

Журнал общей биологии, 2021, T. 82, № 3, стр. 163-174
Экология города: ретроспектива и перспективы изучения
Т. Н. Карманова 1, *, Н. Ю. Феоктистова 1, **, Е.-Е. А. Фетисова 1, ***, А. А. Мосалов 1, 2, ****, А. В. Суров 1, *****
1 Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский просп., 33, Россия
2 Московский педагогический государственный университет
129164 Москва, ул. Кибальчича, 6, Россия
* E-mail: karmanovs95@mail.ru
** E-mail: feoktistovanyu@gmail.com
*** E-mail: e3004e@gmail.com
**** E-mail: rallus@yandex.ru
***** E-mail: surov@sevin.ru
Поступила в редакцию 27.11.2020
После доработки 22.01.2021
Принята к публикации 24.02.2021
Аннотация
В первом десятилетии XXI в. население Земли преимущественно стало городским, а к 2050 г. предполагается, что оно будет составлять уже 68% от общего населения мира. В России к настоящему времени в городах уже проживает 74% населения страны. Однако высокая концентрация населения – это лишь внешнее проявление процесса урбанизации. Ее сопровождают многие глобальные экологические проблемы: нарушение биогеохимических циклов, климата, утрата биоразнообразия, биологические инвазии и пр. Из этого следует особая значимость городов в формировании условий жизни не только для большей части человечества, но и для всех живых существ на планете. Стремительное распространение городских экосистем на планете можно рассматривать как качественно новый этап развития жизни на Земле. Хотя долгое время городская среда считалась недостойной научных исследований в силу своей “искусственности”, сейчас ученые рассматривают города как естественные лаборатории, где можно и нужно решать не только прикладные сугубо градостроительные, но и фундаментальные экологические задачи. В начале 1990-х гг. городская экология приобрела статус самостоятельной науки. В России на актуальность проблем городской экологии ученые обратили внимание еще в 60–70-е годы XX века, но до сих пор ею занимаются лишь единичные исследователи. Обзор посвящен истории формирования городской экологии как биологической науки, адаптациям животных разных таксонов к городским условиям: а) к загрязнениям тяжелыми металлами, шуму, свету и т.д.; б) к дефрагментации мест обитания; в) к кормам антропогенного происхождения. В заключении сформулированы фундаментальные проблемы, которые необходимо решать при изучении урбоценозов, включая оценку скорости эволюции животных и растений в городских условиях.
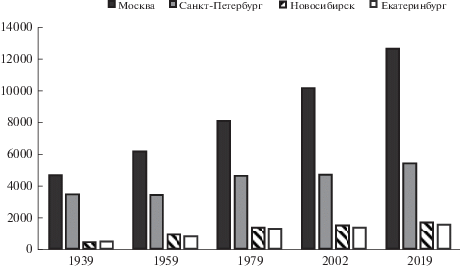
В первом десятилетии XXI в. произошел коренной перелом в соотношении сельского и городского населения Земли – городское стало преобладать над сельским, а к 2050 г. (по данным ООН) будет уже в 2 раза его превышать (United Nations…, 2018). Характерно, что в российских городах, по данным Росстата от 24.01.2020 (https://rosstat.gov.ru/folder/12781), уже сейчас проживает 74% населения страны, при этом особенно быстрыми темпами растет население мегаполисов (рис. 1). Однако высокая концентрация населения – это лишь внешнее проявление процесса урбанизации. Ее сопровождают многие глобальные экологические изменения: нарушение биогеохимических циклов, климата, утрата биоразнообразия, биологические инвазии и т.д., влияние которых распространяется далеко за границы самих городов (Lippe et al., 2005). Поэтому стремительное распространение городских экосистем на планете можно рассматривать как качественно новый этап развития жизни на Земле (Schilthuizen, 2018). Хотя долгое время городская среда считалась недостойной научных исследований в силу своей “искусственности”, сейчас ученые рассматривают ее как естественную лабораторию, где можно и нужно решать не только прикладные сугубо градостроительные, но и фундаментальные экологические задачи (Lahr et al., 2018). Это, например, влияние фрагментации местообитаний (что типично для городской среды) на генетическую структуру видов, ночного освещения на циркадные ритмы людей и животных, зеленых насаждений на здоровье человека, функционирование новых “сугубо городских” сообществ животных и растений, не имеющих естественных аналогов, и многие другие (Dornelas et al., 2014). Таким образом, урбанизацию можно назвать одним из самых глобальных и непреднамеренных экспериментов, который поставило человечество и который позволяет предсказать, как виды будут реагировать на изменения антропогенного характера в масштабах не только городов, но и всей планеты (включая изменения климата) (Lahr et al., 2018).
В начале 1990-х гг. “городская экология” уже приобрела статус самостоятельной науки (Collins et al., 2000; O’Brien, 2012). Ранее городские экосистемы довольно редко становились предметом исследования. Однако в последние десятилетия прослеживается явная тенденция к осознанию городской среды не как к нарушенной, а как к самодостаточной и полноценно функционирующей, где активно протекают эко-эволюционные процессы. Свидетельством того является стремительный рост публикаций на эту тему (с 20 статей в год в 1990-м до 1000 – в 2016‒2017 гг.) (Johnson, Munshi-South, 2017; Rivkin et al., 2019a), появление специализированных журналов (“Environment and Urbanization”, “Journal of Urban Ecology”, “Urban Ecosystems”, “Urban Habitats” и др.), обзорных монографий, ежегодно растущее число научных конференций.
Проведенный нами анализ тематик статей по городской экологии, вышедших в период с 1995 по 2020 год, по базе Web of Science Core Collection (рис. 2) показал, что в отечественной литературе исследований, посвященных этой проблеме очень мало, а работы по эволюции и глобальной экологии единичны, в отличие от зарубежных публикаций. Стоит обратить внимание, что самые ранние работы в области городской экологии были в основном посвящены городскому ландшафту и ботаническим исследованиям. В последнее десятилетие интерес к этим темам сохранился, но также появились статьи, освещающие проблемы глобальной экологии и эволюции (рис. 2а).
Рис. 2.
Распределение по тематикам зарубежных (а) и отечественных (б) статей, относящихся к разделу “городская экология”, выходивших в 1995‒2020 гг., по базе Web of Science Core Collection.
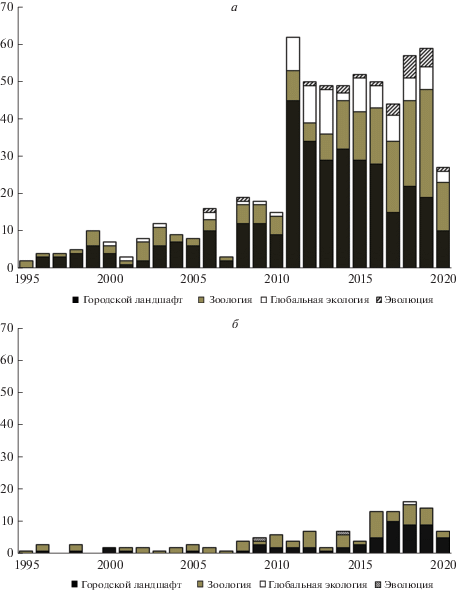
В СССР, а затем и в России, формирование научного интереса к городской экологии связано прежде всего с орнитологическими исследованиями, в частности с изучением врановых. Многие виды этого семейства обитают в урбанизированных ландшафтах, поэтому они оказались удобной моделью для рассмотрения динамики процессов проникновения птиц в города. Подобные работы начались еще в 1960-х гг. и стали основой деятельности Рабочей группы по изучению врановых птиц при Всесоюзном, а затем Мензбировском орнитологическом обществе (Константинов, 2008). Однако следует признать, что, несмотря на активный рост научного интереса к этой теме в мире, в России проблема экологии города получила еще недостаточное освещение в научной литературе – нет ни одного специализированного журнала, в свет выходит мало монографий, посвященных этому вопросу, и, как правило, исследования касаются конкретных городов или таксонов, а не фундаментальных научных проблем. Это подтверждает и анализ публикационной активности отечественных ученых по базе Web of Science (рис. 2б).
Цель данного обзора – привлечь внимание, главным образом отечественных исследователей, к таким проблемам городской экологии, как адаптации растений и животных к существованию в новой экологической нише – городской среде, инвазиям чужеродных видов, эволюционным процессам в условиях города, географии городской среды и др.
ГОРОД КАК НОВАЯ ЭКОЛОГИЧЕСКАЯ НИША ДЛЯ РАСТЕНИЙ И ЖИВОТНЫХ
Для многих видов животных города стали привлекательным местом обитания, так как, несмотря на определенные недостатки, город имеет и преимущества перед естественной средой обитания. Это, например, более мягкие климатические условия, свободные пространства и укрытия, дополнительные (по сравнению с природными) кормовые ресурсы, в том числе за счет насекомых, которых привлекает городское освещение. Считается, что численность насекомых в городах в разы выше, чем в пригороде и в сельской местности (Hall et al., 2017). Экстремальные климатические параметры (температура, влажность) в городах буферизируются, и амплитуды сезонных колебаний сглаживаются. Таким образом, периоды цветения растений и сезоны размножения животных в городах аридной и умеренной зон, как в “псевдотропических пузырьках”, растягиваются (Neil, Wu, 2006). Снижение интенсивности ветра и повышение температуры, особенно в зимнее время, также ведет к повышению биологической продуктивности (Faeth et al., 2011). Большая доступность ограничивающих ресурсов (таких как источники воды в городах аридной зоны) увеличивает и стабилизирует первичную продуктивность, создавая дополнительные ресурсы. Эксперимент, проведенный на сообществах растений в г. Балтиморе (США), показал, что через год после перемещения в городские условия повышается биологическая продуктивность растений, а через пять лет многолетние растения начинают доминировать и вытеснять однолетники (Ziska et al., 2004; George et al., 2009).
Организмы реагируют на урбанизацию на разных уровнях: поведенческом, физиологическом, морфологическом (Le Gros et al., 2016). Фенологическая реакция выражается в более раннем начале цветения, по сравнению с естественной средой обитания, что зафиксировано по крайней мере в 10 городах Европы (Roetzer et al., 2000). Физиологические реакции выражаются в изменении интенсивности фотосинтеза и скорости роста у растений, в изменении численности и устойчивости к высоким температурам у насекомых (Youngsteadt et al., 2015), в сокращении продолжительности спячки у зимоспящих видов животных и т.д. (Surov et al., 2019). В городах меняется динамика численности животных-синурбистов11 – становятся менее выраженными годовые и сезонные пики численности (Тихонова и др., 2012).
Усиление репродуктивных процессов является адаптивной реакцией мелких млекопитающих на увеличение смертности в городе. Так, при общем снижении численности полевки-экономки (Microtus oeconomus) в Сургуте в ответ на застройку естественных местообитаний, отмечено увеличение ее плодовитости (Морозкина, Стариков, 2013). Сходные данные были получены в Оренбурге на домовой мыши (Mus musculus), обыкновенной полевке (Microtus arvalis) и полевой мыши (Apodemus agrarius). Более того, на урбанизированных территориях в размножении начинают участвовать особи более раннего возраста (Шевлюк и др., 2014). Эти данные могут свидетельствовать о популяционном ответе видов на лимитирующие условия городской среды.
Более мягкие климатические условия города формируют некоторые изменения фенологии шмелей. Самки Bombus terrestris, B. pascuorum и B. lapidarius раньше выходят после зимовки по сравнению с загородными популяциями (Попов, 2013).
Изменения условий освещенности трансформируют поведенческие стереотипы у видов-синурбистов. В северных городах Европейской России при длительных зимних ночах серые ворóны (Corvus cornix) используют для поиска корма электрическое освещение улиц. В Москве включение света служит сигналом для массового пролета врановых птиц на ночевку (Константинов, 2015).
Интересно, что животные-синурбисты активно используют “городской материал”, приспосабливая его к своим потребностям. Так, в г. Мехико (Мексика) домовые воробьи (Passer domesticus) и мексиканские чечевицы (Haemorhous mexicanus) используют для гнездостроения окурки. Можно предположить, что таким образом птицы борются с гнездовыми паразитами (Suárez-Rodríguez et al., 2013). Серые вороны в качестве строительного материала для гнезд часто используют предметы хозяйственной деятельности человека. Например, в Архангельске в гнездах ворон находили полиэтиленовую пленку, нитки, стекловату, куски проводов, алюминиевую проволоку, бумагу (Асоскова, Амосов, 2002). Многолетнее гнездо серой вороны высотой около 1 м, снятое с опоры ЛЭП в Москве в 1984 г., в основном было сделано из алюминиевой проволоки. Гнезда сорок, расположенные вблизи радиозавода в Перми, за исключением глиняной чаши состояли из небольших алюминиевых уголков (Константинов, 2015). Похожую картину наблюдали в черте Львова, где сороки использовали для строительства гнезд куски алюминиевой и медной проволоки длиной 20‒50 см (Бокотей, Потапенко, 1990).
Исследуя гнездовой материал в норах обыкновенного хомяка (Cricetus cricetus) в Симферополе, мы также регулярно обнаруживали полиэтиленовые пакеты, елочную мишуру, нетканые салфетки, аккуратно нарезанные животными на ленты и подготовленные для использования в качестве гнездовой подстилки.
Даже эти немногочисленные примеры свидетельствуют о том, что город формирует принципиально новые условия существования, которые не только выявляют скрытые адаптационные возможности вида, но и обуславливают протекание эволюционных процессов, о чем подробнее будет сказано ниже. Таким образом, история городов – это также история новых типов экосистем (Sukopp, 2003), а критическое воздействие урбанистических факторов на климат, гидрологию и почвы в конечном итоге приводит к формированию новых моделей биоразнообразия (Lippe et al., 2005).
ИНВАЗИИ КАК ФАКТОР РАЗВИТИЯ ГОРОДСКИХ ЭКОСИСТЕМ
В последние десятилетия крупные города были признаны очагами фиторазнообразия: флора городов обычно богаче, чем окружающие территории (Pyšek, 1993; McKinney, 2006). Это связано главным образом с увеличением доли адвентивных видов растений, которая в городах Центральной Европы может достигать 40‒50% (Pyšek, 1998; Kühn et al., 2004; Kowarik, 2020) как за счет активного вселения в города инвазивных видов растений (например, с транспортом), так и декоративного озеленения. С другой стороны, большая неоднородность антропогенной среды, особенности городского климата, почв, гидрологии сами по себе способствуют распространению таких видов (Lippe et al., 2005). Было обнаружено, что количество автохтонных видов уменьшается от окраин к городским центрам. Например, в Мехико наблюдалось линейное снижение числа видов растений с 30–80 видов на гектар в пригородах до 3–10 в центре города. Полосы отчуждения железнодорожных путей и автомагистралей усиливают присутствие чужеродных видов в городах (Pickett et al., 2011).
Урбанизация приводит к тому, что большая часть аборигенных видов растений заменяется относительно небольшим числом широко распространенных и агрессивных видов, часто не встречающихся в пригородах. Международный научный консорциум под названием GLUSEEN (Глобальная сеть экологии и образования почв города) провел исследования ДНК почвенных микроорганизмов в городах и природных зонах Африки, Северной Америки и Европы и обнаружил, что видовой состав городских почв на этих континентах практически идентичен. По крайней мере, сообщества 12 000 видов грибов и 3700 видов архей в городах гораздо более сходен, чем в окружающих их биотопах.
В городах Америки (например, в Балтиморе) около 80% высших растений, обитающих около корней уличных деревьев, были идентичны тем, которые встречаются в европейских городах. Все это означает, что урбоценозы по всему миру становятся все более похожими; сообщества растений и животных, грибов, одноклеточных организмов и вирусов постепенно приближаются к единому глобализированному городскому биоразнообразию. Этот процесс носит название биотической гомогенизации (McKinney, 2006; Olden, Rooney, 2006). В дальнейшем адвентивные виды начинают проникать из центра на окраины, вытесняя автохтонные виды уже и в пригородах. Таким образом, города сами становятся источниками распространения инвазивных видов растений (Müller, Werner, 2010).
Несмотря на увеличивающееся видовое богатство и разнообразие, происходящее за счет сосуществования адвентивных и аборигенных видов, флора разных городов становится все более и более сходной.
Естественно, что урбанизация отражается и на биоразнообразии городской фауны. Так, распространение тротуарных муравьев (Tetramorium caespitum) в Северной Америке является классическим примером успешной натурализации в городах. Около 200 лет назад этот вид был завезен в Соединенные Штаты с почвой, которая использовалась в качестве балласта на торговых судах. В настоящее время он является одним из самых распространенных видов муравьев в городах и пригородах вдоль побережья Атлантического океана от Канады до Флориды (King, Green, 1995).
Ожереловые попугаи Крамера (Psittacula krameri) были завезены в Европу и США из Африки и Азии в качестве домашних питомцев. В течение последних десятилетий они, покидая клетки, создавали колонии в городах Старого и Нового Света. Уже из городов попугаи начали расселяться в сельские районы, становясь серьезными вредителями зерновых культур. Более того, одичавшие птицы негативно влияли на гнездование аборигенных видов – удодов, сычей (Butler, 2005; Strubbe, Matthysen, 2007).
Домовый воробей, вероятно, возник на Ближнем Востоке (в настоящее время “дикие” популяции этого вида неизвестны) и распространился вместе с сельским хозяйством по большей части Евразии и в некоторых частях Северной Африки. С середины XIX в. он захватил большую часть мира. Космополитизм этого вида объясняется множеством факторов, включая крупный размер мозга, большое количество поведенческих адаптаций, комменсализм по отношению к человеку и т.д. (Garber, 1998). Воробьи не только адаптировались к жизни в городах, но и стали серьезной проблемой для местной орнитофауны. Споры о ценности этого вида, допустимости его сохранения и вреде, наносимом им, часто выходили за рамки научных дискуссий и стали известны как “война воробьев”. В метафорическом смысле стоял вопрос даже о том, можно ли переносить проблему воробьев-вселенцев на мигрантов в широком смысле (Fine, Christoforides, 1991).
Подобно домовому воробью, есть и другие виды, которые настолько широко распространились по городам мира, что без них уже практически невозможно представить себе городской ландшафт. Это, например, сизые голуби (Columba livia domestica), которых начали одомашнивать как минимум 3–4 тыс. лет назад. Проводилось их селекционное разведение для улучшения отдельных экстерьерных признаков, и выводились породы для практического использования (Kelcey, Rheinwald, 2005). Одомашненные голуби преднамеренно выпускались или сами покидали клетки и распространились в городах по всему миру. Сейчас в центрах многих городов биомасса диких голубей на порядки выше, чем в естественных местообитаниях (Nuorteva, 1971). Среди положительных аспектов можно выделить то, что голуби подъедают остатки пищи человека и тем самым очищают улицы города от болезнетворных организмов. Однако в помете этих птиц было найдено более 60 возбудителей болезней, хотя случаи заражения от голубей человека единичны.
Чайки, как и голуби, за последние десятилетия все активнее осваивают города, но в отличие от последних, они в основном концентрируются на городских свалках. Однако случаи гнездования чаек непосредственно в урбанизированной среде были крайне редки. Но в 1998‒1999 гг. В.А. Зубакин обнаружил гнезда чаек вдали от водоемов и на изначально сухих местах – на залитой гудроном крыше прядильно-ниточной фабрики на берегу р. Клязьмы недалеко от центра Ногинска (Зубакин, 2001). Таким образом, чайки также демонстрируют склонность к синурбанизации.
На протяжении последних 50 лет сохраняется тенденция к увеличению количества и размеров популяций серой вороны в Москве. Интенсивное заселение городов этим видом произошло во второй половине XX в. и охватило многие города Европейской России (Константинов, 2008).
В 1820-х гг. отмечено появление и гнездование черных дроздов (Turdus merula) в Риме. 200 лет назад этот вид являлся исключительно лесным, но с начала XIX в. его ареал начал расширяться из европейских лесных массивов и они начали колонизировать растущие города (Luniak et al., 1990). К концу XIX в. эти птицы стали обычными в городах Центральной Европы, а в XХ в. достигли Лондона, городов Исландии и Северной Скандинавии. В течение всего этого периода распространение городских популяций происходило со средней скоростью 8 км в год. В городах этот вид приобрел ряд новых черт: птицы перестали совершать сезонные миграции и стали размножаться на месяц раньше, чем вне городов. Черные дрозды стали столь обычными в городах, что их предложили называть “городскими черными птицами” (Schilthuizen, 2018).
Таким образом, можно сказать, что у городской фауны и флоры прослеживаются сходные тенденции ‒ происходит увеличение биоразнообразия по сравнению с естественными биотопами.
Безусловно, накопление подобных фактов поможет проследить формирование биологического разнообразия городов и проанализировать связи в сообществах растений и животных. Это даст возможность прогнозировать возможные изменения в городских экосистемах, когда урбанизация будет все больше захватывать пригородные территории и сильнее влиять на сложившиеся местные экосистемы.
ЭВОЛЮЦИЯ В ГОРОДЕ
Как дисциплина, городская экология не может рассматриваться в отрыве от эволюционных процессов, происходящих в урбоценозах. Очевидно, что фрагментация и деградация естественных биотопов в городах ведут к уменьшению размеров и усилению изоляции аборигенных популяций, подобно “островному эффекту” (Faeth, Kane, 1978; Haddad et al., 2015; Феоктистова и др., 2016, 2019). Урбанизация или, точнее, синурбанизация влияет на адаптивные (естественный и половой отбор) и неадаптивные (дрейф генов и перенос генов) эволюционные процессы (Johnson, Munshi-South, 2017). Изменения в биотической и абиотической среде, происходящие по градиенту урбанизации, могут существенно ускорять естественный отбор и стимулировать городскую адаптивную эволюцию (Hendry et al., 2008; Donihue, Lambert, 2015; Alberti et al., 2017; Schilthuizen, 2018; Rivkin et al., 2019b). Фактически города можно рассматривать как глобальные экспериментальные площадки для изучения эволюции, заметной здесь практически невооруженным глазом. По образному выражению голландского исследователя Шильхузена, города являются “скороварками эволюции” (Schilthuizen, 2018).
Классическим примером городской эволюции, который попал во все учебники экологии, является промышленный меланизм березовой пяденицы (Biston betularia), обнаруженный в городах Англии в конце XIX в. (Tutt, 1896; Kettlewell, 1955). При сильном загрязнении копотью березовых стволов всего за 100 лет черная морфа березовой пяденицы сильно потеснила ранее гораздо более широко распространенную белую. Однако после принятия законодательства по ограничению загрязнения воздуха в 1950-х гг. процесс быстро пошел в обратную сторону, и к настоящему времени белые морфы Biston betularia снова преобладают (Cook, Saccheri, 2013). Интересно, что меланизм березовой пяденицы обеспечивается одной мутацией, обусловленной переходом 22000 н.п. из одной хромосомы в другую (Van’t Hof et al., 2016).
Похожее явление формирования меланизма может быть рассмотрено на примере сизого голубя в Москве. По мере роста численности птиц в городе наблюдалась тенденция к увеличению доли меланистических форм и вытеснению светлоокрашенных и пестрых птиц из популяций. Адаптивный характер подобного явления заключается в том, что темные морфы оказались более приспособленными к существованию в больших скоплениях – их гнезда располагались ближе друг к другу, чем у светлых форм (Обухова, 2016). Исследования, проведенные на сизом голубе в ряде европейских стран, показали, что в пере темных голубей содержание цинка на 25% больше, чем у светлоокрашенных. Предполагается, что городские голуби эволюционируют в сторону более темного оперения, благодаря детоксикационным свойствам перьев, наполненных меланином (Chatelain et al., 2016). Но, возможно, история сложнее, потому что гены, вырабатывающие меланин, также участвуют в регуляции гормона стресса и иммунной системы (Schilthuizen, 2018). Таким образом, окраска птиц и содержание тяжелых металлов в окружающей среде могут быть частью более сложной системы, в которой задействованы иммунная и гормональная регуляция.
В то же время у других видов-синурбистов подобные тенденции не отмечены. Так, не выявлена зависимость между частотой проявления лейкизма (частичная утрата пигментации) у серых ворон и уровнем загрязнения на урбанизированных территориях Москвы. Однако частота встречаемости птиц с лейкизмом возрастает с повышением плотности населения птиц и в период вылета молодых особей из гнезда (Фетисова, Мосалов, 2019).
Загрязнения могут быть не только промышленными, но и просто связанными со спецификой городских условий. К ним относятся шумовое, световое, тепловое и другие загрязнения. Очевидно, что к их присутствию животным-синурбистам тоже необходимо приспосабливаться. Так, например, большая синица (Parus major) в Европе, китайский настоящий бюльбюль (Pycnonotus sinensis) в Азии, рыжеошейниковая овсянка (Zonotrichia capensis) в Южной Америке, серебряная белоглазка (Zosterops lateralis) в Австралии и многие другие в городе стали петь громче и на более высокой частоте по сравнению с загородными популяциями, что связывают с необходимостью сохранения функциональности песен в условиях высокого уровня городского шума (Slabbekoorn, Peet, 2003). И такой эффект отмечен не только у птиц, но, например, у австралийской квакши (бурой литории Litoria ewingii) (Parris et al., 2009) и кузнечика Chorthippus biguttulus (Lampe et al., 2014). Более того, отмечено, что птицы в городах начинают петь по утрам в среднем на 50 мин раньше, чем за городом, пока еще не достигнут высокий уровень шума. Считается, что этот показатель связан еще и со световым загрязнением (Nemeth et al., 2013).
Постоянное освещение является одним из серьезных факторов, оказывающих негативное влияние на животных в городе. Однако некоторые виды смогли использовать и этот фактор в “своих интересах”. Например, взрослые самки мостовых пауков (Larinioides sclopetarius) предпочитают искусственно освещенные участки (под фонарями) для строительства ловчих сетей неосвещенным территориям. Пауки в искусственно освещенной “среде обитания” ловили в свою паутину в 4 раза больше добычи, чем те, которые строили ее в темных местах. И такое поведение предопределено генетически: выращенные в лаборатории потомки от самок, отловленных в городе, предпочитали искусственное освещение естественному (Heiling, 1999). А горностаевая моль (Yponomeuta cagnagella) из городских популяций, подвергающихся сильному световому загрязнению в течение поколений, реже летела на свет, чем ее деревенские сородичи. То есть обитающая в городе горностаевая моль развила способность не привлекаться светом, что пока является уникальным примером подобной адаптации для насекомых (Altermatt, Ebert, 2016).
Кроме светового и шумового загрязнений для городов характерно химическое загрязнение, в том числе загрязнение полициклическими ароматическими углеводородами (ПАУ), полихлорированными дифенилами (ПХБ), которые “прилипают” к белку AHR, действующему как регулятор программы развития эмбрионов. ПХБ и ПАУ постоянно вмешиваются в работу AHR, так что программы или включаются слишком рано или вовремя не выключаются. В таких условиях некоторые виды рыб, в частности небольшая хищная рыбка из семейства фундуловых ‒ фундулус гетероклитус (Fundulus heteroclitus), распространенная вдоль Атлантического побережья и в водоемах США, Канады и на Бермудских островах, приспособились выживать в воде с концентрацией этих опасных веществ в 8000 раз превышающих ПДК для других видов рыб (Frederick et al., 2007; Whitehead et al., 2011). Оказалось, что у рыб, обитающих в загрязненных водоемах Нью-Бедфордской гавани в Массачусетсе и в Бриджпорте (США), возникли мутации в генах белков AHR, которые выключили работу участка гена таким образом, что рыбки больше не реагируют на отравляющие вещества. Интересно, что эти мутации варьировали в разных популяциях фундулусов в зависимости от особенностей загрязненных участков. Это может означать, что эволюция неоднократно приводила к формированию устойчивости к ПХБ. Пример с F. heteroclitus стал классическим для демонстрации устойчивости живых организмов к ПХБ и ПАУ, сформировавшейся всего за два десятка поколений (Reid et al., 2016). Другой пример устойчивости к загрязнению вод ПХБ демонстрирует атлантический томкод (Microgadus tomcod). Показано, что 25% особей имеют патологии печени – неопластические узелки и гепатоцеллюлярную карциному, но тем не менее продолжают существовать в условиях загрязнения (Wirgin et al., 2011).
Группа ученых из Норвегии, Ирана и Казахстана провела анализ генов, отвечающих за расщепление крахмала у домового воробья. Было показано, что этот вид в течение примерно 6 тыс. лет сформировал адаптации к существованию в антропогенной среде. В числе прочего, это экспрессия гена AMY2A, который отвечает за усвоение кормов с высоким содержанием крахмала, т.е., по сути, питание обработанными человеком продуктами (хлебом) (Ravinet et al., 2018).
И это не единственный пример адаптаций, возникших у животных, питающихся в городах не типичной для них пищей. У мексиканской чечевицы, обитающей в г. Тусон (США) и питающейся семенами подсолнечника на кормушках для птиц, шел отбор по морфологии клюва – городские особи имели более широкие клювы, чем деревенские птицы, что обеспечивало им необходимую силу сжатия при обработке семян (Badyaev et al., 2008).
В 2017 г. в журнале “Proceedings of the National Academy of Sciences” группа авторов опубликовала глобальный анализ более 1600 случаев “фенотипических” изменений различных организмов и показала, что в городах они происходят быстрее, чем вне городов, и что наиболее сильными движущими силами эволюции являются столкновения второго рода (взаимодействия с самим человеком или с другими организмами, привнесенными им в город) (Schilthuizen, 2018).
Вопросы об эволюционных механизмах, действующих в антропогенно нарушенной среде, требуют более детального обсуждения, а также серьезного изучения на большем количестве видов.
ТРАНСФОРМАЦИЯ ПОВЕДЕНЧЕСКИХ СТЕРЕОТИПОВ В ГОРОДАХ
Существование в городах множества видов-синантропов с устойчивыми поведенческими и экологическими характеристиками, позволяющими им жить рядом с человеком тысячи лет (крысы, мыши, клопы, тараканы и пр.), указывает на то, что эволюция этих видов в городской среде уже произошла (Johnson, Munshi-South, 2017; Thompson et al., 2018). Урбанизация требует разрушения имеющихся поведенческих стереотипов животных. Особенно это касается таких поведенческих характеристик, как толерантность к присутствию человека, внутривидовая агрессия, исследовательское поведение, добывание пищи и др. (Luniak, 2004).
На 42 видах птиц Австралии было показано, что дистанция вспугивания при приближении человека у всех исследованных видов в городе меньше, чем в деревенских биотопах. При этом чем дольше птицы жили в городах, тем сильнее была выражена толерантность к человеку. Например, галки (Corvus monedula), которые колонизировали города еще в 1880-х гг., взлетают при приближении человека с 8 м в условиях города и с 30 м – в сельской местности. С другой стороны, большие пестрые дятлы (Dendrocopos major) начали заселять города с 1970-х гг., и разницы в дистанции вспугивания у них практически одинаковые в городе и в деревне (Symonds et al., 2016). При этом выраженность этого параметра пропорциональна времени с начала освоения видом городских ландшафтов (Minias, Janiszewski, 2016).
На многих видах птиц показано, что за счет повышения плотности населения в городах растет внутривидовая агрессивность. Так, лысухи (Fulica atra) в условиях города становятся агрессивнее друг к другу, охраняя гнезда от сородичей. Подобное отмечалось и у певчего воробья (Melospiza melodia) (Scales et al., 2011). У самцов (но не самок) водяных ящериц (Intellagama lesueurii) агрессивность в городе также была выше, чем в сельской местности, однако сила укуса не увеличивалась, как это обычно бывает у ящериц в условиях высокой плотности (Baxter-Gilbert et al., 2019).
В городе может меняться и стратегия охотничьего поведения. Популяции ястреба-тетеревятника (Accipiter gentilis) начали заселять города СССР с 1980-х гг. Эти птицы в городе начали проявлять нетипичные способы охоты, например, добывая жертвы на чердаках, в сумерках, “мышкованием” среди травы (Еремкин, Очагов, 1998; Матус, 2003).
Особые условия кормообеспеченности в урбоценозах требуют формирования специфических поведенческих адаптаций. Так, черные вороны (Corvus corone) в городах Японии употребляют в пищу широко распространенный здесь японский орех (Juglans ailantifolia), отличающийся особо твердой скорлупой. Чтобы добраться до ядра птицы научились бросать целые орехи под колеса автомобилей, стоящих на светофорах. Впервые этот факт был установлен в 1975 г. в городе Сендай, а затем и во многих других городах Японии. Анализ множества таких случаев привел к заключению, что такое поведение является преднамеренным, и птицы способны обучаться этому навыку друг от друга (“культурные традиции”) (Nihey, 1995).
Другим, ставшим уже харизматическим примером поведенческой адаптации к городским условиям являются обыкновенные лазоревки (Cyanistes caeruleus). В 1930-х гг. в ряде английских городов птицы научились расклевывать крышки на бутылках с молоком и поедать отстоявшиеся сверху сливки (Hinde, Fisher, 1951).
ГОРОДСКАЯ ГЕОГРАФИЯ
Отдельного внимания заслуживает влияние особенностей урбанизированного ландшафта на экологию города. Фрагментация города создает условия, которые изолируют популяции и препятствуют перемещению животных между ними. Теория островной биогеографии использовалась для объяснения изменений биоразнообразия в городах в связи с размерами, сложностью, расстоянием до исходных популяций. Небольшие и удаленные друг от друга демы включают меньше особей из-за ограничения миграции и недостаточности ресурсов для поддержания минимальной численности, что в конечном счете может привести к вымиранию местных популяций (Müller, Werner, 2010).
На городских популяциях животных нередко можно наблюдать “эффект бутылочного горлышка” и “эффект основателя”. Исследование обыкновенного хомяка в Симферополе показало, что, несмотря на то, что единичные особи могут перемещаться по городу на большие расстояния, городская популяция представлена отдельными демами с высокой степенью генетической обособленности и сниженным по сравнению с установленным для города в целом генетическим разнообразием (Феоктистова и др., 2016, 2019).
Фрагментация городской среды приводит к снижению генетического разнообразия и большей генетической дифференциации среди городских популяций белоногого хомячка (Peromyscus leucopus) (Munshi-South et al., 2016) и льнянки обыкновенной (Linaria vulgaris) (Bartlewicz et al., 2015) по сравнению с загородными популяциями.
Преградой для перемещения особей обыкновенной стенной ящерицы (Podarcis muralis) в г. Трире (Германия) является р. Мозель. Здания и дороги обеспечивают ограничение потока генов через город, тогда как городские виноградники и каменистые субстраты, напротив, облегчают его (Beninde et al., 2016).
Отдельные популяции темных (Desmognathus fuscus) в г. Нью-Йорке (Lourenço et al., 2017), красноспинных (Plethodon cinereus) в г. Монреале (Канада; Munshi-South et al., 2013) и огненных (Salamandra salamandra) в г. Овьедо (Испания; Noël et al., 2007) саламандр демонстрируют более низкое генетическое разнообразие, чем в менее урбанизированных районах (Johnson, Munshi-South, 2017).
Нередко из-за сильной гетерогенности ландшафта в городах можно встретить и изолированные от антропогенного воздействия территории, которые также испытывают влияние изоляции. Яркий пример – Бобровый остров, образовавшийся в конце 1960-х гг. в Нагатинской пойме р. Москвы (Пчелкина и др., 2013). Для посетителей он доступен только зимой, когда река замерзает, в остальное время это практически изолированная территория. Здесь, всего в нескольких километрах от Кремля, успешно сосуществуют бобры, ласки, ондатра и многие другие виды млекопитающих и птиц. Такое уникальное место, безусловно, перспективно для разнообразных научных работ.
В некоторых случаях городские парки выступают в качестве источников генетического разнообразия для окружающих территорий. Так, например, популяция большой синицы (Parus major), обитающей в Барселоне (Испания), содержит больше генетических линий в городских парках, чем в близлежащих лесах, и поток генов из городских в пригородные популяции интенсивнее, чем в обратном направлении (Björklund et al., 2010). Лесопарки, расположенные в черте города, являются оазисами для животных, обитающих в загородных лесных массивах, но посещающих город в поисках пропитания. В первую очередь это относится к птицам, так как они более мобильны. Организмы меньших размеров и со слабыми способностями к расселению, вероятно, острее испытывают потерю генетического разнообразия.
Поток генов может поддерживать генетическое разнообразие и уменьшать генетическую дифференциацию между городскими популяциями, но может также замедлять формирование адаптаций к существованию в городе.
ЗАКЛЮЧЕНИЕ
Считается, что городские экосистемы отличаются от своих “естественных” аналогов только степенью влияния человека (Walbridge, 1997). Однако многие экологические процессы в городской среде выражены даже ярче, чем в природе (Niemela, 1999). Города представляют собой уменьшенные модели глобальных экосистем, и благодаря этому глобальное потепление оказывает влияние на городские организмы в более сжатые сроки. Исследование влияния климатических изменений на экосистемы именно в городах позволит в мировом масштабе сохранить ресурсы и время для решения выявленных проблем (Lahr et al., 2018). Так, в городах создаются условия, в которых, вероятно, в будущем будет существовать биосфера. Это, например, повышение содержания диоксида углерода, более высокие температуры, засуха, загрязнения почв. Таким образом, изучение влияния процессов урбанизации на экосистемы городов должно помочь прогнозировать возможные сценарии развития жизни на Земле.
Представляют интерес наблюдения за городскими сообществами растений и животных для оценки влияния климатических изменений в целом на трофические сети. Кроме этого, изучение городских экосистем позволит прогнозировать возможные изменения ареалов некоторых видов животных по мере изменения климатических условий. Так, например, изучение фауны городов Западной Сибири показало, что на юге идет замещение сухолюбивых видов млекопитающих на влаголюбивые в результате развития гидрологической сети в южных городах. На севере наблюдается обратная ситуация в связи с аридизацией местообитаний (Гашев, 2000).
Плохо изучена степень влияния адвентивных видов на эндемичные экосистемы и возможные последствия инвазий для мирового биоразнообразия. Не ясно, являются ли адаптации, возникающие у живых организмов в городской среде прямым или опосредованным следствием обитания в данных условиях. Не установлена связь ограничения потока генов и скорости эволюции в городах и экосистемах в целом.
Работа выполнена в рамках Государственного задания Института проблем экологии и эволюции им. А.Н. Северцова Российской академии наук № АААА-А18-118042690110-1.
Список литературы
Асоскова Н.И., Амосов П.Н., 2002. Изменение поведения серой вороны в урбанизированных ландшафтах севера таежной зоны Архангельской области // Экология врановых птиц в антропогенных ландшафтах. Мат-лы VI науч.-практ. конф. по врановым. Саранск. С. 9‒13.
Бокотей А., Потапенко В., 1990. О гнездовании сорок в металлических гнездах в черте г. Львова // Орнитология. Т. 24. С. 123.
Гашев С.Н., 2000. Млекопитающие в системе экологического мониторинга (на примере Тюменской области). Тюмень: ТюмГУ. 220 с.
Еремкин Г., Очагов Д., 1998. О тенденциях в изменении численности птиц Москвы и ближнего Подмосковья // Природа Москвы. М.: Биоинформсервис. С. 170‒176.
Зубакин В.А., 2001. Необычное гнездование чайковых птиц в Московской области в 1998 и 1999 годах // Орнитология. Т. 29. С. 291‒293.
Константинов В.М., 2008. Краткая история исследований врановых птиц // Рус. орнитол. журн. Т. 17. № 407. С. 415‒425.
Константинов В.М., 2015. Особенности синантропизации и урбанизации врановых птиц // Рус. орнитол. журн. Т. 24. № 1177. С. 2892‒2901.
Матус А., 2003. Ястреб-тетеревятник в Киеве // Беркут. Т. 12. № 1‒2. С. 66‒70.
Морозкина А., Стариков В., 2013. Особенности экологии полевки-экономки города Сургута // Мир науки, культуры, образования. № 5 (42). С. 450‒453.
Обухова Н.Ю., 2016. Городские голуби: полиморфизм и стратегии выживания // Природа. № 9. С. 42‒50.
Попов И.Б., 2013. Фенологические стратегии шмелей (Hymenoptera, Apidae, Bombus Latr.) в условиях различных экосистем Северо-Западного Кавказа // Тр. Рус. энтомол. об-ва. Т. 84. С. 116‒122.
Пчелкина Т.А., Кухта А.Е., Пчелкин А.В., 2013. Формирование биоты на островах кваторий мегаполиса Москвы // Общество. Среда. Развитие (Terra Humana). Т. 2. № 27. С. 243‒250.
Тихонова Г.Н., Тихонов И.А., Суров А.В., Богомолов П.Л., Котенкова Е.В., 2012. Экологические аспекты формирования фауны мелких млекопитающих урбанистических территорий Средней полосы России. М.: КМК. 373 с.
Феоктистова Н.Ю., Мещерский И.Г., Суров А.В., Богомолов П.Л., Товпинец Н.Н., Поплавская Н.С., 2016. Генетическая структура городской популяции обыкновенного хомяка (Cricetus cricetus) // Генетика. Т. 52. № 2. С. 221‒230.
Феоктистова Н.Ю., Мещерский И.Г., Богомолов П.Л., Мещерский С.И., Поплавская Н.С. и др., 2019. Обыкновенный хомяк (Cricetus cricetus) в Предкавказье: генетическая структура городских и пригородных популяций // Генетика. Т. 55. № 3. С. 337‒348.
Фетисова Е.-Е.А., Мосалов А.А., 2019. Проявление лейкизма в популяциях серой вороны (Corvus cornix L.) в г. Москве // Экология врановых птиц в естественных и антропогенных ландшафтах Северной Евразии. Мат-лы XII Всерос. конф. с междунар. участием. Кисловодск, 26‒28 сентября 2019 г. Казань: ООО “Олитех”. С. 164‒166.
Шевлюк Н.Н., Блинова Е.В., Боков Д.А., Обухова Н.В., Сивожелезова Н.А. и др., 2014. Морфофункциональные особенности размножения мелких млекопитающих в условиях урбанизированной среды обитания на примере г. Оренбурга // Изв. ОГАУ. Т. 46. № 2. С. 201‒203.
Alberti M., Correa C., Marzluff J.M., Hendry A.P., Palkovacs E.P. et al., 2017. Global urban signatures of phenotypic change in animal and plant populations // Proc. Natl. Acad. Sci. V. 114. № 34. P. 8951‒8956.
Altermatt F., Ebert D., 2016. Reduced flight-to-light behaviour of moth populations exposed to long-term urban light pollution // Biol. Lett. V. 12. № 4. Art. 20160111.
Andrzejewski R., Babiñska-Werka J., Gliwicz J., Goszczyñski J., 1978. Synurbization processes in an urban population of Apodemus agrarius. I. Characteristics of population in urbanization gradient // Acta Theriol. V. 23. P. 341–358.
Badyaev A.V., Young R.L., Oh K.P., Addison C., 2008. Evolution on a local scale: Developmental, functional, and genetic bases of divergence in bill form and associated changes in song structure between adjacent habitats // Evolution. V. 62. № 8. P. 1951‒1964.
Bartlewicz J., Vandepitte K., Jacquemyn H., Honnay O., 2015. Population genetic diversity of the clonal self-incompatible herbaceous plant Linaria vulgaris along an urbanization gradient // Biol. J. Linn. Soc. V. 116. № 3. P. 603‒613.
Baxter-Gilbert J., Riley J.L., Whiting M.J., 2019. Bold New World: Urbanization promotes an innate behavioral trait in a lizard // Behav. Ecol. Sociobiol. V. 73. № 8. P. 105.
Beninde J., Feldmeier S., Werner M., Peroverde D., Schulte U. et al., 2016. Cityscape genetics: Structural vs. functional connectivity of an urban lizard population // Mol. Ecol. V. 25. № 20. P. 4984‒5000.
Björklund M., Ruiz I., Senar J.C., 2010. Genetic differentiation in the urban habitat: The great tits (Parus major) of the parks of Barcelona city // Biol. J. Linn. Soc. V. 99. № 1. P. 9‒19.
Butler C.J., 2005. Feral parrots in the continental United States and United Kingdom: past, present, and future // J. Avian Med. Surg. V. 19. № 2. P. 142‒149.
Chatelain M., Gasparini J., Frantz A., Chatelain J., 2016. Do trace metals select for darker birds in urban areas? An experimental exposure to lead and zinc // Glob. Change Biol. V. 22. P. 2380–2391.
Collins J., Kinzig A., Grimm N., Fagan W., Hope D. et al., 2000. A new urban ecology // Am. Sci. V. 88. № 5. P. 416‒425.
Cook L., Saccheri I., 2013. The peppered moth and industrial melanism: Evolution of a natural selection case study // Heredity. V. 110. № 3. P. 207‒212.
Donihue C.M., Lambert M.R., 2015. Adaptive evolution in urban ecosystems // Ambio. V. 44. № 3. P. 194‒203.
Dornelas M., Gotelli N.J., McGill B., Shimadzu H., Moyes F. et al., 2014. Assemblage time series reveal biodiversity change but not systematic loss // Science. V. 344. № 6181. P. 296‒299.
Faeth S.H., Kane T.C., 1978. Urban biogeography ‒ city parks as islands for Diptera and Coleoptera // Oecologia. V. 32. № 1. P. 127‒133.
Faeth S.H., Bang C., Saari S., 2011. Urban biodiversity: Patterns and mechanisms // Ann. NY Acad. Sci. V. 1223. № 1. P. 69‒81.
Fine G.A., Christoforides L., 1991. Dirty birds, filthy immigrants, and the English sparrow war – metaphoric linkage in constracting social problems // Symb. Interact. V. 14. № 4. P. 375‒393.
Frederick L.A., Veld P.A., van, Rice C.D., 2007. Bioindicators of immune function in creosote-adapted estuarine killifish, Fundulus heteroclitus // J. Toxicol. Environ. Health A. V. 70. № 17. P. 1433‒1442.
Garber S.D., 1998. The Urban Naturalist. L.: Dover Publications. 256 p.
George K., Ziska L.H., Bunce J.A., Quebedeaux B., Hom J.L. et al., 2009. Macroclimate associated with urbanization increases the rate of secondary succession from fallow soil // Oecologia. V. 159. № 3. P. 637‒647.
Haddad N.M., Brudvig L.A., Clobert J., Davies K.F., Gonzalez A. et al., 2015. Habitat fragmentation and its lasting impact on Earth’s ecosystems // Sci. Adv. V. 1. № 2. P. e1500052.
Hall D.M., Camilo G.R., Tonietto R.K., Ollerton J., Ahrne K. et al., 2017. The city as a refuge for insect pollinators // Conserv. Biol. V. 31. № 1. P. 24‒29.
Heiling A.M., 1999. Why do nocturnal orb-web spiders (Araneidae) search for light? // Behav. Ecol. Sociobiol. V. 46. № 1. P. 43‒49.
Hendry A.P., Farrugia T.J., Kinnison M.T., 2008. Human influences on rates of phenotypic change in wild animal populations // Mol. Ecol. V. 17. № 1. P. 20‒29.
Hinde R.A., Fisher J., 1951. Further observations on the opening of milk bottles by birds // British Birds. V. 44. P. 393–396.
Johnson M.T.J., Munshi-South J., 2017. Evolution of life in urban environments // Science. V. 358. № 6363. P. eaam8327.
Kelcey J.G., Rheinwald G., 2005. Birds in European Cities. St. Katharinen: Ginster Verlag. 450 p.
Kettlewell H.B.D., 1955. Selection experiments on industrial melanism in the Lepidoptera // Heredity. V. 9. № 3. P. 323‒342.
King T., Green S., 1995. Factors affecting the distribution of pavement ants (Hymenoptera: Formicidae) in Atlantic Coast urban fields // Entomol. News. V. 106. № 5. P. 224‒228.
Kowarik I., 2020. Herbert Sukopp ‒ an inspiring pioneer in the field of urban ecology // Urban Ecosyst. V. 23. № 3. P. 445‒455.
Kühn I., Brandl R., Klotz S., 2004. The flora of german cities is naturally species rich // Evol. Ecol. Res. V. 6. P. 749‒764.
Lahr E.C., Dunn R.R., Frank S.D., 2018. Getting ahead of the curve: Cities as surrogates for global change // Proc. Biol. Sci. V. 285. № 1882. P. 20180643.
Lampe U., Reinhold K., Schmoll T., 2014. How grasshoppers respond to road noise: Developmental plasticity and population differentiation in acoustic signalling // Funct. Ecol. V. 28. № 3. P. 660‒668.
Le Gros A., Samadi S., Zuccon D., Cornette R., Braun M.P. et al., 2016. Rapid morphological changes, admixture and invasive success in populations of Ring-necked parakeets (Psittacula krameri) established in Europe // Biol. Invasions. V. 18. № 6. P. 1581‒1598.
Lippe M., von der, Saumel I., Kowarik I., 2005. Cities as drivers for biological invasions-the role of urban climate and traffic // Die Erde. V. 136. № 2. P. 123‒143.
Lourenço A., Álvarez D., Wang I.J., Velo-Antón G., 2017. Trapped within the city: Integrating demography, time since isolation and population specific traits to assess the genetic effects of urbanization // Mol. Ecol. V. 26. № 6. P. 1498‒1514.
Luniak M., 2004. Synurbization ‒ adaptation of animal wildlife to urban development // 4th Int. Urban Wildlife Symp. Tucson: Univ. Arizona. P. 50‒55.
Luniak M., Mulsow R., Walasz K., 1990. Urbanization of the European Blackbird expansion and adaptations of urban population // Urban Ecological Studies in Central and Eastern Europe / Ed. Luniak M. Wroclaw: Ossolineum. P. 87‒199.
McKinney M.L., 2006. Urbanization as a major cause of biotic homogenization // Biol. Conserv. V. 127. № 3. P. 247‒260.
Minias P., Janiszewski T., 2016. Territory selection in the city: Can birds reliably judge territory quality in a novel urban environment? // J. Zool. V. 300. № 2. P. 120‒126.
Müller N., Werner P., 2010. Urban biodiversity and the case for implementing the convention on biological diversity in towns and cities // Urban Biodiversity and Design / Eds Müller N., Werner P., Kelcey J.G. Oxford: Wiley-Blackwell. P. 3‒34.
Munshi-South J., Zak Y., Pehek E., 2013. Conservation genetics of extremely isolated urban populations of the northern dusky salamander (Desmognathus fuscus) in New York City // PeerJ. V. 1. P. e64.
Munshi-South J., Zolnik C.P., Harris S.E., 2016. Population genomics of the Anthropocene: Urbanization is negatively associated with genome-wide variation in white-footed mouse populations // Evol. Appl. V. 9. № 4. P. 546‒564.
Neil K., Wu J., 2006. Effects of urbanization on plant flowering phenology: A review // Urban Ecosyst. V. 9. № 3. P. 243‒257.
Nemeth E., Pieretti N., Zollinger S.A., Geberzahn N., Partecke J. et al., 2013. Bird song and anthropogenic noise: Vocal constraints may explain why birds sing higher-frequency songs in cities // Proc. Biol. Sci. V. 280. № 1754. P. 20122798.
Niemela J., 1999. Ecology and urban planning // Biodivers. Conserv. V. 8. № 1. P. 119‒131.
Nihey Y., 1995. Variations of behaviour of carrion crows Corvus corone using automobiles as nutcrackers // Jap. J. Ornithol. V. 44. № 1. P. 21‒35.
Noël S., Ouellet M., Galois P., Lapointe F.-J., 2007. Impact of urban fragmentation on the genetic structure of the eastern red-backed salamander // Conserv. Genet. V. 8. № 3. P. 599‒606.
Nuorteva P., 1971. The synanthropy of birds as an expression of the ecological cycle disorder caused by urbanization // Ann. Zool. Fennici. № 8. P. 547‒553.
O’Brien K., 2012. Global environmental change II: From adaptation to deliberate transformation // Prog. Hum. Geogr. V. 36. P. 667‒676.
Olden J.D., Rooney T.P., 2006. On defining and quantifying biotic homogenization // Glob. Ecol. Biogeogr. V. 15. № 2. P. 113‒120.
Parris K.M., Velik-Lord M., North J.M.A., 2009. Frogs call at a higher pitch in traffic noise // Ecol. Soc. V. 14. № 1. P. 24.
Pickett S.T.A., Cadenasso M.L., Grove J.M., Boone C.G., Groffman P.M. et al., 2011. Urban ecological systems: Scientific foundations and a decade of progress // J. Environ. Manag. V. 92. № 3. P. 331‒362.
Pyšek P., 1993. Factors affecting the diversity of flora and vegetation in central European settlements // Vegetatio. V. 106. № 1. P. 89‒100.
Pyšek P., 1998. Is there a taxonomic pattern to plant invasions? // Oikos. V. 82. № 2. P. 282‒294.
Ravinet M., Elgvin T.O., Trier C., Aliabadian M., Gavrilov A., Sætre G.-P., 2018. Signatures of human-commensalism in the house sparrow genome // Proc. Roy. Soc. B. V. 285. № 1884. P. 20181246.
Reid N.M., Proestou D.A., Clark B.W., Warren W.C., Colbourne J.K. et al., 2016. The genomic landscape of rapid repeated evolutionary adaptation to toxic pollution in wild fish // Science. V. 354. № 6317. P. 1305‒1308.
Rivkin L., Santangelo J., Alberti M., Aronson M., Keyzer C., de, et al., 2019a. A roadmap for urban evolutionary ecology // Evol. Appl. V. 12. № 3. P. 384‒398.
Rivkin L.R., Santangelo J.S., Alberti M., Aronson M.F.J., Keyzer C.W., de, et al., 2019b. A roadmap for urban evolutionary ecology // Evol. Appl. V. 12. № 3. P. 384‒398.
Roetzer T., Wittenzeller M., Haeckel H., Nekovar J., 2000. Phenology in central Europe – differences and trends of spring phenophases in urban and rural areas // Int. J. Biometeorol. V. 44. № 2. P. 60‒66.
Scales J., Hyman J., Hughes M., 2011. Behavioral syndromes break down in urban song sparrow populations // Ethology. V. 117. № 10. P. 887‒895.
Schilthuizen M., 2018. Darwin Comes to Tows. How the Urban Jungle Drives Evolution. L.: Quercus Edition Ltd. 344 p.
Slabbekoorn H., Peet M., 2003. Ecology: Birds sing at a higher pitch in urban noise ‒ Great tits hit the high notes to ensure that their mating calls are heard above the city’s din // Nature. V. 424. № 6946. P. 267.
Strubbe D., Matthysen E., 2007. Invasive ring-necked parakeets Psittacula krameri in Belgium: Habitat selection and impact on native birds // Ecography. V. 30. № 4. P. 578‒588.
Suárez-Rodríguez M., López-Rull I., Macias Garcia C., 2013. Incorporation of cigarette butts into nests reduces nest ectoparasite load in urban birds: New ingredients for an old recipe? // Biol. Lett. V. 9. № 1. P. 20120931.
Sukopp H., 2003. Flora and vegetation reflecting the urban history of Berlin // Die Erde. V. 134. № 3. P. 295‒316.
Surov A.V., Zaytseva E.A., Kuptsov A.V., Katzman E.A., Bogomolov P.L. et al., 2019. Circle of life: The common hamster (Cricetus cricetus) adaptations to the urban environment // Integr. Zool. V. 14. № 4. P. 383‒395.
Symonds M.R., Weston M.A., Dongen W.F., van, Lill A., Robinson R.W., Guay P.-J., 2016. Time since urbanization but not encephalisation is associated with increased tolerance of human proximity in birds // Front. Ecol. Evol. V. 4. https://doi.org/10.3389/fevo.2016.00117
Thompson K.A., Rieseberg L.H., Schluter D., 2018. Speciation and the City // Trends Ecol. Evol. V. 33. № 11. P. 815‒826.
Tutt J.W., 1896. British Moths. L.: G. Routledge and Sons. 368 p.
United Nations World Urbanization Prospects: The 2018 Revision, 2018. https://www.un.org/en/desa/products/publications
Van’t Hof A.E., Campagne P., Rigden D.J., Yung C.J., Lingley J. et al., 2016. The industrial melanism mutation in British peppered moths is a transposable element // Nature. V. 534. № 7605. P. 102‒105.
Walbridge M., 1997. Urban ecosystems // Urban Ecosyst. V. 1. № 1. P. 1‒2.
Whitehead A., Galvez F., Zhang S.J., Williams L.M., Oleksiak M.F., 2011. Functional genomics of physiological plasticity and local adaptation in killifish // J. Heredity. V. 102. № 5. P. 499‒511.
Wirgin I., Roy N.K., Loftus M., Chambers R.C., Franks D.G., Hahn M.E., 2011. Mechanistic basis of resistance to PCBs and dioxin in Atlantic tomcod from the Hudson River // Science. V. 331. № 6022. P. 1322‒1325.
Youngsteadt E., Dale A.G., Terando A.J., Dunn R.R., Frank S.D., 2015. Do cities simulate climate change? A comparison of herbivore response to urban and global warming // Glob. Change Biol. V. 21. № 1. P. 97‒105.
Ziska L.H., Bunce J.A., Goins E.W., 2004. Characterization of an urban-rural CO2/temperature gradient and associated changes in initial plant productivity during secondary succession // Oecologia. V. 139. № 3. P. 454‒458.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии