

Океанология, 2021, T. 61, № 1, стр. 141-146
Специфика формирования маргинальных фильтров в приливных устьях малых рек арктических морей
И. В. Мискевич 1, *, В. Б. Коробов 1, Д. С. Мосеев 1
1 Институт океанологии им. П.П. Ширшова РАН
Москва, Россия
* E-mail: subarct@gmail.com
Поступила в редакцию 21.05.2020
После доработки 09.06.2020
Принята к публикации 15.08.2020
Аннотация
Проведены исследования для оценки характера функционирования маргинального фильтра в приливных устьях малых рек арктических морей. Для этой цели использованы данные наблюдений в Белом и Баренцевом морях. При наличии обширных приливных осушек и зарослей галофитов в устьях малых рек биогеохимические процессы могут заметно отличаться от условий, описываемых теорией классического маргинального фильтра. Это связано с тем, что она разработана на примере устьев больших рек и не учитывает специфику экосистем устьев малых рек. В них может меняться пространственная схема чередования мутьевой, химической и биологических “пробок”. Обращено внимание на маргинальные фильтры в лагунных устьях арктических рек с наличием водных макрофитов, которые могут препятствовать переносу в море нерастворимых веществ, освобождаемых при таянии мерзлотных грунтов и ледников.
Теория маргинального фильтра как глобального фильтра взвешенного и растворенного вещества, поступающего в океан с континентов, разработанная академиком А.П. Лисицыным, нашла свое подтверждение при исследованиях большого количества устьев рек в различных регионах нашей планеты [1, 5, 6, 16]. Однако почти все они относятся к так называемым большим рекам, под которыми, согласно ГОСТ 19 179-73 “Гидрология суши. Термины и определения”, принято понимать водотоки с площадью водосбора не менее 50 000 км2. Практически во всех публикациях, посвященных этой проблеме, размеры рек не учитываются, что автоматически подразумевает распространение основных положений вышеупомянутой теории на морские устья любых водотоков. Кроме того, существуют неясности в функционировании маргинального фильтра в устьях арктических морей в холодный период при наличии припая на устьевом взморье реки и на прилегающем участке моря из-за крайне малого количества натурных комплексных наблюдений в зимний сезон.
Исследования, проведенные Северо-Западным отделением Института океанологии РАН в 2014–2020 гг. в Белом и Баренцевом морях, выявили заметные отклонения биогеохимических процессов в устьях малых рек от характеристик, присущих устьям крупных рек этих морей [9, 10]. Под малой рекой, согласно ГОСТ 19 179-73 “Гидрология суши. Термины и определения”, принято понимать реку, бассейн которой располагается в одной географической зоне, и гидрологический режим ее под влиянием местных факторов может быть не свойственен для рек этой зоны. При этом площадь водосбора малой реки не должна превышать 2000 км2. Принимая во внимание разнообразие ландшафтов, по которым протекают эти реки [2], строение морских берегов [3] и источников питания данных водотоков, возникает вопрос о возможности специфических механизмов функционирования маргинального фильтра в устьях малых рек приливного моря. Для учета величины прилива мы будем использовать следующие понятия [13]: микроприливные условия – средняя величина прилива в сизигию на устьевом взморье реки не превышает 1.6 м; мезоприливные условия – средняя величина сизигийного прилива превышает 1.6 м, но меньше уровня в 2.8 м; макроприливные условия – средняя величина сизигийного прилива превышает 2.8 м.
К основным отличиям устьев малых рек от больших рек относится соизмеримость ширины русла водотока в малую воду приливного цикла с шириной приливных осушек. Это более характерно для микроприливных условий. Для мезоприливных и особенно для макроприливных условий на устьевых взморьях малых рек ширина русла водотока в малую воду обычно бывает заметно ниже размеров приливной осушки, часто на 1–2 порядка. В подобных условиях процессы смешения речных и морских вод в течение приливного цикла проходят преимущественно в пределах периодически заливаемых устьевыми водами и периодически осушаемых зон. Их донные отложения, как показывают наши наблюдения, как правило, формируются илистыми и глинистыми (илисто-глинистыми) осадками, тогда как неосыхаемое, даже в малую воду приливного цикла, ложе водотока имеет крупнозернистые осадки – пески, гравий и гальку.
Биогеохимические процессы в данной ситуации начинают полностью зависеть от природных условий таких приливных осушек, спецификой которых является наличие маршей с массовыми зарослями растений-галофитов и мощными отложениями иловых наносов. Здесь геохимические процессы, наблюдаемые при смешении речных и морских вод, которые происходят по сценарию классического маргинального фильтра, могут терять свою доминантную позицию при формировании транзита растворимых и нерастворимых веществ из реки в море. Это, в первую очередь, обусловлено влиянием гидробиологических факторов и изменением характера гидрофизических процессов (рис. 1). В рассматриваемой зоне из-за массовых зарослей макрофитов меняется режим теплообмена между атмосферой, водой и донным грунтом, резко ослабевают скорости течений, уменьшается турбулентный обмен, значительно снижается высота ветровых волн и меняется характер вертикальных движений ледяного покрова.
Рис. 1.
Суточная изменчивость солености (а), величины рН (б) и кислородонасыщения (в) в протоке в зарослях галофитов (правый столбец) и на открытой акватории (левый столбец) на устьевом взморье р. Мудьюги в Двинском заливе 27.08.2019 г.
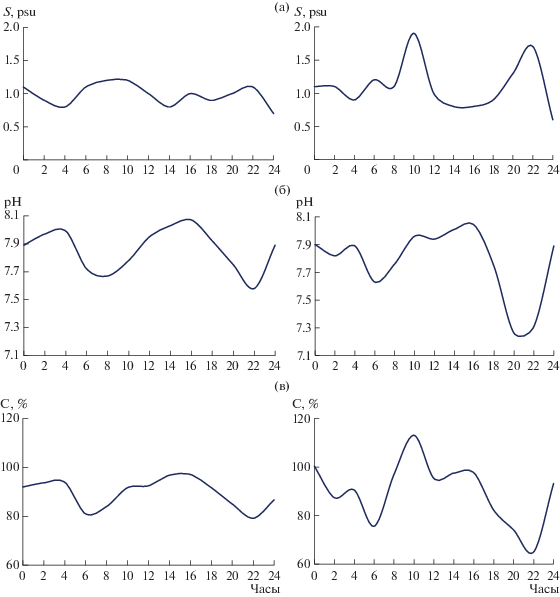
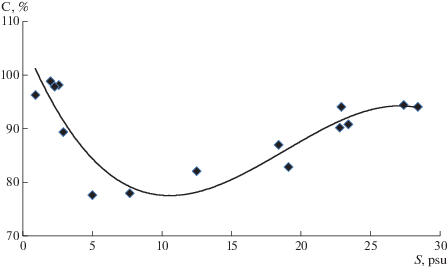
Считается, что эстуарии являются ловушкой для биогенных веществ. При этом чем выше величина прилива, тем выше его продукционный потенциал [5]. Однако, как показывают наши исследования, данное утверждение справедливо лишь в микроприливном диапазоне возрастания величины прилива. При ее дальнейшем увеличении наблюдается нелинейное возрастание интенсивности размыва мелкодисперсных наносов, сопровождающееся значительным замутнением устьевых вод [10]. Если ориентироваться на кислородонасыщение воды как на индикатор интенсивности процессов фотосинтеза, то перенасыщение устьевых вод до 110–140% и более в вегетационный период характерно для микроприливных условий. Заметим, что вне рамок вышеуказанного периода перенасыщение устьевых вод, как правило, не превышает 100–103%. В эстуариях с макроприливными параметрами оно обычно не бывает больше 75–98% (рис. 2).
Рис. 2.
График связи кислородонасыщения вод с распределением солености вод эстуария р. Чеши в Чешской губе Баренцева моря 23.07.2018 г.
Другим фактором, показывающим регламентирующее влияние приливов на функционирование водной экосистемы, является возможность возникновения в северных (арктических) условиях эвтрофикации в микроприливных устьях рек с наличием массовых зарослей галофитов. С другой стороны, с возрастанием величины прилива в ее мезо-макроприливном диапазоне вероятность возникновения эвтрофикации резко снижается.
В вегетационный период заросли галофитов служат мощным источником появления в устьевых водах соединений углерода (органических веществ) за счет усвоения ими атмосферного углекислого газа. Они также способны генерировать поступление в водную среду органического азота за счет деятельности симбиотических микроорганизмов, образующих сообщества с высшими растениями галофитами [12, 19]. Данные микроорганизмы усваивают молекулярный азот из атмосферы, переводя его в соединения, доступные для водных растений. При этом потоки данных органических веществ из устьевых вод в море способны превышать поступление в устье малой реки органики и общего азота с речными водами.
Таким образом, приливные осушки с развитой галофитной растительностью в устьях малых рек могут заметно влиять на гидрологические, гидрохимические и гидробиологические условия. Массовые заросли макрофитов здесь могут формироваться как в начале зоны смешения речных и морских вод, так и в ее середине. Это зависит не только от солености, но и от таких факторов, как ширина приливной осушки и ее защищенность от ветрового волнения и ледовой эрозии.
Здесь необходимо также учитывать, что растительные сообщества галофитов очень разнообразны по морфометрическим параметрам, что накладывает соответствующий отпечаток на характер их влияния на устьевую экосистему, в частности на процессы заиливания приливных осушек [14]. При этом максимальные концентрации фитопланктона и зоопланктона в пространственном отношении за счет формирования благоприятных гидрологических условий для планктона в целом совпадают друг с другом, в отличие от модели маргинального фильтра, где они разнесены по пространству [6].
Необходимо отметить, что при интенсивном развитии эвтрофикации, в годы с жарким летом не исключается гибель донных беспозвоночных в малую воду приливного цикла, когда содержание кислорода может снижаться ниже уровня 2 мг/л. Для арктических морей подобная ситуация является маловероятной, исключением в этом отношении являются южные заливы Белого моря.
Таким образом, биологическая пробка в структуре маргинального фильтра в приливном устье малых рек не замыкает цепочку его активных геохимических фрагментов, а может локализоваться в зарослях водных макрофитов в начале или в середине зоны солоноватых вод.
Массовое развитие галофитов в устьевом водотоке может заметно замедлять процесс смешения речных и морских вод. Это обусловлено ограничением на развитие размеров вихрей и замедлением скоростей течений, определяющих процессы турбулентного перемешивания разнородных водных масс. В данной ситуации эстуарный геохимический барьер смещается к устьевому взморью малой реки, тогда как в устьях более крупных рек в меженных условиях он обычно локализуется на нижнем отрезке устьевого участка реки.
В условиях тундровых ландшафтов, особенно при интенсивном протаивании мерзлых грунтов с очень низкими величинами рН (3.8–5.8) [7], смена кислотных свойств на щелочные при ослаблении турбулентного перемешивания речных и морских вод также будет происходить на устьевом взморье малой реки. Это в значительной мере усиливает интенсивность геохимических процессов в вышеуказанной зоне устья реки, в первую очередь связанных с выпадением в осадок металлов, имеющих высокую миграционную способность при низких величинах рН. Данная ситуация наиболее характерна для железа [18], а также алюминия.
В итоге получаем, что в устьях малых рек приливного моря фрагменты маргинального фильтра (мутьевая, химическая и биологическая пробки) могут занимать различные пространственные позиции. Для устьев более крупных рек подобное нетипично.
Следует отметить, что распространять полученные выводы на устья рек неприливных морей, а также на водотоки с малой шириной приливной осушки (не более 1–2 м) на скалистых побережьях приливных морей, по нашему мнению, некорректно. Например, в некоторых устьях рек Кандалакшского залива Белого моря [8] и в устьях рек Северного острова архипелага Новая Земля с наличием очень узких зон приливных осушек на скалистых берегах в функционировании их экосистем наблюдается соблюдение классической модели маргинального фильтра.
Необходимо обратить внимание на еще один аспект рассматриваемой проблемы. В условиях арктических морей с микроприливными условиями значительное развитие получают опресняемые лагуны. Они, в частности, широко представлены на таких островах, как Колгуев, Вайгач, и на архипелаге Новая Земля [15]. В такие лагуны, поперечные размеры которых могут достигать нескольких километров, обычно впадает 1–3 малые реки, но значительная мелководность лагун, как правило, вызывает их заметное распреснение в безледоставный период. Этому также способствует ограниченный обмен устьевых лагун с морскими водами, который происходит через узкие проливы. Их расширению препятствует здесь накопление в виде продольных валов (кос) отложений, сформированных наносами большой гидравлической крупности (галька, гравий, крупнозернистый песок). На этот процесс на локальном участке около выхода лагунного пролива в море большое влияние оказывает взаимодействие приливов и ветра. При их разнонаправленности происходит существенное изменение параметров ветровых волн, в первую очередь крутизны [4], что влечет за собой возрастание вертикальной скорости воды, вовлеченной в движение по волновой орбите. При обрушении волн возникают разрывные течения с большими скоростями течений – до 1 м/с, вызывающие накопление крупнозернистого материала около лагунного пролива. Подобный материал начинает поддерживать его устойчивость при малом поперечном сечении к размыву при прохождении штормов и весенних паводков на реках. Лагуны в данной ситуации превращаются в ловушку для взвесей, выносимых малыми реками в море, и начинают постепенно заболачиваться, в том числе за счет разрастания водных макрофитов. Данный процесс растягивается на многие десятилетия из-за малых величин стока рассматриваемых рек, но из-за наблюдаемого потепления в европейской Арктике в последние годы он может усилиться. Отметим, что в условиях, когда источником питания малых рек являются ледники, лагунные устья рек могут накапливать редкоземельные металлы и другие, даже очень высокотоксичные вещества, депонированные в ледниках за счет аэрозольного переноса в течение продолжительного периода времени. В данной ситуации, например, не исключается возникновение геохимических аномалий в лагунах Северного острова архипелага Новая Земля, за счет повышенных потоков переноса ряда тяжелых металлов над акваторией Баренцева моря [17]. Таким образом, устья некоторых малых рек могут “замыкать” маргинальный фильтр, препятствуя при разрушении многолетнемерзлых грунтов переносу веществ с талыми водами с территории суши в арктические моря. Учитывая, что водотоки всех арктических архипелагов и островов в пределах Российской Федерации имеют статус малых рек, исследованиям данной проблемы необходимо уделить особое внимание.
Мелководность устьевых взморьев малых рек и наличие на них обширных приливных осушек обуславливает периодическое (на квадратуре) опускание и примерзание льда ко дну водотока практически на всей его площади. Это может кардинально менять пространственный масштаб функционирования маргинального фильтра. Подразумевается, что его протяженность в зоне влияния вод устьев малых рек составляет сотни метров [6]. Однако весной лед с примерзшим к нему верхним слоем донных отложений под действием ветра может выноситься из устья малой реки в море на большие расстояния (километры–десятки километров). В данной ситуации конфигурация и протяженность маргинального фильтра начинают определяться вероятностными характеристиками ветрового режима в весенний период. Он по протяженности может становиться сопоставимым с маргинальными фильтрами средних и больших рек, но интенсивность накопления в зоне его действия осадков должна быть относительно слабой. Исключение могут составлять те устья малых рек, на водосборах которых имеются месторождения полезных ископаемых и ведется их разработка. Отметим, что этот механизм переноса взвесей не работает в лагунных устьях рек из-за ограниченности ледообмена устьевых вод с морскими водами через узкие проливы.
Мелководность устьевых взморьев некоторых малых рек, где заросли галофитов не могут получить массового развития из-за высокой солености и влияния ветрового волнения, обуславливает формирование в их зонах повышенных концентраций взвешенных веществ из-за постоянного взмучивания илов за счет воздействия сильных приливных течений. В данной ситуации получаем возрастание содержания взвесей по мере увеличения солености устьевых вод, что противоречит схеме классического маргинального фильтра.
Можно сделать вывод, что в устьях малых рек приливного моря в зависимости от условий развития приливных осушек, галофитной растительности, их глубин и конфигурации береговой черты пространственная структура маргинального фильтра может кардинально отличаться от таковой в устьях больших и средних рек. С этой точки зрения, устье каждой малой реки следует рассматривать как уникальный водный объект со своим специфическим набором геохимических и других природных процессов, что необходимо учитывать при проведении их прикладных исследований [11]. Подобное утверждение, в первую очередь, справедливо для устьев рек западного сектора российской Арктики (Белое, Баренцево и Карское моря), располагающихся в регионе с ярко выраженными пространственными градиентами климатических и природных характеристик.
Источники финансирования. Работа выполнена в рамках темы госзадания № 0149-2019-0007 “Современные и древние донные осадки и взвесь Мирового океана – геологическая летопись изменений среды и климата: рассеянное осадочное вещество и донные осадки морей России, Атлантического, Тихого и Северного Ледовитого океанов – литологические, геохимические и микропалеонтологические исследования; изучение загрязнений, палеообстановок и процессов в маргинальных фильтрах рек”.
Список литературы
Гордеев В.В. Геохимия системы река–море. М.: ИП Матушкина И.И., 2012. 452 с.
Исаченко А.Г., Шляпников А.А. Ландшафты. М.: Мысль, 1989. 504 с.
Каплин П.А., Леонтьев О.К., Лукьянов С.А., Никифоров Л.Г. Берега. М.: Мысль, 1991. 479 с.
Коробов В.Б., Лавренов И.В. Оценка влияния приливных течений на функции распределения высот ветровых волн // Метеорология и гидрология. 1989. № 11. С. 73–80.
Лисицын А.П. Лавинная седиментация // Лавинная седиментация в океане. Ростов: Изд-во Ростовского университета, 1982. С. 3–59.
Лисицын А.П. Маргинальный фильтр океанов // Океанология. 1994. Т. 34. № 5. С. 735–747.
Лобова Е.В., Хабаров А.В. Почвы. М.: Мысль, 1983. 303 с.
Лукашин В.Н., Кособокова К.Н., Шевченко В.П. и др. Результаты комплексных океанографических исследований в Белом море в июне 2000 г. // Океанология. 2003. Т. 43. № 2. С. 237–253.
Мискевич И.В. Специфика природных процессов в приливных устьях малых рек Белого моря // Проблемы региональной экологии. 2019. № 4. С. 37–41.
Мискевич И.В., Алабян А.М., Коробов В.Б. и др. Исследования короткопериодной изменчивости гидролого-гидрохимических характеристик устья реки Кянда в Онежском заливе Белого моря (28 июля–15 августа 2016 г.) // Океанология. 2018. Т. 58. № 3. С. 369–373.
Мискевич И.В., Коробов В.Б., Алабян А.М. Специфика инженерно-экологических изысканий в приливных устьях малых рек западного сектора российской Арктики // Инженерные изыскания. 2018. Т. 12. № 3-4. С. 50–61.
Мискевич И.В., Чульцова А.Л., Мосеев Д.С. Марши мезо-макроприливных эстуариев рек арктической зоны как источник растворенного органического азота для водных экосистем // Естеств. тех. науки. 2018. № 4. С. 75–81.
Михайлов В.Н. Принципы типизации и районирования устьевых областей рек (аналитический обзор) // Водные ресурсы. 2004. Т. 31. № 1. С. 5–14.
Мосеев Д.С. Динамика развития маршей на юге Онежского залива Белого моря под влиянием галофитной растительности (на примере эстуария р. Кянда) // Материалы XXIII Международной научной конференции (Школы) по морской геологии. Т. III. Москва. 2019. С. 202–206.
Школьный Д.И., Айбулатов Д.Н. Типизация устьевых областей рек острова Южный (архипелаг Новая Земля) // Вестн. Московского ун-та. Сер. 5. География. 2016. № 6. С. 50–58.
Lisitzin A.P. Oceanic sedimentation: lithology and geochemistry. Washington: American Geophysical Union, 1996. 390 p.
Lisitzin A.P., Shevchenko V.P. Glacial-marine sedimentation // Encyclopedia of Marine Geosciences / Harff J., Meschede M., Petersen S., Thiede J. (eds.). Dordrecht: Springer Science + Business Media, 2016. P. 288–294.
Pokrovsky O.S., Schott J. Iron colloids/organic matter associated transport of major and trace elements in small boreal rivers and their estuaries (NW Russia) // Chem. Geol. 2002. V. 190. P. 141–179.
Teske A., Durbin A., Ziervogel K. et al. Microbial community composition and function in permanently cold seawater and sediments from an Arctic Fjord of Svalbard // Appl. Environ. Microbiol. 2011. V. 77. № 6. P. 2008–2018.
Дополнительные материалы отсутствуют.