что в стабильной среде и в среде с небольшими колебаниями условий оказываются успешными независимые популяции, специализированные каждая под свою нишу.
В очень нестабильной среде, в которой часто случаются сильные локальные кризисы, оказывается выгодным единство популяции, занимающей разные ниши: численность особей фенотипического класса, занимающего подвергшуюся кризису ресурсную нишу, быстрее восстанавливается за счет потомков особей других фенотипических классов. Еще более эффективной стратегией является кооперация в форме перераспределения добытых ресурсов по всей популяции, что позволяет сгладить последствия сильных кризисов за счет “обобщения” локальной проблемы в одной нише на всю популяцию. Также успешной для выживания в кризисной среде является генерализация возможностей особей, когда каждая особь способна получать ресурс из нескольких ниш, которые не могут быть одновременно в кризисном состоянии.
Полученные результаты помогают понять механизмы известных корреляций между типом отношений в обществе (кооперативных или конкурентных), типом экономики (централизованное перераспределение благ или совокупность экономически автономных субъектов; Кирдина и др., 2015), типом культуры и ментальности (Александров, Кирдина, 2012). Рассмотренные в работе возможные биологические механизмы влияния нестабильности среды на социальные отношения могут быть предпосылкой формирования соответствующих типов менталитетов, морали и культур (Александров, Александрова, 2009; Александров, Кирдина, 2012; Arutyunova et al., 2016; Кольцова, Журавлев, 2017).
Работа выполнена при поддержке РФФИ (грант № 18-29-22045).
Список литературы
Александров Ю.И., 2008. Активный нейрон // Нейрон: обработка сигналов, пластичность, моделирование: фундаментальное руководство / Под ред. Соколова Е.Н., Филиппова В.А., Черноризова А.М. Тюмень: Изд-во ТюмГУ. С. 33–58.
Александров Ю.И., Александрова Н.Л., 2009. Субъективный опыт, культура и социальные представления. М.: ИП РАН. 320 с.
Александров Ю.И., Кирдина С.Г., 2012. Типы ментальности и институциональные матрицы: мультидисциплинарный подход // Социол. исследования. № 8. С. 3–12.
Алещенко Г.М., Букварева Е.Н., 1991. Модель фенотипического разнообразия популяции в случайной среде // Журн. общ. биологии. Т. 52. № 4. С. 499–508.
Анохин П.К., 1978. Философские аспекты теории функциональной системы. М.: Наука. 400 с.
Апанович В.В., Безденежных Б.Н., Знаков В.В., Самс М., Яаскелайнен И., Александров Ю.И., 2016. Различия мозгового обеспечения индивидуального, кооперативного и конкурентного поведения у субъектов с аналитическим и холистическим когнитивными стилями // Эксперим. психология. Т. 9. № 2. С. 5–22.
Бабурин В.Л., Чистяков П.А., 2007. Пространство циклов: Мир–Россия–регион. М.: Изд-во ЛКИ. 320 с.
Бродский В.Я., 2009. Прямые межклеточные взаимодействия и социальное поведение клеток млекопитающих, протистов и бактерий. Возможные причины многоклеточности // Онтогенез. Т. 40. № 2. С. 97–111.
Букварева Е.Н., 2010. Роль наземных экосистем в регуляции климата и место России в посткиотском процессе. М.: Т-во науч. изд. КМК. 97 с.
Букварева Е.Н., Алещенко Г.М., 1997. Схема усложнения биологической иерархии в случайной среде // Успехи соврем. биологии. Т. 117. № 1. С. 18–32.
Букварева Е.Н., Алещенко Г.М., 2010. Оптимизация разнообразия надорганизменных систем как один из механизмов их развития в экологическом, микроэволюционном и эволюционном масштабах // Успехи соврем. биологии. Т. 130. № 2. С. 115–129.
Букварева Е.Н., Алещенко Г.М., 2013. Принцип оптимального разнообразия биосистем. М.: Т-во науч. изд. КМК. 522 с.
Греченко Т.Н., Харитонов А.Н., Жегалло А.В., Александров Ю.И., 2015. Психофизиологический анализ осцилляторных процессов в поведении биосоциальных систем // Психол. журн. Т. 36. № 5. С. 75–86.
Гумилев Л.Н., 1972. Изменения климата и миграции кочевников // Природа. № 4. С. 44–52.
Загускин С.Л., 2010. Ритмы клетки и здоровье человека. Ростов-на-Дону: Изд-во ЮФУ. 292 с.
Кирдина С.Г., 2014. Институциональные матрицы и развитие России. Введение в Х-Y-теорию. М.; СПб.: Нестор-История. 468 с.
Кирдина С.Г., Кузнецова А.Н., Сенько О.В., 2015. Климат и институциональные матрицы // Социол. исследования. № 9. С. 3–13.
Кольцова В.А., Журавлев А.Л., 2017. Сущностные характеристики и факторы формирования российского менталитета // Психол. журн. Т. 38. № 3. С. 5–17.
Коротаев А.В., Халтурина Д.А., Божевольнов Ю.В., 2015. Законы истории: Вековые циклы и тысячелетние тренды. Демография, экономика, войны. Отв. ред. Крадин Н.Н. Изд. 4-е. М.: ЛЕНАНД. 256 с.
Крылов А.К., 2017. Активность нейрона как способ получения метаболитов // Тр. XIX Междунар. научно-технич. конф. “Нейроинформатика-2017”. Ч. 1. М.: НИЯУ МИФИ. С. 154–163.
Менджел М., Кларк К., 1992. Динамические модели в экологии поведения. М.: Мир. 300 с.
Недорезов Л.В., 2015. Аппроксимация временных рядов по динамике Paramecium caudatum моделями Ферхюльста и Гомпертца: нетрадиционный подход // Биофизика. Т. 60. № 3. С. 564–573.
Нейсбит Р., 2012. География мысли. М.: Астрель. 285 с.
Нефедов С.А., 2012. Неомальтузианство в современной методологии истории // Вестн. РАН. Т. 82. № 12. С. 1092–1107.
Реутов Е.В., Реутова М.Н., Шавырина И.В., 2019. Реципрокность в сетях взаимопомощи (на материалах регионального исследования) // Социол. исследования. № 3. С. 106–117.
Ризниченко Г.Ю., Рубин А.Б., 1993. Математические модели биологических продукционных процессов: Учебное пособие. М.: Изд-во МГУ. 302 с.
Северцов А.С., 2013. Эволюционная экология позвоночных животных. М.: Т-во науч. изд. КМК. 347 с.
Швырков В.Б., 1995. Введение в объективную психологию. Нейрональные основы психики. М.: Ин-т психологии РАН. 162 с.
Alexandrov Yu., Kirdina S., 2013. Toward integration of social mental and institutional models: Systemic approach // Montenegrin J. Econ. V. 9. № 1. P. 7–15.
Andras P., Lazarus J., Roberts G., 2007. Environmental adversity and uncertainty favour cooperation // BMC Evol. Biol. V. 7. № 1. P. 240.
Arutyunova K.R., Alexandrov Y.I., Hauser M.D., 2016. Sociocultural influences on moral judgments: East–west, male–female, and young–old // Front. Psychol. V. 7. P. 1334.
Axelrod R.M., 1984. The Evolution of Cooperation. N.Y.: Basic Books. 241 p.
Brosnan S.F., 2006. Nonhuman species’ reactions to inequity and their implications for fairness // Soc. Justice Res. V. 19. № 2. P. 153–185.
Carja O., Feldman M.W., 2012. An equilibrium for phenotypic variance in fluctuating environments owing to epigenetics // J. R. Soc. Interface. V. 9. P. 613–623.
Childs D.Z., Metcalf C.J.E., Rees M., 2010. Evolutionary bet-hedging in the real world: Empirical evidence and challenges revealed by plants // Proc. Biol. Sci. V. 277. № 1697. P. 3055–3064.
D’Odorico P., Laio F., Ridolfi L., Lerdau M.T., 2008. Biodiversity enhancement induced by environmental noise // J. Theor. Biol. V. 255. P. 332–337.
Davis M.B., Shaw R.G., 2001. Range shifts and adaptive responses to quaternary climate change // Science. V. 292. P. 673–79.
Eisenhauer N., Scheu S., Jousset A., 2012. Bacterial diversity stabilizes community productivity // PLoS ONE. V. 7. № 3. P. 1–5.
Gonzalez A., Holt R.D., 2002. The inflationary effects of environmental fluctuations in source–sink systems // Proc. Natl. Acad. Sci. USA. V. 99. P. 14872–14877.
Henrich J., Heine S.J., Norenzayan A., 2010. The weirdest people in the world? // Behav. Brain Sci. V. 33. № 2–3. P. 61–83.
Isbell F., Craven D., Connolly J., Loreau M., Schmid B. et al., 2015. Biodiversity increases the resistance of ecosystem productivity to climate extremes // Nature. V. 526. № 7574. P. 574–577.
Jaeggi A.V., Gurven M., 2013a. Natural cooperators: Food sharing in humans and other primates // Evol. Anthropol. V. 22. P. 186–195.
Jaeggi A.V., Gurven M., 2013b. Reciprocity explains food sharing in humans and other primates independent of kin selection and tolerated scrounging: A phylogenetic meta-analysis // Proc. R. Soc. B. V. 280. № 1768. P. 20131615.
Kaplan H.S., Schniter E., Smith V.L., Wilson B.J., 2012. Risk and the evolution of human exchange // Proc. R. Soc. B. V. 279. P. 2930–2935.
Kaznatcheev A., Shultz T.R., 2011. Ethnocentrism maintains cooperation, but keeping one’s children close fuels it // Proceedings of the 33rd Annual Conference of the Cognitive Science Society. P. 3174–3179.
Krebs C., Boonstra R., Boutin S., Sinclair A., 2001. What drives the 10-year cycle of snowshoe hares? // BioScience. V. 51. № 1. P. 25–35.
Kussell E., Leibler S., 2005. Phenotypic diversity, population growth, and information in fluctuating environments // Science. V. 309. P. 2075–2078.
Laland K.N., Galef B.G., Jr., 2009. The Question of Animal Culture. Cambridge: Harvard Univ. Press. 360 p.
Magistretti P.J., 2009. Role of glutamate in neuron-glia metabolic coupling // Am. J. Clin. Nutr. V. 90. P. 875–880.
Matias M., Combe M., Barbera C., Mouquet N., 2013. Ecological strategies shape the insurance potential of biodiversity // Front. Microbiol. V. 3. P. 1–9.
Nisbett R.E., Peng K., Choi I., Norenzayan A., 2001. Culture and systems of thought. Holistic versus analytic cognition // Psychol. Rev. V. 108. P. 291–310.
Norberg J., Swaney D.P., Dushoff J., Lin J., Casagrandi R., Levin S.A., 2001. Phenotypic diversity and ecosystem functioning in changing environments: A theoretical framework // Proc. Natl. Acad. Sci. USA. V. 98. № 20. P. 11376–11381.
Peng W., Hsieh G., 2012. The influence of competition, cooperation, and player relationship in a motor performance centered computer game // Comput. Hum. Behav. V. 28. № 6. P. 2100–2106.
Pereda M., Zurro D., Santos J.I., Godino I.B., Alvarez M. et al., 2017. Emergence and evolution of cooperation under resource pressure // Nat. Sci. Rep. V. 7. P. 45574.
Plass J.L., O’keefe P.A., Homer B.D., Case J., Hayward E.O. et al., 2013. The impact of individual, competitive, and collaborative mathematics game play on learning, performance, and motivation // J. Educ. Psychol. V. 105. № 4. P. 1050–1066.
Ranta E., Kaitala V., Lundberg P., 1997. The spatial dimension in population fluctuations // Science. V. 278. № 5343. P. 1621–1623.
Riera J.J., Schousboe A., Waagepetersen H.S., Howarth C., Hyder F., 2007. The micro-architecture of the cerebral cortex: Functional neuroimaging models and metabolism // Neuroimage. V. 40. P. 1436–1459.
Starrfelt J., Kokko H., 2012. Bet-hedging — a triple trade-off between means, variances and correlations // Biol. Rev. V. 87. № 3. P. 742–755.
Stenseth N.C., Chan K.-S., Tong H., Boonstra R., Boutin S. et al., 1999. Common dynamic structure of Canada lynx population within three climatic regions // Science. V. 285. № 5430. P. 1071–1073.
Sznycer D., Lopez Seal M.F., Sell A., Lim J., Porat R. et al., 2017. Support for redistribution is shaped by compassion, envy, and self-interest, but not a taste for fairness // Proc. Natl. Acad. Sci. USA. V. 114. № 31. P. 8420–8425.
Talhelm T., Zhang X., Oishi S., Shimin C., Duan D. et al., 2014. Large-scale psychological differences within China explained by rice versus wheat agriculture // Science. V. 344. P. 603–608.
Tan J., Hare B., 2013. Bonobos share with strangers // PLoS One. V. 8. № 1. P. e51922.
Wienand K., Frey E., Mobilia M., 2017. Evolution of a fluctuating population in a randomly switching environment // Phys. Rev. Lett. V.119. P. 158301.
Yachi S., Loreau M., 1999. Biodiversity and ecosystem productivity in a fluctuating environment: The insurance hypothesis // Proc. Natl. Acad. Sci. USA. V. 96. P. 1463–1468.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии