

Океанология, 2021, T. 61, № 4, стр. 609-619
Палеоокеанология Азовского моря в голоцене (по данным бурения и изучения малакофауны на косе Долгой)
Г. Г. Матишов 1, 2, *, В. В. Польшин 1, Е. П. Коваленко 1, К. С. Григоренко 1
1 Федеральный исследовательский центр Южный научный центр РАН
Ростов-на-Дону, Россия
2 Мурманский морской биологический институт Кольского научного центра РАН
Мурманск, Россия
* E-mail: matishov_ssc-ras@ssc-ras.ru
Поступила в редакцию 18.09.2020
После доработки 11.10.2020
Принята к публикации 19.11.2020
Аннотация
В статье представлены результаты исследований, проводимых сотрудниками ЮНЦ РАН на косе Долгой в 2019–2020 гг. Анализируются данные космоснимков, измерения прибрежных течений, бурения скважин, изучения малакофауны и определения абсолютного возраста береговых отложений. В соответствии с простиранием древних береговых валов, последовательностью их причленения друг к другу выделяются шесть геоморфологических генераций, отражающих ход нимфейской и отчасти более ранней трансгрессий. Данные радиоуглеродного датирования раковин моллюсков позволяют предположить, что основная часть аккумулятивного тела косы была образована за одну тысячу лет, в интервале 2.5–1.5 тыс. л.н. По результатам натурных наблюдений показано, что при слабом восточном ветре скорость течений у западного берега косы в 2–3 раза выше, чем у восточного. Приводится список, включающий порядка 20 видов и надвидовых таксонов моллюсков из береговых отложений косы Долгой. Отмечается, что основную массу (более 90%) ракушечных отложений, слагающих береговые валы, составляют створки раковин Cerastoderma glaucum.
ВВЕДЕНИЕ
Азовское море на 30% представлено мелководьями с глубинами до 3 м. До сих пор нет четких представлений о развитии этого морского водоема в конце плейстоцена и в голоцене. Существуют расхождения во взглядах, касающиеся количества, амплитуды и продолжительности регрессивно-трансгрессивных циклов [1, 2, 5, 6, 8, 9, 12, 14, 20, 22, 27, 29, 30, 32–36, 41, 43, 44]. Среди характерных геоморфологических образований северного, западного и восточного побережий выделяются азовские косы. Осадочная толща кос сформирована преимущественно из створок моллюсков и ракушечного песка [3, 4, 11, 15, 19, 39].
Единственной в своем роде является коса-стрелка Долгая. Ее строение и история развития остаются неясны и требуют уточнения. За всю историю изучения косы Долгой не была опубликована характеристика отложений из карьеров по добыче ракушечного сырья, не выполнялось целенаправленное бурение, не выяснены конкретные источники и пути переноса раковин моллюсков. В ходе работы нам предстояло восстановить палеогеографическую обстановку в морском бассейне, определить таксономический состав малакофауны из отложений косы Долгой, выяснить предпосылки, способствовавшие формированию высокой биопродуктивности донных организмов и воссоздать оптимальную среду обитания для Cerastoderma glaucum Poiret, 1789 – ключевого осадкообразующего вида бентоса.
Цель работы заключалась в том, чтобы, опираясь на геоморфологический анализ морского дна, данные бурения скважин и исследование малакофауны, получить новые представления о палеоокеанологии Азовского моря в голоцене и реконструировать историю развития косы Долгой.
МАТЕРИАЛЫ И МЕТОДЫ
С 2016 г. ЮНЦ РАН проводит планомерное бурение на побережье Таганрогского залива, а именно на косах: Очаковской, Чумбурской, Беглицкой и Долгой [20, 21].
В 2019–2020 гг. нами было пробурено 7 скважин с отбором керна в корневой, центральной и дистальной частях косы Долгой. В 2019 г. было пробурено 2 скважины глубиной 11 м. Глубина выработки скважин, пробуренных в 2020 г., составила: 1 – 19.40 м; 2 – 26.50 м; 3 – 25.50 м; 4 – 20.00 м; 5 – 21.50 м. Кроме этого были изучены 10 обнажений (мощностью от 1.7 до 2.5 м), расположенных в карьерах по добыче ракушечного сырья (рис. 1).
Рис. 1.
Районы экспедиционных исследований ЮНЦ РАН в 2019–2020 гг.: 1 – точки отбора проб; 2 – остров; 3 – станции измерения течений Aanderraa RCM9LW (макс. скорость, см/с); 4 – скорость и направление течений, 5 см/с; 5 – бурение 2020 г.; 6 – бурение 2019 г.; 7 – малакофауна; 8 – маршруты ревизий древних береговых валов; 9 – геоморфологические районы (стадии развития косы Долгой).
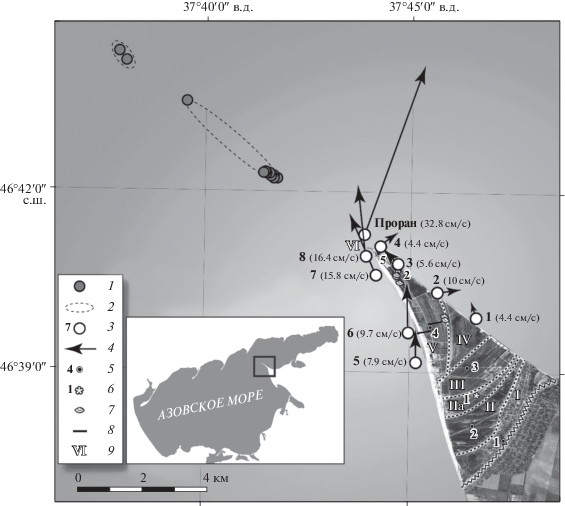
В прибрежных районах моря колонки донных осадков отбирались гравитационной трубкой с НИС “Денеб” и “Профессор Панов”, а также вибрационной трубкой с несамоходного понтона. Особое внимание уделялось отбору раковинного материала по вертикали изученных разрезов. Отбор проб из обнажений, кернов скважин и грунтовых колонок производился послойно, с описанием видовой принадлежности, степени сохранности и фотографированием раковинного материала. В результате в ракушечных отложениях аккумулятивного тела косы было выявлено порядка 20 видов и надвидовых таксонов моллюсков (табл. 1).
Зообентос отбирался в прибрежных районах с поверхностного горизонта дна с помощью дночерпателя Ван-Вина. Пробы бентоса разбирались на месте и фиксировались 70%-ным этанолом. Организмы, попавшие в крупную фракцию (>0.5 см), определяли в живом виде.
В комплекс работ входила закладка контрольных точек геодезическим GNSS приемником “EFТ M4 GNSS” для фиксации точек бурения и привязки аэрофотосъемки, а также измерения течений на участках акватории, прилегающей к косе Долгой. Используемый нами доплеровский измеритель течений Aanderaа RCM 9LW с дискретность измерений 5 мин был укомплектован датчиками температуры и солености. Для сравнения полученных результатов были привлечены данные предыдущих съемок Южного научного центра РАН [18, 19].
Абсолютный возраст отложений определялся радиоуглеродным методом по образцам раковин моллюсков из керна скважин и обнажений в лаборатории геоморфологических и палеогеографических исследований полярных регионов и Мирового океана им. В.П. Кёппена (Санкт-Петербургский государственный университет). Значения календарного возраста были получены с помощью программы “OxCal 4.3” (калибровочная кривая “IntCal 13”).
РЕЗУЛЬТАТЫ
Гидрология. Для современного Азовского моря характерно существование течений, направленных с обеих сторон косы Долгой к ее дистальной части. По результатам экспедиции, проводимой летом 2020 г., было установлено, что в условиях слабого восточного ветра (1–3 м/с) вдольбереговые течения с обеих сторон косы Долгой направлены от основания к ее оконечности. При этом течения у западного берега косы в 2–3 раза сильнее (10–15 см/с), чем у восточного. У восточного берега, в средней части косы, течения имеют разнонаправленный характер и к дистальной части усиливаются до 4–6 см/с. В узостях между островами косы течения имеют максимальные скорости (более 32 см/с) (рис. 1). Даже при слабом ветре из Азовского моря в Таганрогский залив скорость водотока сохраняется. Очевидно, что и на ранних стадиях формирования аккумулятивного тела косы основной поток наносов осуществлялся из юго-восточного района моря [28].
Геоморфология голоцена. Исходя из анализа космических снимков, коса Долгая представляет собой чередование валов и ложбин, фиксирующих стадии последовательного выдвижения аккумулятивной формы в море [45]. Общая протяженность системы коса–острова – более 20 км. Длина собственно косы составляет более 10 км, а ширина ее основания около 4 км. Высота береговых валов над уровнем моря достигает 2.5 м.
В соответствии с простиранием древних береговых валов и лагун, последовательностью их причленения друг к другу выделяются шесть геоморфологических генераций, отражающих осцилляции нимфейской и отчасти более ранней трансгрессий.
Первая стадия развития косы Долгой соотносится с образованием прикорневой террасы вдоль обрыва коренного берега – по возрасту самая ранняя, вероятно 6–7 тыс. л.н. [3].
Вторая стадия – стадия лиманов и лагун, выражена в рельефе системой из порядка 30 вытянутых впадин. Абсолютное превышение перемычек, разделяющих лагуны, составляет 0.6–0.8 м Балтийской системы высот (БСВ). Лагуны возникли на низком стоянии уровня Азовского моря. В периоды высокого стояния уровня, при западных штормах эта часть косы затапливается, что фиксировалось в 1914, 1960, 1969 и 1970 гг. В условиях же низкого стояния уровня лагуны полностью осушались, давая толчок к возникновению растительности [3].
Третья и четвертая стадии возникновения береговых валов отражают различные фазы нимфейской трансгрессии (2.4–1.5 тыс. л.н.). Определенный по 14С возраст моллюсков из ракушечных валов составил 1670–2300 л. [3]. Коса Долгая в это время была гораздо короче и не являлась стрелкой в геоморфологическом смысле.
Пятая стадия развития косы Долгой относится к историческому времени (0.5–0.1 тыс. л.н.). В это время она развивалась по типу косы-стрелки. Современную литодинамику отражает шестая стадия развития косы. С 1890 по 2000 гг. прослеживалась устойчивая тенденция уменьшения длины оконечности косы. Этот процесс интенсивно происходил с 1890 по 1958 гг., когда протяженность косы уменьшилась на 1.35 км. За период с 1964 по 1966 гг. оконечность отступила на юг на 0.75 км, а за 2006–2016 гг. – еще на 50–60 м [3, 4].
К этой же стадии развития косы можно отнести возникновение гряды островов и отмелей, протягивающихся на 12 км к северо-западу от ее дистальной оконечности. В литературе нет описаний этих образований. В ходе экспедиционных исследований нами был выполнен пеший и водный (на маломерном судне) осмотр, аэрофотосъемка, а также заложен шурф глубиной 1.6 м на одном из островов. В результате было установлено, что численность песчаных островов (у одного из них координаты 46°44′14.6″ с.ш. и 37°37′51.9″ в.д.) постоянно изменяется от 15 до 20, в зависимости от штормовых сгонно-нагонных явлений. Островная гряда возвышается над уровнем моря в среднем на 1–2.5 м. Три самых высоких острова имеют длину до 1 км. Сложены острова ракушечным детритом и целыми створками раковин.
Бурение. Геологические работы. Бурение, проведенное в 2019–2020 гг., позволило нам получить представление о строении и литологии отложений косы Долгой. Точки бурения в 2020 г. намечались в соответствии с пятью предварительно обозначенными геоморфологическими районами: скв. 1 − на верхней террасе в 50 м от обрывов коренного берега, скв. 2 − на осушке лагуны, скв. №№ 3 и 4 (координаты 46°38′49.4″ с.ш., 37°46′35.1″ в.д. и 46°39′33.5″ с.ш., 37°45′37.3″ в.д. соответственно) были пробурены на двух береговых валах, а скв. № 5 − на дистальном окончании косы (рис. 1).
Вскрытая скважинами верхняя часть отложений косы Долгой повсеместно сложена ракушечным материалом с примесью глины и суглинка. Мощность ракушечных отложений увеличивается от корневой к дистальной части косы с 4 до 7 м (рис. 2). Мористее оконечности косы залегают новоазовские отложения, представленные песчано-ракушечными осадками, мощность которых может достигать 10 м [41].

В ракушечных отложениях по вертикали разреза отмечается резкое преобладание створок раковин и детрита моллюсков рода Cerastoderma Poli, 1795 (более 95%). Значительно реже встречаются Mytilus galloprovincialis Lamarck, 1819, Chamelea gallina (Linné, 1758), Abra segmentum (Récluz, 1843), Gastrana fragilis (Linné, 1758) и раковины Rissoa sp. и Tritia reticulata (Linné, 1758). Цвет ракушечных отложений меняется от белого и светло-желтого (в верхней части) до серо-черного (в нижней части). Верхняя часть ракушечной толщи уплотнена, а средняя сильно обводнена.
Уровень безнапорных грунтовых вод в районах бурения повышается от корневой к дистальной части косы от 3.5 до 1.6 м. По всей площади косы под ракушечными наносами с прослоями иловатой глины и суглинка прослеживаются коричневые тугопластичные суглинки с включениями гидроокислов Fe, Mn и карбонатов. Местами в суглинках отмечаются прослои мелкозернистого обводненного песка. В нижней части разрезов скважинами №№ 2, 4 и 5 вскрыты обводненные мелкозернистые пески коричневого цвета. Отмечено ожелезнение песчаных зерен по поверхности и редкие прослои тугопластичной глины. В песках, отобранных из низов керна скв. 4, присутствуют включения раковинного материала до 10% от объема отобранной пробы. Здесь массово отмечаются раковины моллюсков: Viviparus sp., Bittium sp., Lithoglyphus sp., Planorbis sp., Dreissena sp., Theodoxus sp., Cerastoderma sp. и Abra sp. Найденные совместно с ними зубы Arvicola cf. mosbachensis и Microtus cf. gregalis (примитивный морфотип) позволяют коррелировать эти слои с началом среднего неоплейстоцена (определения А.С. Тесакова, ГИН РАН).
Исследование малакофауны. В настоящее время ключевую роль в образовании карбонатных осадков в Азовском море играет моллюск Cerastoderma glaucum, характеризующийся высокой продуктивностью и прочностью створки, а также Abra segmentum и Mytilaster lineatus (Gmelin, 1790) [7, 37]. Заметное влияние на этот процесс также оказывают Mytilus galloprovincialis и вселенцы Mya arenaria Linné, 1758 и Anadara kagoshimensis (Tokunaga, 1906). Общая годовая продукция зообентоса в Азовском море оценивается в 19–20 млн т, в том числе продукция Cerastoderma glaucum составляет 13–14 млн т в год, достигая биомассы 2 кг/м2 [7, 13].
При изучении обнажений на косе Долгой нами было отмечено, что ракушечные отложения преимущественно состоят из раковин моллюска Cerastoderma glaucum разной степени сохранности (табл. 1).
Таблица 1.
Малакофаунистический состав береговых отложений косы Долгой
| Таксон | % от общего количества створок и раковин в отложениях | ||||
|---|---|---|---|---|---|
| 0.5 м | 1.0 м | 1.5 м | 2.0 м | 2.5 м | |
| радиоуглеродный возраст раковин моллюсков | |||||
| 1590 ± 100 | 2420 ± 120 | 2300 ± 130 | 2110 ± 110 | 1960 ± 90 | |
| Cerastoderma glaucum | 96.14 | 90.82 | 93.57 | 80.83 | 76.54 |
| Bittium reticulatum | 1.51 | 6.01 | 2.75 | 10.25 | 13.14 |
| Pholadidae gen. sp. | 0.00 | 0.52 | 0.47 | 2.37 | 3.11 |
| Tritia reticulata | 1.51 | 0.94 | 1.18 | 1.33 | 1.14 |
| Hydrobia sp. | 0.00 | 0.00 | 0.16 | 2.47 | 1.14 |
| Chamelea gallina | 0.17 | 0.34 | 0.86 | 0.66 | 1.67 |
| Abra segmentum | 0.00 | 0.17 | 0.24 | 0.09 | 1.14 |
| Theodoxus sp. | 0.00 | 0.86 | 0.63 | 0.47 | 0.76 |
| Mytilidae gen. sp. | 0.67 | 0.00 | 0.08 | 0.38 | 0.76 |
| Lentidium mediterraneum | 0.00 | 0.09 | 0.00 | 0.38 | 0.23 |
| Rissoa sp. | 0.00 | 0.09 | 0.08 | 0.19 | 0.30 |
| Abra sp. | 0.00 | 0.00 | 0.00 | 0.47 | 0.00 |
| Hydrobia acuta | 0.00 | 0.17 | 0.00 | 0.00 | 0.00 |
| Cylichnina sp. | 0.00 | 0.00 | 0.00 | 0.09 | 0.00 |
| Raphitomidae gen. sp. | 0.00 | 0.00 | 0.00 | 0.00 | 0.08 |
| Общее количество створок Bivalvia и раковин Gastropoda | 596 | 1165 | 1275 | 1054 | 1317 |
В дистальной части косы исследовались обнажения в стенках карьеров по добыче ракушечного сырья. Вся толща отложений представляет собой чередование слоев хорошо сохранившихся раковин с прослоями раковинного детрита. Цвет отложений – от светло-желтого до желто-коричневого. Нижние горизонты обводнены, ракуша окрашена в серый и черный цвета и имеет характерный запах сероводорода.
К основанию изученных обнажений в отложениях отмечается увеличение содержания более крупных и массивных створок раковин руководящего вида Cerastoderma glaucum. В виде прослоев и единичных включений также были отмечены крупные, хорошо сохранившиеся, створки Chamelea gallina. Кроме того, на разных глубинах встречаются раковины и обломки Theodoxus sp., Solen vagina Linné, 1758, Bittium reticulatum (Costa, 1799), Tritia reticulata, Pholadidae gen. sp. и сильно разрушенные обломки Mytilidae gen. sp. В отложениях, отобранных на глубине 2.5 м от дневной поверхности, отмечено резкое увеличение количества Bittium reticulatium (до 13.14%) при сокращении доли Cerastoderma (до 76.5%) (табл. 1). Этот морской моллюск обитает на водной растительности и илисто-песчаных грунтах до глубины 20 м и выносит опреснение до 10‰ [40].
Ракушечные отложения центральной части косы в основном сложены имеющими различный размер и толщину целыми и слаборазрушенными створками раковин Cerastoderma glaucum с примесью песчано-илистых частиц. Отмечается переслаивание слоев, преимущественно сложенных целыми створками раковин, с маломощными прослоями, в составе которых преобладает ракушечный детрит. В основании расчистки встречаются крупные раковины Chamelea gallina. На разных горизонтах отмечены единичные раковины Barnea candida (Linné, 1758), Abra segmentum, Lentidium mediterraneum (Costa, 1829), Bittium reticulatum, Tritia reticulata, Rissoa sp., Hydrobia sp. и Theodoxus pallasi Lindholm, 1924.
Исследование островов у оконечности косы Долгой показали, что их толща сложена в основном хорошо сохранившимися створками Cerastoderma glaucum. В меньшем количестве встречаются преимущественно скрепленные раковины Mytilaster lineatus. Много створок моллюсков-вселенцев Mya arenaria и Anadara kagoshimensis, впервые отмеченных в Азовском море в середине прошлого века. На поверхности островов, кроме вышеперечисленных, отмечены створки Solen vagina, Gastrana fragilis, Chamelea gallina, Mytilus galloprovincialis, единичные раковины Bittium reticulatum, Hydrobia sp., свежая раковина Viviparus viviparus (Linné, 1758), на отдельных створках отмечены раковины рака Balanus improvisus Darwin, 1854. Наличие в пробе недавних вселенцев, створок мидий, скрепленных лигаментом, позволяют предположить, что верхняя часть островов сложена современным раковинным материалом.
Таким образом, основную массу ракушечных отложений, образующих аккумулятивное тело косы Долгой, составляют раковины эвригалинных видов Cerastoderma glaucum (содержание в отложениях более 90%), Abra segmentum, Hydrobia sp., способных выносить опреснение до 5‰. Присутствие в отобранных пробах створок умеренно стеногалинного моллюска Gastrana fragilis, который в настоящее время обитает в Черном море, может свидетельствовать о периодическом увеличении солености водоема, а наличие раковин солоноватоводного брюхоногого моллюска рода Theodoxus Monfort, 1810, способного обитать при солености не более 5–7‰ – на его опреснение. Также в исследованных пробах в большом количестве были встречены раковины Barnea candida, образующие устойчивые биоценозы на плотных и твердых грунтах [10, 26].
Предварительный анализ малакофауны из исследованных обнажений, подкрепленный полученными данными абсолютного датирования возраста (по 14С), позволяет предположить, что основная часть аккумулятивного тела косы была образована в интервале времени от 1920 ± 110 (ЛУ-9756) до 2500 ± 150 (ЛУ-9757) лет, во время нимфейской трансгрессии (табл. 1).
Мелководье банок на востоке моря. Важное место в формировании биопродуктивности Азовского моря занимает прибрежный шельф в 100-километровой зоне к западу от Приморска-Ахтарска и Должанской косы, имеющий ряд отличительных геоморфологических черт. Здесь широко развиты крупные подводные гряды – банки Еленина, Железинская, Ахтарская и Ачуевская, длина которых достигает 35–50 км (рис. 3) [16].
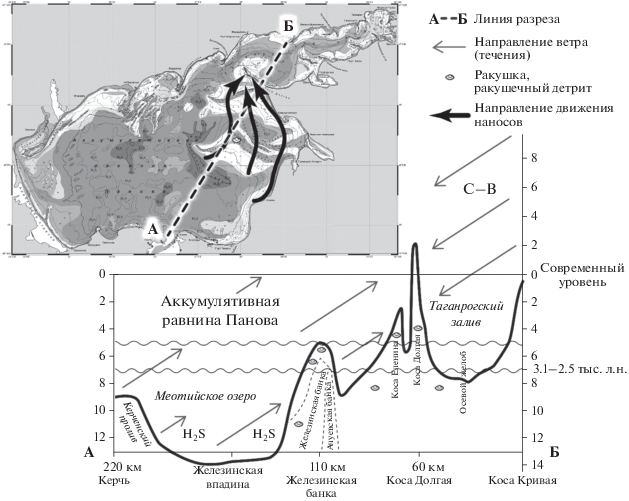
Глубины над вершинами банок минимальные, возрастают от 3 до 7 м, а над банкой Еленина составляют 1–2 м. Высота гряд по отношению ко дну прилегающих ложбин варьирует от 2 до 5 м. На вершинах распространены самые значительные для открытого моря площади песчано-ракушечных отложений (фракция 1.0–0.1 мм >70%), а на подводном склоне банок залегают илистые пески с содержанием фракции 1.0–0.1 мм в пределах 50–70% (рис. 3). Во многих местах донные осадки представляют собой заиленный ракушечник. Железинская и Ахтарская банки разделены между собой сравнительно узкой ложбиной, заполненной илистыми отложениями со значительным включением раковинного материала [16, 17, 31]. По периферии Железинской банки возраст отложений в интервалах 80–90 и 210–220 см составил 5900 ± 140 л.н. (ЛУ – 6799) и 6480 ± 120 л.н. (ЛУ – 7047) соответственно [23].
Опираясь на данные о структуре донных сообществ Азовского моря и закономерностях их распределения, представленные в наших ранних публикациях [24–26, 42], можно отметить, что зонами современного карбонатонакопления остаются заливы Северного Приазовья, район Арабатской стрелки и банок Еленина, Ахтарской и Железинской [17, 38]. Ряд отличительных черт этих районов способствуют формированию благоприятных условий для развития здесь высокопродуктивных бентосных сообществ.
Проведенные исследования показали, что центральную роль в формировании донных сообществ в районах с пересеченным рельефом дна играют два вида моллюсков – Mytilaster lineatus и Cerastoderma glaucum. В распространении зообентоса в обозначенном районе прослеживаются четкие закономерности. Сообщества Mytilaster населяют гребни банок и с увеличением глубины сменяются сообществом Cerastoderma, которое спускается к ложбинам у основания банок, окантовывает их и проникает внутрь расчлененных участков шельфа.
Mytilaster lineatus формирует богатые органикой друзовые сообщества с высокими показателями биомассы, достигающими максимального значения на Железинской банке – 5 кг/м2.
Сообщество Cerastoderma в основном приурочено к алевритовым и алевритово-глинистым илам. В его состав входят пелофильные виды: Hydrobia acuta (Draparnaud, 1805), Nephthys hombergii Savigny in Lamarck, 1818, Neanthes succinea (Leuckart, 1847), биомасса зообентоса не превышает 300 г/м2 [24].
Современный геоморфологический и литологический облик отмеченных мелководий глубиной 1–7 м, расположенных на вершинах банок, создавался в течение последних 2 тыс. лет в ходе нимфейской трансгрессии. В фанагорийскую регрессию или в античное время уровень моря опускался на 6–7 м ниже современного его положения [20]. При этом вся акватория Таганрогского залива, Железинская и другие банки осушались, и вышедший на поверхность участок дна подвергался воздействию денудации.
ОБСУЖДЕНИЕ
Особенности развития косы Долгой. Феноменальным по размеру представляется ракушечное тело косы Долгой: 20 км × 4 км × 4–7 м (длина, ширина, толщина). О масштабе аккумуляции раковинного материала свидетельствует объем его промышленного изъятия (порядка 1.451 млн т с 1956 по 1970 г.) из одного из карьеров, расположенных на оконечности косы [3, 4]. Исходя из данных радиоуглеродного датирования моллюсков, период накопления колоссального объема ракуши охватывал временной промежуток в четыре тысячи лет. Однако основная часть органогенного материала косы сформировалась примерно за одну тысячу лет, в интервале 2.5–1.5 тыс. л.н.
Сочетание на косе Долгой генераций реликтовых береговых валов (порядка 150), залежей ракушняка и ракушечного детрита, видовое разнообразие моллюсков, для которых установлен радиоуглеродный возраст, господство среди видов зообентоса Cerastoderma sp., в совокупности приоткрывают неизвестные черты палеогеографии Азовского моря в голоцене.
Геоморфологические черты азовских кос сформировались в середине голоцена в условиях новочерноморской (4–6 тыс. л.н.) и нимфейской (2.4–1.5 тыс. л.н.) трансгрессий. Эти рубежи подтверждаются датировками абсолютного возраста отложений (по 14С) [20, 22]. Ракушечные образования кос Долгой, Бердянской, Обиточной в открытой части шельфа могли вступить в начальную стадию развития с момента поступления черноморских вод и затопления впадины Азовского моря около 5–6 тыс. л.н.
К концу древнеазовского этапа развития господствовали морские эвригалинные моллюски [27]. Последующее осолонение моря с одновременным поступлением пресных вод привело к массовому развитию донных биоценозов с доминированием Bivalvia Linne, 1758. Это способствовало продуцированию гигантского объема створок моллюсков, которые обеспечили формирование значительных по площади и объему аккумулятивных тел [2, 3].
В эпоху фанагорийской регрессии (Меотийское озеро) уровень моря был ниже современного на 6–7 м, а морской край дельты Дона неоднократно смещался [12, 20]. Наличие на современном шельфе площадей древней суши подтверждается нахождением античных следов ниже современного уровня моря. Артефакты отмечены в ракушечных отложениях Песчаных островов (центральная часть Таганрогского залива), на пляжах косы Долгой, к северу от Керченско-Таманского побережья [2, 3, 20].
Скважины, пробуренные в 2019–2020 гг., дополняют представления о фанагорийских озерно-морских отложениях эпохи “Меотийского озера” (3.1–2.5 тыс.л.н.). В керне, отобранном с глубины от 2 до 5 м, периодически встречались ракушечные отложения темно-серого до черного цвета с запахом сероводорода. Заражение сероводородом – особенность отложений Азовского моря, которые формировались в обстановке дефицита кислорода в условиях лагун, болот и плавней (Меотийское море) (рис. 4). Судя по видовому составу малакофауны, в этот этап развития моря соленость превышала 5–8‰ [26].

С началом нимфейской трансгрессии Меотийское озеро стало превращаться в полноценную морскую акваторию. Активное развитие береговых кос происходило, когда соленость водоема возросла до 11–12‰. В противном случае отложения кос формировали бы тонкостенные и неустойчивые к волновому воздействию раковины двустворчатых моллюсков родов Dreissena Beneden, 1835, Monodacna Eichwald, 1838, Anodonta Lamarck, 1799 и др. [2].
В целом нимфейская трансгрессия ассоциируется с накоплением отложений новоазовского возраста (возраст 3.1–0 тыс. л.н.), которые имеют повсеместное распространение. В центре моря они представлены глинистыми, алеврито-глинистыми и мелкоалевритовыми илами с маломощными (1–2, реже 5–7 см) прослоями ракуши [20, 31].
На косе Долгой вначале формировалась генерация валов у Ейского коренного выступа с простиранием на северо-восток. На Ейском побережье преобладают восточные ветры, но разгон волн невелик. Юго-западные ветры более редки, но имея значительную длину разгона, возбуждают крупные волны. Поскольку отмель косы Долгая играет роль “барьера” на входе в Таганрогский залив, у ее оконечности в проранах возникают штормовые течения, скорость которых может достигать 1–1/5 м/с [3, 19]. Такой силы водотоки способствуют активному размыву песчаных аккумулятивных тел (рис. 1).
В завершающую стадию аккумулятивные дуги косы выстраивались в северо-западном направлении. Господствовали южные ветровые течения и вдольбереговые потоки наносов. В этот же период возникшие ранее валы активно срезались течениями из Таганрогского залива. При двустороннем питании происходило выдвижение оконечности косы в открытое море и формирование стрелки косы.
Основной причиной образования и дальнейшего роста прибрежных кос явилось то, что в береговую зону непрерывно поступал интенсивно продуцируемый органогенный материал из центра Азовского моря [11, 39]. Крупнейший источник наносов для формирования надводно-подводной части косы Долгой очевидно существовал на юго-востоке азовского шельфа в районе Железинской, Ахтарской, Елениной банок. Площадь этих положительных структур составляет порядка 100 км2. Господство южных румбов в Азовском море предопределило вектор перемещения органогенных наносов в сторону косы Долгой.
ЗАКЛЮЧЕНИЕ
Несмотря на большой объем проведенных раннее исследований, вопросы формирования и трансформации кос Азовского моря продолжают оставаться актуальными и в настоящее время. Прежде всего, это относится к вопросам определения возраста этих аккумулятивных форм. Напрямую это пересекается с неопределенностью, касающейся установления границ береговой линии Азовского моря в разные периоды голоценовой истории. Зачастую мнения исследователей по этому вопросу кардинально расходятся. Как нам представляется, причина таких разночтений во многом заключается в том, что проводимые исследования, как правило, не носят комплексный и систематический характер. Между тем, наш опыт показывает, что наилучший результат получается только при совместном анализе одновременно полученных океанологических, геофизических, литологических и биостратиграфических данных. Подобный комплексный подход позволяет взглянуть на научную проблему с разных сторон, что значительно снижает вероятность ошибочной трактовки полученных результатов.
Таким образом, целенаправленное применение на азовских косах комплекса океанологических исследований и бурения в сочетании с расшифровкой по космоснимкам рельефа береговых валов, радиоуглеродными датировками моллюсков и анализом малакофауны открывает новые возможности для реконструкций палеобиопродуктивности водоема и истории голоцена Азово-Черноморского бассейна. При этом полученные результаты могут быть актуальны при освоении акватории Азовского моря. Их всесторонний анализ будет способствовать решению проблем, связанных с управлением и минимизацией экологических рисков и выработки стратегии устойчивого развития региона.
Источники финансирования. Публикация подготовлена в рамках реализации гранта РНФ № 20-17-00196.
Список литературы
Артюхин Ю.В. К геоморфологической интерпретации природных условий античной колонизации вершины Таганрогского залива // Древности Боспора. Т. 14. M.: Таус, 2010. С. 28–38.
Артюхин Ю.В. Факторы изменчивости природных условий существования греческих колоний на берегах Меотийского озера и Киммерийского Боспора в позднеантичную эпоху // Боспорские исследования. Вып. XXXIII. Керчь: Керченская городская типография, 2016. С. 163–185.
Артюхин Ю.В., Артюхина О.И., Родионова Н.Б. Ейское морское побережье: история и проблемы освоения, природные основы реконструкции. Ейск: Фонд науки и образования, 2015. 205 с.
Артюхин Ю.В., Артюхина О.И., Суслов О.Н., Шереметьев В.М. Природные условия северного региона Краснодарского края и проблемы хозяйственного освоения его морского побережья. Ростов-на-Дону: Фонд науки и образования, 2016. 142 с.
Балабанов И.П., Измайлов Я.А. Изменение уровневого и гидрохимического режимов Черного и Азовского морей за последние двадцать тысяч лет // Водные ресурсы. 1988. № 6. С. 54–62.
Барг И.М. О возрасте и образовании Керченского пролива и Азовского моря // Докл. РАН. 2007. Т. 412. № 2. С. 220–222.
Воробьев В.П. Бентос Азовского моря. Симферополь: Крымиздат, 1949. 193 с.
Горецкий Г.И. Аллювиальная летопись великого Пра-Днепра. M.: Наука, 1970. 491 с.
Дикарёв В.А. О фанагорийской регрессии Черного моря // Вестник Московского государственного университета. Сер. 5. География. 2011. № 1. С. 35–40.
Зенкевич Л.А. Биология морей СССР. М.: Изд-во АН СССР, 1963. 740 с.
Зенкович В.П. Берега Черного и Азовского морей. М.: Географгиз, 1958. 374 с.
Иванов В.В., Коротаев В.Н., Римский-Корсаков Н.А. и др. Строение поймы и динамика русла Нижнего Дона // Вестник Московского университета. Серия 5. География. 2013. № 5. С. 60–66.
Ивлиева О.В., Фроленко Л.Н. Биогенное карбонатонакопление донных отложений Азовского моря во второй половине XX века // Изв. ВУЗов. Северо-Кавказский регион. Естественные науки. 2009. № 4. С. 96–100.
Маев Е.Г., Мысливец В.И., Зверев А.С. Строение верхнего слоя осадков и рельеф дна Таганрогского залива Азовского моря // Вестник Московского университета. Серия 5. География. 2009. № 5. С. 78–82.
Мамыкина В.А., Хрусталев Ю.П. Береговая зона Азовского моря. Ростов-на-Дону: Изд-во РГУ, 1980. 176 с.
Матишов Г.Г. Новые данные о геоморфологии дна Азовского моря // Докл. РАН. 2006. Т. 409. № 3. С. 375–380.
Матишов Г.Г. Сейсмопрофилирование и картирование новейших отложений дна Азовского моря // Вестник ЮНЦ РАН. 2007. Т. 3. № 3. С. 32–40.
Матишов Г.Г., Григоренко К.С. Причины осолонения Таганрогского залива // Докл. РАН. 2017. Т. 477. № 1. С. 92–96.
Матишов Г.Г., Григоренко К.С. Динамический режим Азовского моря в условиях осолонения // Докл. РАН. Науки о земле. 2020. Т. 429. № 1. С. 107–112.
Матишов Г.Г., Польшин В.В., Ковалёва Г.В., Титов В.В. Литология и биостратиграфия голоценовых отложений Азовского моря: итоги 15-летних исследований // Наука Юга России. 2019. Т. 15. № 3. С. 24–34.
Матишов Г.Г, Польшин В.В, Титов В.В. Исследования отложений кос Азовского моря (на примере косы Долгой) // Океанология. 2020. Т. 60. № 1. С. 158–161.
Матишов Г.Г., Польшин В.В. Новые результаты по истории Азовского моря в голоцене // Докл. РАН. 2019. Т. 489. № 2. С. 190–194.
Матишов Г.Г., Польшин В.В., Титов В.В., Шевердяев И.В. Голоценовая история азовского шельфа // Наука Юга России. 2019. Т. 15. № 1. С. 42–53.
Матишов Г.Г., Польшин В.В., Шохин И.В. Влияние геоморфологических особенностей дна Азовского моря на распределение бентоса // Вестник ЮНЦ РАН. 2010. Т. 6. № 2. С. 14–20.
Матишов Г.Г., Шохин И.В., Набоженко М.В., Польшин В.В. Многолетние изменения донных сообществ Азовского моря в связи с характером осадконакопления и гидрологическим режимом моря // Океанология. 2008. Т. 48. № 3. С. 425–435.
Набоженко М.В. Реконструкция и динамика таксоценоза двустворчатых моллюсков (Mollusca: Bivalvia) Азовского моря в позднем голоцене в связи с изменением солености // Тр. Зоологич. ин-та РАН. 2013. № 3. С. 182–191.
Невесская Л.А. Позднечетвертичные двустворчатые моллюски Черного моря, их систематика и экология // Тр. Палеонт. ин-та АН СССР. 1965. Т. 105. 390 с.
Определитель Черного и Азовского морей / Под ред. Мордухай-Болтовского Ф.Д. Т. 3. Киев: Наукова Думка, 1972. 340 с.
Панов Д.Г. История развития Азовского моря в голоцене // Океанология. 1965. Т. 5. № 4. С. 673–682.
Панов Д.Г., Хрусталев Ю.П. Об истории развития Азовского моря в голоцене // Докл. АН СССР. 1966. Т. 166. № 2. С. 429–432.
Польшин В.В. Донные отложения позднего голоцена Азовского моря // Геология, география и экология океана / Под ред. Г.Г. Матишова. Ростов-на-Дону: изд-во ЮНЦ РАН, 2009. С. 269–272.
Попов Г.И. Плейстоцен черноморско-каспийских проливов. М.: Наука, 1983. 216 с.
Свиточ А.А. Морской плейстоцен побережий России. М.: ГЕОС, 2003. 362 с.
Свиточ А.А., Янина Т.А. Новые данные по малакофауне морского плейстоцена Маныча // Докл. РАН. 2001. Т. 380. № 4. С. 570–573.
Семененко В.Н., Ковалюх Н.Н. Абсолютный возраст верхнечетвертичных отложений азово-черноморского бассейна по данным радиоуглеродного анализа // Геологический журнал. 1973. Т. 33. № 6. С. 91–97.
Федоров П.В. Послеледниковая трансгрессия Черного моря и проблема изменений уровня океана за последние 15 000 лет // Колебания уровня морей и океанов за 15 000 лет / Под ред. Каплина П.А., Клиге Р.К., Чепалыги А.Л. M.: Наука, 1982. С. 151–156.
Фроленко Л.Н. Зообентос Азовского моря и особенности его развития в современный период // Изв. ВУЗов Сев.-Кав. регион. Естеств. науки. 2000. № 4. С. 62–65.
Хрусталев Ю.П. Закономерности осадконакопления во внутриконтинентальных морях аридной зоны. Л.: Наука, 1989. 261 с.
Хрусталев Ю.П., Щербаков Д.А. Позднечетвертичные отложения Азовского моря и условия их накопления. Ростов-на-Дону: Изд-во Ростовского ун-та, 1974. 148 с.
Чухчин В.Д. Экология брюхорогих моллюсков Черного моря. Киев: Наукова Думка, 1984. 176 с.
Шнюков Е.Ф., Орловский Г.Н., Усенко В.П., Григорьев А.В., Гордиевич В.А. Геология Азовского моря. Киев: Наукова думка, 1974. 248 с.
Шохин И.В., Набоженко М.В., Сарвилина С.В., Титова Е.П. Современное состояние и закономерности распределения донных сообществ Таганрогского залива // Океанология. 2006. Т. 46. № 3. С. 432–441.
Янина Т.А. Неоплейстоцен Понто-Каспия: биостратиграфия, палеогеография, корреляция. М.: Географ. ф-тет МГУ, 2012. 264 с.
Brückner H., Kelterbaum D., Marunchak O. et al. The Holocene sea level story since 7500 BP – Lessons from the Eastern Mediterranean, the Black and the Azov Seas // Quaternary International. 2010. V. 225. № 2. P. 160–179.
World Imagery ESRI. URL: http://goto.arcgisonline. com/maps/World_Imagery (Accessed 12 December 2019).
Дополнительные материалы отсутствуют.