

Журнал общей биологии, 2023, T. 84, № 3, стр. 229-240
Сезонная динамика роста побегов у растений Forsythia ovata Nakai: ритмичность апикального и радиального роста
С. А. Шавнин 1, *, А. А. Монтиле 1, Л. А. Семкина 1, А. И. Монтиле 1
1 Ботанический сад Уральского отделения РАН
620144 Екатеринбург, ул. 8 Марта, 202а, Россия
* E-mail: sash@botgard.uran.ru
Поступила в редакцию 12.10.2022
После доработки 13.02.2023
Принята к публикации 26.02.2023
- EDN: RALCMZ
- DOI: 10.31857/S0044459623020057
Аннотация
Целью исследований являлось установить характер (линейный или нелинейный) и форму (наличие экстремумов и трендов) зависимостей от времени скоростей апикального и радиального видов роста побегов двух типов (ветвления и формирования) у древесных растений на примере интродуцированного на Среднем Урале декоративного раннецветущего вида кустарников – форзиции овальной (Forsythia ovata Nakai). Проведено изучение сезонных динамик морфометрических показателей отличающихся по интенсивности роста и происхождению (вырастающих из верхушечных и спящих почек скелетной ветви) побегов, а также сделана оценка их связи с температурой воздуха и количеством осадков. Оба вида роста сопровождаются колебаниями величин приростов, продолжительность отдельных фаз которых составляет около 7 и более дней, что позволяет отнести данное явление к инфрадианным ритмам роста. Слабая связь характеристик наблюдаемых колебаний с погодными условиями указывает на преимущественно эндогенную природу определяющих их морфогенетических процессов. Рост побегов формирования начинается на 4 нед. позже, чем побегов ветвления, что связано, по-видимому, с временем активации роста спящей почки. Сезонные динамики величин скоростей и ускорений апикального и радиального роста побегов имеют вид не наблюдавшихся ранее, затухающих по амплитуде и изменяющихся по продолжительности фаз колебаний, завершающихся за 1–2 мес. до наступления неблагоприятных для роста условий среды. Вызывающие изменения скорости роста побегов процессы связаны, по-видимому, с действием двух или более разнонаправленных факторов, определяющих особенности и темпоральные обусловленности увеличения объема в различных сегментах побега (в том числе различный вклад деления и растяжения клеток в изменение геометрических размеров доменов апикальной меристемы). У обоих типов побегов существуют отличия сезонных динамик величин апикального и радиального приростов по продолжительности роста (6–8 и 4 нед. соответственно), количеству экстремумов, амплитуде и длительности отдельных фаз колебаний. Кривые динамик скоростей апикального роста имеют два максимума, а радиального – один. Каждый из видов роста, несмотря на задержку его начала у побегов формирования, имеет у разных типов побегов одинаковую общую продолжительность, а формы кривых сезонных изменений величин скоростей и ускорений у разных типов побегов аналогичны. Сходство форм кривых сезонных динамик отдельных видов роста у разных типов побегов указывает на наличие у побегов одинаковых, обусловленных в конечном счете генотипом, “программ” регуляции роста, проявляющихся в наблюдаемых колебаниях. Ритмичность роста побегов обоих типов в течение сезона позволяет предполагать существование взаимосвязи во времени скоростей деления и роста отдельных клеток, находящихся в функционально отличающихся группах клеток зоны апекса. Отличия динамик у разных типов побегов заключаются в том, что у побегов формирования амплитуды колебаний выше, чем у побегов ветвления. Эта особенность объясняет известный факт более высоких величин их суммарных приростов.
Изучение биологических ритмов относится к числу актуальных проблем биологии развития. Одним из ее важных аспектов является исследование периодических процессов роста древесных растений и их связи с образованием разных жизненных форм и ходом морфогенеза (Серебряков, 1962; Сабинин, 1963; Михалевская, 2008). Регуляция этих процессов имеет преимущественно эндогенную природу, включая генетический контроль и гормональный статус, однако на их ход влияют также факторы среды, в том числе отмечаемые в ряде обзорных статей продолжительность светового дня и температура (Luttge, Hertel, 2009; Cooke et al., 2012; Singh et al., 2017, 2020). Наибольшее количество исследований в этой области связано с изучением молекулярно-генетических и биохимических механизмов, а также моделированием ультрадианных (период колебаний менее 24 ч) и циркадных ритмов травянистых растений (McClung, 2006; Farre, 2012; Kim et al., 2017; Edwards et al., 2018; Mahmud et al., 2018, и др.). Проявления инфрадианных ритмов (период колебаний более 24 ч) и механизмов их возникновения изучены в меньшей степени. В годичном цикле древесных растений также наблюдается несколько ритмов, появление которых обусловлено действием в первую очередь эндогенных факторов на фоне влияния внешних условий среды, включая их сезонные изменения (Серебряков, 1952, 1962, 1966; Сабинин, 1963; Luttge, Hertel, 2009; Herrmann et al., 2015; Singh et al., 2017). Указания на ритмичность роста древесных растений связаны, как правило, с наличием внешних морфологических признаков изменений скорости роста на поверхности побега (расположение почечного кольца и зон междоузлий), позволяющих определить количество циклов роста и элементарных побегов (Михалевская, 2008). В этой обзорной работе обсуждаются полученные ранее данные о полимодальности кривых распределения элементарных побегов по числу метамеров (Михалевская, 1987), свидетельствующей о квантированности процесса роста у растений и объясняемой периодичностью активности апикальной меристемы на примере трех видов дуба. Кроме того, в статье указывается, что зависимость скорости видимого роста побега обычно имеет вид одновершинной кривой. Других примеров прямых измерений скоростей роста побегов в течение сезона вегетации в данной работе не приводится. Следует отметить, что наличие периодичности в сезонной динамике апикального роста свидетельствует о существовании инфрадианных ритмов изменения скорости роста побега.
Количественный подход к изучению проблемы морфогенеза растений является основой количественной морфогенетики (Магомедмирзаев, 1990) – научного направления, связанного с выявлением закономерностей морфогенеза биологических объектов с помощью математико-статистического анализа результатов, полученных путем прямых измерений. Его применение дополняет традиционные морфологические методы при решении задач, связанных с изучением проявлений действия генетической программы развития при формировании архитектоники растения в виде изменений морфологических признаков во времени. В рамках количественного подхода изучению сезонной динамики скоростей роста древесных растений до настоящего времени уделялось недостаточное внимание. В частности, несмотря на относительно хорошо изученные механизмы генетического контроля роста и анатомо-цитологическую организацию апикальной меристемы побега, конкретные связи определяющих рост побега процессов с его внешними проявлениями во времени установлены на уровне моделей только для циркадных и ультрадианных ритмов.
Метод количественного анализа морфогенеза был использован при исследовании апикального роста побегов ивы трехтычинковой (Salix triandra) (Афонин, 2019a, в, 2021) и ивы корзиночной (S. viminalis) (Афонин, 2019б). С его помощью на основе гармонического анализа данных была выявлена цикличность сезонной динамики апикальных приростов побегов, которая объясняется взаимодействием инфрадианных ритмов трех уровней. Соответствующие периоды колебаний составляют 9–18, 21–24 и 29–36 сут. На основании полученных результатов сделано заключение, что “цикличность сезонной динамики суточного прироста определяется синхронизаторами неизвестной природы”, а также высказано предположение об определяющей роли синхронизатора при “завершении развития ранних (преформированных) метамеров и начале развития поздних (примордиальных) метамеров” (Афонин, 2021, с. 4). В число предполагаемых синхронизаторов входят температурный и фотопериодический факторы. К сожалению, автором этого цикла работ возможная природа цикличности роста побегов на тканевом уровне не рассматривалась. Для получения более полного представления о механизмах инфрадианных колебаний, наблюдаемых при развитии апикальной меристемы, представляется важным определить, встречаются ли они у других видов растений (аналогичных работ найти не удалось). Кроме того, для дополнения феноменологии явления с целью углубления интерпретации данных необходимо изучить сезонную динамику радиального роста, так как вопрос о его взаимосвязи с апикальным до настоящего времени слабо изучен (Ding et al., 2021).
Перспективными объектами для изучения проявлений ритмичности роста и развития древесных растений, наряду с быстрорастущими ивами, являются виды, у которых активный рост побегов, несмотря на благоприятные условия среды, заканчивается уже в середине сезона вегетации. К их числу относится кустарник форзиция овальная (Forsythia ovata Nakai) (Семкина, Овсянникова, 1998). Раннее цветение и короткий период роста выделяет данный вид среди других при изучении биоритмов. При омоложении кустов древесных растений обрезкой скелетных ветвей из расположенных у основания ветвей спящих почек вырастают побеги формирования (ПФ) (Мазуренко, Хохряков, 1977). ПФ отличаются от побегов ветвления (ПВ) значительно большей величиной годичных апикальных и радиальных приростов, а у многих видов – размерами и количеством листьев. Эта особенность связана с изменениями баланса концентраций ростовых гормонов (в первую очередь ауксинов и цитокининов) в частях стебля, расположенных на разном удалении от апекса (Aloni, 2007; Лутова и др., 2010; Творогова и др., 2012; Медведев, Шарова, 2014; Кулуев, Сафиуллина, 2015; Кулуев, 2017). Данное объяснение основано на преобладающей роли ауксинов в регуляции роста ПВ, а цитокининов – в пробуждении спящих почек после удаления скелетных ветвей и регуляции роста вырастающих из этих почек ПФ. В связи с этим сравнительный анализ морфологических проявлений апикального и радиального роста у разных типов побегов представляется перспективным для выявления сходства и различия между функционированием соответствующих регуляторных систем и выяснения механизмов их работы.
На основании анализа приведенных выше сведений нами были сформулированы следующие рабочие гипотезы: 1) у ПФ и ПВ форзиции овальной существуют инфрадианные ритмы апикального и радиального роста; 2) эти ритмы отличаются у ПВ и ПФ; 3) на ритмы апикального и радиального роста отдельных типов побегов, наряду с эндогенными факторами, оказывают влияние температура воздуха и количество осадков. В связи с проверкой выдвинутых гипотез целью исследований являлось установить характер (линейный или нелинейный) и форму (наличие экстремумов и трендов) сезонных зависимостей от времени скоростей апикального и радиального видов роста побегов двух типов (ветвления и формирования) у древесных растений на примере интродуцированного на Среднем Урале раннецветущего вида – форзиции овальной (Forsythia ovata Nakai). В число основных задач работы входило: 1) определение сезонных динамик скоростей и ускорений апикального и радиального роста отличающихся по интенсивности роста и происхождению побегов ветвления и формирования (вырастающих из верхушечных и спящих почек скелетной ветви соответственно); 2) сравнительный анализ характера и формы сезонных динамик морфометрических показателей роста побегов ветвления и формирования; 3) проведение оценки связи изучаемых морфометрических показателей роста побегов с температурой воздуха и количеством осадков.
МАТЕРИАЛЫ И МЕТОДЫ
Объектом исследований являлась форзиция овальная (Forsythia ovata Nakai), жизненная форма которой – кустарник, обладающий высокой декоративностью благодаря раннему обильному цветению и раскидистой форме кроны, достигающей 2 м и более в диаметре. Ее происхождение – влажные субтропики (Юго-Восточная Азия, Корея). Несмотря на подмерзание верхней части побегов, этот вид интродуцирован и успешно размножается в коллекции Ботанического сада Уральского отделения РАН (г. Екатеринбург, РФ) с 1962 г. Территория сада относится к южно-таежной подзоне Среднеуральской низкогорной провинции Уральской горно-лесной области (Колесников и др., 1974). В результате селекции была получена устойчивая к полному вымерзанию форма. Растения зацветают во второй декаде апреля и продолжают цвести в течение 3–4 нед. На ПВ и ПФ прошлого года распускаются одиночные (иногда 2–3 штуки) ярко-желтые цветки диаметром около 3 см. На последней неделе цветения из верхушечной почки, находящейся между двумя цветочными, развивается один ПВ первого порядка, интенсивный рост которого длится около 5 нед. Цветочные почки имеют заостренно-яйцевидную форму с плотно прилегающими темно-коричневыми чешуями. Ростовые почки небольшие, конусовидные, покрытые светло-коричневыми чешуями. К началу июля ПВ достигают длины 12–20 см и почти прекращают свой рост, закладывая цветочные и ростовые почки будущего года. На однолетних ПВ развивается 5–7 ярусов листьев, размеры которых незначительно изменяются в течение всей вегетации. ПФ образуются на нижней части скелетной ветви после гибели или удаления ее верхней части. За вегетационный период они могут достигать длины 60–170 см. На следующий год из почек в верхней части туриона развиваются ПВ. Растение легко переносит омоложение обрезкой скелетных ветвей, образующиеся при этом ПФ не изменяют общее состояние куста, и их появление не является результатом раневого роста. Созревание семян происходит в середине октября. Кусты достигают высоты 1.5–2 м и состоят из 5–7 скелетных ветвей. Основной рост куста осуществляется за счет ПВ и появления новых скелетных ветвей, формирующихся из спящих или адвентивных почек. Для наблюдений использовали 6 произрастающих в коллекции кустов. Возраст растений составляет около 15 лет.
В 2019 г. у каждого из исследуемых растений на высоте 100–150 см от поверхности были выбраны 16 ПВ, растущих из терминальных почек отдельных скелетных ветвей (2–3 ПВ на кусте). При этом на каждом кусте примерно за 1 мес. до начала цветения была произведена выборочная обрезка 1–2 скелетных ветвей под пень для получения ПФ из спящих почек, расположенных на высоте 3–5 см от поверхности почвы. После начала роста ПФ из них были выбраны 15 экземпляров для исследований (2–4 побега на кусте). У всех побегов определяли высоту (общая длина побега) и диаметр у основания побега. Измерения проводили у ПВ и ПФ с интервалом 6–10 дней в течение вегетационного периода – с начала роста ПВ в мае до начала сентября. Величина интервала была установлена с учетом предварительно определенного периода роста побегов (7–8 нед.). Длину побега измеряли от его основания с момента начала роста, а диаметр – у его базальной части с точностью 1 и 0.01 мм соответственно.
Обработку данных проводили в пакетах Excel (Microsoft, 2007) и Statistica 8.0 (StatSoft Inc., 2007). Для построения графиков сезонных динамик средних приростов (апикальных и по радиусу) побегов форзиции данные за каждую дату измерения усредняли для ПВ и ПФ по отдельности и рассчитывали стандартные ошибки. Ускорения изменений приростов рассчитывали как разности между приростами за две смежные даты измерений, деленные на промежуток времени между ними по формуле ΔV/Δt, где V – величина прироста, ΔV = Vt + 1 – Vt, Δt – время между измерениями, которое постоянно и равно одной неделе. На рисунках представлены стандартные сглаживающие кривые графиков, получаемые в пакете Excel. Ввиду того, что период измерений составлял одну неделю, а также с целью облегчения восприятия текста, на рисунках и в тексте приведены номера недель, начиная от начала измерений. Для выявления сезонных трендов роста отдельных побегов строилась аппроксимация ростовых кривых участком S-образной функции, имеющей вид
Для установления влияния погодных условий вегетационного периода на приросты применяли однофакторный дисперсионный анализ ANOVA, в котором фактором выступала либо температура, либо уровень осадков, эффект фактора принимался достоверным при уровне значимости p < 0.05. При выполнении анализа значения средних температур и осадков за неделю, предшествующую дате измерения приростов, были разбиты на уровни (градации фактора) с интервалом изменения 2.5°C для температуры и 2 мм для осадков. Реакцию на изменения уровня температурного фактора оценивали на основании анализа величин отклонений приростов от соответствующих значений на кривой сезонного роста (анализ на основе непосредственных величин приростов влияния температуры не выявил). Для каждого побега эту кривую строили посредством аппроксимации данных о его сезонном росте. При дисперсионном анализе влияния уровня осадков использовали непосредственно величины приростов.
РЕЗУЛЬТАТЫ
Рост ПВ начался в последнюю неделю цветения и продолжался около 5 нед. Рост ПФ начался на этапе завершения активного роста ПВ и продолжался также около 5 нед. Апикальный рост ПВ начался в период 16.05.19–23.05.19. Динамика зависимости приростов от времени (рис. 1) свидетельствует о быстром увеличении скорости роста в первую неделю, которая достигает максимальных за сезон значений. На второй неделе этот показатель снижается, а затем восстанавливается на третьей неделе.
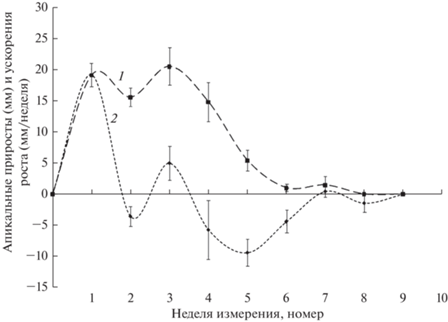
В течение последующих 3 нед. происходит последовательное уменьшение скорости апикального роста ПВ до его полной остановки на 6–7-й неделе. Таким образом, сезонная динамика в целом имеет нелинейный характер. В ней наблюдается два максимума, а общая продолжительность интенсивного апикального роста ПВ составляет около 6 нед.
При дальнейшем анализе данных о зависимости скоростей апикального роста ПВ от времени с целью непосредственной оценки изменения скоростей роста были рассчитаны соответствующие значения средних ускорений роста. Анализ полученных результатов (рис. 1) показал, что ускорения изменяются волнообразно и их динамика во времени имеет вид уменьшающихся по амплитуде и изменяющихся по периоду колебаний, полностью затухающих на седьмой неделе. Первое полное колебание завершается через 2.5, а второе – через 4.5 нед. Таким образом, сопоставление данных о временных динамиках скоростей и ускорений апикального роста ПВ свидетельствует о колебательном характере данного процесса, который продолжается в течение 6–7 нед.
Радиальный рост ПВ начался позже апикального – на второй неделе проведения наблюдений (рис. 2). Его скорость сразу достигла максимальных значений, в последующие 3 нед. радиальные приросты постепенно уменьшаются до минимума, далее сохраняются на этом уровне еще 2 нед., после чего рост прекращается. Таким образом, зависимость радиальных приростов от времени имеет вид одновершинной кривой, общая продолжительность радиального роста ПВ составляет около 7 нед., а его наиболее интенсивной стадии – 4 нед.
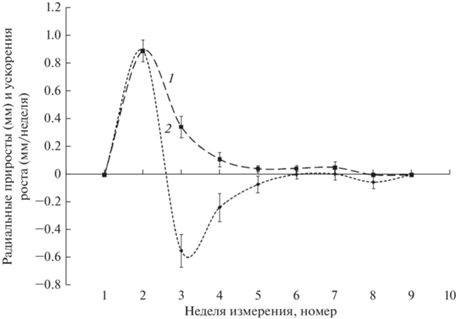
При анализе ускорений радиального роста ПВ использовали подход, примененный ранее для апикального роста. Анализ полученных результатов (рис. 2) показал, что ускорения, как и в случае апикального роста ПВ, изменяются волнообразно. При этом наблюдается только одна волна, у которой с течением времени уменьшаются амплитуды и изменяются длительности отдельных последовательных фаз колебаний. Полное прекращение колебаний происходит на шестой неделе наблюдений. Сопоставление данных о динамике во времени скоростей и ускорений радиального роста ПВ позволяет заключить, что этот процесс, как и апикальный рост, имеет колебательный характер. Интенсивный рост продолжается около 4 нед., а общая продолжительность роста составляет 6–7 нед.
Апикальный рост ПФ начался на четвертой неделе периода наблюдений (29.05.19–06.06.19). В течение этой недели достигается максимальная скорость роста (рис. 3). На пятой неделе наблюдений приросты уменьшаются более чем на 80%, после чего на 2 нед. вновь увеличиваются (примерно до 40% от максимальных значений), а затем постепенно снижаются на протяжении следующих 5 нед. до полной остановки на 12–13-й неделе наблюдений. Следует отметить, что апикальный рост ПФ начинается на 4 нед. позже, чем у ПВ, а величины его максимальных приростов выше примерно в 15 раз. В целом сезонная динамика приростов характеризуется наличием двух максимумов, а общая продолжительность апикального роста ПФ составляет около 9 нед.
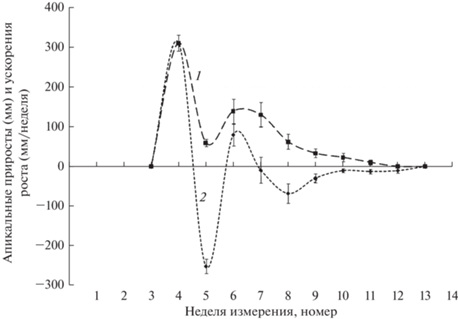
Анализ величин ускорений апикального роста ПФ (рис. 3) показал, что кривая сезонных изменений этого показателя, как и описанных ранее, имеет вид уменьшающихся по амплитуде и изменяющихся по продолжительности отдельных фаз колебаний. Следует отметить, что продолжительности фаз изменяются в меньшей степени, чем у ростовых характеристик ПВ, и составляют 0.5–1 нед. Изменения ускорения прекращаются на 10–11-й неделе наблюдений. Первая полная волна длится около 2.5, а вторая – 4.5 нед. Процесс роста длится 6–7 нед.
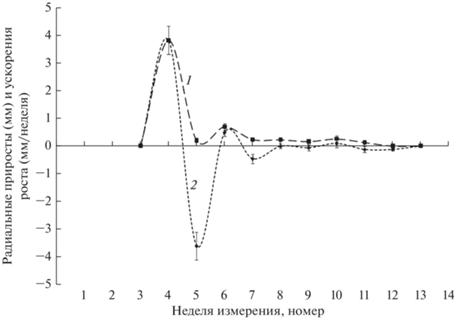
Зависимость радиальных приростов от времени имеет вид двухвершинной кривой, на которой первый максимум значительно выше второго (рис. 4). Общая продолжительность радиального роста ПФ составляет около 8 нед., а активного роста – 4 нед. Анализ результатов расчета ускорений радиального роста ПФ наглядно свидетельствует о волнообразном характере изменений этого показателя. В его сезонной динамике наблюдается примерно 1.5 волны, у которых с течением времени уменьшаются амплитуды и остаются относительно неизменными длительности отдельных фаз колебаний. Полное прекращение роста произошло через 5 нед. после начала радиального роста. Сопоставление данных о сезонной динамике скоростей и ускорений радиального роста ПВ и ПФ позволяет заключить, что этот процесс представляет собой затухающее колебание.
Важной для описания инфрадианных ритмов апикального и радиального роста побегов особенностью является более высокая согласованность (синхронность) фаз колебаний у разных ПФ по сравнению с ПВ. Этот факт продемонстрирован на примере сезонных динамик ускорений апикального роста отдельных изучавшихся ПФ и ПВ (рис. 5). Аналогичная закономерность наблюдается также у остальных исследованных характеристик.
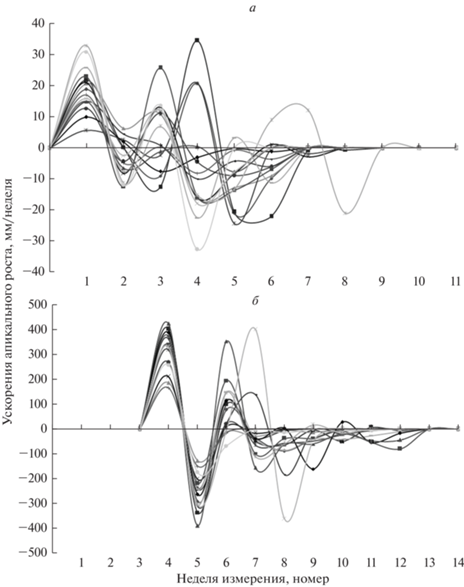
Рост побегов связывают с температурой среды и фотопериодической регуляцией (Jackson, 2009; Triozzi et al., 2018; Miskolczi et al., 2019). Задержка начала роста ПФ по сравнению с ПВ составляет 4–5 нед. и свидетельствует о задержке процесса активации спящих почек по сравнению с терминальными у ПВ. Этот эффект является одним из проявлений “коррелятивной стимуляции”, заключающейся в активации роста побега растущим корнем, обеспечивающей “сбалансированность в развитии надземной и подземной частей растения” (Медведев, Шарова, 2014, с. 138).
Анализ полученных результатов позволяет предполагать, что нелинейный характер процессов роста связан с действием эндогенных и/или экзогенных (погодных) факторов. С целью оценки влияния факторов среды (сведения приведены в табл. 1) на изучаемые характеристики роста ПВ и ПФ были проведены корреляционный и дисперсионный ANOVA анализы. Корреляционный анализ не выявил статистически значимых (на уровне p = 0.05) корреляций приростов ПФ и ПВ с температурой воздуха и количеством осадков. Результаты дисперсионного анализа свидетельствуют о том, что влияние температурного фактора на отклонения апикальных приростов от сезонного тренда достоверно как для ПФ, так и для ПВ (табл. 2).
Таблица 1.
Величины средних за предшествующую неделю температур и количества осадков в период проведения исследований (по данным метеостанции г. Екатеринбурга (Булыгина и др., 2020))
| Номера недель и даты измерений | Температура, °C | Осадки, мм | |
|---|---|---|---|
| 1 | 16.05.2019 | 16.6 | 0.5 |
| 2 | 23.05.2019 | 7.0 | 3.3 |
| 3 | 29.05.2019 | 13.9 | 0.8 |
| 4 | 06.06.2019 | 15.6 | 2.6 |
| 5 | 14.06.2019 | 17.1 | 0.5 |
| 6 | 19.06.2019 | 13.8 | 0.9 |
| 7 | 28.06.2019 | 17.2 | 3.0 |
| 8 | 05.07.2019 | 18.1 | 3.5 |
| 9 | 12.07.2019 | 17.9 | 1.0 |
| 10 | 18.07.2019 | 22.8 | 0.1 |
| 11 | 25.07.2019 | 21.4 | 10.5 |
| 12 | 31.07.2019 | 16.2 | 5.7 |
| 13 | 09.08.2019 | 13.6 | 3.1 |
| 14 | 15.08.2019 | 18.6 | 0.4 |
| 15 | 20.08.2019 | 18.9 | 1.4 |
| 16 | 26.08.2019 | 13.2 | 4.6 |
| 17 | 05.09.2019 | 9.5 | 3.6 |
Таблица 2.
Результаты дисперсионного анализа влияния температуры воздуха и количества осадков на апикальный и радиальный рост ПФ и ПВ
| Фактор, вид роста | Оцениваемый параметр | Число степеней свободы | SS | Средняя дисперсия | F-критерий | p-значение |
|---|---|---|---|---|---|---|
| Температура, апикальный рост | ПФ | |||||
| Влияние фактора | 3 | 102 736.0 | 34 245.34 | 27.255 | 0.0000 | |
| Случайная ошибка | 81 | 101 775.5 | 1256.49 | |||
| ПВ | ||||||
| Влияние фактора | 3 | 203.941 | 67.980 | 2.8585 | 0.0413 | |
| Случайная ошибка | 91 | 2164.178 | 23.782 | |||
| Температура, радиальный рост | ПФ | |||||
| Влияние фактора | 3 | 2.339 | 0.7795 | 4.4683 | 0.0058 | |
| Случайная ошибка | 85 | 14.828 | 0.174 | |||
| ПВ | ||||||
| Влияние фактора | 2 | 0.011 | 0.011 | 0.5231 | 0.4714 | |
| Случайная ошибка | 91 | 1.903 | 0.0209 | |||
| Уровень осадков, апикальный рост | ПФ | |||||
| Влияние фактора | 1 | 241 138 | 241 138 | 19.7919 | 0.000025 | |
| Случайная ошибка | 87 | 1 059 979 | 12 184 | |||
| ПВ | ||||||
| Влияние фактора | 1 | 330.66 | 330.66 | 3.0998 | 0.0821 | |
| Случайная ошибка | 81 | 8640.35 | 106.67 | |||
| Уровень осадков, радиальный рост | ПФ | |||||
| Влияние фактора | 2 | 11.208 | 5.604 | 2.7323 | 0.0709 | |
| Случайная ошибка | 84 | 172.280 | 2.051 | |||
| ПВ | ||||||
| Влияние фактора | 1 | 0.330 | 0.330 | 2.0645 | 0.1561 | |
| Случайная ошибка | 58 | 9.272 | 0.1599 | |||
Изучение действия температурного фактора на радиальные приросты показало, что эффект влияния температуры статистически значим для ПФ и не достоверен для ПВ. Результаты, полученные с помощью ANOVA по фактору осадков для приростов ПФ и ПВ (табл. 2), свидетельствуют о том, что его действие на апикальные приросты статистически значимо только у ПФ, а действие на радиальные приросты ПВ и ПФ – недостоверно.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Анализ совокупности полученных данных свидетельствует о наличии у форзиции овальной инфрадианных ритмов роста ПФ и ПВ (колебаний скоростей и ускорений как апикального, так и радиального роста). В отличие от относительно устойчивых инфрадианных ритмов апикального роста побегов ив, наблюдаемых в течение всего периода вегетации (Афонин, 2021), изменения характеристик апикального роста ПВ и ПФ у форзиции овальной происходят в течение более короткого интервала времени и имеют аналогичный по форме вид затухающих по амплитуде колебаний. По-видимому, указанные отличия сезонных динамик роста между данными видами связаны с особенностями их развития. У радиального роста ПФ и ПВ сезонные динамики также аналогичны, однако отличаются по форме кривых от динамик апикального роста, что объясняется отличием процессов развития апикального и базального участков апикальной меристемы. При этом динамики скоростей апикального и радиального роста имеют сходство в первые недели роста, а в дальнейшем они отличаются. Аналогичная закономерность (корреляция этих видов роста на начальной стадии) наблюдалась также в динамике количественных характеристик роста деревьев Picea meyery (Ding et al., 2021). Совпадение продолжительностей как апикального, так и радиального роста у ПФ и ПВ позволяет предполагать, что механизмы регуляции наблюдаемых инфрадианных ритмов включают также учет количества прошедшего с начала роста дней.
На ритмичность морфогенеза форзиции овальной указывают следующие факты: 1) наличие последовательной смены фаз развития (цветение–апикальный и радиальный рост ПВ); 2) волнообразность кривых зависимостей величин скоростей и ускорений апикального и радиального роста от времени; 3) затухающий характер и конечность продолжительностей колебаний у ПФ и ПВ; 4) согласованность времен окончания роста ПВ и начала роста ПФ (рост ПФ начинается на стадии завершения роста ПВ); 5) отличие продолжительностей интенсивного апикального и радиального роста побегов (6–8 и 4 нед. соответственно), несмотря на то, что условия среды в конце июля–августе не лимитируют ростовые процессы; 6) одинаковость продолжительностей каждого из видов роста у ПФ и ПВ, в результате чего прекращение интенсивного роста у двух типов побегов происходит в разное время (в июне у ПВ и в июле у ПФ); 7) аналогичность форм кривых сезонных динамик у отдельных видов роста у ПФ и ПВ, при этом формы кривых колебаний апикального и радиального роста отличаются. Следует отметить, что радиальный рост ПВ начинается на одну неделю позже, чем апикальный, а у ПФ эти виды роста начинаются синхронно. Наблюдаемая более высокая синхронность фаз ритмов роста отдельных ПФ по сравнению с ПВ (рис. 5) объясняется, по-видимому, более простой (менее протяженной и разветвленной) системой транспорта гормонов и метаболитов к ростовым меристемам ПФ. Эта система у ПФ не связана с другими побегами и функционирует на основе значительных количеств ростовых гормонов и накопленных ассимилятов в корнях, что обеспечивает меньшую зависимость действия этих регуляторов от дополнительных факторов. Таким образом, гипотезы 1 и 2 подтверждены.
Результаты корреляционного и дисперсионного анализов связей морфометрических характеристик роста побегов с погодными условиями среды указывают на слабое влияние температуры воздуха и почти полное отсутствие влияния количества осадков. Эти данные соответствуют известным представлениям о преимущественно эндогенной природе ритмичности роста побегов и второстепенном влиянии факторов среды, и гипотеза 3 подтверждена частично.
В основе представлений о ритмике роста растений лежат сведения о развитии апикальной меристемы побега, в которой выделяют центральную, подстилающую и периферическую зоны и примордий листа. Клетки этих частей апекса делятся периклинально и антиклинально, проходя последовательно этапы от вхождения в состав организующего центра до дифференцировки, и имеют разные длительности клеточных циклов. Например, клетки покоящегося центра (аналога организующего центра) в апексе корня у кукурузы делятся с периодичностью примерно 200 ч, а клетки проксимальной меристемы – 20 ч (Лутова и др., 2010). При этом регуляция развития апикальной меристемы находится под контролем системы генов и происходит с участием гормонов. В связи с приведенными сведениями, сходный характер сезонных динамик апикального (а также радиального) роста ПФ и ПВ позволяет предположить наличие у них одинаковых механизмов возникновения инфрадианных ритмов, в основе функционирования которых лежат собственная и относительная динамики процессов деления и роста клеток. При этом авторы считают, что на регуляцию ростовых процессов побега непосредственно влияют величины распределенных по объему концентраций фитогормонов, в первую очередь ауксинов и цитокининов, а также количество поступающих в меристемы ассимилятов. Сочетание концентраций различных фитогормонов и ассимилятов в конкретной “точке” обусловливает согласование клеточных циклов в разных зонах апикальной меристемы и ритмичность роста побега в целом. Ранее не описанный в известной нам литературе факт наличия инфрадианных сезонных изменений скоростей апикального роста ПВ и ПФ в форме затухающих псевдогармонических колебаний указывает на то, что они являются результатом взаимодействия как минимум двух факторов, происходящего в ходе вегетационного периода. В отличие от ив, у форзиции этот процесс приводит к завершению роста в течение нескольких недель. Эта особенность и приведенные выше факты ритмичности морфогенеза побегов форзиции овальной позволяют считать перспективной постановку задачи разработки качественной модели динамики роста побега.
ЗАКЛЮЧЕНИЕ (ВЫВОДЫ)
1. В сезонной динамике роста побегов ветвления и побегов формирования растений Forsythia ovata Nakai наблюдаются колебания величин апикального и радиального приростов. Продолжительность отдельных фаз колебаний составляет около 7 и более дней, что позволяет отнести данное явление к инфрадианным ритмам роста. Слабая связь характеристик наблюдаемых колебаний с температурой воздуха и количеством осадков указывает на эндогенный характер определяющих их морфогенетических процессов.
2. На примере форзиции овальной впервые установлено, что сезонные динамики величин скоростей апикального и радиального роста побегов и их изменений (ускорений) могут иметь вид затухающих по амплитуде и изменяющихся по продолжительности фаз колебаний. Эти колебания не относятся к классу автоколебаний, а являются, по-видимому, результатом наложения действий как минимум двух факторов или процессов.
3. Как у побегов ветвления, так и у побегов формирования существуют отличия сезонных динамик величин апикального и радиального приростов по продолжительности роста (6–8 и 4 нед. соответственно) и по форме описывающих их кривых, включая количество экстремумов и величины амплитуд и длительностей отдельных фаз колебаний. Особенности сезонных динамик роста побегов указывают на отличия механизмов, вызывающих колебания величин апикальных и радиальных приростов у изучавшихся двух типов побегов.
4. У отдельных видов роста (апикального и радиального) их продолжительности одинаковы как у побегов ветвления, так и у побегов формирования, а формы кривых сезонных изменений величин скоростей и ускорений аналогичны, что указывает на единство механизма процессов, определяющего соответствующие виды роста у этих типов побегов.
Список литературы
Афонин А.А., 2019a. Ритмичность линейного прироста однолетних побегов ивы трехтычинковой // Соврем. наука: актуальные проблемы теории и практики. Сер. Естеств. и техн. науки. № 1. С. 10–16.
Афонин А.А., 2019б. Сезонная динамика нарастания побегов ивы корзиночной (Salix viminalis) // Изв. высших учебных заведений. Поволж. регион. Естеств. науки. № 4 (28). С. 26–34. https://doi.org/10.21685/2307-9150-2019-4-3
Афонин А.А., 2019в. Структурный анализ ритмов развития однолетних побегов ивы трехтычинковой // Бюлл. науки и практики. Т. 5. № 1. С. 22–32. https://doi.org/10.5281/zenodo.2539541
Афонин А.А., 2021. Инфрадианные ритмы динамики нарастания побегов в клонах ивы трехтычинковой (Salix triandra) // Аграр. вестн. Урала. Т. 205. № 2. С. 2–11. https://doi.org/10.32417/1997-4868-2021-205-02-2-11
Булыгина О.Н., Разуваев В.Н., Александрова Т.М., 2020. Описание массива данных суточной температуры воздуха и количества осадков на метеорологических станциях России и бывшего СССР (TTTR). http://meteo.ru/data/162-temperature-precipitation#описание-массива-данных
Колесников Б.П., Зубарева Р.С., Смолоногов Е.П., 1974. Лесорастительные условия и типы лесов Свердловской области. Свердловск: Изд-во УНЦ АН СССР. 176 с.
Кулуев Б.Р., 2017. Регуляторы деления и пролиферации клеток в растениях // Биомика. Т. 9. № 2. С. 119–135.
Кулуев Б.Р., Сафиуллина М.Г., 2015. Регуляция роста клеток растяжением в растениях // Успехи соврем. биологии. Т. 135. № 2. С. 148–163.
Лутова Л.А., Ежова Т.А., Додуева И.Е., Осипова М.А., 2010. Генетика развития растений: для биологических специальностей университетов. 2-е изд., перераб. и доп. СПб.: Изд-во Н-Л. 432 с.
Магомедмирзаев В.А., 1990. Введение в количественную морфогенетику. М.: Наука. 229 с.
Мазуренко М.Т., Хохряков А.П., 1977. Структура и морфогенез кустарников. М.: Наука. 160 с.
Медведев С.С., Шарова Е.И., 2014. Биология развития растений. Т. 2. Рост и морфогенез. Учебник. Нижневартовск: Изд-во Нижневарт. гос. ун-та. 235 с.
Михалевская О.Б., 1987. Ритмичность процессов роста и морфогенеза побегов в роде Quercus L. // Морфогенез и ритм развития высших растений. М.: Изд-во МГПИ. С. 33–38.
Михалевская О.Б., 2008. Ритмы роста на разных этапах морфогенеза побега у древесных растений // Онтогенез. Т. 39. № 2. С. 85–93.
Сабинин Д.А., 1963. Физиология развития растений. М.: Изд-во АН СССР. 196 с.
Семкина Л.А., Овсянникова О.М., 1998. Морфобиологические особенности форзиции яйцевидной при интродукции на Среднем Урале // Экология и акклиматизация растений. Екатеринбург: Изд. УрО РАН. С. 113–119.
Серебряков И.Г., 1952. Морфология вегетативных органов высших растений. М.: Сов. наука. 391 с.
Серебряков И.Г., 1962. Экологическая морфология растений. М.: Высш. шк. 378 с.
Серебряков И.Г., 1966. Соотношение внутренних и внешних факторов в годичном ритме развития растений // Бот. журн. Т. 41. № 7. С. 923–928.
Творогова В.Е., Осипова М.А., Додуева И.Е., Лутова Л.А., 2012. Взаимодействие транскрипционных факторов и фитогормонов в регуляции активности меристем у растений // Экол. генетика. Т. 10. № 3. С. 28–40.
Aloni R., 2007. Phytohormonal mechanisms that control wood quality formation in young and mature trees // The Compromised Wood Workshop / Eds Entwistle K., Harris P., Walker J. Christchurch: The Wood Technology Research Centre, Univ. of Canterbury. P. 1–22.
Cooke J.E.K., Eriksson M.E., Junttila O., 2012. The dynamic nature of bud dormancy in trees: environmental control and molecular mechanisms // Plant Cell Environ. V. 35. № 10. P. 1707–1728. https://doi.org/10.1111/j.1365-3040.2012.02552.x
Ding X., Jiang Y., Xue F., Zhang Y., Wang M. et al., 2021. Intra‑annual growth dynamics of Picea meyeri needles, shoots, and stems on Luya Mountain, North‑central China // Trees. V. 35. № 2. P. 637–648. https://doi.org/10.1007/s00468-020-02065-9
Edwards K.D., Takata N., Johansson M., Jurca M., Novak O. et al., 2018. Circadian clock components control daily growth activities by modulating cytokinin levels and cell division-associated gene expression in Populus trees // Plant Cell Environ. V. 41. № 6. P. 1468–1482. https://doi.org/10.1111/pce.13185
Farre E.M., 2012. The regulation of plant growth by the circadian clock // Plant Biol. V. 14. № 3. P. 401–410. https://doi.org/10.1111/j.1438-8677.2011.00548.x
Herrmann S., Recht S., Boenn M., Feldhahn L., Angay O. et al., 2015. Endogenous rhythmic growth in oak trees is regulated by internal clocks rather than resource availability // J. Exp. Bot. V. 66. № 22. P. 7113–7127. https://doi.org/10.1093/jxb/erv408
Jackson S.D., 2009. Plant responses to photoperiod // New Phytol. V. 181. № 3. P. 517–531. https://doi.org/10.1111/j.1469-8137.2008.02681.x
Kim J.A., Kim H.-S., Choi S.-H., Jang J.-Y., Jeong M.-J., Lee S.I., 2017. The importance of the circadian clock in regulating plant metabolism // Int. J. Mol. Sci. V. 18. № 12. https://doi.org/10.3390/ijms18122680
Luttge U., Hertel B., 2009. Diurnal and annual rhythms in trees // Trees. V. 23. № 4. P. 683–700. https://doi.org/10.1007/s00468-009-0324-1
Mahmud K.P., Holzapfel B.P., Guisard Y., Smith J.P., Nielsen S., Rogiers S.Y., 2018. Circadian regulation of grapevine root and shoot growth and their modulation by photoperiod and temperature // J. Plant Physiol. V. 222. P. 86–93. https://doi.org/10.1016/j.jplph.2018.01.006
McClung C.R., 2006. Plant circadian rhythms. Historical perspective essay // Plant Cell. V. 18. № 4. P. 792–803.
Miskolczi P., Singh R.K., Tylewicz S., Azeez A., Maurya J.P. et al., 2019. Long-range mobile signals mediate seasonal control of shoot growth // PNAS. V. 116. № 22. P. 10852–10857. https://doi.org/10.1073/pnas.1902199116
Singh R.K., Bhalerao R.P., Eriksson M.E., 2020. Growing in time: Exploring the molecular mechanisms of tree growth // Tree Physiol. V. 41. № 4. P. 657–678. https://doi.org/10.1093/treephys/tpaa065
Singh R.K., Svystun T., AlDahmash B., Jonsson A.M., Bhalerao R.P., 2017. Photoperiod- and temperature-mediated control of phenology in trees – a molecular perspective // New Phytol. V. 213. № 2. P. 511–524. https://doi.org/10.1111/nph.14346
Triozzi P.M., Ramos-Sanchez J.M., Hernandez-Verdeja T., Moreno-Cortes A., Allona I., Perales M., 2018. Photoperiodic regulation of shoot apical growth in poplar // Front. Plant Sci. V. 9. P. 1–9. https://doi.org/10.3389/fpls.2018.01030
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии