

Океанология, 2023, T. 63, № 1, стр. 108-123
Плейстоценовые океанологические условия в районе дрифта Иоффе (Южная Атлантика) по комплексам бентосных фораминифер
Е. А. Овсепян 1, *, Н. О. Гречихина 1, 2
1 Институт океанологии им. П.П. Ширшова РАН
Москва, Россия
2 Геологический институт РАН
Москва, Россия
* E-mail: eovsepyan@ocean.ru
Поступила в редакцию 21.12.2021
После доработки 26.09.2022
Принята к публикации 28.10.2022
- EDN: AGEWQF
- DOI: 10.31857/S0030157423010124
Аннотация
Контуритовый дрифт Иоффе, расположенный в юго-западной Атлантике к северо-востоку от канала Вима, был открыт в 32-м рейсе НИС “Академик Иоффе” в 2010 г. Проведенный ранее комплексный анализ сейсмоакустических, биостратиграфических, литологических, гранулометрических, геохимических и рентгено-флюоресцентных данных по шести колонкам донных осадков позволил установить плейстоценовый возраст верхней части дрифта, а также доказать контуритовый генезис отложений. Количественные анализы комплексов бентосных фораминифер, проведенные по поднятой вблизи вершины дрифта Иоффе колонке АИ-2436 (26°51.6′ ю.ш., 34°01.40′ з.д., глубина моря 3800 м), позволили реконструировать относительные вариации потока и периодичности поступления органического вещества на дно, изменения скоростей придонных течений, а также предположить смену омывавших район исследования водных масс в плейстоцене. Визуальная оценка раковин бентосных фораминифер позволила выделить три группы предположительно разных по генезису повреждений. Первая группа деформаций раковин связана с разрушением их целостности в результате ударов перемещающихся частиц осадка в условиях повышенных скоростей придонных течений, вторая объединяет повреждения, появившиеся в результате транспорта самих раковин на небольшие расстояния, третья включает в себя деформации, полученные при воздействии агрессивной придонной среды, приводившей к растворению карбонатных микрофоссилий. Таким образом, характерные особенности внешнего вида раковин БФ в комплексе с традиционными критериями, вероятно, можно использовать в будущем в качестве дополнительного индикатора при диагностике контуритов, однако, такая методика требует детальной разработки критериев применения и проверки на соответствующем материале.
ВВЕДЕНИЕ
Бентосные фораминиферы (БФ) являются важными индикаторами ряда параметров окружающей среды (например, [18, 21, 24, 36, 49]), поэтому количественный анализ видового состава сообществ из глубоководных районов часто используется при реконструкциях палеоокеанологических обстановок разного временно́го масштаба. С его помощью восстанавливают колебания биопродуктивности поверхностных вод, потока органического вещества на дно и периодичности его поступления (например, [1, 13, 23]), изменения содержания кислорода на границе вода–осадок [10, 50], интенсивности придонных течений [1, 36, 49], степени агрессивности придонных вод по отношению к карбонатным компонентам осадка [36, 44] и смену водных масс [2, 36] в прошлом для разных районов Мирового океана.
Современные исследования комплексов БФ в колонках глубоководных донных осадков сосредоточены в основном в районах континентальных склонов, где относительно небольшие глубины и/или высокие скорости осадконакопления способствуют сохранению палеоокеанологической летописи с временны́м разрешением от тысяч до сотен лет. Для центральных частей океанов количество таких работ существенно меньше из-за медленного осадконакопления и, следовательно, низкого временно́го разрешения получаемых данных, а также вследствие близости лизоклина и критической глубины карбонатонакопления, влияющих на сохранность карбонатных микрофоссилий.
В юго-западной Атлантике по комплексам БФ были реконструированы изменения биопродуктивности в котловине Сантус в течение последних 570 тыс. лет [23] и 769 тыс. лет [13]. Установлено, что в котловине Сантус сезонная поставка фитодетрита на дно происходила в интервалы оледенений [13]. Восточнее, на плато Риу-Гранди, усиление сезонного контраста в поступлении органического вещества на дно в течение оледенений и стадиалов, а также увеличение среднегодовой биопродуктивности поверхностных вод в течение межледниковий и межстадиалов восстановлено по комплексам БФ [1] в интервале восьми последних изотопно-кислородных стадий [29]. На южном уступе плато Сан-Паулу хорошая сохранность карбонатных микрофоссилий зафиксирована в интервалах межледниковий и межстадиалов, в то время как интенсивное растворение отмечено в пределах стадиалов [44]. Авторы связывают колебания сохранности раковин с вертикальным смещением верхней границы агрессивных водных масс субантарктического и антарктического происхождения в течение двух последних климатических циклов. Ледниково-межледниковая смена водных масс реконструирована в экваториально-тропической Атлантике в интервале среднего плейстоцена–голоцена [32]. Позднеплейстоцен–голоценовая динамика глубинной циркуляции реконструирована по комплексам БФ в огибающем плато Риу-Гранди с востока канале Хантер [2].
Контуритовый дрифт Иоффе расположен к северу от плато Риу-Гранди и к северо-востоку от канала Вима – основного пути проникновения Антарктических придонных вод из Южной Атлантики в тропические широты (рис. 1). Эта аккумулятивная структура была открыта в ходе 32-го рейса НИС “Академик Иоффе” в 2010 г. [41]. Комплексный анализ сейсмоакустических, биостратиграфических, литологических, гранулометрических, геохимических и рентгено-флюоресцентных данных, проведенный по шести колонкам (рис. 1), позволили установить плейстоценовый возраст верхней части дрифта, а также доказать контуритовый генезис отложений [9, 25–29, 42].
Рис. 1.
Карта района исследования, гидрология (А) и положение мест отбора колонок донных осадков на дрифте Иоффе (Б). При построении карты использовалась батиметрическая основа GEBCO [17]. Гидрология района по [8] и ссылки в публикации. Серые стрелки – пути распространения глубинных вод североатлантического происхождения; черные стрелки – направления движения глубинных и придонных вод антарктического происхождения. ВЦГВ – Верхняя циркумполярная глубинная вода, НЦГВ – Нижняя циркумполярная глубинная вода, ААДВ – Антарктическая донная вода.

Целью данной работы является реконструкция палеоокеанологических условий по комплексам БФ по колонке АИ-2436 в сочетании с полученными ранее данными [9, 25–29, 42] по этому же разрезу в течение плейстоцена.
ФИЗИКО-ГЕОГРАФИЧЕСКОЕ ПОЛОЖЕНИЕ И ОКЕАНОЛОГИЧЕСКИЕ ОБСТАНОВКИ
Район исследования расположен в южной части Бразильской котловины в юго-западной Атлантике, к северу от подводного плато Риу-Гранди между 26 и 28° ю.ш. и 33 и 36° з.д. (рис. 1). Контуритовый дрифт Иоффе покрывает центральную часть узкого тектонического хребта, ограниченного с юга и севера разломами глубиной около 250 м [8]. Описанный комплекс геоморфологических структур является частью зоны разломов Флорианополис [38], расположенной к востоку и северо-востоку от изучаемого района. Хребет простирается с юго-запада на северо-восток и возвышается над дном котловины более чем на 700 м. Склоны хребта асимметричны: южный склон более пологий, чем северный. Вершина подводного поднятия с учетом покрывающего его контуритового дрифта располагается на глубине 3750 м.
Глубинная циркуляция Южной Атлантики определяется взаимодействием распространяющейся с севера на юг Северо-Атлантической глубинной воды (САГВ) и следующими с юга на север водами антарктического и субантарктического происхождения (Верхней (ВЦГВ) и Нижней (НЦГВ) циркумполярной глубинной воды и Антарктической донной воды (ААДВ) [40]. Вершина дрифта Иоффе омывается НЦГВ, характеризующейся потенциальной температурой 0.2–2°С [40]. Согласно натурным измерениям в каналах севернее дрифта, скорости придонных течений составляют более 20 м/с [3].
В настоящее время район исследования характеризуется низкой биопродуктивностью фотического слоя, главным образом, из-за дефицита нитратов и нитритов [37], необходимых для развития фитопланктона. Концентрация хлорофилла-а в поверхностном слое составляет 0.1–0.3 мг/м3, величина первичной продукции также низка и соответствует 200–400 мгC/м2/день (или ~73–146 гС/м2/год) [37]. Придонные воды насыщены кислородом, его концентрация составляет 5.25–5.5 мл/л [16].
Фораминиферовый лизоклин и критическая глубина карбонатонакопления в районе канала Вима и плато Риу-Гранди составляют 4050 и 4500 м соответственно [39].
МАТЕРИАЛ И МЕТОДЫ
Колонка АИ-2436 (26°51.6′ ю.ш., 34°01.40′ з.д., глубина моря 3800 м) была поднята вблизи вершины дрифта Иоффе [26]. Хотя колонка отобрана ударной трубкой длиной 6 м, ее длина составляет 714 см из-за растяжения при извлечении осадка из трубы на борту судна методом гидровыталкивания. Судя по рейсовым фотографиям колонки, наиболее деформированные участки находятся выше 139 см [26]. Разрез сложен нано-фораминиферовым илом с прослоями песка, представленного фораминиферами хорошей сохранности, мощностью от 1 до 6 см [26]. Железомарганцевые конкреции шаровидной и цилиндрической формы 1–3 см в диаметре встречены в верхних 250 см колонки и на уровне 668–670 см. Плейстоценовый возраст вскрытых осадков был определен ранее методом зональной стратиграфии по планктонным фораминиферам (ПФ) и нанопланктону [25].
Образцы на фораминиферовый анализ мощностью 2 см были отобраны через каждые 10 см по всей длине колонки, промыты через сита с ячеей 0.5, 0.25 и 0.1 мм и высушены при комнатной температуре. БФ были изучены в каждой фракции; результаты сравнивались между собой. Однако, из-за небольшого количества раковин БФ размером >0.5 мм, подсчеты во фракциях >0.5 и 0.5–0.25 мм были объединены, и распределение БФ анализировалось во фракции >0.25 мм. Отдельно были проведены подсчеты во фракции 0.25–0.1 мм, а затем было проанализировано распределение БФ в суммарной фракции >0.1 мм. В каждой пробе было подсчитано 250–300 экземпляров БФ, где это возможно. Однако, в случае малого обилия раковин БФ в образце 100 подсчитанных экземпляров считались статистически значимым количеством для проведения палеоокеанологических реконструкций [22]. При высокой концентрации БФ проба делилась на равные части микросплиттером Отто. Из-за отсутствия данных о плотности и массе сухого осадка расчеты фораминиферового числа (количество экземпляров на 1 г сухого непромытого осадка) и примерных скоростей аккумуляции раковин произведены не были. Визуальные оценки показывают, что БФ составляют крайне малую часть (около 1%) фракции во всех изученных пробах.
В полученном для каждого образца комплексе были определены преобладающие и второстепенные виды с содержанием >15% и 15–5% от общего количества подсчитанных раковин хотя бы в одном образце. Интерпретация колебаний изменчивости каждого вида проводилось на основании экологических предпочтений видов БФ (табл. 1 ). Некоторые раковины БФ были определены до рода с формированием сборных родовых групп, таких как Quinqueloculina spp., Lagena spp., Oolina spp., Fissurina spp. и др., вследствие слабой изученности их экологии и крайне низкого обилия отдельных видов.
Таблица 1.
Экологические предпочтения основных видов бентосных фораминифер по литературным данным
| Вид | Экология | Ссылки | |
|---|---|---|---|
| Nutallides umbonifer | Эпифауна/ поверхностная инфауна |
В море Уэдделла преобладает на глубинах >3500 м | [35] |
| В Южной Атлантике этот вид обилен в областях распространения придонных вод, агрессивных по отношению к карбонату кальция и насыщенных кислородом. Распространен в осадках между кальцитовым лизоклином и критической глубиной карбонатонакопления | [36, 49] | ||
| При доминировании E. exigua в сообществах этот вид указывает на сильный контраст в поступлении органического вещества на дно, при этом сам поток характеризуется как низкий или умеренный | [20] | ||
| Приспособлен к разной периодичности поступления органического вещества на дно, но, по-видимому, предпочитает условия резко выраженного сезонного контраста | [33] | ||
| Вероятно, благоприятными факторами среды для развития этого вида являются олиготрофные условия и отсутствие свежего (лабильного) фитодетрита | [11, 48] | ||
| Globocassidulina subglobosa | Инфауна | В море Уэдделла доминирует на глубинах 2500–3500 м | [35] |
| В Южной Атлантике доминирует в районах с потоком органического вещества на дно ≤1 гС/м2/год | [36] | ||
| Предпочитает условия низкой сезонности поступления органического вещества на дно | [33] | ||
| В Южной Атлантике этот вид обитает в условиях повышенной гидродинамической активности придонных вод и низким потоком органического вещества на дно. Найден в песках с низким содержанием органического углерода и высоким содержанием карбоната кальция | [49] | ||
| Oridorsalis umbonatus | Эпифауна/ поверхностная и промежуточная инфауна |
Поверхностная инфауна, обитает на глубинах >2000 м в условиях стабильно низкого потока сильно преобразованного органического вещества на дно | [36, 46] |
| Вид предпочитает условия низкого потока органического вещества на дно и высокого содержания кислорода на границе вода–осадок | [47] | ||
| В восточной части Индийского океана обнаружен в холодноводных условиях (<3°С) с высоким содержанием кислорода (>3.5 мл/л); вид обитает в условиях низкого потока органического вещества на дно (<3 гC/м2/год) | [43] | ||
| Предпочитает условия умеренно выраженной сезонности (несколько эпизодов в течение года) поступления органического вещества на дно | [33] | ||
| В Южной Атлантике обитает в условиях низкого, но относительно постоянного потока органического вещества на дно | [36] | ||
| В море Уэдделла встречается вместе с E. exigua выше карбонатного лизоклина | [35] | ||
| Cibicidoides wuellerstorfi | Эпифауна | Обитает в условиях стабильно низкого потока органического вещества на дно (<1 гС/м2/год). Доминирование этого вида в сообществах отмечается в областях влияния САГВ | [36] |
| Питается взвешенным органическим веществом, предпочитает селиться на возвышенном субстрате | [34] | ||
| Обитает в областях с низким потоком органического вещества на дно (<2 гС/м2/год). В Индийском океане присутствует в комплексе с фитодетиритовым видом E. exigua | [5] | ||
| Предпочитает условия умеренно выраженной сезонности (несколько эпизодов в течение года) поступления органического вещества на дно | [33] | ||
| В то же время, в Южной Атлантике обилие вида коррелирует с низким, но относительно постоянным потоком органического вещества на дно. Избегает сильно агрессивных по отношению к карбонатам условий | [36] | ||
| Вид способен адаптироваться к широкому диапазону условий среды | [15] | ||
| В юго-восточной Атлантике распределение вида коррелирует с распространения ядра САГВ | [48] | ||
| Epistominella exigua | Поверхностная инфауна | Оппортунистический вид, быстро размножающийся и заселяющий субстрат при наличии фитодетрита | [18] |
| Предпочитает условия низкой гидродинамики | [36] | ||
| В восточной части Индийского океана предпочитают холодноводные условия (<3°С) с высоким содержанием кислорода вблизи дна (>3.5 мл/л). Обилие этого вида отрицательно коррелирует с разнообразием комплекса | [43] | ||
| Присутствие этого вида может указывать на периодическое (сезонное) поступление органического вещества на дно. Способен быстро размножаться при обилии фитодетрита | [18, 43] | ||
| Обитает в условиях высокого содержания кислорода вблизи дна и хорошо адаптируется к олиготрофным условиям с низким потоком органического вещества на дно | [36, 49] | ||
| Pullenia bulloides | Поверхностная инфауна | Предпочитает условия низкой сезонности поступления органического вещества на дно | [33] |
| Pullenia spp. | Поверхностная инфауна | Найден в осадках, обогащенных фитодетритом | [18] |
| Виды рода Pullenia (преимущественно P. bulloides) считаются индикатором интенсивного поступления органического вещества на дно и высокой (изменчивой) биопродуктивности поверхностных вод | [36] | ||
| Pyrgo murrhina | Эпифауна | В восточной части Индийского океана предпочитают холодноводные условия (<3°С) с высоким содержанием кислорода (>3.5 мл/л); обитают в условиях низкого потока органического вещества на дно (<3 гC/м2/год). Обилие этого вида отрицательно коррелирует с разнообразием комплекса | [43] |
| В колонках и Индийского океана присутствие этого вида наблюдается при хорошей сохранности всего комплекса карбонатных микрофоссилий | [20] | ||
| Gyroidina orbicularis, Gyroidina altiformis | Поверхностная- промежуточная инфауна | В Средиземном море обильны в областях с потоком свежего лабильного органического вещества менее 2.5 гC/м2/год | [15] |
| Oolina spp., Lagena spp. | Инфауна | Детальная экология представителей этих групп до конца не определена. Предполагается, что представители этих групп обитают в широком диапазоне величин потока органического вещества на дно | [15] |
| Quinqueloculina spp. | Эпифауна- поверхностная инфауна | Предпочитает фитодетрит в качестве пищи | [19] |
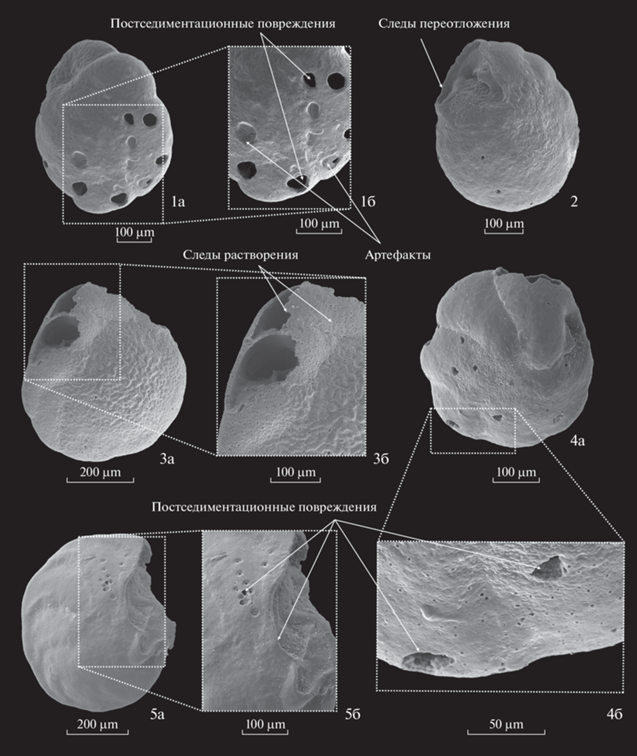
Раковины БФ были также проанализированы на предмет деформаций и характера поверхности. Согласно предыдущим исследованиям, эти характеристики могут оказаться полезными при оценке степени растворения раковин или влияния придонных течений на осадконакопление [11, 45, 51]. Фотографии раковин с характерными повреждениями сделаны на сканирующем электронном микроскопе TESCAN VEGA-II XMU в Палеонтологическом институте им. А.А. Борисяка РАН.
РЕЗУЛЬТАТЫ
В результате количественных анализов комплексов БФ по колонке АИ-2436 определено 36 видов и 18 сборных родовых групп.
Преобладающими видами в суммарной фракции >0.1 мм являются Nutallides umbonifer (25–69%) и Globocassidulina subglobosa (8–35%), к второстепенным видам и сборным родовым группам относятся Cibicidoides wuellerstorfi (0.3–8.8%), Oridorsalis umbonatus (1–12.3%), Epistominella exigua (2.3–14.4%), Pullenia bulloides (1–9.6%), Fissurina spp. (0.4–6%), Lagena spp. (0.6–7.4%) В небольших количествах встречены Ioanella tumidula (0–1.3%), Gyroidinoides soldanii (0–2%), Gyroidina lamarkiana (0–3.6%). Процентные содержания других сборных родовых групп составляют: Quinqueloculina spp. (0–2.5%), Oolina spp. (0–0.5%).
Во фракции >0.1 мм N. umbonifer доминирует по всей колонке АИ-2436 (рис. 2), G. subglobosa демонстрирует повышенные значения процентного содержания в нижней части колонки (714–440 см), в то время как O. umbonatus и E. exigua варьируют по разрезу без видимых закономерностей. Представители I. tumidula и G. lamarkiana практически полностью отсутствуют в интервале 714–600 см и появляются в малом количестве в верхней части колонки. Экологические предпочтения основных индикаторных видов представлены в таблице.
Рис. 2.
Распределение основных видов бентосных фораминифер и содержания органического углерода [26] во фракции >0.1 мм в колонке АИ-2436. Стратиграфическое расчленение по [25]. ПФ – планктонные фораминиферы, НФ – нанофоссилии, ОВ – органическое вещество. Волнистыми линиями показаны перерывы в геологической летописи колонки (выпадения стратиграфических интервалов из разреза).

Сопоставление данных во фракциях >0.25, 0.25–0.1 и >0.1 мм показало, что доминирующими видом во всех трех фракциях является N. umbonifer (рис. 3). Структура комплексов основных видов БФ в суммарной фракции >0.1 мм практически полностью повторяет таковую во фракции 0.1–0.25 мм, поэтому кривые для этой фракции на рис. 3 не показаны. Во фракции >0.25 мм при абсолютном преобладании N. umbonifer отмечены вариации процентных содержаний видов C. wuellerstorfi, O. umbonatus и Pyrgo spp., которые наиболее заметно проявляются в интервале 541–131 см (рис. 3).
Рис. 3.
Распределение процентного содержания бентосных фораминифер во фракциях >0.1 и >0.25 мм. Для расшифровки сокращений см. подпись к рис. 2.

Анализ состава сообществ БФ показал, что комплексы состоят, главным образом, из видов с толстостенными раковинами. Отмечено значительное число повреждений на раковинах видов N. umbonifer и C. wuellerstorfi по всей длине колонки. На многих раковинах обнаружены хаотично расположенные сколы, вмятины, царапины, отверстия (рис. 4). Часто у таких представителей отсутствует последняя камера, причем сколотые края, как правило, окатанные. Наряду с раковинами, имеющими вышеописанные черты, встречаются экземпляры, стенка которых характеризуется свежими сколами и относительно равномерным распределением мелких отверстий, не являющихся порами.
Рис. 4.
Фотографии бентосных фораминифер со следами переотложения и растворения. 1а, 1б – Nuttallides umbonifer, 600–602 см, 2, 3а, 3б – Nuttallides umbonifer, 550–552 см, 4а, 4б – Nuttallides umbonifer, 620–622 см, 5а, 5б – Cibicidoides wuellerstorfi, 550–552 см. Стрелочками показаны: постседиментационные повреждения, которые интерпретированы как результат ударов перемещающихся под действием придонных течений вдоль поверхности дна других компонентов осадка; следы переотложения – окатанные сколы поврежденной раковины; следы растворения – неокатанные сколы поврежденной раковины и вторичная пористость; артефакты подготовки к сканированию – вероятное загрязнение раковины, поверх которого легло напыление.
ОБСУЖДЕНИЕ
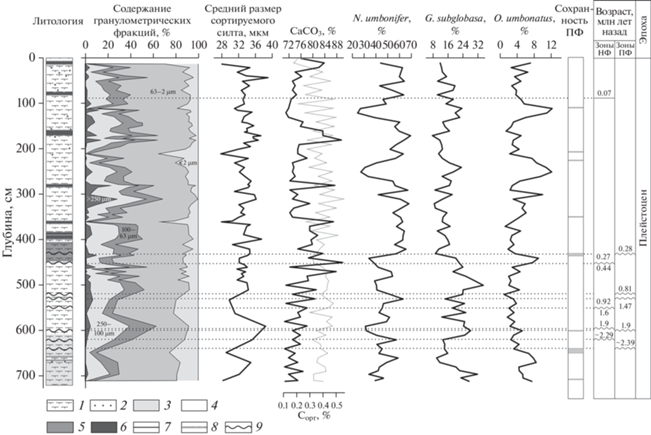
Биопродуктивность поверхностных вод и поступление органического вещества на дно. Встреченные в пробах колонки АИ-2436 комплексы видов БФ свидетельствуют о преобладании олиготрофных условий в районе исследования, начиная с раннего плейстоцена (рис. 2). В частности, абсолютное доминирование вида N. umbonifer, обитающего при низком потоке органического вещества на дно [11, 48], указывает на дефицит пищи для донных микроорганизмов в течение всего времени накопления изученных осадков. Виды БФ O. umbonatus, C. wuellerstorfi и G. subglobosa также являются индикаторами ограниченного поступления органического вещества из фотического слоя на дно [5, 43, 47, 49], что свидетельствует в пользу низкой биопродуктивности поверхностных вод и подтверждается малым (<1%) содержанием органического углерода в осадке (рис. 5, [26, 42]). С одной стороны, низкий поток свежего органического вещества мог быть обусловлен низкой продукцией фитопланктона в фотическом слое вследствие дефицита биогенных элементов. Действительно, удаленное положение района исследования от океанских (зон апвеллинга) и континентальных (устьев рек и областей устойчивых ветровых потоков) источников биогенных элементов, может объяснить низкую биологическую продуктивность над дрифтом Иоффе в течение плейстоценового времени. С другой стороны, сейсмоакустические, литологические, гранулометрические, биостратиграфические и рентгено-флюоресцентные данные по этой и соседним (АИ-3316, АИ-3317, АИ-3318 и АИ-3655; рис. 1) колонкам указывают на ведущую роль придонных течений в процессе формирования дрифта [9, 26, 27, 42], поэтому часть вертикально осажденного органического вещества могла вымываться из осадка при высоких скоростях течений.
Рис. 5.
Гранулометрический состав [26], доля сортируемого силта [42], содержание органического углерода и карбонатность осадка [26], процентное содержание индикаторных видов бентосных фораминифер и интервалы растворения [25]. Стратиграфическое расчленение по [25]. ПФ – планктонные фораминиферы, НФ – нанофоссилии. Условные обозначения: состав: 1 – нано-фораминиферовый ил, 2 – железо-марганцевые конкреции; цвет: 3 – бледно-оранжевый, 4 – серовато-оранжевый, 5 – темно-желтовато-оранжевый, 6 – желтовато-серый; контакт: 7 – резкий, 8 – постепенный, 9 – эрозионный.
Некоторые исследователи отмечают, что отсутствие свежего фитодетрита является благоприятным фактором для развития N. umbonifer [11, 48] и O. umbonatus [36, 46], поэтому доминирование этих видов, вероятно, отражает преобладание преобразованного органического вещества на дне в течение всего времени накопления осадка. По-видимому, то небольшое количество фитодетрита, которое образовывалось в поверхностном слое в условиях дефицита биогенных элементов, подвергалось бактериальной деградации в процессе осаждения через водную толщу и поступало на дно уже в переработанном виде. С другой стороны, придонные течения могли не только вымывать осевшие органические частицы, но и приносить частично переработанное органическое вещество в составе нефелоидных потоков.
Влияние придонных течений на осадконакопление было ранее подтверждено сейсмоакустическими и гранулометрическими данными [42]. Интерпретируя полимодальные гранулометрические распределения по этой колонке, И.О. Мурдмаа и соавторы [42] предположили, что тонкая фракция осадка (<0.063 мм), представленная, в основном, фрагментами раковин ПФ и кокколитами, могла поступать на дрифт Иоффе с придонными течениями в виде суспензии. Этот материал, вероятно, представлял собой продукт эрозии донных осадков, отлагавшихся ниже лизоклина в удаленных районах, предположительно, в канале Вима и/или глубоководных желобах и впадинах, окружающих дрифт [42].
Несмотря на преобладание N. umbonifer – вида, способного питаться преобразованным органическим веществом, – в комплексе БФ в небольшом количестве содержатся виды, которые в современных условиях используют свежий фитодетрит в качестве пищи (рис. 2, таблица). К ним относятся E. exigua [18, 43], Pullenia spp. [18], Quinqueloculina spp. [19]. Хотя суммарная доля этих видов сравнительно мала, тем не менее, их присутствие в комплексах БФ может указывать на наличие небольшой доли свежего органического вещества в осадке. Хотя E. exigua и Pullenia spp. встречаются в районах с интенсивным сезонным поступлением фитодетрита на дно [18, 36, 43], развитие фауны этих видов в типично олиготрофном комплексе БФ, вероятно, свидетельствует об их хорошей приспособляемости к разным трофическим условиям и, соответственно, о широком диапазоне обитания.
Влияние придонных течений. Толщина стенки и характер поверхностей раковин некоторых представителей БФ могут указывать на воздействие придонных течений на комплексы микрофоссилий в процессе накопления осадка [45, 51]. В частности, замечено, что в энергоактивных зонах, где накапливается крупнозернистый неконсолидированный осадок, комплекс БФ формируется, главным образом, видами с большой толщиной стенки раковин [51]. Основную долю видов в комплексе БФ в колонке АИ-2436 составляют толстостенные представители N. umbonifer, C. wuellerstorfi, G. subglobosa, O. umbonatus, Gyroidina spp., Pyrgo spp., что может указывать на гидродинамическую активность придонных вод в районе дрифта Иоффе в течение всего времени накопления осадка.
Обилие раковин N. umbonifer и C. wuellerstorfi с многочисленными постседиментационными повреждениями поверхностей, таких как царапины, углубления, отверстия, сколы, может свидетельствовать о влиянии придонных течений на компоненты донных отложений на границе вода–осадок [45, 51]. Деформации на поверхности раковин могут быть разделены на три группы, предположительно разные по генезису.
Первая группа включает в себя вмятины, углубления и отверстия, размер которых намного больше характерных для данных таксонов пор (рис. 4, фиг. 1, 4). Такие повреждения могли образовываться в результате ударов перемещающихся под действием придонных течений других частиц осадка, например, более легких и округлых раковин ПФ и их обломков. На рис. 4 (фиг. 1, 4) показаны раковины вида N. umbonifer, на которой отверстия сконцентрированы на одной стороне. Вероятно, представители эпифауны/поверхностной инфауны N. umbonifer во время накопления осадка были частично погружены в субстрат, таким образом, возвышающиеся над поверхностью дна участки раковин оказывались уязвимыми к ударам перемещающихся частиц.
Ко второй группе деформаций можно отнести повреждения из первой группы в сочетании с царапинами, окатанными сколами, сглаженной орнаментацией и разрушенными стенками камер (рис. 4, фиг 2, 5). Такие признаки свидетельствуют о переотложении раковин БФ придонными течениями, как было экспериментально доказано зарубежными коллегами [45]. На основании гранулометрических данных И.О. Мурдмаа и соавторы [42] показали, что основную часть аллохтонного материала, приносимого придонными течениями, составляла фракция мелкого силта (силт: 2–63 мкм, мелкий силт 2–10 мкм по [7]), включающая в себя нанофоссилии и их обломки. Раковины БФ, которые в данной колонке представлены видами с толстой стенкой, являются слишком тяжелыми для дальнего переноса, но, учитывая характерные деформации, их локальное перемещение (волочение) на небольшие расстояния в условиях подвижных придонных вод, вероятно, происходило.
Третья группа повреждений включает в себя неокатанные края разрушенных камер и истонченные стенки раковины. Такой тип деформаций характерен для агрессивной среды, способствующей растворению карбонатных компонентов осадка [11]. В колонке АИ-2436 экземпляры с подобными характерными чертами были найдены в небольшом количестве по всему разрезу, с незначительным увеличением содержания в интервалах плохой сохранности раковин ПФ (рис. 5, [28]), что, с одной стороны, подтверждает эпизодическое усиление растворения на дрифте Иоффе в течение плейстоцена. С другой стороны, встречаемость раковин с деформациями третьего типа по всей длине колонки показывает, что постепенное разрушение раковин под действием придонной морской воды, по-видимому, происходило непрерывно в процессе формирования осадка, что, скорее всего, было обусловлено крайне низкими скоростями седиментации.
Несмотря на то, что данной работе подсчет раковин отдельных видов с соответствующими деформациями произведен не был, анализ внешнего вида раковин может оказаться перспективным для разработки вспомогательного индикатора для диагностики контуритовых отложений. Данный подход требует детальной разработки методологии и последующей проверки на материале с доказанным контуритовым генезисом.
Кроме характера поверхности раковин, изменения процентного содержания вида G. subglobosa, обитающего, в том числе в условиях повышенной гидродинамической активности придонных вод [49], позволяют судить об относительных изменениях скоростей придонных течений в плейстоценовое время. Увеличение доли этого вида в нижней части колонки (интервал ~2.29–0.44 млн л.н.) может свидетельствовать о повышенной активности придонных течений в течение раннего и среднего плейстоцена. Крайне низкие процентные содержания видов I. tumidula и Gyrodina spp., питающихся фитодетритом [14, 52], могут подтвердить данное предположение. Скорее всего, практически полное отсутствие указанных таксонов в комплексах БФ в нижней части колонки отражает уменьшение количества фитодетрита на дне не столько за счет вариаций потока органического вещества на дно, сколько вследствие его вымывания сильными придонными течениями в раннеплейстоценовое время.
Такая интерпретация комплексов БФ согласуется с ранее опубликованными выводами [26, 28, 41, 42]. Авторы показали, что увеличение количества перерывов в геологической летописи (отсутствие и сокращение мощности некоторых стратиграфических зон и резкие высокоамплитудные пики магнитной восприимчивости) фиксируется именно в нижней части колонки и указывает на пульсационный характер придонных течений в раннем плейстоцене [4, 28, 42]. Согласно гипотезе о природе перерывов [4, 28, 42], увеличение скоростей придонных течений до величин, способных эродировать накопившийся осадок, приводило к размыву верхней толщи осадочного чехла. При этом хроностратиграфический объем уничтоженных эрозией отложений не обязательно соответствовал длительности эрозионного процесса [4, 28, 42]. Как отмечают авторы [4, 28, 42], продолжительность эрозии могла быть кратковременной, в том числе и геологически мгновенной.
Палеоокеанологические условия на дрифте Иоффе в течение плейстоцена. На основании анализа комплексов БФ выделено четыре палеоокеанологически контрастных временны́х интервала, которые описывают плейстоценовую историю формирования осадочного чехла на вершине дрифта Иоффе.
Интервал до ~2.39 млн л.н. (начало плейстоцена) характеризовался низким потоком органического вещества на дно, судя по типично олиготрофным комплексам БФ (рис. 2). Повышенные процентные содержания G. subglobosa, C. wuellerstorfi, P. bulloides, обитающие в условиях постоянного и/или высокопериодичного поступления органического вещества на дно (таблица), указывают на слабовыраженную сезонность биопродуктивности поверхностных вод над дрифтом Иоффе в начале плейстоцена. Этот вывод согласуется с результатами С. Кендера и соавторов [30], которые показали, что сезонность поступления органического вещества на дно могла являться важным фактором, обусловившим глобальное распределение глубоководных БФ в течение раннего плейстоцена и последующего среднеплейстоценового перехода (~1.25–0.7 млн л.н.).
Повышенная доля G. subglobosa может указывать не только на количество и характер поступления органического вещества на дно, но и свидетельствовать об усилении придонных течений в раннем плейстоцене. Присутствие раковин БФ с характерными повреждениями также указывает на влияние придонных течений на процесс накопления осадка на дрифте Иоффе.
Кроме сезонности поступления органического вещества на дно, агрессивность придонных вод по отношению к карбонатным микрофоссилиям также, по-видимому, влияла на состав комплексов БФ на дрифте Иоффе. Пониженные концентрации вида N. umbonifer, который обычно доминирует при более экстремальных условиях, может указывать на установление слабоагрессивной придонной среды в раннем плейстоцене. Действительно, многочисленные опубликованные данные свидетельствуют о том, что в теплую эпоху позднего плиоцена и раннего плейстоцена усиление Атлантической меридиональной циркуляции приводило к экспансии на юг менее агрессивной палео-САГВ, присутствие которой было реконструировано на глубинах до 4350 м в Юго-Восточной Атлантике [6]. Судя по структуре комплекса БФ (рис. 2), вероятно, что палео-САГВ омывала и дрифт Иоффе на глубине отбора колонки (современная глубина 3800 м) в это время. Однако такое предположение требует дальнейших исследований и проверки независимыми методами.
Интервал ~2.39–1.6/0.92 млн л.н. по нанофоссилиям (или ~2.39–1.47/0.81 млн л.н. по ПФ) характеризуется сходными с предыдущим интервалом условиями (рисунки 2, 3 и 5). Однако увеличение доли G. subglobosa может свидетельствовать об усилении придонных течений на дрифте Иоффе в это время. Данный вывод подтверждается найденными поврежденными раковинами N. umbonifer и C. wuellerstorfi. Согласно опубликованным данным ([6] и ссылки в работе), между 2.0 и 1.5 млн л.н. происходило максимально интенсивное формирование аналога САГВ в Северной Атлантике, что, по-видимому, сопровождалось усилением придонных течений на глубинах распространения этой водной массы в юго-западной Атлантике и, возможно, на дрифте Иоффе. В пользу усиления придонных течений во время максимальной активности формирования палео-САГВ свидетельствует и перерыв, маркирующий окончание интервала и образовавшийся, по-видимому, в результате размыва большей части среднеплейстоценовых осадков [25, 42].
Интервал 0.92 (0.81)–0.44/0.27 млн. л.н. отмечен появлением крупных представителей видов C. wuellerstorfi, Pyrgo spp., Quinqueloculina spp. Lagena spp. во фракции >0.25 мм наряду с постоянным их присутствием в более мелкой фракции (рис. 3). Обилие разных размеров раковин одного вида говорит об установлении благоприятных условий для его существования. В настоящий момент перечисленные виды обитают в условиях нерегулярного (пульсационного) потока органического вещества на дно, так что их присутствие в комплексе БФ в интервале 0.92 (0.81)–0.44/0.27 млн л.н., скорее всего, свидетельствует в пользу более ярко выраженного сезонного характера формирования первичной продукции над дрифтом Иоффе в этом время. Такой вывод согласуется с результатами С. Кендера и соавторов [30], которые предположили глобальное усиление сезонности в образовании органического вещества в фотическом слое после среднеплейстоценового перехода. Переход к выраженному сезонному характеру цветения фитопланктона происходил на фоне установления 100-тысячелетней периодичности ледниково-межледниковых циклов (например, [12]) и ростом ледниковых щитов в Северном полушарии, что обусловило увеличение сезонного контраста температур воздуха и поверхностных вод. Усиление сезонного контраста в Южной Атлантике в среднем-позднем плейстоцене предполагают и другие авторы на основании микропалеонтологического анализа [1, 13]. Несмотря на усиление сезонности, среднегодовой поток органического вещества оставался низким, как следует из доминирования вида-индикатора низкой продуктивности N. umbonifer.
Увеличение процентных содержаний N. umbonifer также может свидетельствовать о более агрессивной по отношению к карбонатным микрофоссилиям среде в интервале 0.92 (0.81)–0.44/ 0.27 млн л.н. по сравнению с рассмотренными ранее интервалами. Это может быть связано с усилением влияния палео-НЦГВ/ААДВ и сокращением продукции палео-САГВ в течение оледенений [6].
Максимальные процентные содержания G. subglobosa могут отражать усиление придонных течений в интервале 0.92 (0.81) –0.44/0.27 млн л.н.. Это предположение согласуется с предыдущими выводами об увеличении продукции ААДВ между 0.7 и 0.4 млн л.н. за счет усиления конвекции в Южном океане и интенсивного формирования льда на антарктическом шельфе [31]. Суммируя высказанное, можно предположить, что после среднеплейстоценового перехода в результате глобальной перестройки термохалинной циркуляции вершину дрифта Иоффе стали омывать воды антарктического происхождения.
Интервал последних 0.28 млн лет характеризовался более спокойной, холодной и стабильной гидродинамической обстановкой, судя по монотонному распределению видов БФ. Общее похолодание климата, установившееся после среднеплейстоценового перехода (например, [6]), могло способствовать уменьшению биологической продуктивности поверхностных вод и дальнейшему усилению сезонности формирования органического вещества над дрифтом Иоффе, что видно по сокращению доли видов-индикаторов низкого сезонного контраста в комплексах БФ. Более спокойная гидродинамическая обстановка реконструируется на основании уменьшения процентного содержания индикаторного вида G. subglobosa и по присутствию I. tumidula, не характерного для районов распространения сильных придонных течений (рис. 2; табл. 1). При этом вариации гранулометрического состава в этой же части колонки предполагают наличие трех циклов усиления и ослабления придонных течений (рис. 5, [42]). Данное расхождение может быть связано с небольшим градиентом изменчивости скоростей течений после ~0.28 млн лет, который, с одной стороны, приводил к вымыванию более тонких частиц из осадка, но, с другой стороны, был недостаточно сильным, чтобы оказывать существенное влияние на структуру комплексов бентосных организмов.
ЗАКЛЮЧЕНИЕ
Количественные анализы сообществ бентосных фораминифер по колонке АИ-2436 позволили реконструировать относительные вариации потока и периодичности поступления органического вещества на дно, а также изменения скоростей придонных течений. Кроме того, удалось предположить смену водных масс, омывавших вершину дрифта Иоффе в юго-западной Атлантике в плейстоценовое время. Показано, что поток органического вещества был низким в течение всего времени накопления осадка. Трансформированное органическое вещество, по-видимому, преобладало над лабильным, скорее всего, как за счет низкой биопродуктивности фотического слоя, так и вследствие вымывания тонкой фракции придонными течениями. Образование органического вещества в результате цветения фитопланктона происходило в условиях низкого сезонного контраста до среднеплейстоценового перехода и с выраженной сезонной периодичностью после него.
Доминирование толстостенных видов в комплексе бентосных фораминифер и характерных черт деформации раковин (вмятины, отверстия, сколы, царапины) некоторых видов по всей длине колонки послужило еще одним подтверждением гипотезы контуритовой природы осадков дрифта Иоффе. Выделены три группы деформаций, обнаруженных при визуальной оценке раковин видов N. umbonifer и C. wuellerstorfi и предположительно разного генезиса: (1) повреждения, вероятно, связанные с ударами о раковину перемещающихся компонентов осадка; (2) окатанные сколы и разрушенные последние камеры, по-видимому, обусловленные перемещением экземпляров на небольшие расстояния под действием придонных течений; (3) неокатанные сколы и истончение раковин, являющиеся результатом воздействия агрессивной придонной среды на карбонатные микрофоссилии. Результаты данного исследования показали хороший потенциал использования бентосных фораминифер в качестве вспомогательного индикатора при выделении контуритов. Однако это предположение требует проверки на других материалах из областей формирования контуритовых тел и осадочных волн.
По комплексам бентосных фораминифер в плейстоценовой истории контуритового дрифта Иоффе выделяется четыре этапа: (1) начало плейстоцена (до ~2.39 млн л.н.), характеризовавшееся, вероятно, слабовыраженным сезонным контрастом в поступлении органического вещества на дно, доминированием менее агрессивной по отношению к карбонатным компонентам осадка водной массы и изменчивыми скоростями придонных течений; (2) интервал ~2.39–1.6/0.92 млн л.н. по нанофоссилиям (или ~2.39–1.47/0.81 млн л.н. по планктонным фораминиферам), отличающийся от предыдущего усилением придонных течений за счет активного формирования палео-САГВ в Северной Атлантике; (3) интервал 0.92 (0.81)– 0.44/0.27 млн л.н., характеризующийся усилением сезонного контраста в поступлении органического вещества на дно, повышенными скоростями придонных течений, а также, вероятно, преобладанием более агрессивной палео-НЦГВ/ААДВ в районе исследования за счет увеличения продукции вод антарктического и субантарктического происхождения в течение длительных интервалов оледенений при установлении 100-тысячелетней ледниково-межледниковой цикличности; и (4) период последних 0.28 млн лет, когда при доминировании палео-НЦГВ/ААДВ установились более стабильные и спокойные условия с пониженными скоростями придонных течений.
Источники финансирования. Работа выполнена в рамках Госзадания ИО РАН (проект № 0128-2021-0006).
Благодарности. Авторы благодарят сотрудников Кабинета приборной аналитики ПИН РАН и М.А. Зенину за помощь с фотографированием видов, Е.В. Иванову и И.О. Мурдмаа за плодотворные дискуссии и конструктивную критику, а также рецензента статьи за ценные замечания и полезные рекомендации, позволившие улучшить данную статью.
Список литературы
Кулешова Л.А., Овсепян Е.А. Палеоокеанологические реконструкции для юго-западной части Атлантического океана в среднем-позднем плейстоцене на основе комплексов бентосных фораминифер // Вестник Московского Университета. Серия 5. География. № 3. С. 72–82.
Лукашина Н.П. Глубоководная циркуляция в канале Хантер (Юго-Западная Атлантика) в позднем плейстоцене и голоцене по бентосным фораминиферам // Океанология. 2019. Т. 59. № 1. С. 133–142.
Морозов Е.Г., Тараканов Р.Ю. Вытекание Антарктической донной воды из канала Вима // Докл. РАН. 2014. Т. 456. № 2. С. 227–230.
Мурдмаа И.О. Перерывы (hiatuses) в разрезах глубоководных отложений: вырванные придонными течениями страницы геологической летописи // Материалы XXIII Международной научной конференции (Школы) по морской геологии. М.: ИО РАН, 2019. Т. I. С. 136–139.
Altenbach A.V., Pflaumann U., Schiebel R. et al. Scaling percentages and distributional patterns of benthic foraminifera with flux rates of organic carbon // Journal of Foraminiferal Research. 1999. V. 29. P. 173–185.
Bell D.B., Jung S.J.A., Kroon D. The Plio-Pleistocene development of Atlantic deep-water circulation and its influence on climate trends // Quaternary Science Reviews. 2015. V. 123. P. 265–282.
Blatt H., Middleton G., Murray R. Origin of sedimentary rocks. New Jersey, Englewood Cliffs: Prentice-Hall Inc., 1980. 782 p.
Borisov D., Murdmaa I., Levchenko O., Frey D. Regional setting // The Ioffe Drift / Murdmaa I.O., Ivanova E.V. (Eds.). Switzerland: Springer, 2021. P. 7–20.
Borisov D., Levchenko O., Libina N. The geomorphology and seismic structure // The Ioffe Drift / Murdmaa I.O., Ivanova E.V. (Eds.). Switzerland: Springer, 2021. P. 37–52.
Cannariato K.G., Kennett J.P. Climatically related millennialscale fluctuations in strength of California margin oxygen-minimum zone during the past 60 k.y. // Geology. 1999. V. 27. № 11. P. 975–978.
Carman M.R., Keigwin L.D. Preservation and color differences in Nuttallides umbonifera // Journal of Foraminiferal Research. 2004. V. 34. № 2. P. 102–108.
Clark P.U., Archer D., Pollard D. et al. The middle Pleistocene transition: characteristics, mechanisms, and implications for long-term changes in atmospheric pCO2 // Quaternary Science Reviews. 2006. V. 25. P. 3150–3184.
de Almeida F.K., de Mello R.M., Costa K.B., Toledo F.A.L. The response of deep-water benthic foraminiferal assemblages to changes in paleoproductivity during the Pleistocene (last 769.2 kyr), western South Atlantic // Palaeogeography, Palaeoclimatology, Palaeoecology. 2015. V. 440. P. 201–212.
de Rijk S., Jorissen F.J., Rohling E.J., Troelstra S.R. Organic flux control on bathymetric zonation of Mediterranean benthic foraminifera // Marine Micropaleontology. 2000. V. 40. P. 151–166.
Diz P., Barker S. Approaches and constrains to the reconstruction of paleoproductivity from Cape Basin abyssal benthic foraminifera (South Atlantic) // Journal of Micropaleontology. 2016. V. 35. P. 195–204.
Garcia H.E., Locarnini R.A., Boyer T.P. et al. World Ocean Atlas 2013, V. 3: Dissolved Oxygen, Apparent Oxygen Utilization, and Oxygen Saturation / Levitus S. (Ed.). NOAA Atlas NESDIS 75, 2014. 27 p.
GEBCO Compilation Group (2021) GEBCO 2021 Grid (doi:).https://doi.org/10.5285/c6612cbe-50b3-0cff-e053-6c86abc09f8f
Gooday A.J. Deep-sea benthic foraminifera species which exploit phytodetritus: characteristic features and controls on distribution // Marine Micropaleontology. 1993. V. 22. P. 187–205.
Gooday A.J., Malzone M.G., Bett B.J., Lamont P.A. Decadal-scale changes on shallow-infaunal foraminiferal assemblages at the Porcupine Abyssal Plain, NE Atlantic // Deep-Sea Research II. 2010. V. 57. P. 1362–1382.
Gupta A.K., Thomas E. Initiation of Northern Hemisphere glaciation and strengthening of the northeast Indian monsoon: Ocean drilling program site 758, eastern equatorial Indian Ocean // Geology. 2003. V. 31. P. 47–50.
Fariduddin M., Loubere P. The surface ocean productivity response of deeper water benthic foraminifera in the Atlantic Ocean // Marine Micropaleontology. 1997. V. 32. P. 289–310.
Fatela F., Taborda R. Confidence limits of species proportions in microfossil assemblages // Marine Micropaleontology. 2002. V. 45. P. 169–174.
Ferreira F., Frontalini F., Leão C.J., Leipnitz I.I. Changes in the water column structure and paleoproductivity in the western South Atlantic Ocean since the middle Pleistocene: evidence from benthic and planktonic foraminifera // Quaternary International. 2014. V. 352. P. 111–123.
Jorissen F.J., Fontanier C., Thomas E. Paleoceanographical proxies based on deep-sea benthic foraminiferal assemblage characteristics // Proxies in Late Cenozoic Paleoceanography. V. 1. Developments in Marine Geology / C. Hillaire-Marcel, A. de Vernal (Eds.). New York: Elsevier, 2007. P. 263–325.
Ivanova E., Dmitrenko O. Micropaleontology and biostratigraphy // The Ioffe Drift / Murdmaa I.O., Ivanova E.V. (Eds.). Switzerland: Springer, 2021. P. 99–130.
Ivanova E., Murdmaa I., Borisov D. et al. Late Pliocene-Pleistocene stratigraphy and history of formation of the Ioffe calcareous contourite drift, Western South Atlantic // Marine Geology. 2016. V. 372. P. 17–30.
Ivanova E., Borisov D., Dmitrenko O., Murdmaa I. Hiatuses in the late Pliocene-Pleistocene stratigraphy of the Ioffe calcareous contourite drift, western South Atlantic // Marine and Petroleum Geology. 2020. V. 111. P. 624–637.
Ivanova E., Borisov D., Murdmaa I. Hiatuses and core correlations // The Ioffe Drift / Murdmaa I.O., Ivanova E.V. (Eds.). Switzerland: Springer, 2021. P. 145–160.
Ivanova E.V., Borisov D.G., Murdmaa I.O., Ovsepyan E.A., Stow D. Contourite systems around the northern exit from the Vema Channel // Marine Geology. 2022. V. 449. 106835.
Kender S., McClymont E.L., Elmore A.C. et al. Mid Pleistocene foraminiferal mass extinction coupled with phytoplankton evolution // Nature Communication. 2016. V. 7. 11970.
Kuijpers A. Southern Ocean circulation and global climate in the Middle Pleistocene (early Bruhnes) // Palaeogeography. Palaeoclimatology. Palaeoecology. 1989. V. 76. P. 67–83.
Kuleshova L.A., Bashirova L.D., Matul A.G. et al. Changing sea-surface and deep-water conditions in the southern Cape Verde Basin during the mid-Pleistocene to Holocene // Paleogeography. Palaeoclimatology. Palaeoecology. 2022. V. 594. 110921.
Loubere P., Fariduddin M. Quantitative estimation of global patterns of surface ocean biological productivity and its seasonal variation on timescales from centuries to millennia // Global Geochemical Cycles. 1999. V. 13. № 1. P. 115–133.
Lutze G.F., Thiel H. Cibicidoides wuellerstorfl and Planulina ariminensis, elevated epibenthic Foraminifera // Beobachtungen an Benthos-Foraminiferen / Altenbach A.V., Lutze G.F., Weinholz P. (Eds.). Ber. Sonderforschungsbereich 313, Univ. Kiel. 1987. V. 6. P. 17–30.
Mackensen A., Grobe H., Kuhn G., Fütterer D.K. Benthic foraminiferal assemblages from the eastern Weddell Sea between 68 and 73°S: distribution, ecology and fossilization potential // Marine Micropaleontology. 1990. V. 16. P. 241–283.
Mackensen A., Schmiedl G., Harloff J., Giese, M. Deep-sea foraminifera in the South Atlantic Ocean: Ecology and assemblage generation // Micropaleontology. 1995. V. 41. P. 342–358.
Marañón E., Holligan P.M., Varela M. et al. Basin-scale variability of phytoplankton biomass, production and growth in the Atlantic Ocean // Deep Sea Research I. 2000. V. 47. P. 825–857.
Meisling K.E., Cobbold P.R., Mount V.S. Segmentation of an obliquely rifted margin, Campos and Santos basins, southeastern Brazil // Am. Assoc. Petrol. Geol. Bull. 2001. V. 85. P. 1903–1924.
Melguen M., Thiede J. Facies distribution and dissolution depths of surface sediment components from the Vema Channel and the Rio Grande Rise (Southwest Atlantic Ocean) // Marine Geology. 1974. V. 17. P. 341–353.
Morozov E.G., Demidov A.N., Tarakanov R.Y., Zenk W. Abyssal Channels in the Atlantic Ocean. Dordrecht: Springer, 2010. 266 p.
Murdmaa I.O., Borisov D.G., Ivanova E.V. et al. The Ioffe calcareous contourite drift, Western South Atlantic // Book of Abstracts, 2nd Deep-Water Circulation Congress: the Contourite Log-book / Van Rooij D., Rüggeberg A. (Eds.). Ghent, Belgium, 10–12 September 2014. VLIZ Special Publication 69. Ghent University, Department of Geology and Soil Science – Vlaams Instituut voor de Zee – Flanders Marine Institute (VLIZ), Oostende, Belgium. 2014. P. 75–76.
Murdmaa I. Ivanova E., Borisov D. History of the Ioffe Drift // The Ioffe Drift / Murdmaa I.O., Ivanova E.V. (Eds.). Switzerland: Springer, 2021. P. 161–182.
Murgese D.S., De Dekker P. The distribution of deep-sea benthic foraminifera in core tops from the eastern Indian Ocean // Marine Micropaleontology. 2005. V. 56. P. 25–49.
Ovsepyan E.A., Ivanova E.V. Glacial-interglacial interplay of southern- and northern-origin deep waters in the São Paulo Plateau area of the western South Atlantic // Palaeogeography, Palaeoclimatology, Palaeoecology. 2019. V. 514. P. 349–360.
Peebles M.W., Lewis R.D. Surface textures of benthic foraminifera from San Salvador, Bahamas // Journal of Foraminiferal Research. 1991. V. 21. № 4. P. 285–292.
Poli M.S., Meyers P.A., Thunell R.C., Capodivacca M. Glacial-interglacial variations in sediment organic carbon accumulation and benthic foraminiferal assemblages on the Bermuda Rise (ODP Site 1063) during MIS 13 to 10 // Paleoceanography. 2021. V. 27. PA3216.
Rathburn A.E., Corliss B.H. The ecology of living (stained) deep-sea benthic foraminifera from the Sulu Sea // Paleoceanography. 1994. V. 9. № 1. P. 87–150.
Smart C.W., Gooday A.J. Recent benthic foraminifera in the abyssal Northeast Atlantic Ocean: relation to phytodetritus inputs // Journal of Foraminiferal Research. 1997. V. 27. № 2. P. 85–92.
Schmiedl G., Mackensen A. Late Quaternary paleoproductivity and deep water circulation in the eastern South Atlantic Ocean: Evidence from benthic foraminifera // Palaeogeography, Palaeoclimatology, Palaeoecology. 1997. V. 130. P. 43–80.
Tetard M., Licari L., Ovsepyan E. et al. Toward a global calibration for quantifying past oxygenation in oxygen minimum zones using benthic Foraminifera // Biogeosciences. 2021. V. 18(9). P. 2827–2841.
Wetmore K.L. Correlations between test strength, morphology and habitat in some benthic foraminifera from the coast of Washington // Journal of Foraminiferal Research. 1987. V. 17. P. 1–13.
Wollenburg J.E., Mackensen A. Living benthic foraminifers from the central Arctic Ocean: faunal composition, standing stock and diversity // Marine Micropaleontology. 1998. V. 34. P. 153–185.
Дополнительные материалы отсутствуют.