

Океанология, 2020, T. 60, № 5, стр. 663-679
Мезомасштабная изменчивость океана в северной части моря Уэдделла
Е. Г. Морозов 1, *, Д. И. Фрей 1, А. А. Полухин 1, В. А. Кречик 1, В. А. Артемьев 1, А. В. Гавриков 1, В. В. Касьян 2, Ф. В. Сапожников 1, Н. В. Гордеева 1, С. Г. Кобылянский 1
1 Институт океанологии им. П.П. Ширшова РАН
Москва, Россия
2 Национальный научный центр морской биологии ДВО РАН
Владивосток, Россия
* E-mail: egmorozov@mail.ru
Поступила в редакцию 15.05.2020
После доработки 21.05.2020
Принята к публикации 22.06.2020
Аннотация
Анализируются результаты мезомасштабной съемки в северной части моря Уэдделла (бассейн Пауэлла) 19–20 февраля 2020 г. Съемка состояла из 24 CTD зондирований (6 × 4) с интервалом 6 морских миль. На станциях выполнялись измерения профилей прозрачности и флюоресценции, пелагические траления и планктонные ловы сетями Джедди и Бонго. Съемка покрывала пространство гидрологического фронта. Фронт проявляется по резким горизонтальным градиентам гидрологических и химических параметров. Он существует за счет различий в свойствах воды, характерной для всего моря Уэдделла, и воды из западной части моря, сформированной на мелководье, покрытом льдом и айсбергами, что не дает возможности солнечной радиации прогреть верхний слой. Фронт в районе мезомасштабной съемки, возможно, появляется за счет отрыва меандров от крупномасштабного фронта в северной части моря Уэдделла и перемещения в район съемки оторвавшегося ринга с охлажденной водой на возвышенности. Различия в структуре вод к северу и югу от фронта проявляются также в химических параметрах, оптических и биологических характеристиках верхнего слоя и, в частности, в концентрациях и видах фитопланктона, зоопланктона и пелагических рыб.
ВВЕДЕНИЕ
Мезомасштабная изменчивость океанологических процессов относится к пространственным масштабам от сотни метров до километров [5]. Как правило, мезомасштабная изменчивость определяется такими явлениями, как вихри, меандрирование фронтов, низкочастотные внутренние волны. Вихри открытого океана определяются волнами Россби и бароклинной неустойчивостью крупномасштабных течений [7]. Такие вихри отличаются от фронтальных вихрей тем, что в вихрях открытого океана содержится такая же вода, что и в окружающем их пространстве. Фронтальные вихри, или ринги, несут захваченную воду с другой стороны фронтального раздела. Структура изотермических, изохалинных и изопикнических поверхностей в синоптических вихрях такова, что в вихрях изоповерхности либо поднимаются, либо заглубляются. При этом форма θ,S-кривых не меняется. Несмотря на то, что ринги или фронтальные вихри подчиняются тем же физическим законам баланса сил давления и Кориолиса, форма θ,S-кривых отлична от кривых для окружающих вод, поскольку они несут захваченную воду, отличающуюся по своим термохалинным, химическим и прочим характеристикам от окружающих вод.
Орбитальные скорости в таких вихрях могут быть много больше, чем скорости средних течений [5]. Возникает важный вопрос о роли мезомасштабной изменчивости в горизонтальном обмене свойств океанской воды, массы, энергии и импульса. Важно понять, как изменяются физические и химические характеристики вод, а также биологические объекты в разных водах на горизонтальных расстояниях, соответствующих мезомасштабной изменчивости. Для понимания процессов, происходящих в океане, нужна оценка степени изменчивости концентрация растворенных веществ, примеси и наличие планктона в разных водах. Такие задачи, относящиеся к физическим процессам, решались в разных районах океана [2, 3, 8, 10–12]. Например, фронтальный вихрь в Антарктическом циркумполярном течении исследовался в работе [4].
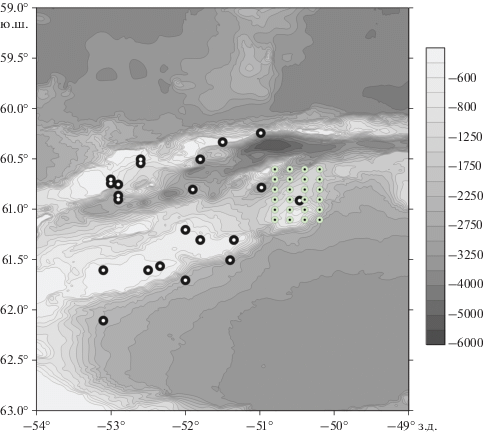
Для изучения изменчивости физических, химических и биологических характеристик на пространственных масштабах около 25–30 морских миль в 79-м рейсе НИС “Академик Мстислав Келдыш” была запланирована мезомасштабная съемка 5 на 5 станций размером около 25 × 25 миль с расстоянием 6 миль между станциями. По результатам измерений на первых галсах план съемки был изменен. В результате была выполнена съемка 4 на 6 станций четырьмя меридиональными разрезами длиной 30 миль. Съемка планировалась таким образом, чтобы ее юго-западный угол оказался на подводной горе с вершиной на глубине около 600 м, а также, чтобы съемка частично перекрывала фронтальную зону, которая разделяет воды морей Скотия и Уэдделла и носит название Weddell-Scotia Confluence Zone (WSCZ) [3, 38]. Схема съемки 19–20 февраля 2020 г. изображена на рис. 1. Отметим, что наша съемка оказалась недостаточно протяженной на север, чтобы достигнуть этой крупномасштабной фронтальной зоны. Район съемки накрыл вторичное фронтальное образование между фронтальной зоной WSCZ и водой, формирующейся на возвышенности в западной части моря Уэдделла. Этот вторичный фронт существует за счет различий в свойствах воды, характерной для всего моря Уэдделла (к югу от WSCZ) и воды из западной части моря, которая формируется на мелководье, покрытом льдом и айсбергами. Оказалось, что вода моря Уэдделла с южной стороны WSCZ располагалась в северной части съемки, а вода с возвышенности в юго-западной части съемки.
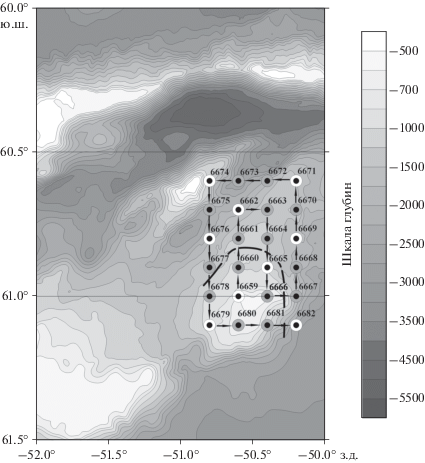
Рис. 1.
Схема мезомасштабной съемки 19–20 февраля 2020 г. Показаны номера станций. Станции на разрезах делались по широтам 60°36, 60°42, 60°48, 60°54, 61°00, 61°06 ю.ш. и по долготам: 50°48, 50°36, 50°24, 50°12 з.д. Ловы сетью Бонго выполнялись на станциях 6659, 6662, 6665, 6669, 6671, 6674, 6676, 6679, 6682 (белые кружки). Стрелки показывают обход станций полигона. Шкала глубин в метрах. Черная штриховая линия схематично показывает линию фронта.
ГИДРОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА РАЙОНА СЪЕМКИ
Термохалинные свойства вод в море Уэдделла описаны в работе [23]. Холодная зимняя вода занимает верхний слой до 100–300 м глубины. Летом верхний слой до 50–100 м прогревается и перемешивается. Глубже залегает слой Теплой глубинной воды, который определяет максимум температуры и солености на промежуточных глубинах около 600 м. Далее залегает слой Глубинной воды моря Уэдделла или в другой терминологии Антарктической донной воды (потенциальная температура θ < 2°С). Под ней находится слой Донной воды моря Уэдделла с температурой менее –0.7°С.
При выполнении съемки выяснилось, что даже на таком малом масштабе съемки на разных станциях структура вод оказалась совершенно различной. В северной части съемки структура вод была типичной для моря Уэдделла [6, 9]. В верхних 100–150 м располагался прогретый летний поверхностный слой с сильным вертикальным градиентом температуры. Под ним на глубине около 200 м расположен летний холодный промежуточный слой с подповерхностным минимумом температуры с отрицательными значениями около –0.5°С (рис. 2). Зимой верхний слой за счет зимней конвекции и ветрового перемешивания превращается в однородный холодный слой. Далее залегает слой теплой глубинной воды с температурой до +0.4°С на глубине около 600 м; далее ко дну температура уменьшается. Теплая глубинная вода наблюдается в море Уэдделла вплоть до 70° ю.ш. [6, 9].
Рис. 2.
Вертикальное распределение температуры на станциях 6660 (толстая линия) и 6673 (тонкая штриховая линия).
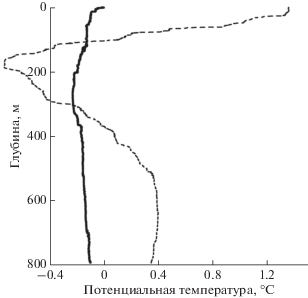
Второй тип воды характеризуется малым изменением температуры с глубиной. Весь столб воды имеет температуру, близкую к отрицательной (рис. 2). Такая структура вод характерна для возвышенности в западной части моря Уэдделла к западу от меридиана 50° W между широтами 60°30 ю.ш. и 62°00 ю.ш. Глубины моря в этом районе находятся в диапазоне 400–700 м. В этот район из южной части моря Уэдделла идет большой вынос льда и айсбергов, которые охлаждают воду до отрицательной температуры, а сильные внутренние волны над возвышенностью перемешивают весь слой воды от поверхности до дна. Эти воды носят название антарктических шельфовых вод. Толщина перемешанного слоя достигает 600 м. Надо отметить, что в экспедиции 79-го рейса НИС “Академик Мстислав Келдыш” мы смогли сделать станции только в северной части района этих вод до 63° ю.ш. Далее к югу были ледовые поля.
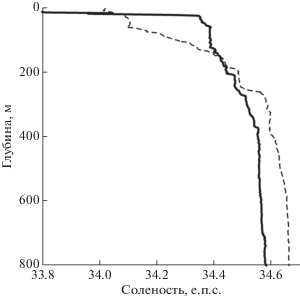
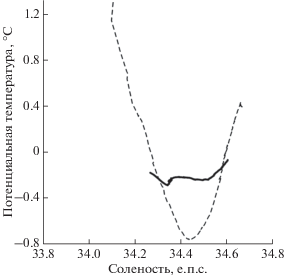
На всех станциях соленость в целом увеличивается с глубиной, тем самым определяя стратификацию вод (рис. 3). В глубинной части соленость на северных станциях несколько выше солености в юго-западной части съемки. Соленость в верхнем слое на станциях над горой (например, станция 6659), которые оказались во фронтальном ринге, близка к 33.9 епс. Такие значения солености отсутствуют на северном разрезе съемки в районе к северу от фронта. Там минимальная соленость составляет 34.0 епс. Очень сильны различия в температурно-соленостных зависимостях (рис. 4).
Рис. 3.
Вертикальное распределение солености на станциях 6659 (толстая линия) и 6671 (тонкая штриховая линия).
Из массива базы данных WOD18 были выделены станции в районе моря Уэдделла, на которых структура вод оказалась близкой к той, что обнаружена на юго-западных станциях мезомасштабной съемки над подводной горой (рис. 5). Все эти станции расположены в северо-западной части моря Уэдделла над возвышенностью. Таким образом, источники холодной воды находятся на возвышенности в западной части бассейна Пауэлла в северо-западной части моря Уэдделла, где вода охлаждается за счет присутствия льда и айсбергов. Плавающий лед не дает возможности солнечной радиации нагревать воду в летнее время.
Рис. 5.
Батиметрия западной части моря Уэдделла. Черные кружки с белой точкой – положения станций из базы данных WOD18, на которых структура водных масс аналогична структуре в юго-западной части мезомасштабной съемки. Светлые кружки с черной точкой показывают положение станций мезомасштабной съемки. Справа дана шкала глубин в метрах в оттенках серого.
Таким образом, представляется, что при проведении мезомасштабной съемки были зафиксированы две различные по структуре воды. Более теплая вода с подповерхностным холодным слоем, типичная для моря Уэдделла, и другая вода адвективного происхождения, обнаруженная на юго-западных станциях съемки и поступившая в район наших работ из западной более мелководной части бассейна Пауэлла (северо-западная часть моря Уэдделла).
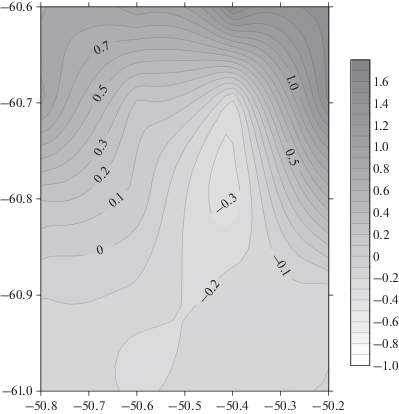
Для подтверждения этой гипотезы рассмотрим данные спутниковой альтиметрии на 19 февраля, то есть на даты выполнения мезомасштабной съемки (рис. 6). На рис. 6 также изображена изобата 1000 м, которая близка к очертаниям меандра по спутниковой динамической топографии. Эта взаимосвязь показывает зависимость положения меандра от донной топографии.
Рис. 6.
Спутниковая альтиметрия 19 февраля 2020 г. Показана абсолютная динамическая топография (Absolute dynamic topography, ADT) Фронтальная зона (ADT = 114–118 см) выделена более темным серым цветом. Справа дана шкала динамической топографии в оттенках серого. На карту альтиметрии нанесена изобата 1000 м (толстая черная линия) и точки станций мезомасштабной съемки (24 черных кружка с белой точкой). Светлый ромб (61°40 ю.ш., 45°45 з.д.) соответствует Южным Оркнейским островам.
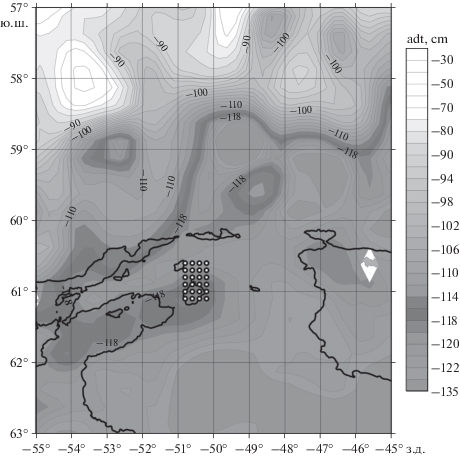
В северной части бассейна Пауэлла находятся относительно теплые воды. Четко выраженная фронтальная зона между этими водами и более холодными водами на юге расположена в диапазоне широт 58°–61°. Фронтальная зона простирается в направлении с юго-запада на северо-восток. В районе 61° ю.ш., 55° з.д. от нее отделяется антициклонический меандр. Представляется, что этот меандр образован влиянием топографии и является квазистационарным. Меандр обтекает возвышенность с глубинами меньше 1000 м, что хорошо видно на совмещенной карте глубин и альтиметрии. Захваченная вода в меандре подвержена сильному охлаждению за счет присутствия большого количества льда и айсбергов в этом районе. Охлаждаясь, эта вода приобретает другие термохалинные свойства. Самой восточной частью этот меандр достигает района нашей мезомасштабной съемки, принося туда холодную воду с другими свойствами. Возможно, этот меандр распадается на несколько отдельных рингов, которые цепочкой расположены над возвышенностью в широтной зоне от 61° ю.ш. до 62° ю.ш. Надо полагать, что подводная гора, расположенная в юго-западной части мезомасштабной съемки, лишь частично изменяет циркуляцию в этом районе. Эта гора является концом возвышенности, над которой располагается меандр.
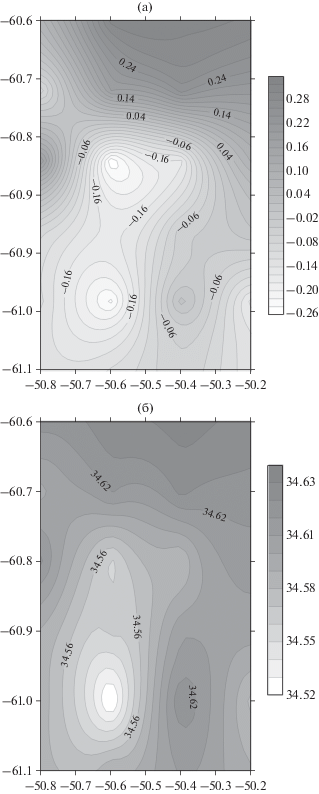
Рассмотрим пространственную изменчивость температуры и солености в районе съемки (рис. 7). Температура и соленость в районе подводной горы несколько меньше, чем в других частях полигона. Однако, как это обычно бывает в океане, такое уменьшение температуры и солености не означает, что изотермы и изохалины, а следовательно, и изопикны прогибаются вниз над горой под воздействием антициклонического вращения. Этот эффект за счет опускания изоповерхностей более слабый, чем заток более холодной и менее соленой воды в район нашей мезомасштабной съемки с юго-запада. Эффект присутствия подводной горы больше сказывается в формировании более холодной воды над возвышенностью, частью которой и является эта подводная гора. Антициклонической вращение создается находящимся над горой меандром или уже оторвавшимся рингом.
Рис. 7.
Карты распределения температуры, °С (а), и солености, епс (б), на глубине 500 м во время выполнения мезомасштабной съемки 19–20 февраля 2020 г. Справа показаны шкалы температуры в оттенках серого, °С, и солености, епс.
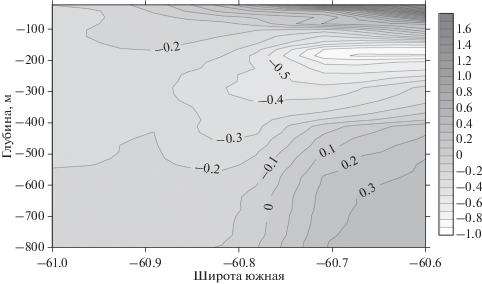
Меридиональный разрез температуры через мезомасштабную съемку наглядно показывает различия в водной структуре на небольшом расстоянии полигона (рис. 8). В южной части обнаруживается почти однородный слой холодной воды, а в северной части – структура вод с прогретым поверхностным слоем, подповерхностным промежуточным холодным слоем и более теплой глубинной водой.
Рис. 8.
Меридиональный разрез температуры по долготе 50°36 з.д. Справа показана шкала температуры в оттенках серого, °С.
Гидрологический фронт очень сильно проявляется в температурных градиентах в поверхностном слое. Перепад по температуре на фронте в верхнем слое достигает 2°С (рис. 9).
Рис. 9.
Карта температуры верхнего слоя океана (50 м) во время мезомасштабной съемки. Справа показана шкала температуры в оттенках серого, °С.
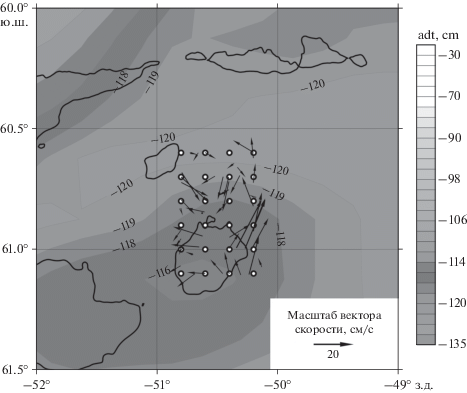
Скорости течения, измеренные погружаемым и бортовым профилографами, показывают удовлетворительное согласие как с геострофическими скоростями, рассчитанными по данным спутниковой альтиметрии, так и между собой. Мгновенные скорости, измеренные профилографами, были подвергнуты обработке для удаления приливных течений, оцененных по модели Университета штата Орегон (США) TPХО 9 [21]. На подходах к станции 6666 и после нее к востоку от подводной горы бортовой допплеровский профилограф течений показывал высокую скорость течения на север во всем слое воды, что характерно для восточной периферии антициклонического вихря. Погружаемый профилограф скорости также показывал высокие скорости около 50 см/с на север. К западу от подводной горы высокие скорости были направлены на запад. Вектора течений на горизонте 100 м показаны на рис. 10.
Рис. 10.
Векторы скоростей течений в районе мезомасштабной съемки по данным бортового профилографа течений. Рисунок показывает фрагмент рисунка 6 с данными спутниковой альтиметрии на 19 февраля 2020 г. Шкала динамической топографии (см) в оттенках серого показана справа. На карту нанесена изобата 1000 м (толстая черная линия) и точки станций мезомасштабной съемки (24 черные кружка с белой точкой).
РАЗЛИЧИЯ В ХИМИЧЕСКИХ ПАРАМЕТРАХ ПО РАЗНЫЕ СТОРОНЫ ФРОНТА
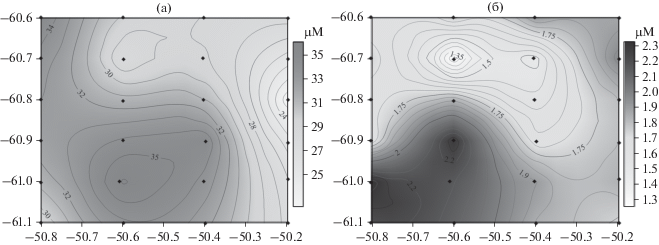
Различия в свойствах вод к северу и югу от фронта подтверждаются химическими определениями содержания нитратов, силикатов и фосфатов. В распределении фосфатов и силикатов в поверхностном слое (3–5 м) выделяются четкие максимумы в юго-западной части полигона, на станции 6659 (рис. 11), что выражается в увеличении концентрации биогенных элементов на поверхности, а также в уменьшении содержания кислорода. Это связано с подъемом глубинных вод над возвышенностью к западу от мезомасштабной съемки там, где находится источник воды из юго-западной части съемки. Высокие значения нитратов и фосфатов на поверхности характерны для вод к югу от фронта и, по всей видимости, для вод над возвышенностью к западу от мезомасштабной съемки. Этот процесс хорошо известен и неоднократно отмечался в других районах Мирового океана [7, 19, 39].
Рис. 11.
Карты распределения нитратов (а) и фосфатов (б) на поверхности в районе мезомасштабной съемки. Шкалы концентрации нитратов и фосфатов (μM) в оттенках серого показаны справа.
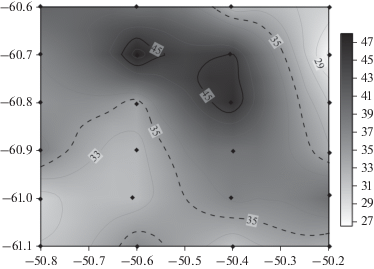
По соотношению кремний/фосфор отчетливо выделяется фронтальный раздел между водами моря Уэдделла и водами над возвышенностью к западу от мезомасштабной съемки (рис. 12). На фронтальном разделе соотношение кремния к фосфору более 35, что характерно для высокоширотной модификации водной массы моря Уэдделла [1]. Южнее фронтальной зоны это соотношение ниже 35, что указывает на воды фронтальной зоны. Кроме того, эти воды наблюдаются именно в юго-западной части полигона, над подводной возвышенностью. Антициклонический фронтальный вихрь переместил сюда воду с запада. Распределение фосфатов и растворенного кислорода в толще воды также указывает на различные водные массы, характерные для района Атлантического сектора Южного океана [17].
Рис. 12.
Карта поверхностного распределения отношения концентраций силикатов и фосфатов. Шкала отношения в оттенках серого показана справа.
На основании полученных гидрохимических данных можно сделать вывод, что в районе проведенных исследований наблюдается значительная изменчивость содержания биогенных элементов, вызванная наличием вихря над подводной возвышенностью. Адвекция холодных вод с юго-запада и сопутствующее ей поднятие глубинных вод вдоль склона горы приводит к увеличению содержания фосфатов и силикатов. В перспективе это может привести к активизации продуцирования органического вещества [19, 22] и других биологических процессов. Дальнейшее взаимодействие двух масс воды с разными свойствами усиливает изменчивость гидрохимических параметров в толще вод, что характерно для фронтальных зон Южного океана [36].
ОПТИЧЕСКИЕ ХАРАКТЕРИСТИКИ ВЕРХНЕГО СЛОЯ
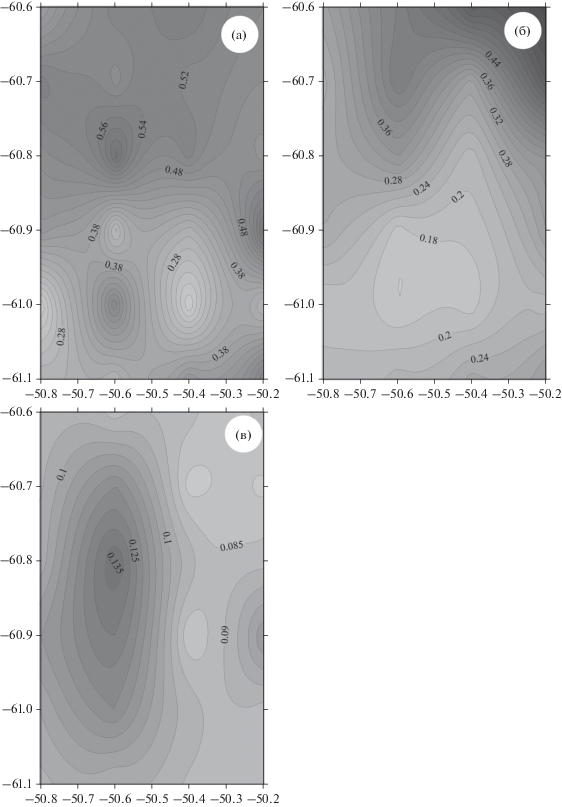
Различия в структуре вод по разные стороны фронта проявляются и в оптических характеристиках. Показатель ослабления света (который является индикатором концентрации взвеси и характеризует мутность воды) в поверхностном слое (5–7 м), к северу от фронта значительно выше (0.6 м–1), чем к югу (0.2–0.3 м–1). Аналогичная изменчивость наблюдается и по распределению флюоресценции. Флюоресценция к северу от фронта значительно выше, чем к югу от фронта: соответственно 0.50 и 0.14 ед. (рис. 13). В целом, в этом слое распределения показателя ослабления хорошо коррелируют с температурой, и в меньшей степени, с распределением флюоресценции, что, вероятно, связано с изменением видового состава биогенной компоненты взвеси: в юго-западной части полигона поверхностная температура меньше на ~ 0.5–1.0°С, чем в северо-восточной. На глубине 190 м значения показателя ослабления возрастают над подводной горой до 0.15 м–1, что в два раза выше, чем на других станциях мезомасштабной съемки (рис. 13). Это связано с взмучиванием донных отложений течениями при взаимодействии со склоном, что становится заметно уже на глубине 190 м. Вершина горы находится на глубине около 600 м.
Рис. 13.
Карты оптических характеристик во время мезомасштабной съемки. Показатель ослабления м–1, горизонт 5–7 м (а); Хлорофилл о.е., горизонт 25 м (б); Показатель ослабления м–1, горизонт 190 м (в).
Различия оптических характеристик наглядно проявляются и на их вертикальных распределениях на станциях 6660 (над горой) и 6671 (северо-восточная часть) (рис. 14). На ст. 6660 в приповерхностном тонком 12–14-метровом слое наблюдается типичная ситуация для зоны таяния льда. Температура более высокая за счет летнего прогрева. Изменения показателя ослабления и флюоресценции происходят практически синхронно на фоне увеличения солености с глубиной, связанного с поверхностным распреснением, вызванным таянием льда и айсбергов. В зонах таяния морского льда в воду попадают диатомовые водоросли, которые вызывают возрастание флюоресценции и показателя ослабления. Ниже этой зоны значения температуры и флюоресценции выходят на квазипостоянные значения, которые практически не меняются до глубин около 200 м. Значения показателя ослабления монотонно снижаются до 0.15 м–1.
Рис. 14.
Вертикальные распределения флюоресценции (тонкая линия) и показателя ослабления (толстая линия) на станциях 6660 (над горой) и 6671 (северо-восточный угол).
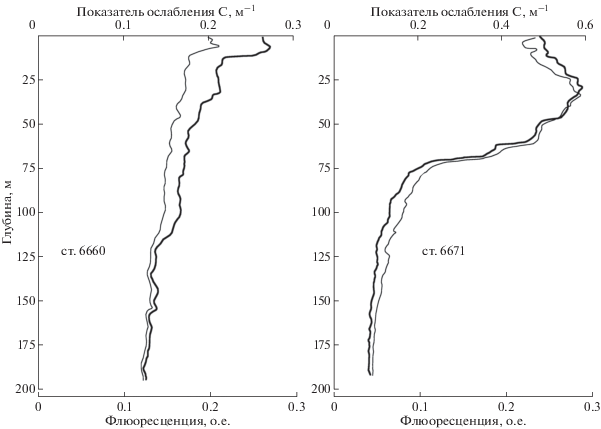
На ст. 6671 в северо-восточной части съемки картина иная. Здесь расположен мощный квазиоднородный по температуре верхний слой прогретой воды толщиной ~70 м. На глубине 25 м сформировался максимум показателя ослабления и флюоресценции, соответственно 0.58 м–1 и 0.65 о.е. Температура опускается ниже нуля на глубинах более 100 м, однако значения флюоресценции остаются значимыми (0.17 о.е.) и выходят на фоновые значения глубже 150 м. На глубине 190 м значения показателя ослабления опускаются до 0.087. На ст. 6663 в этой части мезополигона, значения составляют 0.07 м–1, что является рекордом прозрачности для всех станций 79 рейса. Высокая корреляционная зависимость между показателем ослабления и флюоресценцией говорит о том, что в составе взвеси доминирует биогенная компонента. Оптические данные подтверждают различия гидрологических характеристик разных частей мезополигона.
ФИТОПЛАКТОН К СЕВЕРУ И К ЮГУ ОТ ФРОНТА МЕЗОПОЛИГОНА
Наблюдения за состоянием (составом, структурой и пространственной организацией) фитопланктона на станциях мезомасштабной съемки показали ряд отличий сообществ к югу от гидрологического фронта – от таковых, изученных к северу от фронта. Эти отличия коснулись как набора видов, представленных свободноплавающими клетками, так и тех видов, что образуют эпибионтные агрегаты на поверхности хет (щетинок) панцирей крупных планктонных диатомей.
К югу от фронта сообщества были образованы крупными видами свободноплавающих тихопелагических диатомей, такими как Corethron pennatum, Chaetoceros criophilum, Ch. convolutus, Stellarima microastris и Thalassiothrix antarctica. Все эти виды обычны для антарктических вод [29, 40]. В эуфотическом слое, в целом, преобладали Corethron pennatum и Ch. criophilum. Эпибионтные комплексы на поверхности щетинок, строящиеся на основе матрикса колоний гетеротрофных бактерий, были образованы тремя видами мелких гетеротрофных жгутиконосцев и колониальными видами диатомей из числа ледовой флоры [14, 30, 33]: Fragilariopsis nana, F. curta, F. cylindrus, изредка – F. kerguelensis, а также небольшими колониями Chaetoceros debilis и Ch. laciniosus. В приповерхностном слое, на глубинах около 4 м – согласно материалам, полученным с помощью проточной системы, резко доминировал Th. antarctica (с небольшой примесью C. pennatum и Ch. criophilum). На основе сплетений их длинных клеток, фрагментов колоний и мертвых панцирей формировались парусовидные бактериальные колонии, на матриксе которых в массе обитали уже упомянутые жгутиконосцы и, изредка, короткие лентовидные колонии видов Fragilariopsis (F. nana и F. kerguelensis), а также единичные клетки Licmophora spp.
На станциях к северу от фронта основными структурными элементами фитопланктона по всему эуфотическому слою стали два колониальных вида диатомей из рода Pseudonitzschia (P. antarctica и P. turgidula). Здесь им сопутствовали массовые C. pennatum и Ch. criophilum, а на глубинах около 4 м преобладал все тот же Th. antarctica, а также часто встречались клетки C. pennatum на разных стадиях перехода к половому размножению. Тем не менее, состав эпибионтных комплексов на хетах Corethron и клетках Th. antarctica (по сути, целиком выглядящих как тонкие щетинки) здесь существенно изменился. Теперь на матриксе колониальных бактерий в массе развивались колонии Pseudonitzschia, мелкие жгутиконосцы стали более массовыми и число их видов достигло шести, отмечались массивные колониальные поселения мелких бентосных видов из родов Amphora, Halamphora и Nitzschia (суммарно 10 видов), частый массовый рост Licmophora spp., а также регулярно попадались клетки динофлагелляты Protoperidinium defectum. Колонии Fragilariopsis spp. также присутствовали, но их роль в структуре эпибионтных комплексов была выражена уже в меньшей степени.
В целом, можно говорить о существенной перестройке фитопланктонных ценозов к северу от фронта. Изменения выражались в общем увеличении разнообразия флоры, появлении в составе сообществ массовых видов из рода Pseudonitzshia, а также в существенной реструктуризации и росте разнообразия эпибионтных комплексов, формирующихся на панцирях крупных диатомей, в частности, на их щетинковидных структурах, обеспечивающих парение клеток в толще воды.
Эпибионтные комплексы представляют особый интерес, поскольку, как показали наши наблюдения, служат основой рациона криля в этих водах.
ЗООПЛАНКТОН В РАЙОНЕ МЕЗОМАСШТАБНОЙ СЪЕМКИ
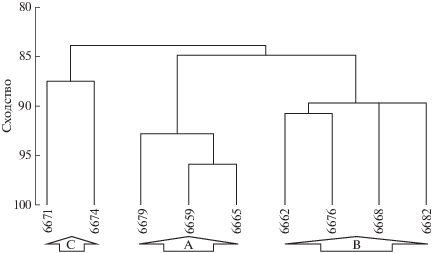
Исследования зоопланктона в бассейне Пауэлла в северной части моря Уэдделла показали существенные различия сообществ по видовому составу, структуре и пространственному распределению в направлении от юга гидрологического фронта к его северу. С помощью кластерного анализа все станции разделились на три группы (рис. 15).
Рис. 15.
Дендрограмма сходства между станциями (пробами) (метрика – коэффициент сходства Брэя–Кертиса, метод средней связи [16]. Буквами (А, В, С) обозначены группы (сообщества) зоопланктона. По оси абсцисс отложены номера станций.
Первая группа (A) отмечена южнее района фронта в водах меандра с отрицательной температурой и характеризовалась самой низкой численностью (рис. 16) и видовым разнообразием сообществ зоопланктона. В планктоне преобладали копеподы родов Metridia, Oithona, Oncea и Triconia – виды, характерные для прибрежных районов Антарктического полуострова [25, 26]. Глубже 200 м единично встречались крупные копеподы родов океанического комплекса – Calanus, Calanoides, а также Ostracoda. Обычно копеподы родов Calanus и Calanoides встречаются на глубине 0–250 м летом и 500–1000 м зимой [18] и являются доминирующими по численности в летний период [31].
Рис. 16.
Изменение средней численности А (%), В (экз./м3) основных таксонов зоопланктона в выделенных сообществах.
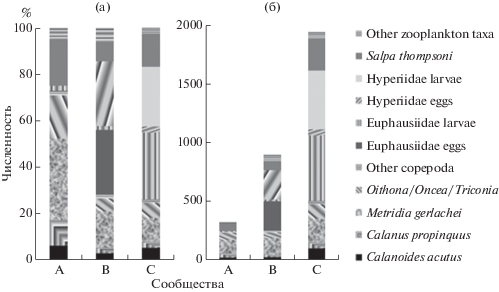
В районе фронта во второй группе (B) численность и видовое разнообразие зоопланктона увеличились. В летнем прогретом поверхностном слое отмечено появление большого количества (свыше 250 ± 76 зкз/м3) яиц Hyperiidae и Euphausiidae (рис. 16). В меньшем количестве встречались взрослые особи гипериид (Hyperiidae), птеропод (Clione) и личинки (calyptopes) эвфаузиид. Единично были отмечены не встречающиеся ранее копеподы Paraeuchaeta antarctica, Rhincalanus gigas, а также Siphonophorae (Diphyes) и copepod nauplii. Крупные копеподы (т.к. Calanus propinquus, Calanoides acutus), которые встречались южнее района фронта в холодном слое 200–500 м, в этом районе были обнаружены выше 200 м. В районе фронта холодные антарктические воды наблюдались в подповерхностном слое выше 200 м, где создавались условия для скопления крупных копепод.
Сообщества (C), расположенные севернее района фронта в типичной воде для моря Уэдделла [6], характеризовались высокой численностью (более 500 ± 103 экз./м3) личинок (calyptopes and furcilia) Euphausiidae и Hyperiidae. Личинки Euphausiidae концентрировались в поверхностном теплом слое (0–200 м), в то время как личинки Hyperiidae были сосредоточены в слое Теплой глубинной воды (200–500 м). В этой группировке встречались копеподы родов Calanus, Calanoides, Ctenocalanus, Metridia, Oithona, Oncea, Paraeuchaeta, Triconia, Rhincalanus, а также Chaetognatha, Euphausiidae (Thysanoessa), Pteropoda (Clione), Ostracoda, Siphonophorae (Diphyes) и личинки Polychaeta. Здесь была отмечена самая высокая численность и разнообразие сообществ зоопланктона, превосходящая таковую в других исследованных районах в 3–5 раз (рис. 16). Повышенная численность личинок в сообществах, расположенных севернее района фронта показывает, что эта акватория может служить важной областью откорма китов, которые, как известно, активно охотятся на эвфаузиид и гипериид [27].
Известно, что эвфаузииды Euphausiidae – важный компонент сообщества макрозоопланктона в прибрежных экосистемах Антарктиды [20, 24]. Криль Euphausia superba, питающийся фитопланктоном и мелким зоопланктоном, служит пищей для усатых китов, тюленей-крабоедов, пелагических рыб и птиц [41, 42]. В исследуемом районе особи E. superba не образовывали плотных скоплений. Единичные экземпляры E. superba были отмечены в сообществах, расположенных в районе фронта и севернее от него, а небольшое увеличение концентрации (до 23 мг/м3) наблюдалось южнее района фронта. Обнаруженный криль был представлен особями размером от 38 до 52 мм, при средней длине 36.6 мм. Самки были представлены на ранних стадиях развития IIIB и IIIA, в то время как самцы были преимущественно на IIIA стадии развития. Соотношение полов было 1.0 : 1.3 с незначительным преобладанием самцов.
В последние десятилетия сальпа Salpa thompsoni демонстрирует повышенную распространенность в высоких широтах Южного океана и в настоящее время часто является численно доминирующим таксоном зоопланктона в водах Антарктического полуострова. Предполагается, что увеличение численности этого вида в водах высоких широт связано с потеплением океана [32]. В нашем исследовании наибольшее количество S. thompsoni было отмечено в сообществах, расположенных в районе фронта и севернее от него, с концентрацией биомассы 0.66 и 1.62 г/м3 соответственно. При этом количественные показатели криля E. superba были незначительными. Возможно, это обусловлено тем, что происходит выедание сальпами яиц и личинок младших возрастных стадий рачков. Сальпы могут играть важную роль в биогеохимических циклах Южного океана, но они мало связаны с более высокими трофическими уровнями – пелагическими рыбами, морскими птицами, китами и т.д.
Известно, что в регионе Антарктического полуострова продолжают происходить резкие изменения климата, в том числе повышение температуры поверхности моря [13] и уменьшение протяженности морского льда зимой [43]. В сочетании с этими гидрофизическими изменениями первичная продукция, структура сообществ фитопланктона и зоопланктона в толще воды, вероятно, продолжат изменяться, открывая возможности для увеличения популяционной плотности других ранее немногочисленных видов [28, 32, 44].
Таким образом, структура зоопланктонных сообществ в бассейне Пауэлла в северной части моря Уэдделла тесно связана с распределением водных масс. По видовому составу, структуре и пространственному распределению сообществ было выделено три группы, связанные с двумя типами водных масс: типичная вода для моря Уэдделла и холодная вода над возвышенностью. Наиболее высокая численность и видовое разнообразие отмечены для типичной воды моря Уэдделла, наиболее низкая для холодной воды, принесенной с возвышенности.
В районе фронта в летнем поверхностном прогретом слое наблюдалось массовое размножение эвфаузиид и гипериид, а севернее района фронта – скопление их личинок. Личинки Euphausiidae концентрировались в поверхностном теплом слое, в то время как личинки Hyperiidae были сосредоточены слое Теплой глубинной воды.
В целом, по численности преобладали Copepoda (Calanus propinquus, Calanoides acutus and Metridia gerlachei), личинки и яйца Hyperiidae и Euphausiidae, по биомассе – Salpida. Численность и видовое разнообразие зоопланктона увеличивались в направлении через фронт к северу.
Криль E. superba не образовывал плотных скоплений в изученном районе, а наибольшее количество сальпы S. thompsoni было отмечено в сообществах, расположенных в районе фронта и севернее от него. Обилие рачков антарктического криля E. superba будет определяться целым комплексом процессов, среди которых основными являются интенсивность нереста в пределах области размножения, интенсивность транспорта личинок течениями и выедания хищниками.
ПЕЛАГИЧЕСКИЕ РЫБЫ ПО РАЗНЫЕ СТОРОНЫ ФРОНТА
Ихтиологические исследования с помощью пелагического траления южнее и севернее фронтальной зоны показали различия в видовом составе пойманных рыб. Пелагические тралы брались не только в районе мезомасштабной съемки, но к северу и к югу от него.
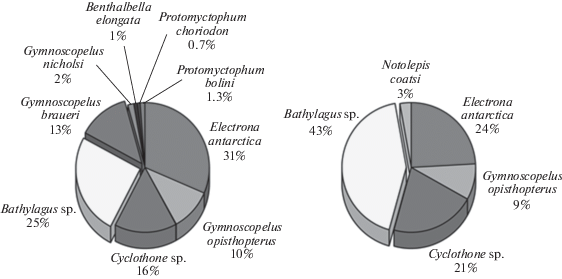
Глубоководные ихтиологические материалы (ихтиопланктон и взрослые мезопелагические рыбы) получены при помощи разноглубинного не замыкающегося трала Айзекса-Кидда в модификации Самышева-Асеева (РТАКСА) с площадью раскрытия около 6 м2. Поскольку наибольшая эффективность РТАКСА для ихтиологических сборов была отмечена в период работ на глубинах больше 500 м, сравнение проводилось между группами станций с примерно одинаковыми горизонтами лова: 2212-1100-0 м на станциях южнее линии фронта (60°42′ ю.ш.) и 2062-634-0 м – на станциях, расположенных севернее фронта. На шести “северных” станциях (ст. 6684–6688 и 6690; 59°42′–60°39′ ю.ш., 48°04′–50°38′ з.д.) было получено 220 экз. рыб, тогда как на пяти “южных” (ст. 6607, 6617, 6619, 6653 и 6655; 61°06′–62°29′ ю.ш., 48°47′–53°16′ з.д.) – 149 экз. без учета личинок, относящихся в обоих случаях по большей части к сем. Paralepididae. В уловах были представлены основные глубоководные виды антарктического региона [15, 35, 37], включая светящихся анчоусов Electrona antarctica, Gymnoscopelus opisthopterus, Gymnoscopelus braueri, Gymnoscopelus nicholsi, Protomyctophum choriodon, Protomyctophum bolini (сем. Myctophidae), гоностомовых рыб рода Cyclothone (сем. Gonostomatidae), батилагов (Bathylagus sp., сем. Bathylagidae), а также единичные экземпляры веретенниковых (Notolepis coatsi, сем. Paralepididae) и скопеларховых рыб (Benthalbella elongata, сем. Scopelarchidae) (рис. 17). На северных станциях число пойманных видов оказалось достоверно выше, чем на южных (в среднем 6 против 4; непараметрический U-тест Манна-Уитни [34]: p = 0.0191). Кроме того, на южных станциях в уловах заметно доминировали батилаги Bathylagus sp. (в среднем 43% против 25%, см. рис. 17), а также совершенно отсутствовал вид Gymnoscopelus braueri, различия в частотах встречаемости которого между двумя группами станций оказались статистически значимы (тест Манна–Уитни: p = 0.0095). В сборах на южных станциях также отсутствовали такие виды светящихся анчоусов, как Gymnoscopelus braueri, Gymnoscopelus nicholsi, Protomyctophum choriodon и Protomyctophum bolini, однако различия в частотах их встречаемости не нашли статистического подтверждения в силу малого числа пойманных экземпляров.
ВЫВОДЫ
Фронт, обнаруженный в северной части моря Уэдделла, существует за счет различий в свойствах воды, характерной для всего моря Уэдделла и воды из западной части моря, которая формируется на мелководье, покрытом льдом и айсбергами, что не дает возможности солнечной радиации прогреть верхний слой. Появление фронта в исследуемом районе происходит за счет существования меандров крупномасштабного фронта в северной части моря Уэдделла и возможного перемещения оторвавшегося ринга на полигон мезомасштабной съемки. Резкие горизонтальные градиенты температуры и солености внутри фронтальной зоны проявляются в дифференциации оптических характеристик и гидрохимических параметров к югу и северу от фронтального раздела. Различия в концентрации биогенных элементов и распределении гидрофизических показателей, в свою очередь, находят отражение в численности и видовом составе фитопланктона, зоопланктона и пелагических рыб. Структура зоопланктонных сообществ в бассейне Пауэлла в северной части моря Уэдделла тесно связана с распределением водных масс. В частности, криль Euphausia superba, важнейший элемент трофической цепи вод Антарктики, отмечался в сообществах, расположенных в районе фронта и севернее от него.
Источник финансирования. Работа выполнена по государственному заданию 0128-2019-0008. Анализ CTD-данных поддержан грантом РНФ 16-17-10 149. Вычислительные работы поддержаны грантом РФФИ 19-57-60 001.
Список литературы
Аржанова Н.В., Артамонова К.В. Гидрохимическая структура вод в районах промысла антарктического криля Euphausia superba Dana // Труды ВНИРО. 2014. Т. 152. С. 118–132.
Берестов А.Л., Иванов Ю.А., Корт В.Г., Кошляков М.Н., Морозов Е.Г., Морошкин К.В., Овчинников И.М., Шадрин И.Ф., Шаповалов С.М., Щербинин А.Д., Яремчук М.И. Мезомасштабные вихри верхнего слоя океана // Океанология. 1987. Т. 27. № 6. С. 889–895.
Богданов М.А., Орадовский С.Г., Солянкин Е.В., Хвацкий Н.В. О фронтальной зоне в море Скоша // Океанология. 1969. Т. 11. № 6. С. 966–974.
Иванов Ю.А., Краснопевцев А.Ю., Морозов Е.Г., Шадрин И.Ф., Щербинин А.Д., Циклонический фронтальный вихрь в Антарктическом циркумполярном течении // Океанология. 1985. Т. 25. № 1. С. 32–36.
Каменкович В.М., Кошляков М.Н., Монин А.С. Синоптические вихри в океане // Л: Гидрометеоиздат, 1982. 264 с. (Kamenkovich V.M., Koshlyakov M.N., Monin A.S. Synoptic Eddies in the Ocean, Springer. 1986).
Клепиков В.В. Гидрология моря Уэдделла // Труды Сов. Антарктической. Экспедиции. 1963. Т. 17. С. 45–93.
Кошляков М.Н. Открытие и исследование вихрей открытого океана // Изв. РАН. Серия ФАО. 2002. Т. 38. № 6. С. 770–783.
Куклев С.Б., Силкин В.А., Часовников В.К., Зацепин А.Г., Паутова Л.А., Подымов О.И., Баранов В.И., Куклева О.Н., Федоров А.В., Лифанчук А.В. Результаты исследования влияния субмезомасштабного вихря на пространственное распределение гидрохимических параметров и структурно-функциональные характеристики фитопланктона в шельфовой зоне Черного моря // Экология гидросферы. 2019, № 1. С. 24–40. URL: http://hydrosphere-ecology.ru/139
Макеров Ю.В. Основные черты гидрологического режима антарктических вод // Сб. Антарктика. Л.: Гидрометеоиздат. 1956. С. 16–34.
Морозов Е.Г., Никитин С.В., Шаповалов С.М. Мезомасштабная изменчивость по измерениям на буйковых станциях в Ново-Зеландском районе Южного океана, Океанологические исследования № 39. 1985. 53–64.
Морозов Е.Г., Пелиновский Е.Н., Талипова Т.Г. Частота повторяемости внутренних волн на Мезополигоне-85 в Атлантике // Океанология. 1998. Т. 38. № 4. С. 521–527.
Морозов Е.Г., Нейман В.Г. Спектральные характеристики мезомасштабной изменчивости придонных океанских течений // Докл. РАН. 2016. Т. 471. № 1. С. 98–101.
Anisimov O.A., Vaughan D.G., Callaghan T.V., Furgal C. and others. Polar regions (Arctic and Antarctic). In: Parry M.L., Canziani O.F., Palutikof J.P., van der Linden P.J., Hanson C.E. (eds). Climate Change 2007: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press. 2007. P. 653− 685.
Cefarelli A.O., Ferrario M.E., Almandoz G.O., Atencio A.G., Akselman R., Vernet M. Diversity of the diatom genus Fragilariopsis in the Argentine Sea and Antarctic waters: morphology, distribution and abundance // Polar Biology. 2010. V. 33. P. 1463–1484. https://doi.org/10.1007/s00300-010-0794-z
Christiansen H., Dettai A., Heindler F.M., Collins M.A., Duhamel G., Hautecoeur M., Steinke D., Volckaert F.A.M., Van de Putte A.P., Diversity of mesopelagic fishes in the Southern Ocean – A phylogeographic perspective using DNA barcoding // Frontiers in Ecology and Evolution. 2018. V. 6. P. 1–19.
Clarke K.R., Gorley R.N. PRIMER: Getting started with v6. PRIMER-E Ltd: Plymouth. 2005. 122 p.
Dafner E.V., Mordasova N.V. Influence of biotic factors on the hydrochemical structure of surface water in the Polar Frontal Zone of the Atlantic Antarctic // Marine Chemistry. 1994. № 1–2 (45). P. 137–148.
Delgado, L.E., Jana R., Marin V.H. Testing hypotheses on life-cycle models for Antarctic calanoid copepods, using qualitative, winter, zooplankton samples // Polar Biology. 1998. V. 20. P. 74–76. https://doi.org/10.1007/s003000050278
Doblin M.A. et al. Nutrient uplift in a cyclonic eddy increases diversity, primary productivity and iron demand of microbial communities relative to a western boundary current // PeerJ. 2016. 2016. № 4. https://doi.org/10.7717/peerj.1973
Driscoll R.M., Reiss C.S., Hentschel B.T. Temperature-dependent growth of Thysanoessa macrura: inter-annual and spatial variability around Elephant Island, Antarctica // Mar. Ecol. Prog. Ser. 2015. V. 529. P. 49–61.
Egbert G.D., Erofeeva S.Y. Efficient inverse modeling of barotropic ocean tides // J. Atmospheric and Oceanic Technology. 2002. V. 19. № 2. P. 183–204. https://doi.org/10.1175/1520-0426(2002)019<0183: EIMOBO>2.0.CO;2
Ellwood M.J. et al. Distinct iron cycling in a Southern Ocean eddy // Nature Communications. 2020. № 1 (11). P. 1–8.
Fahrbach E., Rohardt G., Scheele N, Schriider M., Strass V., Wisotzki A., Formation and discharge of deep and bottom water in the northwestern Weddell Sea // Journal of Marine Research. 1995. V. 53. P. 515–538.
Flores H., Atkinson A., Kawaguchi S., Krafft B.A., Milinevsky G., Nicol S. et al., Impact of climate change on Antarctic krill // Mar. Ecol. Prog. Ser. 2012. V. 458. P. 1–19. https://doi.org/10.3354/meps09831
Foster B.A. Time and depth comparisons of sub-ice zooplankton in McMurdo Sound, Antarctica // Polar Biology. 1989. V. 9. P. 431–435.
Fransz H.G. Vernal abundance, structure and development of epipelagic copepod populations of the eastern Weddell Sea (Antarctica) // Polar Biology. 1988. V. 9. P. 107–114.
Hempel G. Antarctic marine food web. In: Siegfried W.R., Condy P.R., Laws R.M. (eds.) Antarctic nutrient cycles and food webs. Berlin: Springer. 1985a. P. 266–270.
Hewes C.D., Reiss C.S., Holm-Hansen O. A quantitative analysis of sources for summertime phytoplankton variability over 18 years in the South Shetland Islands (Antarctica) region // Deep-Sea Res. 2009. V. 56. P. 1230− 1241.
Kang S.-H., Fryxell G.A. Phytoplankton in the Weddell Sea, Antarctica: composition, abundance and distribution in water-column assemblages of the marginal ice-edge zone during austral autumn // Marine Biology. 1993. V. 116. P. 335–348.
Kang S.-H., Fryxell G.A., Roelke D.L. Fragilariopsis cylindrus compared with other species of the diatom family Bacillariaceae in Antarctic marginal ice-edge zones // Nova Hedwigia Beih, 1993. V. 106. P. 335–352.
Lee W., Kim S., Kang S., Bang H., Lee K., Kwak I. Distribution and abundance of zooplankton in the Bransfield Strait and the western Weddell Sea during austral summer // Ocean and Polar Res. 2004. V. 26. № 4. P. 607–618. https://doi.org/10.4217/OPR.2004.26.4.607
Loeb V.J., Santora J.A. Population dynamics of Salpa thompsoni near the Antarctic Peninsula: Growth rates and interannual variations in reproductive activity (1993–2009) // Progr. in Oceanog. 2012. V. 96. № 1. P. 93–107. https://doi.org/10.1016/j.pocean.2011.11.001
Lundholm N., Rytter H.G. Are Fragilariopsis cylindrus and Fragilariopsis nana bipolar diatoms? – Morphological and molecular analyses of two sympatric species // Nova Hedwigia, Beiheft. 2008. V. 133. P. 231–250.
Mann H.B., Whitney D.R. On a test of whether one of two random variables is stochastically larger than the other // Annals of Mathematical Statistics. 1947. № 18. P. 50–60.
Martin G., White M.G., Piatkowski U. Abundance, horizontal and vertical distribution of fish in eustern Weddell Sea micronekton, Polar Biology. 1992. V. 13. P. 41–53.
Maximenko N., Yamagata T. Submesoscale anomalies in the North Pacific Subarctic Front // J. Geophysical Research. 1995. V. 100 № C9. P. 18 459–18 469.
Parker M.L., Fraser W.R., Ashford J., Patarnello T., Zane L., Torres J.J., Assemblages of micronektonic fishes and invertebrates in a gradient of regional warming along the Western Antarctic Peninsula // J. Marine Systems. 2015. V. 152. P. 18–41.
Patterson S.L., Sievers H.A. The Weddell-Scotia confluence // J. Phys. Oceanog. 1980. V. 10. № 10. P. 1584– 1610.
Sapozhnikov V.V., Ivanova O.S., Mordasova N.V. Identification of local upwelling zones in the Bering Sea using hydrochemical parameters // Oceanology. 2011. V. 51. № 2. P. 247–254. https://doi.org/10.1134/S0001437011020135
Schloss I.R., Ferreyra G.A., Ruiz-Pino D. Phytoplankton biomass in Antarctic shelf zones: a conceptual model based on Potter Cove, King George Island // Journal of Marine Systems. 2002. V. 36. P. 129–143.
Siegel V., Watkins J.L. Distribution, Biomass and Demography of Antarctic Krill, Euphausia superba. In: Siegel V. (eds.) Biology and Ecology of Antarctic Krill. Advances in Polar Ecology. Springer, Cham. 2016. P. 21–100. https://doi.org/10.1007/978-3-319-29279-3_2
Sprong I., Schalk P.H. Acoustic observations on krill spring-summer migration and patchiness in the northern Weddell Sea. In: Hempel G. (eds.) Weddell Sea Ecology. Springer, Berlin, Heidelberg. 1992. P. 261–268. https://doi.org/10.1007/978-3-642-77595-6_30
Stammerjohn S.E., Martinson D.G., Smith R.C., Yuan X., Rind D. Trends in Antarctic annual sea ice retreat and advance and their relation to El Nino-Southern Oscillation and Southern Annular Mode variability // J. Geophys. Res Oceans. 2008. V. 113. P. C03S90.
Voronina N.M. Comparative abundance and distribution of major filter-feeders in the Antarctic pelagic zone // J. of Mar. Sys. 1998. V. 17. № 1–4. P. 375–390. https://doi.org/10.1016/S0924-7963(98)00050-5
Дополнительные материалы отсутствуют.
Инструменты
Океанология