

Журнал общей биологии, 2020, T. 81, № 4, стр. 243-256
Совместная динамика популяций черники и брусники в заповедном послепожарном сосняке-зеленомошнике. Модель с осредненными вероятностями перехода
А. А. Маслов 1, *, Д. О. Логофет 1, 2, **
1 Институт лесоведения РАН
143030 Московская обл., п/о Успенское, Ул. Советская, 21, Россия
2 Институт физики атмосферы им. А.М. Обухова РАН
119017 Москва, Пыжевский пер., 3, Россия
* E-mail: amaslov@ilan.ras.ru
** E-mail: danilal@postman.ru
Поступила в редакцию 27.02.2020
После доработки 22.03.2020
Принята к публикации 15.04.2020
Аннотация
Исследована совместная динамика популяций брусники (Vaccinium vitis-idaea) и черники (V. myrtillus) в заповедном сухом сосняке-зеленомошнике (зона смешанных лесов) на средних и поздних стадиях послепожарной сукцессии. Особенностями полевого эксперимента являлись: период наблюдений в 25 лет, большой объем выборки на постоянных трансектах, интервал учетов – 5 лет, учет встречаемости видов на площадках 0.2 × 0.2 м. В качестве модели наблюдаемой динамики предложена дискретная марковская цепь из четырех состояний: отсутствие двух видов на площадке, присутствие только брусники, присутствие только черники, совместное присутствие видов. Установлено, что на каждом интервале учетов в сосняке-зеленомошнике реализуются все возможные переходы между состояниями, что дает полный орграф цепи. Частота переходов из одного состояния в другое варьирует между интервалами наблюдений, однако для массовых вариантов перехода расчет критерия Мантеля–Хензеля показал однородность выборок, что позволило перейти от осреднения переходных матриц к осреднению вероятностей. Показано, что для трех состояний цепи существует однонаправленный тренд изменений во времени: перманентно возрастает доля площадок только с черникой и доля площадок с двумя видами, убывает доля площадок с отсутствием данных видов. Четвертое состояние (площадки только с брусникой) имеет локальный максимум на средних стадиях сукцессии, после чего его доля снижается. Стационарным результатом динамики ожидается распределение площадок по состояниям, где 29.1% площадок занято только черникой, 15.2% – только брусникой, на 44.4% площадок будут присутствовать оба вида, а 11.3% площадок будут незаняты. Результаты подтверждают возможность устойчивого сосуществования черники и брусники на заключительных стадиях сукцессии. Описаны несколько стадий сукцессии: сначала – с малым участием кустарничков, затем – с преобладанием брусники и наконец – с доминированием черники (хотя преобладают площадки, где оба вида произрастают совместно). Полученные результаты позволяют оценить общую продолжительность послепожарной сукцессии в сухом сосняке-зеленомошнике в зоне смешанных лесов в 140 лет (с достижением состояния, отличного от стационарного менее чем на 5%). С позиций лесной типологии, результат моделирования означает постепенное превращение сосняка лишайниково-зеленомошного в сосняк чернично-брусничный и далее – в сосняк бруснично-черничный.
Сосновые леса на бедных и дренированных песчаных почвах (так называемые боровые сосняки) – широко распространенный тип сообществ зоны бореальных лесов (Hytteborn et al., 2005; Рысин, Савельева, 2008). В центре Русской равнины такие леса особенно часто встречаются по повышенным элементам рельефа на территории флювиогляциальных равнин – в Мещёре и Верхневолжской низменности (Курнаев, 1969; Огуреева и др., 1996).
Сукцессионные стадии послепожарной динамики в таких сосняках довольно подробно изучены в лесах северной и средней тайги (Горшков, 1995; Ипатов и др., 1995; Самойлов, Ипатов, 1995; Кулешова и др., 1996; Hautala et al., 2005), тогда как подобных работ по соснякам в южной тайге и в зоне смешанных лесов очень мало (Коротков, 2002; Маслов, 2002). На ранних стадиях послепожарной сукцессии в боровых сосняках преобладают лишайники, затем им на смену приходят зеленые мхи (Pleurozium schreberi, виды р. Dicranum), на более поздних стадиях преобладают кустарнички – брусника (Vaccinium vitis-idaea), черника (V. myrtillus) и (только на Севере) вороника (Empetrum nigrum). Однако соотношение видов кустарничков в ходе сукцессии и в ее завершающей фазе до сих пор является неизученным. Дискуссионным, в частности, остается вопрос, возможен ли переход сообщества из типа леса “сосняк лишайниково-зеленомошный” в тип леса “сосняк бруснично-черничный” (Маслов, 2002).
Ранее была построена марковская модель совместной динамики популяций брусники и черники в ходе послепожарной сукцессии на примере сырого сосняка-долгомошника (Маслов, Логофет, 2016; Logofet, Maslov, 2019). Модель показала, что в ходе сукцессии на поздних стадиях заметно снижается доля площадок только с брусникой, но возрастает доля площадок, где оба вида произрастают совместно. Оставалось неясным, характерен ли данный тип динамики только для одного (сравнительно редкого) типа сырых сосняков или же данный тип отношений между черникой и брусникой носит универсальный характер и характерен для сосняков разных типов. Для настоящего исследования взят распространенный и сравнительно хорошо известный зеленомошный тип боровых сосняков. Методы полевых наблюдений, регулярность и продолжительность учетов здесь полностью соответствуют подходам, описанным ранее, что облегчает сопоставление результатов, полученных в двух сообществах.
Наряду с количественной оценкой характеристик устойчивого (финального) состояния популяций черники и брусники большой интерес для теории фитоценологии представляет вопрос оценки скорости послепожарной сукцессии. Наконец, в методическом плане по-прежнему актуальным остается вопрос осреднения данных о переходах между различными состояниями марковской цепи в том случае, когда в природе был проведен ряд регулярных учетов.
В предыдущей работе (Логофет, Маслов, 2018) мы указали, что главной причиной неизбежных различий переходных матриц являются флуктуации численности побегов изучаемых видов – как в результате воздействия погодных факторов, так и вследствие нарушений различного рода, включая воздействие фитофагов. Принимая гипотезу о случайной изменчивости матриц, мы перешли к вопросу об усредненной матрице и технологии ее построения. Согласно концепции структурно-геометрического среднего неотрицательных матриц (Logofet, 2018), средняя переходная матрица марковской цепи была получена путем извлечения матричного корня 5-й степени из произведения пяти исходных матриц (Logofet, Maslov, 2019).
При всей математической элегантности такой подход не лишен и определенных недостатков. Во-первых, геометрическое осреднение матриц оперирует матрицами как целостными сущностями, тогда как конкретные переходы из одного состояния в другое имеют характер отдельных случайных событий. Во-вторых, осреднение переходных матриц предполагает работу с выборкой, число площадок в которой неизменно и сохраняется в каждом учете. Такое жесткое требование приводит к необходимости исключения из итоговой выборки все новых и новых площадок на каждом шаге повторных учетов из-за появления валежа на разных площадках в разных учетах. В-третьих, геометрическое осреднение матриц требует нетривиальных математических операций по извлечению матричного корня степени k, где k – число повторных учетов. С ростом k сложность такой задачи все более возрастает.
В настоящей работе мы предлагаем от осреднения матриц перейти к осреднению вероятностей перехода из одного состояния в другое, что соответствует вероятностному подходу к оценке случайных событий. Такое решение позволяет, во-первых, отдельно работать с конкретными переходами (используя методы статистики), не требует, во-вторых, наличия одного и того же числа площадок в каждом учете (что сохраняет максимально возможное число площадок на каждом шаге учетов), и в-третьих, не требует извлечения корня из матриц, что позволяет делать расчеты модели в программе MS Excel.
Цель настоящей работы – анализ совместной динамики популяций брусники и черники на средних и поздних стадиях послепожарной сукцессии в типичном боровом сосняке-зеленомошнике при помощи однородной (во времени) марковской модели. В задачи работы входили: количественная оценка стационарного (финального) распределения вероятностей состояний, оценка средней длительности стадий и характеристик цикличности. В методическом плане при построении усредненной модели по нескольким интервалам наблюдений предлагается вместо осреднения матриц (Логофет, Маслов, 2018) использовать осреднение вероятностей перехода. Биологическая интерпретация результатов моделирования должна ответить на вопросы о соотношении численности популяций брусники и черники на разных стадиях сукцессии, включая вопрос о потенциальном конкурентном вытеснении одного вида другим.
ОБЪЕКТЫ И МЕТОДЫ
Объекты. Полевые исследования проводились в заповедном лесном участке № 63 на севере Клинского района Московской обл. (зона смешанных лесов). Постоянная пробная площадь размером 50 × 40 м была заложена в послепожарном сосняке-зеленомошнике – типичном сообществе Верхне-Волжской низменности (Маслов, 2002). Год сплошного пожара – 1894; следов более поздних низовых пожаров не обнаружено. Участок занимает верхнюю часть пологой гряды со слабовыраженным микрорельефом. Почва – песчаная, крайне бедная элементами минерального питания.
Характеристика сообщества на момент начала наблюдений в 1980 г. опубликована в монографии А.А. Маслова (1990). Древостой состоит из послепожарной сосны с единичной примесью березы и ели, средний возраст сосны – 70 лет. Характерно наличие в древостое больших окон. Редкий разновысотный еловый подрост неравномерно распределен по площади. В травяно-кустарничковом ярусе доминировали брусника и черника, однако суммарное их покрытие не превышало 30%. Из других видов сравнительно часто встречалась только овсяница овечья (Festuca ovina). Мхи и лишайники имели покрытие более 90%. Доминировали зеленые мхи Pleurozium schreberi и Dicranum rugosum; отдельные пятна формировали лишайники Cladina arbuscula и C. rangiferina. Большую часть территории занимали чисто зеленомошные участки. На момент начала наблюдений сообщество относилось к типу леса “сосняк лишайниково-зеленомошный”. Изменения в нижних ярусах за 15 лет наблюдений показали снижение роли лишайников и постепенное возрастание роли брусники и черники (Маслов, 2002).
Метод наблюдений, характер данных. Учеты видов нижних ярусов проводили на постоянных трансектах, состоящих из закрепленных на местности площадок размером 0.2 × 0.2 м; общее число площадок на пробной площади составило 2000. На площадках отмечали факт присутствия укоренившихся побегов трав и кустарничков и присутствие видов мхов. В качестве счетных единиц у черники и брусники выступали парциальные побеги. Отдельно отмечали также наличие на поверхности почвы валежа – стволов и крупных ветвей. Подробнее полевая методика описана в первой публикации по теме (Маслов, Логофет, 2016) и в монографии (Маслов, 1990).
Продолжительность наблюдений на постоянных трансектах составила 25 лет: с 1980 по 2005 год с проведением перечетов каждые пять лет. Всего имеется шесть учетов с картами видов, что позволяет отследить изменения на площадках за пять интервалов: 1980 → 1985, 1985 → 1990, 1990 → 1995, 1995 → 2000, 2000 → 2005. Общее число площадок, задействованное в расчетах, составило 12 000.
Марковская цепь как модель наблюдаемого процесса. Если обозначить (как это было при полевых учетах) присутствие на площадке черники буквой V, а брусники буквой W, то в двувидовой системе для каждой площадки в любой момент времени возможно всего четыре состояния: 1) ∅ – оба вида отсутствуют (такие площадки для краткости называем далее “незанятыми”), 2) V – есть только черника, 3) W – есть только брусника, 4) VW – присутствуют оба вида. Определенные выше состояния естественно отождествляются с состояниями марковской цепи, а переменными состояния модели служат конечные распределения x(t) вероятностей состояний цепи в момент наблюдения t = 1980, 1985, …, 2005 (Маслов, Логофет, 2016).
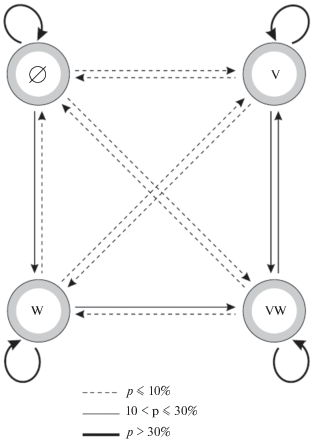
Постулат марковости означает, что распределение вероятностей переходов из любого состояния S (S = ∅, V, W, VW) во все остальные целиком определяется текущим состоянием и не зависит от того, каким путем цепь пришла в это состояние. Из априорных биологических представлений следует, что в природе возможны все типы переходов между четырьмя перечисленными состояниями, и если принять постулат марковости, то схему переходов можно отобразить в виде полного ориентированного графа (рис. 1), в котором присутствуют дуги, ведущие из любого узла графа в любой другой, включая тот же самый. Дополнительно на рис. 1 разными типами линий показан количественный уровень соответствующей вероятности перехода из одного состояния в другое, но этот вопрос мы рассмотрим при обсуждении результатов.
Рис. 1.
Полный граф переходов между четырьмя состояниями учетной площадки за один временной шаг Δt = 5 (по: Маслов, Логофет, 2016, с изменениями). Состояния: ∅ – оба вида отсутствуют, V– есть только Vaccinium myrtillus, W – есть только V. vitis-idaea, VW – присутствуют оба вида. Тип линии показывает диапазон значений вероятности p соответствующего перехода (см. табл. 2).
Формально, если распределение вероятностей перехода из некоторого состояния S (S = ∅, V, W, VW) в каждое из состояний представлено стохастическим вектором-столбцом [p∅S, pVS, pWS, pVW,S]T, а вероятности самих состояний в текущий момент времени t – стохастическим вектором-столбцом x(t) = [x∅, xV, xW, xVW]Т, то элементарная формула теории вероятностей дает вероятность состояния S в следующий момент, равную
(1)
$\begin{gathered} {{x}_{S}}(t + \Delta t) = {{x}_{\emptyset }}(t){{p}_{S}}_{\emptyset } + {{x}_{{\text{V}}}}(t){{p}_{S}}_{{\text{V}}} + \\ + \,\,{{x}_{{\text{W}}}}(t){{p}_{S}}_{{\text{W}}} + {{x}_{{{\text{VW}}}}}(t){{p}_{{S,{\text{VW}}}}},\,\,\,\,S = ~\,\,\emptyset ,{\text{V}},{\text{W}},{\text{VW}}. \\ \end{gathered} $Ненулевые элементы pij переходной матрицы P = [ pij] – имеющие смысл вероятности перехода из состояния j в состояние i за один шаг Δt = 5 годам (i, j = 1, …, 4) и отражающие относительную скорость процесса замещения видов на площадках – зависят от момента времени t, однако такой зависимостью пренебрегают, принимая гипотезу однородности цепи.
Если гипотеза справедлива, то совокупность всех четырех уравнений (1) с учетом орграфа (рис. 1) принимает следующий векторно-матричный вид:
(2)
$\begin{gathered} x(t + \Delta t) = \left[ {\begin{array}{*{20}{c}} {{{p}_{{11}}}}&{{{p}_{{12}}}}&{{{p}_{{13}}}}&{{{p}_{{14}}}} \\ {{{p}_{{21}}}}&{{{p}_{{22}}}}&{{{p}_{{23}}}}&{{{p}_{{24}}}} \\ {{{p}_{{31}}}}&{{{p}_{{32}}}}&{{{p}_{{33}}}}&{{{p}_{{34}}}} \\ {{{p}_{{41}}}}&{{{p}_{{42}}}}&{{{p}_{{243}}}}&{{{p}_{{44}}}} \end{array}} \right]x(t), \\ t = 1980,\,\,1985,\,\,...\,\,2000, \\ \end{gathered} $Время первого достижения представляет собой случайную величину, которая может принимать бесконечное (счетное) множество значений (1, 2, …, ∞) с разными вероятностями в зависимости от разных “маршрутов” достижения, возможных согласно орграфу (рис. 1). Математическое ожидание этой случайной величины и есть среднее время первого достижения (Маслов, Логофет, 2016, формула (А2)). Вместе со средней длительностью Mj каждого j-го состояния времена первого достижения представляют собой временны́е характеристики случайного процесса, выраженные в числе шагов. При известной величине временнóго шага эти характеристики получают выражение и в хронологических единицах (годах). Математические подробности соответствующих вычислений представлены в наших предыдущих работах (Логофет, Маслов, 2018, приложение А; Logofet, Maslov, 2019).
Для оценки длительности хода сукцессии было предложено (Маслов, Логофет, 2016) на каждом шаге по времени оценивать показатель несходства – меру, Δ(x(t), x*), отличия текущего распределения x(t) от стационарного x* по Кейфицу (Keyfitz, 1968, p. 47),
(4)
$\Delta (x,t),x{\text{*)}}\,\,{\text{ = }}\,\,\frac{1}{2}\sum\limits_{j = 1}^n {\left| {{{x}_{j}}\left( t \right) - x_{j}^{*}} \right|} ,$Метод калибровки. Характер имеющихся данных – наличие повторных учетов на постоянных трансектах в сочетании с большим размером выборки – предоставляет (довольно редкую в экологии) возможность вычислить все параметры модели однозначно и достоверно. Параметрами служат 16 переходных вероятностей pij из уравнения (2), точнее 12 переходных и 4 вероятности pjj сохранения текущего состояния за один шаг. О вероятностях мы судим по фактической картине переходов (табл. 1) и в качестве их значений принимаем частоты состоявшихся событий перехода и сохранения состояния. Тогда четыре калибровочных уравнения (1) удовлетворяются автоматически.
Таблица 1.
Данные о переходах между состояниями на шаге [t, t + 5] и о распределении состояний площадок в сосняке-зеленомошнике за 25 лет наблюдений; V – черника, W – брусника, VW – черника вместе с брусникой, ∅ – отсутствие обоих видов
| Год t | Переходы в | Исходящие состояния в год t | Итого в год t + 5 | |||
|---|---|---|---|---|---|---|
| ∅ | V | W | VW | |||
| 1980 | ∅ | 904 | 6 | 29 | 0 | 939 |
| V | 46 | 67 | 7 | 19 | 139 | |
| W | 222 | 2 | 242 | 3 | 469 | |
| VW | 16 | 58 | 81 | 207 | 362 | |
| Итого в год t | 1188 | 133 | 359 | 229 | 1909 | |
| 1985 | ∅ | 642 | 8 | 22 | 2 | 674 |
| V | 44 | 85 | 7 | 78 | 214 | |
| W | 205 | 0 | 325 | 6 | 536 | |
| VW | 26 | 45 | 100 | 269 | 440 | |
| Итого в год t | 917 | 138 | 454 | 355 | 1864 | |
| 1990 | ∅ | 464 | 18 | 37 | 6 | 525 |
| V | 52 | 146 | 6 | 93 | 297 | |
| W | 124 | 5 | 377 | 11 | 517 | |
| VW | 20 | 39 | 91 | 314 | 464 | |
| Итого в год t | 660 | 208 | 511 | 424 | 1803 | |
| 1995 | ∅ | 360 | 13 | 23 | 1 | 397 |
| V | 54 | 189 | 4 | 45 | 292 | |
| W | 93 | 5 | 384 | 22 | 504 | |
| VW | 23 | 73 | 97 | 372 | 565 | |
| Итого в год t | 530 | 280 | 508 | 440 | 1758 | |
| 2000 | ∅ | 277 | 21 | 35 | 11 | 344 |
| V | 32 | 194 | 22 | 192 | 440 | |
| W | 71 | 6 | 362 | 30 | 469 | |
| VW | 21 | 76 | 96 | 345 | 538 | |
| Итого в год t | 401 | 297 | 515 | 578 | 1791 | |
Метод осреднения. Принимая гипотезу о случайной изменчивости матриц, мы принимаем, что в ряду наблюдений частота перехода из одного состояния в другое, зафиксированная в текущем учете, является фактической реализацией случайного числа переходов. Для каждого типа переходов частоты меняются по годам наблюдений (Приложение А, Б). Логично предположить, что осреднение частот переходов, полученных в разных выборках (интервалах учетов) за весь период наблюдений, позволит более точно оценить истинную вероятность перехода из одного состояния в другое. Таким образом, от проблемы осреднения переходных матриц (Логофет, Маслов, 2018) мы переходим к проблеме осреднения вероятностей перехода.
Известен критерий (тест) Мантеля–Хензеля (Mantel–Haenszel test) для оценки возможности осреднения вероятностей через оценку степени однородности материала в разных выборках. Как правило, этот тест применяется в медицинских исследованиях, где проверка однородности материала в разных выборках имеет критическое значение (Borenstein et al., 2009). В марковской модели сукцессии применение теста на однородность мы предлагаем использовать как дополнительный аргумент в пользу случайности изменения вероятностей перехода за весь период исследований.
Осреднение вероятностей для каждого перехода из состояния j в состояние i проводили по стандартной формуле (Справочник…, 1989, с. 378):
(5)
$\overline {{{p}_{{ij}}}} = \frac{{\sum\limits_{l = 1}^k {{{n}_{l}}(j \to i)} }}{{\sum\limits_{l = 1}^k {{{N}_{l}}} }},$Математические подробности вычисления теста Мантеля–Хензеля и пример расчета средневзвешенных вероятностей даны в Приложении В. Все виды расчетов (как на этапе подготовки данных, так и в процессе моделирования) выполнены путем автоматизации вычислений в программе MS Excel, включая операции матричной алгебры.
РЕЗУЛЬТАТЫ
Вероятности переходов и их осреднение. Данные о фактических переходах между состояниями площадок за пять интервалов учетов представлены в абсолютных числах в табл. 1. Чтобы убрать влияние фактора пространственного исключения и фактора нарушения, при построении матриц переходов из расчетов были исключены площадки с живыми и сухостойными стволами деревьев и площадки с наличием валежа. В результате размеры выборки варьируют в разные интервалы учетов от 1758 до 1909.
Из 80 элементов всех пяти переходных матриц только два элемента (VW → ∅ за 1980 → 1985 и V → W за 1985 → 1990) оказались нулевыми, что подтверждает предположение о реализации всех теоретически возможных переходов между состояниями и сохранении текущих состояний за пятилетние интервалы между учетами. Одновременно это свидетельствует о правильном выборе длины временнóго шага и количества пробных площадок: за каждые пять лет в сообществе реализовались практически все переходы, что подтверждает полноту орграфа переходов (рис. 1) в сосняке-зеленомошнике.
Результаты расчета критерия Мантеля–Хензеля для всех типов переходов позволили доказать статистически достоверную однородность материала для 8 из 16 типов перехода (табл. 2). Среди оставшихся переходов часть составляют очень редкие (например, ∅ → VW или V → W; см. табл. 3), а остальные приходятся на переходы c очень сильным варьированием частот (Приложение Б), но без какого-то тренда к увеличению или снижению их значений. Таким образом, в целом результаты расчетов критерия Мантеля–Хензеля (с учетом малого числа интервалов для осреднения) не противоречат принятой нами гипотезе о случайном характере варьирования частот переходов за весь период полевых наблюдений, что позволяет считать корректным расчет осредненных вероятностей переходов по формуле (5).
Таблица 2.
Результаты вычисления критерия Мантеля–Хензеля для каждого типа переходов в ряду из k = 5 интервалов (гипотеза об однородности материала принимается при значении критерия < 2k)
| Куда переход | Исходящие состояния | |||
|---|---|---|---|---|
| ∅ | V | W | VW | |
| ∅ | 5.2 | 4.1 | 6.8 | 11.2 |
| V | 32.1 | 5.8 | 19.6 | 83.0 |
| W | 6.1 | 3.2 | 2.4 | 14.8 |
| VW | 21.5 | 20.2 | 4.1 | 30.9 |
Таблица 3.
Матрица усредненных вероятностей переходов (за пять пятилетних интервалов) и ее предельный вектор x*; все значения даны в процентах
| Куда переход | Исходящие состояния | Предельный вектор x* | |||
|---|---|---|---|---|---|
| ∅ | V | W | VW | ||
| ∅ | 71.6 | 6.2 | 6.2 | 1.0 | 11.3 |
| V | 6.2 | 64.5 | 2.0 | 21.1 | 29.1 |
| W | 19.3 | 1.7 | 72.0 | 3.5 | 15.2 |
| VW | 2.9 | 27.6 | 19.8 | 74.4 | 44.4 |
Осредненные вероятности переходов. Калиброванная переходная матрица, вычисленная по данным табл. 1, представлена в табл. 3. Ее элементы (переходные вероятности), найденные как средневзвешенные частоты соответствующих событий по формуле (5), показаны для наглядности в форме процентов. На рис. 1 градации этих же вероятностей показаны разными типами линий. Наибольшими значениями (> 60%) за пятилетний интервал характеризуются только диагональные элементы матрицы – вероятности сохранения каждого из четырех состояний. Это говорит о медленном ходе сукцессии. Как следует из рис. 1, основными направлениями изменения состояний (с вероятностью от 10 до 30%) являются переходы ∅ → W, W → VW и переходы из VW → V и обратно. Все остальные типы переходов, показанные пунктиром, характеризуются малыми (< 10%) вероятностями.
Стационарное распределение вероятностей состояний. В последнем столбце табл. 3 показан предельный (равновесный) вектор вероятностей x*, полученный как решение уравнения (3) с матрицей из табл. 3. Иными словами, предельным результатом сукцессии (при гипотезе однородности во времени) должно быть распределение состояний, где 15.2% площадок будет занято только брусникой, 29.1% – только черникой, на 44.4% площадок будут присутствовать оба вида, а 11.3% площадок будут существовать без кустарничков.
В табл. 4 представлены показатели несходства между распределениями в конкретные годы и предельным распределением, а рис. 2 дает представление о сходимости начального распределения x(1980) к равновесию x* по пятилетним шагам. Видно, что компоненты начального распределения достаточно медленно приближаются к предельным значениям. Так, распределение первого года учетов (1980) отличается от стационарного на 54.5%, а последнего года учетов (2005) – на 18.9%. Отличие менее 5% достигается только на 11-м шаге, который соответствует 2035 г. Важно отметить, что показатель несходства уменьшается на каждом шаге, что свидетельствует о направленности процесса в целом и в пользу состоятельности калиброванной матрицы из средневзвешенных вероятностей.
Таблица 4.
Показатели несходства между распределениями x(t) в конкретные годы и равновесием x*
| Год | Показатель, % |
|---|---|
| 1980 | 54.5 |
| 1985 | 45.9 |
| 1990 | 37.7 |
| 1995 | 30.3 |
| 2000 | 24.1 |
| 2005 | 18.9 |
| 2010 | 12.4 |
| 2015 | 11.4 |
| 2020 | 8.8 |
| 2025 | 6.7 |
| 2030 | 5.1 |
| 2035 | 3.9 |
| 2040 | 2.9 |
| 2045 | 2.2 |
Рис. 2.
Динамика распределения состояний площадок по пятилетним шагам в усредненной модели от начала наблюдений до достижения равновесного состояния.
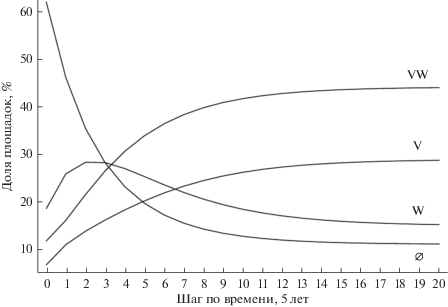
Таким образом, в двувидовой системе продолжительность вторичной послепожарной сукцессии (с достижением состояния, отличного от стационарного менее чем на 5%) составляет от момента начала полевых наблюдений 55 лет (11 пятилетних модельных шагов). К этому времени необходимо добавить 85 лет от года пожара до начала учетов в 1980 г., и, таким образом, время сукцессии составит 140 лет.
Как следует из рис. 2, в ходе сукцессии возрастает доля площадок только с черникой (V). Еще быстрее возрастает доля площадок, где оба вида произрастают совместно (WV). Напротив, доля “незанятых” площадок (∅) быстро снижается. В отличие от монотонного возрастания/убывания для V, WV и ∅, число площадок только с брусникой (W) сначала быстро растет, а затем плавно снижается – кривая имеет локальный максимум.
Расчет по шагам сходимости начального распределения состояний x(1980) к равновесному состоянию x* (рис. 2) позволяет воспроизвести и динамику популяций каждого вида отдельно в традиционной для биологов форме. Действительно, если Vm обозначить как число всех площадок с черникой, а Vv – как число всех площадок с брусникой, то на каждом шаге Vm = V + WV, а Vv = W + WV.
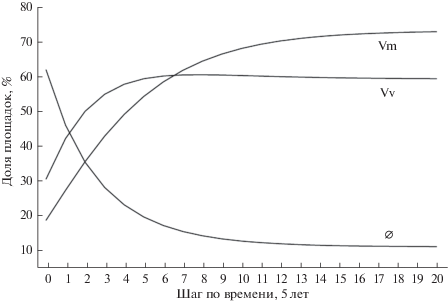
На рис. 3 представлена динамика популяций брусники и черники по шагам усредненной модели. Из рисунка (и табл. 3) следует, что в стационарном состоянии абсолютно преобладают площадки с черникой, так как на них суммарно приходится 73.5% площадок. Несмотря на существенное снижение доли площадок только с брусникой на поздних стадиях (рис. 2), суммарная доля брусники на поздних стадиях сначала выходит на максимум (на 7–8-м шаге достигая значения 60.8%), а затем незначительно уменьшается, составляя в стационарном состоянии 59.6% (табл. 3, рис. 3). Таким образом, согласно модели, на средних стадиях сукцессии преобладает брусника (рис. 3), а на поздних и в равновесном состоянии – черника.
Рис. 3.
Динамика популяций Vaccinium myrtillus (Vm) и V. vitis-idaea (Vv) по шагам в усредненной модели от начала наблюдений до достижения равновесного состояния. Vm = V + VW, Vv = W + VW, ∅ – оба вида отсутствуют.
Длительность стадий и цикличность процесса. Матрица средних времен первого достижения любого состояния (из любого другого и по различным “маршрутам”) содержит соответствующие времена, выраженные в количестве временны́х шагов цепи, а с учетом величины шага Δt = 5 годам они оцениваются приближенно, как показано в табл. 5. Например, для достижения состояния W из V требуется в среднем 111 лет, а для достижения V из W – 50 лет. В совокупности эти показатели отражают цикличность случайного процесса. Так, одни и те же состояния повторяются в среднем за разное (от 11 до 44) количество лет (см. диагональные ячейки табл. 5), а площадка состояния VW достигает состояния ∅ в среднем за 153 года.
Таблица 5.
Средние времена первого достижения/возвращения, выраженные приближенно в годах, и средняя длительность состояния до его смены
| Конечное состояние | Начальное состояние | |||
|---|---|---|---|---|
| ∅ | V | W | VW | |
| ∅ | 44 | 139 | 136 | 153 |
| V | 55 | 17 | 50 | 29 |
| W | 53 | 111 | 33 | 113 |
| VW | 42 | 23 | 29 | 11 |
| Средняя длительность | 18 | 14 | 18 | 20 |
Результаты по средней длительности состояния до его смены приведены в последней строке табл. 5. Видно, что дольше всех – 20 лет – сохраняется сосуществование видов, а наименьшую длительность имеет состояние только с черникой.
ОБСУЖДЕНИЕ
Как отмечалось выше, вопросы построения осредненной модели не получили до сих пор однозначного решения в экологической литературе. В теоретических построениях (например, Tuljapurkar, 1990; Caswell, 2001) существование средней матрицы просто постулируется, а в практических приложениях авторы используют по умолчанию арифметическое среднее (Logofet, 2013), не вдаваясь в смысл осреднения. Принимая гипотезу о случайном характере изменения вероятностей перехода в ряду наблюдений, подобное осреднение следует признать ошибочным, так как оно никак не учитывает различия в размере выборки между разными переходами.
Как было показано ранее, одним из возможных решений проблемы является геометрическое осреднение матриц (Логофет, Маслов, 2018; Logofet, Maslov, 2019). Однако такое решение корректно только при уменьшении числа площадок для расчетов с увеличением числа учетов. Происходит это из-за того, что идеология осреднения и технология калибровки марковской модели предполагают работу с выборкой, число площадок в которой неизменно и сохраняется в каждом учете. По факту число площадок меняется от учета к учету, так как меняется число исключаемых из расчетов площадок с валежом и стволами. Единственным способом добиться равенства числа площадок на всех интервалах учета является исключение площадок, где валеж и стволы встречались хотя бы раз за все интервалы учета.
Поиск геометрического среднего k переходных матриц осложняется также необходимостью извлечения матричного корня k-й степени из их произведения. Уже для k = 5 задача не была тривиальной и сталкивалась с необходимостью привлекать жорданову нормальную форму для корректного вычисления матричного корня (Логофет, Маслов, 2018). В последней версии MATLAB (R2019b) задача решается уже рутинным образом для k = 5, однако корректность рутины при бóльших k остается под вопросом.
Избавлен от подобных трудностей подход, когда от осреднения матриц мы переходим к осреднению вероятностей перехода. На каждом интервале учетов сохраняется максимально возможное число площадок. Так, в изученном сосняке-зеленомошнике число площадок на разных шагах перехода составило от 1758 до 1909 (табл. 1), что позволяет с бóльшей точностью оценить вероятности переходов. При осреднении матриц на каждом шаге размер выборки был бы существенно меньше – 1677. Кроме того, осреднение вероятностей не требует извлечения матричного корня k-й степени из произведения матриц, что позволяет производить расчеты по модели более доступными средствами.
Стохастическая природа марковской модели позволяет примирить такие, казалось бы, противоречивые категории, как направленность и цикличность процесса. Направленность выявляется как сходимость исходного распределения x(1980) к стационарному вектору x* (табл. 3), который оказывается и предельным, а цикличность продемонстрирована расчетами средней длительности состояния площадки до его смены и средних времен возвращения к прежнему состоянию (табл. 5). Средние времена возвращения характеризуют круговорот состояний площадок. Наименьшее среднее время первого достижения/возвращения (11 лет) наблюдается у площадок, где оба вида произрастают совместно (VW), – и это понятно, так как состояние VW максимально преобладает в заключительной стадии. Наибольшее среднее время (153 года) – у перехода из состояния VW в состояние ∅, что тоже понятно, так как состояние ∅ преобладает на самых ранних стадиях.
Наименьшая средняя длительность состояния до его смены (14 лет) наблюдается у площадок только с черникой, но это не связано с временем жизни побегов. По последним данным (Прокопьева и др., 2000; Полянская и др., 2000), продолжительность жизни парциальных кустов брусники составляет 3–5 лет, а черники – 5–8 (до 30 лет). Относительно малое время средней длительности состояния V связано с тем, что такие площадки довольно быстро переходят в состояние VW, а затем снова в V.
Фактические данные и результаты моделирования подтверждают тот факт, что между черникой и брусникой в ходе сукцессии не происходит конкурентного исключения, хотя экологические ниши видов очень близки (Баландина, Вахрамеева, 1978, 1980; Makipaa, 1999) и совпадают на 83% (Grime et al., 1988). Напротив, площадки, где оба вида произрастают совместно, преобладают на поздних стадиях сукцессии (рис. 2). Аналогичный результат был получен в сыром сосняке-долгомошнике (Маслов, Логофет, 2016). Подтверждается сделанный ранее вывод, что сосуществование черники и брусники наблюдается на самых разных уровнях пространственной организации растительного покрова: от самого мелкого до географического (Маслов, 1990).
Причины положительных связей между обилием побегов черники и брусники на мелких площадках в настоящее время неясны. Идеология марковских моделей не требует знания причинных механизмов изучаемого процесса. Такая модель лишь “помогает выявить области, где это понимание особенно важно, т.е. задает и ориентир, и стимул для дальнейшего исследования” (Jeffers, 1978, p. 92). Одним из нескольких потенциально возможных механизмов положительных связей может быть то, что побеги черники, растущие рядом с брусникой, не так охотно поедаются фитофагами. Например, черника является главным зимним кормом красносерой полевки (Clethrionomys rufocanus), тогда как бруснику полевки употребляют значительно реже (Dahlgren et al., 2007).
Результаты моделирования позволяют подробнее описать этапы послепожарной сукцессии на трех разных уровнях детализации: а) динамика синузии кустарничков, б) динамика популяций черники и брусники, в) динамика состояний площадок, отражающих раздельную и совместную встречаемость черники и брусники.
Оценить общее разрастание кустарничков в ходе сукцессии мы можем, просуммировав данные по чернике и бруснике (рис. 3). В момент начала наблюдений (1980 г.) суммарная доля площадок, занятых кустарничками, составляла 37.8%. Через 25 лет, согласно модели, доля площадок с кустарничками возрастает до 80.1% (что отличается от фактического значения всего на 0.7%). В финальном (устойчивом) состоянии доля площадок с кустарничками составит 88.7%. Эти оценки довольно близки к результатам, полученным для сухих сосняков Северной Финляндии, где проективное покрытие кустарничков в разных вариантах сукцессии через 150 лет после пожара составляло 70–80% (Ruokolainen, Salo, 2009).
На уровне популяций отдельных видов мы можем подробнее описать соотношение видов кустарничков в ходе послепожарной сукцессии (рис. 3). С момента начала наблюдений брусника преобладает над черникой на протяжении 6 шагов (30 лет); при этом встречаемость того и другого вида довольно быстро растет (с 31 до 60% у брусники и с 19 до 59% у черники). Действительно, как было показано в экспериментах (Hautala et al., 2009), восстановление у брусники после различного рода нарушений происходит быстрее, чем у черники.
Через 30 лет черника обгоняет бруснику, и разрыв между видами постепенно увеличивается. На 20-м шаге (100 лет) встречаемость черники – 73%, а брусники – 60% (рис. 3). Это хорошо согласуется с выводами В.С. Ипатова с соавторами (1995), относивших чернику и бруснику к группе “нормально-климаксовых” видов, но отмечавших при этом, что брусника (в отличие от черники) является характерным видом не только финальных, но и более ранних стадий сукцессии в сосняках на бедных песчаных почвах. С позиций лесной типологии, такая динамика означает постепенный переход из типа леса “сосняк лишайниково-зеленомошный” в тип леса “сосняк чернично-брусничный” и наконец – в “сосняк бруснично-черничный”, где “возможна смена сосны елью” (Рысин, 1975, с. 126).
Анализ динамики состояний площадок позволяет раскрыть механизмы динамики популяций с момента начала наблюдений. На первых фазах (3 шага – 15 лет) преобладают площадки только с брусникой, на втором месте – площадки с брусникой и черникой, на третьем месте – площадки только с черникой (рис. 2). Через 10 лет процент площадок только с брусникой достигает максимума в 28.5%, после чего начинает снижаться. Напротив, число площадок с черникой и брусникой продолжает быстро расти (рис. 2). На 7-м шаге (35 лет) число площадок только с черникой обгоняет число площадок только с брусникой, и разрыв между ними продолжает быстро расти. По результатам динамики состояний площадок (рис. 2) и с учетом наиболее вероятных направлений переходов (рис. 1) можно выделить четыре стадии совместной динамики популяций кустарничков в сосняке-зеленомошнике, начиная с момента начала полевых наблюдений.
I) Колонизация брусникой “незанятых” площадок (чисто зеленомошных участков) – преобладают площадки “незанятые” и только с брусникой – 2 шага (до 10 лет).
II) Колонизация брусникой “незанятых” площадок еще продолжается, но на ранее захваченные участки подселяется черника – преобладают площадки, где оба вида произрастают совместно – шаги 3–7 (от 10 до 35 лет).
III) Процесс колонизации “незанятых” участков близится к завершению, явно преобладают площадки с черникой и брусникой, и растет число переходов из состояния VW в состояние V (и обратно), процент площадок только с черникой выходит на второе место, и продолжает снижаться число площадок только с брусникой – шаги 8–11 (от 35 до 55 лет).
IV) Стационарное состояние: почти вся территория сосняка захвачена кустарничками, абсолютно преобладают площадки с черникой и брусникой и только с черникой – шаг 12 и далее (с 55 лет). Преобладающий тип переходов VW → V и обратно – V → VW.
Общее время послепожарной сукцессии в двувидовой модели составляет примерно 140 лет: 85 лет от года пожара до начала учетов плюс 55 лет до достижения состояния, отличного от стационарного менее чем на 5%. Для сравнения: в сыром сосняке-долгомошнике время сукцессии после низового пожара (по данным аналогичной модели) составило только 45 лет (Маслов, Логофет, 2016). Различия в скорости изменений, по всей вероятности, связаны с более медленным накоплением гумуса в сухих условиях зеленомошника по сравнению с заболоченным долгомошником. При сравнении одинаковых типов сосняков важно иметь в виду, что в северной части таежной зоны время сукцессий существенно больше из-за низких температур и меньшей продолжительности вегетационного сезона (Hautala et al., 2005; Ruokolainen, Salo, 2009).
ЗАКЛЮЧЕНИЕ
Исследование совместной динамики популяций брусники (Vaccinium vitis-idaea) и черники (V. myrtillus) на средних и поздних стадиях послепожарной сукцессии в заповедном сосняке-зеленомошнике с помощью марковской модели позволило выявить пространственные и временны́е характеристики наблюдаемого процесса. Показано, что частота переходов из одного состояния площадок в другое меняется на разных интервалах наблюдений, однако для массовых вариантов перехода расчет критерия Мантеля–Хензеля показал однородность выборок, что позволило рассчитать усредненные вероятности и перейти к однородной модели.
Показано, что для трех состояний цепи существует однонаправленный тренд изменений во времени: перманентно возрастает доля площадок только с черникой и доля площадок с двумя видами, убывает доля площадок с отсутствием данных видов. Четвертое состояние (площадки только с брусникой) имеет локальный максимум на средних стадиях сукцессии, после чего его доля снижается. Стационарным результатом динамики ожидается распределение площадок по состояниям, где 29.1% площадок занято только черникой, 15.2% – только брусникой, на 44.4% площадок будут присутствовать оба вида, а 11.3% площадок будут незаняты. Это подтверждает возможность устойчивого сосуществования черники и брусники на заключительных стадиях сукцессии и в данном типе сообществ. Преобладание в ходе динамики того или другого вида и различных состояний площадок позволило описать несколько стадий сукцессии: сначала – с малым участием кустарничков, затем – с преобладанием брусники и наконец – с доминированием черники (хотя преобладают площадки, где оба вида произрастают совместно).
Полученные результаты позволяют оценить общую продолжительность послепожарной сукцессии для типа леса “сухой сосняк-зеленомошник” в зоне смешанных лесов в 140 лет (с достижением состояния, отличного от равновесного менее чем на 5%). Данное время почти в 3 раза больше, чем продолжительность послепожарной сукцессии, полученная по данным аналогичной модели в сыром сосняке-долгомошнике (Маслов, Логофет, 2016). С позиций лесной типологии, результат моделирования означает постепенное превращение сосняка лишайниково-зеленомошного в сосняк чернично-брусничный и далее – в сосняк бруснично-черничный.
Таблица 6.
Таблица А.1. Данные о частоте переходов между состояниями в сосняке-зеленомошнике за 25 лет наблюдений, %
| Годы учетов | Куда переход | Переходная матрица | |||
|---|---|---|---|---|---|
| ∅ | V | W | VW | ||
| 1980 ↓ 1985 |
∅ | 76.1 | 4.5 | 8.1 | 0.0 |
| V | 3.9 | 50.4 | 1.9 | 8.3 | |
| W | 18.7 | 1.5 | 67.4 | 1.3 | |
| VW | 1.3 | 43.6 | 22.6 | 90.4 | |
| 1985 ↓ 1990 |
∅ | 70.0 | 5.8 | 4.9 | 0.5 |
| V | 4.8 | 61.6 | 1.5 | 22.0 | |
| W | 22.4 | 0.0 | 71.6 | 1.7 | |
| VW | 2.8 | 32.6 | 22.0 | 75.8 | |
| 1990 ↓ 1995 |
∅ | 70.3 | 8.6 | 7.2 | 1.4 |
| V | 7.9 | 70.2 | 1.2 | 21.9 | |
| W | 18.8 | 2.4 | 73.8 | 2.6 | |
| VW | 3.0 | 18.8 | 17.8 | 74.1 | |
| 1995 ↓ 2000 |
∅ | 67.9 | 4.6 | 4.5 | 0.2 |
| V | 10.2 | 67.5 | 0.8 | 10.2 | |
| W | 17.5 | 1.8 | 75.6 | 5.0 | |
| VW | 4.3 | 26.1 | 19.1 | 84.6 | |
| 2000 ↓ 2005 |
∅ | 69.1 | 7.1 | 6.8 | 1.9 |
| V | 8.0 | 65.3 | 4.3 | 33.2 | |
| W | 17.7 | 2.0 | 70.3 | 5.2 | |
| VW | 5.2 | 25.6 | 18.6 | 59.7 | |
Таблица 7.
Таблица Б.1. Коэффициенты вариации для частот переходов за 25 лет наблюдений, %
| Конечное состояние | Начальное состояние | |||
|---|---|---|---|---|
| ∅ | V | W | VW | |
| ∅ | 4.5 | 28.5 | 24.5 | 98.4 |
| V | 37.1 | 12.3 | 70.5 | 53.0 |
| W | 10.2 | 59.9 | 4.4 | 58.0 |
| VW | 44.5 | 32.0 | 10.6 | 15.2 |
Работа частично поддержана грантом РФФИ № 16-05-00762. Авторы выражают признательность А.В. Рубановичу (ИОГен РАН) за советы из области прикладной статистики.
Приложение А
Данные о частоте переходов между состояниями в сосняке-зеленомошнике за 25 лет наблюдений представлены в табл. А.1.
Приложение В
Пример расчета критерия Мантеля–Хензеля
Согласно критерию Мантеля–Хензеля, выборки можно объединять в одну, если
(В.1)
$\frac{{\sum\limits_{l = 1}^k {{{N}_{l}}{{{({{p}_{l}} - \bar {p})}}^{2}}} }}{{\bar {p}}} < 2k,$Рассчитаем критерий Мантеля–Хензеля на примере переходов из состояния W в состояние VW. Для этого подставим данные из табл. 1 о числе наблюдений и объеме выборки на каждом шаге в табл. В.1, вычислим вероятности переходов (pl) на каждом шаге и просуммируем число наблюдений и объемы выборки согласно формуле (5).
Таблица 8.
Таблица В.1. Данные для расчета критерия Мантеля–Хензеля
| Номер (шаг) перехода, l $ | Число переходов на шаге l, nl | Объем выборки, Nl – 1 | Частота, pl |
|---|---|---|---|
| 1 | 81 | 359 | 0.2256 |
| 2 | 100 | 454 | 0.2203 |
| 3 | 91 | 511 | 0.1781 |
| 4 | 97 | 508 | 0.1910 |
| 5 | 96 | 515 | 0.1864 |
| Итого: | 465 | 2347 |
Тогда, согласно формуле (5), значение средневзвешенной частоты $\bar {p}$ = 465/2347 = 0.1981. А подстановка частот pl и значения $\bar {p}$ в формулу (В.1) дает значение теста Мантеля–Хензеля – 4.02, что меньше 2k = 10. Вывод: объединенная выборка однородна.
Список литературы
Баландина Т.П., Вахрамеева М.Г., 1978. Брусника обыкновенная // Биол. флора Моск. обл. Т. 4. М.: Изд-во МГУ. С. 167–178.
Баландина Т.П., Вахрамеева М.Г., 1980. Черника обыкновенная // Биол. флора Моск. обл. Т. 5. М.: Изд-во МГУ. С. 132–146.
Гантмахер Ф.Р., 1967. Теория матриц. М.: Наука. 576 с.
Горшков В.В., 1995. Послепожарное восстановление мохово-лишайникового яруса в сосновых лесах Кольского полуострова // Экология. № 3. С. 179–183.
Ипатов В.С., Герасименко Г.Г., Кирикова Л.А. и др., 1995. Автогенные сукцессии в сосняке лишайниково-зеленомошном. 1. Фитоценологический анализ видового состава // Ботан. журн. Т. 80. № 9. С. 61–75.
Коротков В.Н., 2002. Биоразнообразие растительного покрова при разных типах пожарных нарушений: результаты наблюдений на стационарных пробных площадях в Окском заповеднике // Мониторинг сообществ на гарях и управление пожарами в заповедниках. М.: ВНИИприроды. С. 47–56.
Кулешова Л.В., Коротков В.Н., Потапова Н.А. и др., 1996. Комплексный анализ послепожарных сукцессий в лесах Костомукшского заповедника (Карелия) // Бюл. МОИП. Отд. биол. Т. 101. № 4. С. 3–15.
Курнаев С.Ф., 1969. Основные типы леса боровых сосняков южной тайги Ярославской области // Сосновые боры подзоны южной тайги и пути ведения в них лесного хозяйства. М.: Наука. С. 11–64.
Логофет Д.О., Маслов А.А., 2018. Анализ мелкомасштабной динамики двух видов-доминантов в сосняке чернично-бруснично-долгомошном. II. Неоднородная марковская цепь и осредненные показатели // Журн. общ. биологии. Т. 79. № 2. С. 135–147.
Маслов А.А., 1990. Количественный анализ горизонтальной структуры лесных сообществ. М.: Наука. 160 с.
Маслов А.А., 2002. Сукцессионная динамика древостоя и нижних ярусов в послепожарном 100-летнем сосняке лишайниково-зеленомошном // Лесоведение. № 2. С. 23–29.
Маслов А.А., Логофет Д.О., 2016. Анализ мелкомасштабной динамики двух видов-доминантов в сосняке чернично-бруснично-долгомошном. I. Однородная марковская цепь и показатели цикличности // Журн. общ. биологии. Т. 77. № 6. С. 423–433.
Огуреева Г.Н., Микляева И.М., Суслова Е.Г., Швергунова Л.В., 1996. Растительность Московской области. Пояснительный текст к карте (м-б 1 : 200 000). М.: Экор. 45 с.
Полянская Т.А., Жукова Л.А., Шестакова Э.В., 2000. Онтогенез черники обыкновенной (Vaccinium myrtillus L.) // Онтогенетический атлас лекарственных растений. Учебное пособие. Т. 2. Йошкар-Ола: Марийский гос. ун-т. С. 51–60.
Прокопьева Л.В., Жукова Л.А., Глотов Н.В., 2000. Онтогенез брусники обыкновенной (Vaccinium vitis-idaea L.) // Онтогенетический атлас лекарственных растений. Учебное пособие. Т. 2. Йошкар-Ола: Марийский гос. ун-т. С. 39–46.
Рысин Л.П., 1975. Сосновые леса европейской части СССР. М.: Наука. 212 с.
Рысин Л.П., Савельева Л.И., 2008. Сосновые леса России. М.: Т-во науч. изд. КМК. 290 с.
Самойлов Ю.И., Ипатов В.С., 1995. Пирогенные сукцессии напочвенного покрова сухих сосняков на песках // Вестн. СПбГУ. Сер. 3. Биол. № 4. С. 58–67.
Справочник по прикладной статистике, 1989 / Под ред. Ллойда Э., Ледермана У. Т. 1. М.: Финансы и статистика. 508 с.
Borenstein M., Hedges L.V., Higgins J.P.T., Rothstein H.R., 2009. Introduction to Meta-Analysis. Oxford: Wiley. 421 p.
Caswell H., 2001. Matrix Population Models: Construction, Analysis, and Interpretation. 2nd ed. Sunderland, MA: Sinauer Associates. 722 p.
Dahlgren J., Oksanen L., Sjödin M., Olofsson J., 2007. Interactions between gray-sided voles (Clethrionomys rufocanus) and bilberry (Vaccinium myrtillus), their main winter food plant // Oecologia. V. 152. № 3. P. 525–532.
Grime J.P., Hodgson J.G., Hunt R., 1988. Comparative Plant Ecology: A Functional Approach to Common British Species. L.: Unwin Hyman. 742 p.
Hautala H., Tolvanen A., Nuortila C., 2009. Regeneration strategies of dominant boreal forest dwarf shrubs in response to selective removal of understorey layers // J. Veg. Sci. V. 12. № 4. P. 503–510.
Hautala H., Kuuluvainen T., Hokkanen T.J., Tolvanen A., 2005. Long-term spatial organization of understorey vegetation in boreal Pinus sylvestris stands with different fire histories // Community Ecol. V. 6. № 2. P. 119–130.
Hytteborn H., Maslov A.A., Nazimova D.I., Rysin L.P., 2005. Boreal forests of Eurasia // Coniferous forests. Amsterdam: Elsevier. P. 23–99.
Jeffers J.N.R., 1978. An Introduction to Systems Analysis: With Ecological Applications. L.: Edward Arnold. 198 p. Русский перевод: Дж. Джефферс, 1981. Введение в системный анализ: применение в экологии. М.: Мир. 252 с.
Keyfitz N., 1968. Introduction to the Mathematics of Population. Reading, MA: Addison-Wesley. 490 p.
Logofet D.O., 2013. Projection matrices in variable environments: λ1 in theory and practice // Ecol. Model. V. 251. P. 307–311.
Logofet D.O., 2018. Averaging the population projection matrices: Heuristics against uncertainty and nonexistence // Ecol. Complex. V. 33. № 1. P. 66–74.
Logofet D.O., Maslov A.A., 2019. Bilberry vs. cowberry in a Scots pine boreal forest: Exclusion or coexistence in a post-fire succession? // Ecol. Model. V. 401. P. 134–143.
Makipaa R., 1999. Response patterns of Vaccinium myrtillus and V. vitis-idaea along nutrient gradients in boreal forest // J. Veg. Sci. V. 10. № 1. P. 17–26.
Ruokolainen L., Salo K., 2009. The effect of fire intensity on vegetation succession on a sub-xeric heath during ten years after wildfire // Ann. Bot. Fenn. V. 46. № 1. P. 30–42.
Tuljapurkar S.D., 1990. Population Dynamics in Variable Environments. N.Y.: Springer. 154 p.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии