

Палеонтологический журнал, 2019, № 1, стр. 87-102
Ископаемые древесины Xenoxylon (Coniferales) из юрских и меловых отложений Сибири и российского Дальнего Востока
a Федеральный научный центр биоразнообразия наземной биоты Восточной Азии ДВО РАН
Владивосток, Россия
b Ботанический институт им. В.Л. Комарова РАН
Санкт-Петербург, Россия
* E-mail: afmaxim@inbox.ru
Поступила в редакцию 08.12.2017
Принята к публикации 23.03.2018
Аннотация
По анатомическим признакам ископаемой древесины из местонахождений мелового возраста Сибири и российского Дальнего Востока описаны представители морфорода Xenoxylon: X. latiporosum, X. phyllocladoides, X. japonicum, X. meisteri и X. nariwaense. Виды X. japonicum и X. nariwaense указываются впервые для России, а X. meisteri – впервые для российского Дальнего Востока. Расширено представление о стратиграфическом распространении X. japonicum и X. nariwaense, которые ранее были описаны только из верхнетриасовых отложений. Приводится полный ревизованный список ископаемых древесин Xenoxylon, описанных из юрских и меловых отложений Сибири и российского Дальнего Востока.
ВВЕДЕНИЕ
Морфород Xenoxylon Gothan был установлен В. Готаном (Gothan, 1905) для ископаемых древесин хвойных, которые впервые были описаны С. Крамером (Cramer, 1868) как Pinites latiporosus Cramer и P. pauciporosus Cramer из юрских отложений о-ва Западный Шпицберген (Норвегия). Находки ископаемых древесин Xenoxylon многочисленны и указываются из мезозойских отложений по всему Северному полушарию. Хвойные с древесиной типа Xenoxylon были широко распространены начиная с карния (поздний триас) по маастрихт (поздний мел) включительно (Philippe et al., 2013). К сожалению, несмотря на большое количество данных по Xenoxylon, его систематическое положение среди хвойных до сих пор остается дискуссионным. В.Р. Мюллер-Штоль (Muller-Stoll, 1987) и некоторые другие исследователи даже выражали сомнение по поводу отнесения Xenoxylon к хвойным. А.В. Ярмоленко (1933), И.В. Бэйли (Bailey, 1953) и Р. Худайбердыев с соавт. (1971) на основании сходства в анатомическом строении древесины предполагали, что представители Xenoxylon имеют близкие родственные связи с какими-то древними хвойными, давшими начало семейству Sciadopityaceae. Такого же мнения придерживался и В.Д. Нащокин (1968). Он, однако, полагал, что Xenoxylon включает также и ископаемые древесины, имеющие черты сходства с древесиной некоторых современных представителей Podocarpaceae (роды Dacrydium Solander и Phyllocladus Rich et Mirbel). Отчасти это подтверждают и данные органо-геохимических исследований. Так Л. Мариновский с соавт. (Marynowski et al., 2008) на основе химического анализа образцов Xenoxylon из юрских отложений Польши установили, что Xenoxylon очень близок к современным хвойным Cupressaceae s. l. и Podocarpaceae. М. Филипп с соавт. (Philippe et al., 2013) высказали предположение, что Xenoxylon могут принадлежать вымершим хвойным Miroviaceae, на основании совместных находок и сходного географического и стратиграфического распространения этих ископаемых растений.
В разные годы было описано около 30 видов Xenoxylon (Philippe et al., 2013). Филипп с соавт. (Philippe et al., 2013) провели таксономическую ревизию этого морфорода и разделили его на три неформальные анатомические группы, включив следующие виды: группа Latiporosum (виды X. conchylianum Fliche, X. latiporosum, X. peideensis Zheng et Zhang, X. yixianense Zhang et Shang, X. watarianum M. Nishida et Nishida), группа Phyllocladoides (виды X. hopeiense Chang, X. huttonianum (Witham) Philippe et Hayes, X. jakutiense Shilkina, X. phyllocladoides Gothan) и группа Meisteri (виды X. fuxinense Ding, X. huolinhense Ding, X. japonicum Vogellehner ex Suzuki et Terada, X. liaoningense Duan et Wang, X. meisteri Palibin et Jarmolenko, X. nariwaense Yamazaki, Tsunada et Koike, X. suljuctense Shilkina et Chudajberdiev). Эти три группы Xenoxylon в основном отличаются по характеру поровости на стенках трахеид (Philippe et al., 2013). Для представителей группы Latiporosum характеры строго однорядные уплощенные и сомкнутые поры на радиальных стенках трахеид, очень редко и только в узких трахеидах – однорядные округлые, сближенные и свободно расположенные. У видов группы Phyllocladoides встречаются, в основном, однорядные округлые, свободно расположенные поры на радиальных стенках трахеид, и только местами – однорядные уплощенные и сомкнутые. Представители группы Meisteri часто имеют однорядные уплощенные и сомкнутые поры на радиальных стенках трахеид, которые расположены в группах; иногда у них встречаются двурядные очередные и супротивные поры на радиальных стенках трахеид. Кроме того, для Meisteri, в отличие от Latiporosum и Phyllocladoides, характерны многочисленные одно-, дву- и трехрядные поры на тангентальных стенках трахеид.
Особое место среди представителей Xenoxylon занимает X. parvipunctatum Vogellehner. От всех вышеперечисленных групп Xenoxylon этот вид отличается наличием однорядных, небольших, слегка уплощенных пор на радиальных стенках трахеид, расположенных в непрерывных рядах, и двурядных очередных пор на радиальных стенках трахеид, а также полным отсутствием пор на тангентальных стенках трахеид (Philippe et al., 2013).
Позднее из верхнетриасовых отложений Китая были описаны новые находки Xenoxylon, X. guangyuanense Tian, Wang et Philippe (Tian et al., 2016) и X. junggarensis Wan, Zhou, Tang, Liu et Wang (Wan et al., 2016), которые по анатомическому строению близки к группе Meisteri.
До настоящего времени из юрских и меловых отложений Сибири и российского Дальнего Востока (РДВ) были описаны виды X. barberi Seward (Шилкина, Худайбердыев, 1971), X. hopeiense (Шилкина, Худайбердыев, 1971; Афонин, 2008), X. jakutiense (Шилкина, 1986), X. jurassicum (Eckhold) Kräusel (Нащокин, 1968), X. latiporosum (Нащокин, 1962, 1968; Дорофеюк, Филин, 1969; Шилкина, Худайбердыев, 1971; Афонин, 2008), X. meisteri (Палибин, Ярмоленко, 1932), X. phyllocladoides (Afonin, Philippe, 2014), X. tomiense Larishchev (Ларищев, 1957) и X. watarianum (Nishida, Nishida, 1986).
В представленной работе описываются новые находки, и дается обзор всех ископаемых древесин Xenoxylon из юрских и меловых отложений Сибири и РДВ. Кроме того, приводятся результаты ревизии древесных остатков Xenoxylon из юрских и меловых отложений Сибири по коллекциям И.А. Шилкиной (место хранения – Ботанический институт им. В.Л. Комарова (БИН) РАН, г. Санкт-Петербург) и Нащокина (место хранения – Институт леса им. В.Н. Сукачева (ИЛ) СО РАН, г. Красноярск).
Автор выражает благодарность д-ру М. Филиппу (Лионский ун-т, г. Лион, Франция) за консультации в ходе исследования, В.С. Маркевич, Е.В. Бугдаевой, Е.Б. Волынец (ФНЦ Биоразнообразия ДВО РАН, г. Владивосток), Л.П. Эйхвальд (“Хабаровскгеология”, г. Хабаровск), С.А. Медведевой (Ин-т тектоники и геофизики ДВО РАН им. Ю.А. Косыгина, г. Хабаровск) за предоставленный для исследования материал и литературу по геологии местонахождений, Н.В. Носовой (БИН РАН, г. Санкт-Петербург) и В.Е. Беньковой (ИЛ СО РАН, г. Красноярск) за помощь в работе с коллекциями Шилкиной и Нащокина, А.П. Ляховой (Дальневосточный геологический институт ДВО РАН, г. Владивосток) за изготовление шлифов, анонимным рецензентам настоящей рукописи за ценные замечания.
Работа поддержана грантом РФФИ № 17-04-01582 а.
МАТЕРИАЛ И МЕТОДЫ
Исследованные ископаемые древесины Xenoxylon были собраны в разные годы автором и Е.Б. Волынец (ФНЦ Биоразнообразия ДВО РАН, г. Владивосток) на западном побережье оз. Ханка, п-ове Де-Фриз и п-ове Муравьева-Амурского в Южном Приморье, Л.П. Эйхвальд (“Хабаровскгеология”, г. Хабаровск) на левом берегу р. Амур в Нижнем Приамурье и Е.В. Бугдаевой (ФНЦ Биоразнообразия ДВО РАН, г. Владивосток) в Гусиноозерской впадине в Забайкалье (табл. 1).
Таблица 1.
Местонахождения исследованных ископаемых древесин Xenoxylon из меловых отложений Сибири и российского Дальнего Востока
| Местонахождение | № образца | Размеры образцов: длина, в см/в поперечнике, в см | Кол-во шлифов | Свита/подсвита | Ярус | |
|---|---|---|---|---|---|---|
| Западное побережье оз. Ханка, Южное Приморье, российский Дальний Восток | 30/1-1–30/1-3* | 8/5 × 6 | 10 | галенковская | альб | |
| Мыс Тихий, п-ов Де-Фриз, Южное Приморье, российский Дальний Восток | 32C/2 | 15/2 × 8 | 3 | галенковская | альб | |
| 32C/3 | 28/10 × 15 | 3 | ||||
| п-ов Муравьева-Амурского, Южное Приморье, российский Дальний Восток | Правый берег р. Песчанка | 32B/1 | 16/12 × 15 | 3 | галенковская | альб |
| 32B/2 | 14/8 × 10 | 3 | ||||
| 32B/5 | 18/7 × 8 | 3 | ||||
| 32B/6 | 23/14 × 18 | 3 | ||||
| 32B/8 | 12/6 × 9 | 3 | ||||
| 32B/11-1–32B/11-23* | 64/7 × 10 | 24 | ||||
| 32B/12 | 11/4 × 6 | 3 | ||||
| 32B/15 | 10/3 × 5 | 3 | ||||
| Мыс Клыкова | 32/6 | 10/6 × 8 | 3 | липовецкая | апт | |
| 32/9 | 12/8 × 11 | 3 | ||||
| 32/18 | 12/4 × 7 | 7 | ||||
| 32/25 | 7/4 × 5 | 3 | ||||
| 32/56-1–32/56-7* | 58/14 × 16 | 12 | ||||
| Мыс Фирсова | 32A/5 | 14/11 × 12 | 3 | липовецкая | апт | |
| 32A/8 | 12/6 × 7 | 3 | ||||
| 32A/13 | 8/4 × 9 | 3 | ||||
| 32A/16-1–32A/16-11* | 26/15–18 × 28–35 | 16 | ||||
| Левый берег р. Амур, междуречье рек Дурал и Хальзанка, Нижнее Приамурье, российский Дальний Восток | 27-1 | 12/4 × 6 | 7 | ситогинская/ нижнеситогинская | апт-альб | |
| Гусиноозерская впадина, Забайкалье, Восточная Сибирь | Tb1 | 16/4 × 5 | 3 | селенгинская/ верхнеселенгинская | баррем | |
На западном побережье оз. Ханка ископаемые древесины содержатся в нескольких слоях туфовых алевролитов и туфов галенковской свиты, возраст которой по палеоботаническим (Красилов, 1967; Волынец, 2006) и палинологическим (Маркевич, 1995) данным установлен как альб.
На п-ове Де-Фриз древесные остатки были собраны на мысе Тихий из слоя мелкозернистых песчаников и алевролитов галенковской свиты.
На п-ове Муравьева-Амурского ископаемые древесины были обнаружены на правом берегу р. Песчанка (рядом с шоссе Владивосток-Уссурийск) в мелкозернистых песчаниках и алевролитах галенковской свиты, на мысе Клыкова в крупно- и среднезернистых песчаниках липовецкой свиты и на мысе Фирсова в мелкозернистых песчаниках липовецкой свиты. Возраст липовецкой свиты по палеоботаническим (Красилов, 1967) и палинологическим (Маркевич, 1995) данным установлен как апт. По мнению Волынец (2006), возраст липовецкой свиты – апт–начало раннего альба.
На левом берегу р. Амур (1.5 км выше устья р. Дурал, междуречье рек Дурал и Хальзанка) древесные остатки были собраны из слоя цемента конгломератов нижнеситогинской подсвиты ситогинской свиты. Нижнеситогинская подсвита на основании изучения морских фоссилий датируется апт-альбом (Жаров и др., 2004).
В Гусиноозерской впадине (рядом с сопкой Баян-Зурхе) в Забайкалье ископаемые древесины происходят из песчанистых алевролитов верхнеселенгинской подсвиты селенгинской свиты. Возраст верхнеселенгинской подсвиты по палинологическим данным и на основании изучения пресноводных фоссилий установлен как баррем (Скобло и др., 2001).
Изученные ископаемые древесины очень плотные, минерализованные, часто окремнелые, от серого до черного цвета, и представляют собой фрагменты стволов или побегов. Размеры этих фрагментов от 0.07 до 0.64 м длиной (табл. 1).
При исследовании ископаемых древесин была использована методика изготовления прозрачных шлифов из плотной минерализованной древесины, изложенная в работе А.Ф. Гаммерман с соавт. (1946). Всего было изготовлено и исследовано 124 шлифа (табл. 1). Микроскопическое изучение шлифов и микрофотографирование анатомических структур проводилось с помощью световых биологических микроскопов “Primo Star” и “AxioScop-40”, а также фотокамеры “AxioCamHR” производства “Carl Zeiss”. Описание анатомического строения древесины выполнено с использованием терминологии, изложенной в работе А.А. Яценко-Хмелевского (1954) и “IAWA List …” (2004). Определение ископаемых древесин проводилось с помощью опубликованных работ с ключами и описанием ископаемых древесин Xenoxylon: Шилкина, Худайбердыев, 1971; Philippe, Bamford, 2008; Philippe et al., 2013 и др. Изученные образцы ископаемой древесины из коллекций №№ 27, 30, 32, 32A, 32B, 32C, Tb хранятся в ФНЦ Биоразнообразия ДВО РАН (г. Владивосток).
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
CONIFERALES
Морфород Xenoxylon Gothan, 1905
Группа Latiporosum
Xenoxylon latiporosum (Cramer) Gothan, 1905
Описание (рис. 1, а–е). Годичные кольца отчетливые, с одним-тремя слоями поздней древесины; переход от ранней древесины к поздней резкий (рис. 1, а). Ширина годичных колец 0.8–4.0 мм. Смоляные ходы и тяжевая паренхима отсутствуют.
Рис. 1.
Xenoxylon latiporosum (Cramer) Gothan, обр. № 30/1-1, зап. побережье оз. Ханка, российский Дальний Восток, галенковская свита, альб: а – поперечный срез, годичные кольца, резкий переход от ранней древесины к поздней; б – радиальный срез, однорядные поры на стенках трахеид; в, г – радиальный срез, однорядные поры на стенках трахеид и оконцевые поры на полях перекреста; д, е – тангентальный срез, однорядные лучи, однорядные поры на стенках трахеид и тиллы в трахеидах.

Лучи однорядные, 1–24 клетки высотой, чаще 5–18 клеток (рис. 1, д, е). Горизонтальные и тангентальные стенки лучей гладкие, непористые. Поры на тангентальных стенках трахеид встречаются редко; поры однорядные, округлые и свободно расположенные, 10–11 мкм в диаметре (рис. 1, д, е).
Поры на радиальных стенках трахеид ранней древесины однорядные, уплощенные и сомкнутые, изредка слегка округлые, свободно расположенные (рис. 1, б–г); размер пор – 18–22 × 20–37 мкм. Крассулы и лучевые трахеиды отсутствуют. На полях перекреста одна (реже две) крупные, простые поры оконцевого типа (рис. 1, в, г); размер пор – 18–25 × 24–45 мкм. Тиллы иногда встречаются в трахеидах (рис. 1, д, е).
Сравнение. Среди ископаемых древесин Xenoxylon сходство у исследованных нами древесных остатков наблюдается с древесиной X. latiporosum из группы Latiporosum (Philippe et al., 2013). Это сходство отмечается в наличии крупных, однорядных, в основном уплощенных, сомкнутых, иногда в свободном расположении пор на радиальных стенках трахеид ранней древесины, изредка пористых тангентальных стенок трахеид, а также однорядных лучей (до 24 клеток высотой) без травматических смоляных ходов. Небольшое отличие от типового материала наблюдается лишь по некоторым количественным признакам (размер пор, высота лучей).
Материал. Три образца от одного фрагмента ископаемой древесины, №№ 30/1-1 – 30/1-3.
Местонахождение. Западное побережье оз. Ханка, российский Дальний Восток, галенковская свита, нижний мел, альб.
Группа Phyllocladoides
Xenoxylon phyllocladoides Gothan, 1906
Xenoxylon barberi: Шилкина, Худайбердыев, 1971, с. 125–127, табл. 79 , фиг. 1–3.
Описание (рис. 2, а–ж). Годичные кольца отчетливые, приблизительно 1.0–4.8 мм шириной, часто сильно смяты; переход от ранней древесины к поздней резкий (рис. 2, а). Смоляные ходы и тяжевая паренхима отсутствуют.
Рис. 2.
Xenoxylon phyllocladoides Gothan, обр. № 32/18, мыс Клыкова, п-ов Муравьева-Амурского, российский Дальний Восток, липовецкая свита, апт: а – поперечный срез, годичные кольца, резкий переход от ранней древесины к поздней; б–д – радиальный срез, однорядные поры на стенках трахеид и оконцевые поры на полях перекреста; е – тангентальный срез, однорядные лучи, однорядные поры на стенках трахеид и тиллы в трахеидах; ж – тангентальный срез, однорядные лучи, луч с двумя травматическими карманами и тиллы в трахеидах.
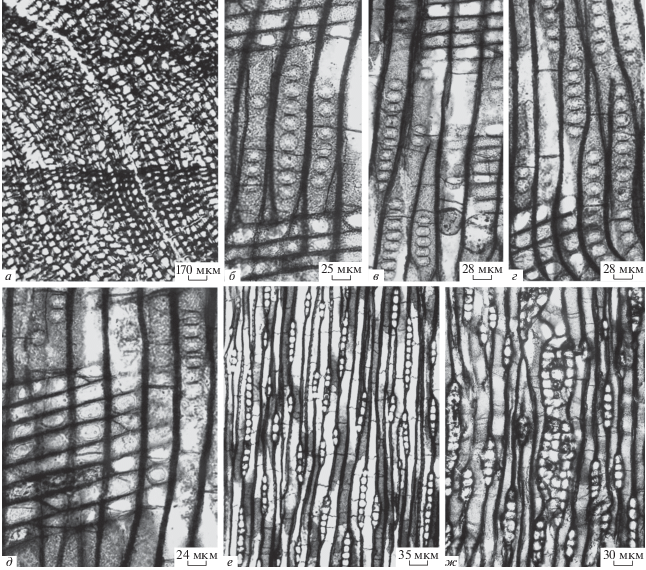
Лучи однорядные, изредка с короткими двурядными участками (один-четыре слоя), 1–14(25) клеток высотой (рис. 2, е, ж). Небольшая травматическая область обнаружена на тангентальном сечении, которая содержит несколько лучей с травматическими карманами (рис. 2, ж). Горизонтальные и тангентальные стенки клеток лучей гладкие, непористые. На тангентальных стенках трахеид изредка встречаются однорядные, округлые, свободно расположенные поры (рис. 2, е, ж), 8–12 мкм в диаметре.
На радиальных стенках трахеид ранней древесины имеются однорядные, округлые, овальные, свободно и сближено распложенные поры (рис. 2, б–д), 18–22 мкм в диаметре; местами встречаются уплощенные поры (рис. 2, б–д), 14–18 × 22–28 мкм в размере. Крассулы и лучевые трахеиды отсутствуют. На полях перекреста одна, изредка две крупные, простые или слегка окаймленные поры оконцевого типа (рис. 2, б–д); размер пор 12–17 × 19–35 мкм. Тиллы иногда встречаются в трахеидах (рис. 2, е, ж).
Сравнение. Среди ископаемых древесин Xenoxylon исследованная древесина имеет сходство с X. phyllocladoides из группы Phyllocladoides (Philippe et al., 2013). Этот вид характеризуется наличием округлых, свободно расположенных, изредка округлых, сближенных или уплощенных, сомкнутых пор на радиальных стенках трахеид, редких пор на тангентальных стенках трахеид, простых или слегка окаймленных пор оконцевого типа на полях перекреста, которые в два (три) раза больше по ширине, чем по высоте, а также отсутствием тяжевой паренхимы и крассул между порами на радиальных стенках трахеид (Philippe et al., 2013).
Материал. Обр. № 32/18.
Местонахождение. Мыс Клыкова, п‑ов Муравьева-Амурского, российский Дальний Восток, липовецкая свита, нижний мел, апт.
Группа Meisteri
Xenoxylon japonicum Vogellehner ex Suzuki et Terada, 1992
Описание (рис. 3, а–е; 4, а–е). Годичные кольца отчетливые, с одним-тремя слоями поздней древесины, 1–2.4 мм шириной; переход от ранней древесины к поздней резкий (рис. 3, а). Смоляные ходы и тяжевая паренхима отсутствуют.
Рис. 3.
Xenoxylon japonicum Vogellehner ex Suzuki et Terada, обр. № T-b1, Гусиноозерская впадина, Забайкалье, селенгинская свита, верхнеселенгинская подсвита, баррем: а – поперечный срез, годичные кольца, резкий переход от ранней древесины к поздней; б, в – радиальный срез, однорядные поры на стенках трахеид и тиллы в трахеидах; г – радиальный срез, однорядные поры на стенках трахеид, оконцевые поры на полях перекреста и тиллы в трахеидах; д – тангентальный срез, однорядные лучи и тиллы в трахеидах; е – тангентальный срез, однорядные лучи, двурядный луч и тиллы в трахеидах.

Лучи однорядные, изредка с короткими двурядными участками (один–четыре слоя), 1–54 клетки высотой, чаще 10–30 клеток (рис. 3, д, е). Обнаружен один двурядный луч до 15 слоев клеток высотой (рис. 3, е). Иногда встречаются очень высокие однорядные лучи, до 60 клеток высотой (рис. 4, д, е). Горизонтальные и тангентальные стенки клеток лучей гладкие, непористые. Поры на тангентальных стенках трахеид встречаются довольно часто; поры однорядные, округлые, свободно расположенные, 10–14 мкм в диаметре. Иногда встречаются многочисленные однорядные, двурядные, изредка трехрядные поры на тангентальных стенках трахеид (рис. 4, е).
Рис. 4.
Xenoxylon japonicum Vogellehner ex Suzuki et Terada, обр. № 32C/2, м. Тихий, п-ов Де-Фриз, российский Дальний Восток, галенковская свита, альб: а – поперечный срез, годичные кольца, резкий переход от ранней древесины к поздней; б, в – радиальный срез, однорядные поры на стенках трахеид и тиллы в трахеидах; г – радиальный срез, однорядные поры на стенках трахеид и оконцевые поры на полях перекреста; д – тангентальный срез, однорядные лучи и тиллы в трахеидах; е – тангентальный срез, однорядные лучи, поры на стенках трахеид, тиллы в трахеидах.

Поры на радиальных стенках трахеид ранней древесины однорядные, часто уплощенные, сомкнутые (рис. 3, б–г), 14–18 × 24–36 мкм в размере, изредка округлые, свободно расположенные или в группах (рис. 3, б–г), 20–23 мкм в диаметре. Обычно по краям групп поры более мелкие и округлые. Крассулы и лучевые трахеиды отсутствуют. На полях перекреста одна, изредка две крупные, простые поры оконцевого типа (рис. 3, г); размер пор 18–24 × 25–50 мкм. Тиллы часто встречаются в трахеидах (рис. 3, б–е).
Сравнение. Сравнение изученного материала с ранее описанными древесинами Xenoxylon показало, что наибольшее сходство наблюдается с древесиной X. japonicum, которая впервые была описана М. Шимакурой (Shimakura, 1936) как X. latiporosum из верхнетриасовых отложений Северной Кореи. Основными признаками этого вида являются наличие однорядных, сближенных, уплощенных, изредка округлых, свободно расположенных или в группах пор на радиальных стенках трахеид, многочисленных пор на тангентальных стенках трахеид и местами очень высоких однорядных лучей (Vogellehener, 1968; Suzuki, Terada, 1992; Philippe et al., 2013). Следует отметить, что М. Сузуки и К. Терада (Suzuki, Terada, 1992) выбрали лектотип для X. japonicum, который характеризуется отсутствием поровости на тангентальных стенках трахеид. Образцы, отнесенные Д. Фогеленером (Vogellehener, 1968) к этому виду, имеют, однако, обильную тангентальную поровость.
Материал. Три образца, №№ T-b1, 32C/2, 32C/3.
Местонахождения. Гусиноозерская впадина, Забайкалье, Восточная Сибирь, селенгинская свита, верхнеселенгинская подсвита, нижний мел, баррем; п-ов Де-Фриз, российский Дальний Восток, галенковская свита, нижний мел, альб.
Xenoxylon meisteri Palibin et Jarmolenko, 1932
Xenoxylon latiporosum: Шилкина, Худайбердыев, 1971, с. 129–131, табл. 76 , фиг. 1–5, табл. 77 , фиг. 3–5; Нащокин, 1962, с. 284, табл. 26 , фиг. 4–22; 1968, с. 64, табл. 11, 12 .
Описание (рис. 5, а–е; 6, а–е). Годичные кольца отчетливые, с одним-тремя слоями трахеид поздней древесины; переход от ранней древесины к поздней резкий (рис. 5, а). Ширина годичных колец от 0.8 до 3.8 мм. Смоляные ходы и тяжевая паренхима отсутствуют.
Рис. 5.
Xenoxylon meisteri Palibin et Jarmolenko, обр. №32B/6, прав. берег р. Песчанка, п-ов Муравьева-Амурского, российский Дальний Восток, галенковская свита, альб: а – поперечный срез, годичные кольца, резкий переход от ранней древесины к поздней; б, в – радиальный срез, однорядные поры на стенках трахеид, оконцевые поры на полях перекреста и тиллы в трахеидах; г – радиальный срез, однорядные поры на стенках трахеид; д, е – тангентальный срез, однорядные лучи, поры на стенках трахеид, тиллы в трахеидах.

Лучи однорядные, изредка с двурядными участками (один-шесть слоев клеток), 1–37 клеток высотой, чаще 7–18 клеток (рис. 5, д, е). Горизонтальные и тангентальные стенки лучей гладкие, непористые. На тангентальных стенках трахеид многочисленные, однорядные, двурядные и изредка трехрядные, округлые поры, 8–16 мкм в диаметре (рис. 5, д, е). Однорядные поры свободно расположенные и сближенные; двурядные и трехрядные – очередные и субсупротивные.
Поры на радиальных стенках трахеид однорядные (рис. 5, б–г). Поры в основном уплощенные и сомкнутые, 15–21 × 22–37 мкм в размере или изредка округлые, свободно расположенные или в группах, 20–22 мкм в диаметре. Обнаружен небольшой участок двурядных пор на радиальных стенках трахеид; поры округло-многоугольные, очередные, 18–24 мкм в диаметре (рис. 6, в). Крассулы и лучевые трахеиды отсутствуют. На полях перекреста одна, изредка две крупные, простые поры оконцевого типа (рис. 5, б). Размер пор 14–24 × 25–46 мкм. Тиллы часто встречаются в трахеидах (рис. 5, б–е).
Рис. 6.
Xenoxylon meisteri Palibin et Jarmolenko, обр. № 32A/16-1, м. Фирсова, п-ов Муравьева-Амурского, российский Дальний Восток, липовецкая свита, апт: а – поперечный срез, годичные кольца, резкий переход от ранней древесины к поздней, б – радиальный срез, однорядные поры на стенках трахеид и тиллы в трахеидах; в – радиальный срез, однорядные и двурядные поры на стенках трахеид и тиллы в трахеидах; г – радиальный срез, однорядные поры на стенках трахеид, оконцевые поры на полях перекреста и тиллы в трахеидах; д – тангентальный срез, однорядные лучи и тиллы в трахеидах; е – тангентальный срез, однорядные лучи, поры на стенках трахеид, тиллы в трахеидах.

Сравнение. Среди ископаемых древесин Xenoxylon наибольшее сходство у изученных нами древесных остатков наблюдается с древесиной X. meisteri, которая впервые была описана из меловых отложений Забайкалья (Палибин, Ярмоленко, 1932). Отличие от типового материала X. meisteri наблюдается лишь в высоте лучей. Палибин и Ярмоленко (1932) указывают для X. meisteri в основном однорядные лучи от трех до восьми клеток высотой, местами до 18 клеток. У изученных нами ископаемых древесин однорядные лучи в основном от 3 до 20 клеток высотой, изредка до 37 клеток (табл. 2). Несколько образцов имеют однорядные лучи до 42 или 52 клеток высотой (табл. 2). Ряд исследованных образцов характеризуются наличием невысоких лучей как у X. meisteri, но очень обильной поровости на тангентальных стенках трахеид как у X. japonicum (табл. 2). По-видимому, X. meisteri и X. japonicum из группы Meisteri очень близки и их отличительные признаки выражены слабо.
Материал. 13 образцов, №№ 32/6, 32/9, 32/25, 32A/5, 32A/8, 32A/13, 32B/1, 32B/2, 32B/5, 32B/6, 32B/8, 32B/12, 32B/15; семь образцов от одного фрагмента, №№ 32/56-1–32/56-7; 11 образцов от одного фрагмента, №№ 32A/16-1–32A/16-11; 23 образца от одного фрагмента, №№ 32B/11-1–32B/11-23.
Местонахождения. П-ов Муравьева-Амурского, российский Дальний Восток: м. Фирсова и м. Клыкова, липовецкая свита, нижний мел, апт; правый берег р. Песчанка, галенковская свита, нижний мел, альб.
Таблица 2.
Сравнительная характеристика высоты лучей и поровости на тангентальных стенках трахеид образцов ископаемой древесины X. meisteri из нижнемеловых отложений п-ова Муравьева-Амурского, российский Дальний Восток
| № образца | Лучи | Поры на тангентальных стенках трахеид | |||||
|---|---|---|---|---|---|---|---|
| высота (в клетках) |
двурядные участки (в клетках) | однорядные | двурядные | трехрядные | встречаемость | ||
| 32/6 | 1–15(22) | 1–3(7) | + | + − | − | много | |
| 32/9 | 1–12(20) | 1–3 | + | + − | − | мало | |
| 32/25 | 1–10(28) | 1–2 | + | + − | − | мало | |
| один ствол | 32/56-2 | 1–20(52) | 1–4(6) | + | + | − | мало |
| 32/56-4 | 1–18(30) | 1–3 | + | + | − | мало | |
| 32/56-7 | 1–20(34) | 1–5 | + | + | − | мало | |
| 32A/5 | 1–10(33) | 1–2 | + | + − | − | мало | |
| 32A/8 | 1–10(20) | 1–3 | + | + | + − | много | |
| 32A/13 | 1–10(28) | 1–2 | + | + | + − | много | |
| один ствол | 32A/16-1 | 1–15(23) | 1–4 | + | + | + − | много |
| 32A/16-4 | 1–12(25) | 1–3 | + | + | + − | много | |
| 32A/16-11 | 1–12(27) | 1–3 | + | + | + − | много | |
| 32B/1 | 1–15(26) | 1–3 | + | + − | − | мало | |
| 32B/2 | 1–18(24) | 1–3 | + | + − | − | мало | |
| 32B/5 | 1–10(28) | 1–3(6) | + | + − | − | мало | |
| 32B/6 | 1–18(37) | 1–3(6) | + | + | + − | много | |
| 32B/8 | 1–15(42) | 1–6 | + | + − | + − | много | |
| 32B/12 | 1–14(32) | 1–3 | + | + − | + − | много | |
| 32B/15 | 1–15(34) | 1–6(14) | + | + | + − | много | |
| один ствол | 32B/11-1 | 1–15(37) | 1–3 | + | + | + − | много |
| 32B/11-4 | 1–12(23) | 1–3 | + | + | + − | много | |
| 32B/11-5 | 1–14(28) | 1–4 | + | + | − | много | |
| 32B/11-14 | 1–15(35) | 1–2 | + | + | − | много | |
| 32B/11-15 | 1–14(32) | 1–3 | + | + | + − | много | |
Xenoxylon nariwaense Yamazaki, Tsunada et Koike, 1980
Описание (7, а–ж). Годичные кольца отчетливые, с одним-тремя слоями трахеид поздней древесины; переход от ранней древесины к поздней резкий (рис. 7, а). Ширина годичных колец варьирует от 1 до 3 мм. Смоляные ходы и тяжевая паренхима отсутствуют.
Рис. 7.
Xenoxylon nariwaense Yamazaki, Tsunada et Koike, обр. № 27-1, лев. берег р. Амур, междуречье рек Дурал и Хальзанка, российский Дальний Восток, ситогинская свита, нижнеситогинская подсвита, апт–альб: а – поперечный срез, годичные кольца, резкий переход от ранней древесины к поздней; б–д – радиальный срез, однорядные и двурядные поры на стенках трахеид; е – радиальный срез, однорядные поры на стенках трахеид и оконцевые поры на полях перекреста; ж – тангентальный срез, однорядные лучи, однорядные поры на стенках трахеид и тиллы в трахеидах.

Лучи однорядные, изредка с короткими двурядными участками (один–три слоя клеток), 1–15 клеток высотой, чаще 4–8 клеток (рис. 7, ж). Горизонтальные и тангентальные стенки лучей гладкие, непористые. На тангентальных стенках трахеид поры сохранились плохо; видимые поры однорядные, округлые, свободно расположенные и сближенные, изредка двурядные, очередные, 8–10 мкм в диаметре (рис. 7, ж).
Поры на радиальных стенках трахеид однорядные и двурядные (рис. 7, б–д). Однорядные поры округлые, свободно расположенные и сближенные, 19–24 мкм в диаметре или уплощенные, сомкнутые и в группах, 15–19 × 21–29 мкм в размере. Двурядные поры округлые, округло-многоугольные, очередные, ложносупротивные или супротивные, 15–26 мкм в диаметре; местами поры образуют параллельные ряды почти округлых пор (рис. 7, б–д). Двурядные поры иногда располагаются по всей длине стенок трахеид. Крассулы и лучевые трахеиды отсутствуют. На полях перекреста одна, изредка две крупные, простые, оконцевые поры (рис. 7, е), 14–25 × 20–54 мкм в размере. Тиллы иногда встречаются в трахеидах (рис. 7, ж).
Сравнение. Среди ископаемых древесин Xenoxylon наибольшее сходство наблюдается с древесиной X. nariwaense, которая впервые была описана из верхнетриасовых отложений Японии (Yamazaki et al., 1980; Yamazaki, Tsunada, 1982). Исследованная нами древесина отличается от типового материала X. nariwaense наличием более обильной двурядной поровости на радиальных стенках трахеид.
Материал. Образец № 27-1.
Местонахождение. Левый берег р. Амур, междуречье рек Дурал и Хальзанка, российский Дальний Восток, ситогинская свита, нижнеситогинская подсвита, нижний мел, апт–альб.
ОБСУЖДЕНИЕ
До наших исследований из Сибири и РДВ было описано девять видов Xenoxylon: X. barberi из юрских и нижнемеловых отложений Восточной Сибири (Шилкина, Худайбердыев, 1971), X. hopeiense из нижнемеловых отложений Восточной Сибири (Шилкина, Худайбердыев, 1971) и РДВ (Афонин, 2008), X. jakutiense из нижнемеловых отложений Восточной Сибири (Шилкина, 1986), X. jurassicum (Eckhold) Kräusel из юрских отложений Западной Сибири (Нащокин, 1968), X. latiporosum из юрских и меловых отложений Сибири (Нащокин, 1962, 1968; Дорофеюк, Филин, 1969; Шилкина, Худайбердыев, 1971), а также из меловых отложений РДВ (Афонин, 2008), X. meisteri из верхнемеловых (?) отложений Восточной Сибири (Палибин, Ярмоленко, 1932), X. phyllocladoides из нижнемеловых отложений РДВ (Afonin, Philippe, 2014), X. tomiense из юрских (?) отложений Западной Сибири (Ларищев, 1957) и X. watarianum из верхнемеловых отложений РДВ (Nishida, Nishida, 1986).
Образцы ископаемой древесины X. barberi (№№ 6г, 10г, 278, 1008, 1024, 22е), описанные Шилкиной и Худайбердыевым (1971) из юрских и нижнемеловых отложений Восточной Сибири, характеризуются наличием округлых, свободно или сближено расположенных, изредка уплощенных, сомкнутых пор на радиальных стенках трахеид, редких пор на тангентальных стенках трахеид, простых или слегка окаймленных пор оконцевого типа на полях перекреста, которые в два (три) раза больше по ширине, чем по высоте, а также отсутствием тяжевой паренхимы и крассул между порами на радиальных стенках трахеид. Такой набор признаков характерен для вида X. phyllocladoides из группы Phyllocladoides (Philippe et al., 2013), к которому и следует отнести эти образцы (табл. 3). Необходимо отметить, что вид X. barberi в настоящее время упразднен и рассматривается как синоним вида X. huttonianum (Philippe et al., 2013).
Таблица 3.
Результаты переопределения ископаемых древесин Xenoxylon из юрских и меловых отложений Сибири (коллекции И.А. Шилкиной и В.Д. Нащокина)
| Название таксона | Местонахождение | Литературный источник | Определение в настоящей работе | Примечание |
|---|---|---|---|---|
| Xenoxylon barberi (Seward) Kräusel | Восточная Сибирь: р. Лена, Хр. Орулган, нижняя юра (образец № 6г); р. Анабар, нижняя юра (образец № 10г); о. Большой Бегичев, средняя юра (образец № 22e); р. Зырянка, нижний мел (образцы №№ 278, 1008, 1024) | Шилкина, Худайбердыев, 1971 | Образцы №№ 6г, 10г, 278, 1008, 1024 – отнесены к Xenoxylon phyllocladoides; образец № 22e отнесен к Xenoxylon cf. phyllocladoides, очень низкие лучи (1–4 клетки), возможно древесина корня или ветки | Образцы и шлифы хранятся в БИН РАН, г. Санкт-Петербург |
| Xenoxylon hopeiense Chang | Восточная Сибирь, р. Оленек, нижний мел (образец № 87/7) | Шилкина, Худайбердыев, 1971 | Xenoxylon hopeiense | Образцы и шлифы хранятся в БИН РАН, г. Санкт-Петербург |
| Xenoxylon jurassicum (Eckhold) Kräusel | Западная Сибирь, р. Бахта, юра (образец № 116) | Нащокин, 1968 | Плохая сохранность, не имеет сходства с Xenoxylon (на основании описания и микрофотографий в работе Нащокина (1968) | Образец и шлифы № 116 потеряны; ИЛ СО РАН, г. Красноярск |
| Xenoxylon latiporosum (Cramer) Gothan | Восточная Сибирь: р. Бэйнчимэ, нижняя юра (образец № 3020б); бассейн р. Лена, юра (образцы №№ 615, 1032); р. Попигай, юра (образцы №№ 469, 216a); р. Нуолон, средняя юра (образец № 546ф); р. Анабар, средняя юра (образец № 17у); Хараулахский хребет, юра-нижний мел (образцы №№ 1179, 1181); р. Оленек, нижний мел (образец № 142з); Хребет Орулган, нижний мел (образец № 182б); р. Зырянка, нижний мел (образцы №№ 1031, 364) | Шилкина, Худайбердыев, 1971 | Образцы №№ 3020б, 615, 1032, 469, 216a, 546ф, 17у, 1179, 1181, 182б, 1031, 364 – отнесены к Xenoxylon meisteri; образец № 142з отнесен к Xenoxylon latiporosum | Образцы и шлифы хранятся в БИН РАН, г. Санкт-Петербург |
| Западная Сибирь: р. Енисей, юра (образец № 271); р. Бахта, юра (образец № 117); р. Кас, нижний мел (образцы №№ 8, 9, 10, 102) | Нащокин, 1962, 1968 | Образцы №№ 10, 102 отнесены к Xenoxylon meisteri | Образцы и шлифы №№ 10, 102 хранятся в ИЛ РАН, г. Красноярск; образцы и шлифы №№ 8, 9, 117, 271 потеряны |
Отнесение ископаемой древесины X. jurassicum (обр. № 116) из юрских отложений Западной Сибири (Нащокин, 1968) к Xenoxylon представляется нам весьма сомнительным. Исходя из приведенного Нащокиным (1968) очень краткого описания и микрофотографий, эта ископаемая древесина очень плохой сохранности, многие анатомические структуры смяты и характерные диагностические признаки Xenoxylon не видны. К сожалению, образец № 116 и его микропрепараты из коллекции Нащокина потеряны.
Большую часть образцов ископаемой древесины X. latiporosum (№№ 3020б, 615, 1032, 469, 216a, 546ф, 17у, 1179, 1181, 182б, 1031, 364 и др.) из юрских и меловых отложений Сибири, описанных в работах Нащокина (1968), а также Шилкиной и Худайбердыева (1971), мы относим к X. meisteri из группы Meisteri (табл. 3). Исследованные образцы часто имеют однорядные уплощенные поры на радиальных стенках трахеид, расположенные в группах, однорядные поры на тангентальных стенках трахеид и не высокие однорядные лучи, что характерно для X. meisteri. Ископаемые древесины Xenoxylon из группы Latiporosum, в том числе X. latiporosum, не имеют пор на радиальных стенах трахеид, распложенных в группах (Philippe et al., 2013). Образец № 32A/5 из нижнемеловых отложений РДВ, который был описан автором (Афонин, 2008) как X. latiporosum, также отнесен к виду X. meisteri.
По данным Филиппа с соавт. (Philippe et al., 2013), ископаемая древесина X. tomiense, описанная из юрских (?) отложений Западной Сибири (Ларищев, 1957), скорее всего, не имеет сходства с Xenoxylon; типовой материал это вида потерян.
Из местонахождений мелового возраста Сибири и РДВ нами описаны ископаемые древесины X. latiporosum, X. phyllocladoides, X. japonicum, X. meisteri и X. nariwaense (табл. 4). При этом X. japonicum и X. nariwaense указываются впервые для России, а X. meisteri – впервые для РДВ. Кроме того, X. japonicum и X. nariwaense впервые обнаружены в меловых отложениях, ранее эти виды были описаны только из верхнетриасовых отложений.
Таблица 4.
Ревизованный список ископаемых древесин Xenoxylon из юрских и меловых отложений Сибири и российского Дальнего Востока (с учетом новых и литературных данных)
| Название таксона | Местонахождение | Литературный источник | Название таксона, принятое в настоящей работе |
|---|---|---|---|
| Xenoxylon barberi | Восточная Сибирь, юра, нижний мел | Шилкина, Худайбердыев, 1971 | X. phyllocladoides |
| Xenoxylon hopeiense | Восточная Сибирь, нижний мел | Шилкина, Худайбердыев, 1971 | X. hopeiense |
| Российский Дальний Восток, нижний мел | Афонин, 2008 | X. hopeiense | |
| Xenoxylon jakutiense | Восточная Сибирь, нижний мел | Шилкина, 1986 | X. jakutiense |
| Xenoxylon japonicum | Восточная Сибирь, российский Дальний Восток, нижний мел | новые данные | X. japonicum |
| Xenoxylon jurassicum | Западная Сибирь, юра | Нащокин, 1968 | не имеет сходства с Xenoxylon |
| Xenoxylon latiporosum | Восточная Сибирь, мел (?) | Дорофеюк, Филин, 1969 | X. meisteri (Philippe et al., 2013) |
| Восточная Сибирь, юра, нижний мел | Шилкина, Худайбердыев, 1971 | X. meisteri, X. latiporosum | |
| Западная Сибирь, юра, нижний мел | Нащокин, 1962, 1968 | X. meisteri | |
| Российский Дальний Восток, нижний мел | Афонин, 2008 | X. meisteri | |
| Российский Дальний Восток, нижний мел | новые данные | X. latiporosum | |
| Xenoxylon meisteri | Восточная Сибирь, верхний мел (?) | Палибин, Ярмоленко, 1932 | X. meisteri |
| Российский Дальний Восток, нижний мел | новые данные | X. meisteri | |
| Xenoxylon nariwaense | Российский Дальний Восток, нижний мел | новые данные | X. nariwaense |
| Xenoxylon phyllocladoides | Российский Дальний Восток, нижний мел | Afonin, Philippe, 2014 | X. phyllocladoides |
| Российский Дальний Восток, нижний мел | новые данные | X. phyllocladoides | |
| Xenoxylon tomiense | Западная Сибирь, юра (?) | Ларищев, 1957 | не имеет сходства с Xenoxylon (Philippe et al., 2013) |
| Xenoxylon watarianum | Российский Дальний Восток, верхний мел | Nishida, Nishida, 1986 | X. watarianum |
Таким образом, с учетом новых и литературных данных, а также результатов ревизии древесных остатков Xenoxylon из коллекций Шилкиной и Нащокина, по анатомическим признакам ископаемой древесины из юрских и меловых отложений Сибири и РДВ установлено восемь представителей морфорода Xenoxylon: X. latiporosum, X. watarianum, X. hopeiense, X. jakutiense, X. phyllocladoides, X. japonicum, X. meisteri и X. nariwaense (табл. 4). Проведенные нами исследования показывают, что представители трех основных анатомических групп Xenoxylon (Latiporosum, Phyllocladoides, Meisteri) встречались в течение юрского и мелового периодов на территории современной Северной Азии. Полученные результаты подтверждают выводы Филиппа с соавт. (Philippe et al., 2009), о том, что в мезозое в этом регионе, по-видимому, был основной центр видообразования Xenoxylon.
Список литературы
Афонин М.А. Первые находки ископаемых древесин Xenoxylon latiporosum (Cramer) Cothan и X. hopeiense Chang на Дальнем Востоке России // Вестн. ДВО РАН. 2008. № 4. С. 126–132.
Волынец Е.Б. Апт-сеноманская флора Приморья. Статья 2. Корреляция флористических комплексов // Стратигр. Геол. корреляция. 2006. Т. 14. № 2. С. 105–116.
Гаммерман А.Ф., Никитин А.А., Николаева Л.Т. Определитель древесин по микроскопическим признакам. М.; Л.: Изд-во АН СССР, 1946. 143 с.
Дорофеюк Н.И., Филин В.Р. О Xenoxylon latiporosum (Cramer) Gothan из верхнемеловых отложений Забайкалья // Бюл. МОИП. Отд. биол. 1969. Т. 74. № 4. С. 72–87.
Жаров А.Э., Кириллова Г.Л., Маргулис Л.С. и др. Геология, геодинамика и перспективы нефтегазоносности осадочных бассейнов Татарского пролива. Владивосток: Дальнаука, 2004. 220 с.
Красилов В.А. Раннемеловая флора Южного Приморья и ее значение для стратиграфии. М.: Наука, 1967. 364 с.
Ларищев А.А. О некоторых ископаемых остатках древесины из Западной Сибири // Тез. докл. VII науч. конф. Томского гос. ун-та. Томск, 1957. С. 13–15.
Маркевич В.С. Меловая палинофлора севера Восточной Азии. Владивосток: Дальнаука, 1995. 200 с.
Нащокин В.Д. Ископаемые древесины из меловых отложений // Биостратиграфия мезозойских и третичных отложений Западной Сибири. Л.: Гостоптехиздат, 1962. С. 282–294 (Тр. СНИИГГиМС. Вып. 22).
Нащокин В.Д. Ископаемые древесины из меловых, третичных и четвертичных отложений Средней Сибири. М.: Наука, 1968. 175 с.
Палибин И.В., Ярмоленко А.В. Новый вид ископаемой древесины с Витимского плоскогорья // Изв. Всес. геол.-разв. объед. 1932. Т. 51. Ч. 8. С. 177–179.
Скобло В.М., Лямина Н.А., Руднев А.Ф., Лузина И.В. Континентальный верхний мезозой Прибайкалья и Забайкалья (стратиграфия, условия осадконакопления, корреляция). Новосибирск: Изд-во СО РАН, 2001. 332 с.
Худайбердыев Р., Гомолицкий Н.П., Лобанова А.В. Материалы к юрской флоре Южной Ферганы // Палеоботаника Узбекистана. Т. II. Ташкент: Изд-во ФАН Узб. ССР, 1971. С. 3–57.
Шилкина И.А. Ископаемые древесины Северной Евразии // Проблемы палеоботаники. Л.: Наука, 1986. С. 150–157.
Шилкина И.А., Худайбердыев Р. Новые находки и обзор родов Protocedroxylon и Xenoxylon // Палеоботаника Узбекистана. Т. II. Ташкент: Изд-во ФАН Узб. ССР, 1971. С. 117–134.
Ярмоленко А.В. Опыт применения анатомии вторичной древесины ствола к объяснению филогении хвойных // Сов. ботаника. 1933. № 6. С. 46–63.
Яценко-Хмелевский А.А. Основы и методы анатомического исследования древесины. М.-Л.: Изд-во АН СССР, 1954. 337 с.
Afonin M., Philippe M. Fossil woods from the Lower Cretaceous (Albian) of Kamchatka Peninsula, Russian Far East // Cret. Res. 2014. V. 50. P. 110–119.
Bailey I.W. Evolution of the tracheary tissue of land plants // Amer. J. Bot. 1953. V. 40. № 1. P. 4–8.
Cramer C. Fossile Hölzer der Arktischen Zone // von Heer. O. (Ed.) Flora fossilis arctica – Die fossile Flora der Polarländer. Zürich, 1868. P. 167–180.
Gothan W. Zur Anatomie lebender und fossiler Gymnospermen Hölzer // Abh. Preuss. Geol. Land. 1905. Bd 44. S. 1–108.
Gothan W. Fossile Hölzer aus dem Bathonien von Russisch-Polen // Verh. Russ.-Kais. Mineral. Ges. 1906. Bd 44. S. 435–458.
IAWA List of Microscopic Features for Softwood Identification // IAWA J. 2004. V. 25. № 1. P. 1–70.
Marynowski L., Philippe M., Zaton M., Hautevelle Y. Systematic relationships of Mesozoic wood genus Xenoxylon: integrative biomolecular and palaeobotanical approach // N. Jb. Geol. Paläontol. Abh. 2008. V. 247. P. 177–189.
Muller-Stoll W.R. Evolutionary trends in gymnospermous wood structures during the Mesozoic. Protopinaceous woods in the German Jurassic // Palaeobotanist. 1987. V. 35. P. 233–235.
Nishida M., Nishida H. Petrified plants from the Upper Cretaceous of Saghalien (1) // Bot. Mag. Tokyo. 1986. V. 99. P. 191–204.
Philippe M., Bamford M.K. A key to morphogenera used for Mesozoic conifer-like woods // Rev. Palaeobot. Palynol. 2008. V. 148. P. 184–207.
Philippe M., Jiang H.-E., Kim K., Oh C. et al. Structure and diversity of the Mesozoic wood genus Xenoxylon in Far East Asia: implications for terrestrial palaeoclimates // Lethaia. 2009. V. 42. № 4. P. 393–406.
Philippe M., Thévenard F., Nosova N. et al. Systematics of a palaeoecologically significant boreal Mesozoic fossil wood genus, Xenoxylon Gothan // Rev. Palaeobot. Palynol. 2013. V. 193. P. 128–140.
Shimakura M. Studies on fossil woods from Japan and adjacent lands (1) // Sci. Rep. Tohoku Imp. Univ. Sendai. 1936. Ser. 2 (Geol.). V. 18. P. 267–298.
Suzuki M., Terada K. Xenoxylon fossil woods from the Lower Cretaceous Akaiwa Subgroup of Shiramine, Central Japan // J. Phytogeo. Taxon. 1992. V. 40. P. 91–97.
Tian N., Wang Y., Philippe M. et al. New record of fossil wood Xenoxylon from the Late Triassic in the Sichuan Basin, southern China and its paleoclimatic implications // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2016. V. 464. P. 65–75.
Vogellehner D. Zur Anatomie und Phylogenie mesozoischer Gymnospermenhölzer, Beitrag 7: Prodromus zu einer Monographie der Protopinaceae. I. Dieprotopinoiden Hölzer des Jura // Palaeontogr. Abt. B. 1968. Bd 124. S. 125–162.
Wan M., Zhou W., Tang P. et al. Xenoxylon junggarensis sp. nov., a new gymnospermous fossil wood from the Norian (Triassic) Huangshanjie Formation in northwestern China, and its palaeoclimatic implications // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2016. V. 441. P. 679–687.
Yamazaki S., Tsunada K. Some fossil woods from the Upper Triassic Nariwa and Mine Groups, the Inner Zone of Southwest Japan // J. Geol. Soc. Japan. 1982. V. 88. № 7. P. 595–611.
Yamazaki S., Tsunada K., Koike N. Some fossil woods from the Upper Triassic Nariwa Group, Southwest Japan // Mem. School Sci. Engin., Waseda Univ. 1980. V. 44. P. 91–131.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал