

Палеонтологический журнал, 2019, № 1, стр. 73-86
Ящерицы семейств Eoxantidae, Ardeosauridae, Globauridae и Paramacellodidae (Scincomorpha) из апта–альба Монголии
В. Р. Алифанов *
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: valifan@paleo.ru
Поступила в редакцию 17.10.2017
Принята к публикации 12.04.2018
Аннотация
В работе обсуждена проблема классификации ящериц инфраотряда Scincomorpha. Более подробно в ней освещены история и проблемы изучения таких его ископаемых семейств, как Eoxantidae, Ardeosauridae, Globauridae и Paramacellodidae. Состав названных групп пополнен новыми видами из апт-альбcких отложений Центральной Монголии.
ВВЕДЕНИЕ
Отложения местонахождения раннемеловых наземных тетрапод Хобур (Ховур, Гучин-Ус) выходят на поверхность в пределах обширного поля песчано-глинистых пород озерного и озерно-речного генезиса в Центральной Монголии (Увэрхангайский аймак). Они принимаются как часть хухтыкского горизонта без формального отнесения к той или иной выделенной в Монголии свите и датируются либо аптом и альбом включительно (Shuvalov, 2000), либо поздним аптом и всем альбом (Lucas, 2006). По нашим предварительным данным, хухтыкский горизонт соответствует интервалу времени, в который входят поздний апт и ранний альб.
Первые направленные сборы в Хобуре проводила в течение нескольких полевых сезонов (начиная с 1969 г.) Совместная Советско-Монгольская палеонтологическая экспедиция. Успехом стало открытие редких на тот момент остатков раннемеловых млекопитающих. Однако основу разнообразия достаточно богатого комплекса наземных тетрапод местонахождения формируют пресмыкающиеся: черепахи, крокодилы, птерозавры и динозавры. Последние три группы в сборах представлены, главным образом, зубами. Наиболее разнообразный компонент фауны наземных тетрапод Хобура – ящерицы. В коллекции их остатки представлены обычно изолированными элементами скелета, среди них преобладают челюстные кости с зубами.
Ящерицам Хобура посвящено несколько относительно недавно опубликованных работ (Daza et al., 2012; Алифанов, 2016), включающих результаты переизучения древнейшего представителя семейства Gekkonidae s.l. – Hoburogekko suchanovi Alifanov, 1989, а также данные (в том числе, описание новых видов) по семейству Hodzhakuliidae (Scincomorpha). Наши предварительные определения указывают на наличие в Хобуре девяти семейств ящериц. Часть из них – это эндемики раннемеловой Центральной Азии (Eoxantidae, Hodzhakuliidae, Xenosauridae, Gekkonidae s.l., Temujiniidae), а часть (Paramacellodidae, Ardeosauridae, Globauridae, Dorsetisauridae) представляют исторический этап, во время которого Центральная Азия входила в единый блок северных континентов.
Настоящая публикация посвящена самой разнообразной в Хобуре группе ящериц – инфраотряду Scincomorpha. В ней поднимаются вопросы классификации этого таксона в целом, а также проблемы, связанные с изучением входящих в него ископаемых семейств, в частности, Eoxantidae, Ardeosauridae, Globauridae и Paramacellodidae. В составе этих групп описаны новые виды.
Работа поддержана грантом РФФИ, № 16-05-00408.
ОБЩИЕ И ЧАСТНЫЕ ПРОБЛЕМЫ КЛАССИФИКАЦИИ ЯЩЕРИЦ ИНФРАОТРЯДА SCINCOMORPHA
Общая проблематика. Текущую классификацию ящериц отражают многочисленные варианты кладистического переизложения системы Ч. Кэмпа (Camp, 1923). И первые из них (напр., Estes et al., 1988), и последние (напр., Conrad, 2008; Gauthier et al., 2012) представляют обсуждаемую группу членами таксона Squamata, “продвинутая” часть которого дает начало амфисбенам и змеям. Этот взгляд, подразумевающий парафилетичность ящериц, построен на формальном подходе к проблеме параллелизмов (однонаправленным изменениям признаков или их комплексов у родственных форм), которые для группы исключительно распространены. С нашей точки зрения, проблема параллелизмов имеет ключевое значение, поскольку без ее специального рассмотрения стабильную и отвечающую реальному разнообразию классификацию создать невозможно. При этом внимания требуют не столько простые и мозаически проявляемые преобразования, сколько их более сложные формы, например, параллелизмы комплексного типа. Некоторые из них определяют появление похожих морфологически форм. Очевидные примеры последнего рода демонстрируют такие семейства, как Anniellidae (Anguimorpha), Dibamidae, Acontiidae (Scincomorpha), Pygopodidae (Gekkota). Для их представителей характерен роющий образ жизни и связанные с ним признаки: утрата конечностей и сходные преобразования черепа (удлинение затылочно-теменной области, редукция мембранной части мозговой коробки, укорочение ринального отдела, приобретение массивности слуховых костей, сокращение числа челюстных зубов, и др.).
Сравнение перечисленных выше групп указывает на значимые для классификации и реконструкции филогенеза элементы строения. К их числу относятся, прежде всего, признаки небного комплекса, в котором удается выявить два распространенных типа: платинотопалатинарный (латеральные отростки небных и наружных крыловидных формируют контакт медиальнее верхнечелюстных костей, которые исключены из обрамления суборбитальных окон) и диплоглоссопалатинарный (латеральные отростки небных и наружных крыловидных костей расположены дорсальнее супрадентального гребня верхнечелюстных костей, которые входят в обрамление суборбитальных окон). Оба состояния представлены во всех крупных стволах ящериц и не могут быть сведены друг к другу прямо из-за разного их положения в разветвленном и полистадийном семогенетическом (трансформационном) ряду (Алифанов, 2000, 2012, 2016). На примере строения небного комплекса можно говорить о том, что у ящериц имеют место примеры регулярно проявляемых параллелизмов.
Регулярные параллелизмы, по нашим представлениям, – результат интенсивной радиации при ограничениях по основной морфо-физиологической адаптации. Для ящериц таковой является кинетизм черепа, за которым стоит возможность изменения общей конфигурации дерматокрания относительно нейрокрания с фокусированием подвижности в крыше черепа, где формируются мезо- и метакинетическая оси (амфикинетизм). При работе челюстной аппарат ящериц в целом приобретает возможность к манипулированию небольшой в сравнении с собственными размерами добычей как пинцетом или большим и указательным пальцами человеческой руки (Иорданский, 1966, 1990), в результате чего снижается риск ее потери в ущерб возможностям силовой обработки.
Амфикинетизм – уникальная и сложная морфо-физиологическая адаптация, которая не может быть автоматически сведена к формам кинетизма черепа других сквамат (к акинетизму амфисбен или гиперкинетизму змей), каждая из которых сопровождается особым типом строения конструкционных узлов черепа и композицией челюстных мускулов. В свете этих данных квалификация ящериц в качестве предков амфисбен и змей выглядит сомнительно, поскольку исключает из обсуждения вопросы о трансформировании амфикинетизма в другие формы или о независимом формировании всех форм кинетизма сквамат. Филогенетическое следствие рассмотрения этих проблем может быть выражено в гипотезе происхождения всех групп сквамат от общего и неспециализированного по типу кинетизма предка.
Учесть указанные выше обстоятельства и данные мы попытались при построении новой классификации ящериц с помощью приемов и методов эволюционной таксономии. Она была опубликована нами ранее (Алифанов, 2000, 2012). В настоящей статье приведен несколько модифицированный по сравнению с последним новый вариант (рис. 1, а). По нему, как и раньше, предполагается монофилия ящериц и их первоначальная дивергенция на два ствола (подотряды): Scincogekkonomorpha и Iguania. Подотряды характеризуются специфическим строением височных дуг: у первых развит преимущественно вентральный, а у вторых – медиальный отросток окципитального конца чешуйчатых костей). Каждый ствол распадается на три ветви (инфраотряды): Anguimorpha, Scincomorpha, Lacertognatha (Scincogekkonomorpha), Iguanota, Pachyglossa, Agamognatha (Iguania). Для каждого из инфраотрядов сцинкогеккономорф удается установить особый тип строения нижних челюстей (ангвиморфный, лацерто-геккотовый, сцинкоморфный). Для игуаний также отмечается три типа: два для Iguanota (мозазавро-тейюморфный и игуаноморфный) и один для Pachyglossa и Agamognatha (агамо-хамелеоморфный). Типы определены по комбинации степени развития и положения отростков заднего края зубных костей, а иногда – с учетом степени развития и положения лабиального отростка венечных костей. Данные признаки связаны с укреплением элементов ветвей нижней челюсти друг с другом на фоне повышения на челюстной аппарат нагрузок, видимо, обусловленных развитием челюстной мускулатуры при расширении предпочтений в выборе корма (Алифанов, 2000).
Рис. 1.
Распределение двух основных морфотипов небного комплекса у ящериц и использование феномена периодичности для реконструкции филогенеза: а – макротаксонов (по: Алифанов, 2012, с изменениями), б – семейств инфраотряда Scincomorpha (по: Алифанов, 2016). Обозначения: P – платинотопалатинарность, D – диплоглоссопалатинарность.
Типичные сцинкоморфы характеризуются наличием короноидного отростка зубных костей, налеганием последнего на лабиальную поверхность дорсального и лабиального (если выражен) отростков венечной кости, редукцией (обычно значительной или полной) супраангулярного отростка зубных костей, окципитальным разрастанием ангулярного отростка зубных костей до пересечении им уровня переднего супраангулярного отверстия и формированием обширной вырезки между короноидным и окципитальным отростками.
Типология небного комплекса и нижней челюсти позволяет ревизовать состав большинства инфраотрядов, включая Scincomorpha. Последний традиционно содержит наибольшее число семейств, сомнительных с точки зрения их родства с типичными представителями этой группы. К первой категории следует отнести современных Gymnophthalmidae, Lacertidae, Feyliniidae (Lacertia), Teiidae (Teiumorpha) и ископаемых (позднемеловых) Polyglyphanodontidae, Adamisauridae (Polyglyphanodontia), Gilmoreteiidae, Cherminsauridae, Mongolochamopidae и Pyramicephalosauridae (Macrocephalosauria). Филогенетические связи всех перечисленных семейств нам удается определить за рамками несомненных сцинкоморф. Например, Lacertidae и Gymnophthalmidae (Lacertia) могут быть представлены в общем таксоне (Lacertognatha) с диплоглоссопалатинарными Gekkota, а макротейиды (Teiumorpha) и макротейидообразные ископаемые Polyglyphanodontia и Macrocephalosauria возможно представить обладающими платинотопалатинарностью микроотрядами в составе подотряда Iguania в рамках инфраотрядов Iguanota, Agamognatha и Chamaeleomorpha, соответственно (рис. 1, а).
Судя по сцинкоморфному типу строения нижней челюсти, к истинным сцинкоморфам можно уверенно отнести ряд современных (Dibamidae, Xantusiidae, Cordylidae, Acontiidae и Scincidae) и ископаемых (Ardeosauridae, Carusiidae, Eoxantidae, Globauridae, Paramacellodidae, Slavoiidae и Hodzhakuliidae) семейств. Типология неба, о которой говорилось выше, позволяет в рамках собственно сцинкоморф выявить две группы семейств (рис. 1, б): платинотопалатинарных Xantusiidae, Dibamidae, Slavoiidae, Eoxantidae и, видимо, Hodzhakuliidae (микроотряд Xantusiomorpha), а также диплоглоссопалатинарных Acontiidae, Scincidae, Cordylidae, Ardeosauridae, Carusiidae, Globauridae, Paramacellodidae (Leptoglossa).
Представленные выше общие данные дают возможность далее перейти к критическому рассмотрению проблем изучения четырех ископаемых семейств. В составе именно этих групп, как уже отмечалось, описаны новые виды.
Семейство Eoxantidae. Единственный род семейства – Eoxanta Borsuk-Białynicka, 1988 (рис. 2, а–в) – известен из позднего мела Монголии. М. Борсук-Бялыницка (Borsuk-Białynicka, 1988) небезосновательно предположила его связь с современными североамериканскими Xantusiidae. Последняя группа краниологически наиболее архаична в составе ксантузиоморф, судя по отсутствию у ее представителей верхних височных, а иногда и суборбитальных окон (палеоплатинотопалатинарность) в строении небного комплекса. Для ксантузиид характерна также постплатинотопалатинарность (утрата связи латеральных отростков небных и наружных крыловидных костей, расположенных медиальнее верхнечелюстных костей). Кроме того, последняя группа обладает продвинутым (тубулярным) типом строения зубных костей. Этот комплекс признаков отмечается и у Gekkota. Однако ключевое отличие ксантузиид, как и всех сцинкоморф, состоит в особом (сцинкоморфном) типе строения их нижней челюсти. К Xantusiidae наиболее близок Tepexisaurus (Scincomorpha fam. indet.: Reynoso, Callison, 2000) из раннего мела Мексики. Данный род позволяет говорить о североамериканском происхождении обсуждаемого современного семейства.
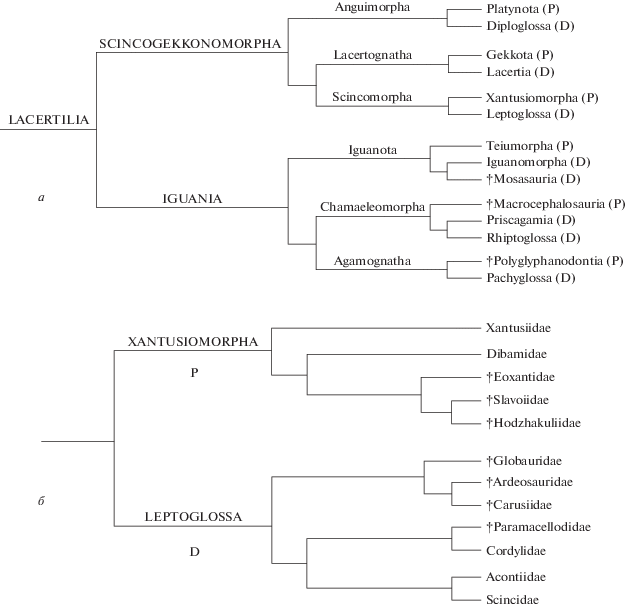
Рис. 2.
Представители семейства Eoxantidae: а–в – Eoxanta lacertifrons Borsuk-Białynicka, 1988 из позднего мела Монголии; реконструкция черепа дорсально (а), нижней челюсти лабиально (б) и лингвально (в); г – Palaeoxanta conicodentatus gen. et sp. nov. (Eoxantidae, Scincomorpha), голотип ПИН, № 3334/520, левая зубная кость лингвально; Монголия, Увэрхангайский аймак, местонахождение Хобур; нижний мел, верхний апт–нижний альб, хухтыкский горизонт.
Нами для классификации Eoxanta Borsuk-Białynicka,1988 было предложено семейство Eoxantidae (Алифанов, 2000), до выделения которого эта форма рассматривалась в семействах Xantusiidae (Алифанов, 1993а, б) и Slavoiidae (Алифанов, 2000). Ее сходство со Slavoia Sulimski, 1984 из позднего мела Монголии, ускользнувшее от внимания других исследователей, можно подчеркнуть платинотопалатинарностью и другими признаками. Например, в отличие от Xantusiidae, Eoxanta и Slavoia характеризуются сходной комбинацией продвинутых и архаических особенностей орбитального региона и височных дуг. К этой же ветви центральноазиатских по происхождению сцинкоморф принадлежат и раннемеловые Hodzhakuliidae (Алифанов, 2016).
Согласно другим точкам зрения, для Eoxanta отмечались невозможность определения точнее, чем Scincomorpha incertae sedis (Gao, Norell, 2000), связь с Globauridae (Gauthier et al., 2012), а также с Myrmecodaptria Gao et Norell, 2000 (Ardeosauridae; см. ниже) из позднего мела Монголии в рамках семейства Bainguidae (Conrad, 2008; по нашим данным, это архаичное подсемейство семейства Anguidae).
Семейство Ardeosauridae. Данная группа имела широкое распространение в прошлом. Она стала известной по находкам близкородственных (Estes, 1983) Ardeosaurus Meyer, 1860 и Eichstaettisaurus Kuhn, 1958 (рис. 3, а, б) из поздней юры Германии. К семейству мы относим всех ящериц из серии Джехол (нижний мел) в Северо-Восточном Китае. К ним же, с нашей точки зрения, принадлежит неназванная ящерица (Lacertilia indet.: Аверьянов, Федоров, 2004) из ходжиабатской свиты (неоком) в Киргизии. К числу ардеозаврид предварительно могут быть причислены и известные по неполным остаткам формы из нижнемеловых отложений Монголии (Norellius Conrad et Daza, 2015) и Японии (Sakurasaurus Evans et Manabe, 1999). Семейство нами устанавливается также в позднем мелу Монголии судя по Myrmecodaptria Gao et Norell, 2000 (рис. 3, в). Это, видимо, позднейший для Центральной Азии и всего мира представитель ардеозаврид.
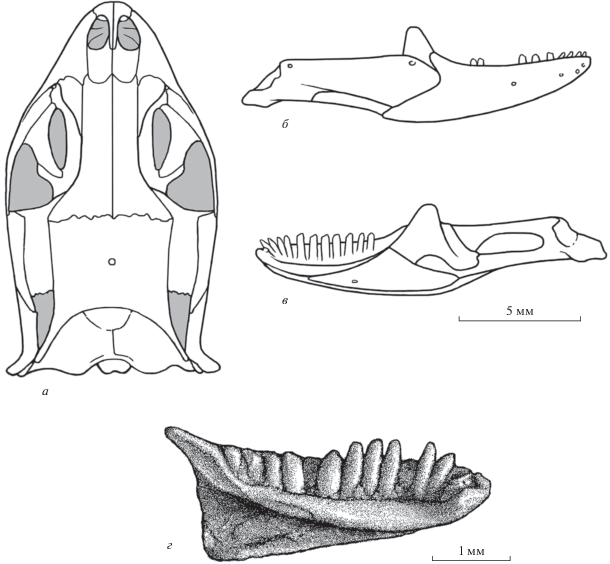
Рис. 3.
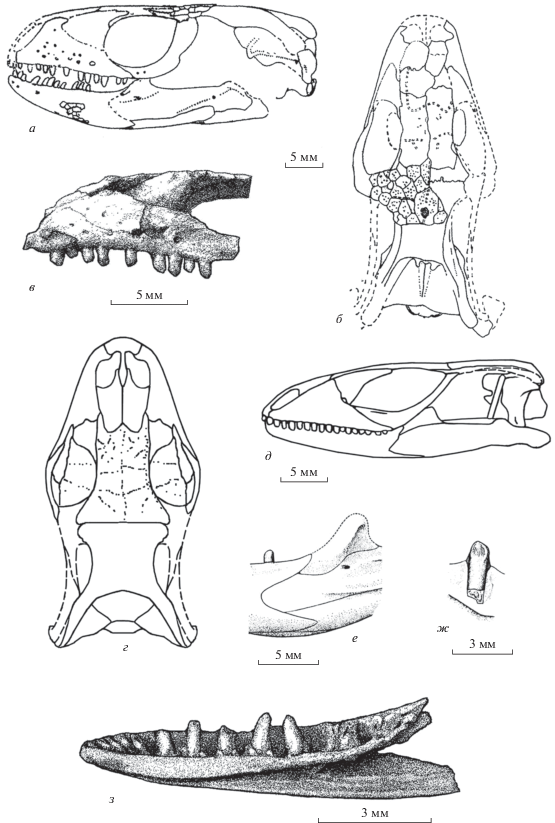
Представители семейства Ardeosauridae: а – Eichstaettisaurus schroederi Broili, 1938, череп дорсально (по: Estes, 1983, фиг. 12, B); поздняя юра Германии; б – Ardeosaurus brevipes Meyer, 1860, фрагмент задней части крыши черепа и височная дуга дорсально (по: Mateer, 1982, текст-фиг. 2, часть); поздняя юра, Германия; в – Myrmecodaptria microphagosa Gao et Norell, 2000, череп дорсально (по: Gao, Norell, 2000, фиг. 14, B); поздний мел Монголии; г, д – Dalinghosaurus longidigitus Ji, 1998, череп дорсально (г) и центральная часть левой ветви нижней челюсти лабиально (д) (по: Evans, Wang, 2005, фиг. 2, А; Evans et al., 2007, текст-фиг. 3, Е); ранний мел Китая; е–и – Gurvelus khangaicus gen. et sp. nov., голотип ПИН, № 3334/518; нижняя челюсть лабиально (е), лингвально (ж) и реконструкция нижнечелюстного зуба лабиально (з) и рострально (и); местонахождение Хобур, Увэрхангайский аймак, Монголия; нижний мел, верхний апт–нижний альб, хухтыкский горизонт. Обозначения костей: J – скуловая, F – лобная, P– теменная, Pf – заднелобная, Po – заглазничная, Sq – чешуйчатая.
Ardeosauridae оказались сложны для определения их филогенетических связей из-за недостаточной сохранности первых находок. Противоречивость мнений в отношении этой группы имела место у ранних авторов и присуща современным специалистам. Однако все определения основаны либо на отсутствии данных по строению деталей нижней челюсти, либо на недооценке их значения для построения гипотез родства. Чаще всего ардеозаврид сближают с Gekkota (иногда в составе кладистического таксона Gekkonomorpha: Camp, 1923; Estes, 1983; Gauthier et al., 2012; Conrad, Daza, 2015; Simões et al., 2017). Кроме этого, отдельные представители группы воспринимаются базальными сцинкогеккономорфами (Eichstaettisaurus: Conrad, 2008; Yabeinosaurus: Evans, Wang, 2012) и даже классифицируются в составе ангвиморф (Dalinghosaurus: Evans, Wang, 2005).
По представленному в работе Н. Матира (Mateer, 1982) черепу, для Ardeosaurus Meyer, 1860 устанавливается диплоглоссопалатинарность. Эта особенность противоречит классификации данного рода и всего семейства в составе или в родстве с Gekkota, для современных представителей которых характерна платинотопалатинарность (иногда переходит, как у Xantusiidae, в постплатинотопалатинарность). Ардеозавриды специфичны по строению височных дуг: заглазничные кости входят в состав обрамления глазниц и обычно несут отчетливо выраженный медиальный отросток, расположенный ростральнее заднелобных костей (рис. 3, а, б). Напомним, что у современных Gekkota заглазничные кости редуцированы, а их наличие даже у ископаемых представителей не установлено. Данные по строению нижней челюсти у Norellius Conrad et Daza, 2015 из раннего мела Монголии или Dalinghosaurus Ji, 1998 из раннего мела Китая (рис. 3, г, д) характеризуют семейство как типичных сцинкоморф. Представителям ардеозаврид свойственно также отсутствие свободных остеодерм. Остеодермальная скульптура на покровных костях крыши черепа может быть выражена (тип Eichstaettisaurus) или отсутствовать (тип Ardeosaurus).
Нами семейство Ardeosauridae рассматривается в составе диплоглоссопалатинарных сцинкоморф (микроотряд Leptoglossa). С ним проявляют сестринское сходство Carusiidae (включает роды Carusia Borsuk-Białynicka, 1987 и Shinisauroides Borsuk-Białynicka, 1985 из позднего мела Монголии). Последнее характеризуется срастанием заднелобных и заглазничных костей, узкими лобными костями, окципитальной экспансией ангулярного отростка зубных костей, многочисленностью тонких и высоких зубов, их притупленной верхушкой, гиперплевродонтным прикреплением, а также рядом других признаков. Иногда карусиид в составе одного рода – Carusia Borsuk-Białynicka, 1987 – ошибочно сближают с ангвиморфами семейства Xenosauridae s.l. (Gao, Norell, 1998, 2000; Conrad, 2008).
Семейство Globauridae. Это еще одно семейство надсемейства Ardeosauroidea, у представителей которого отсутствует остеодермальный панцирь и развит диплоглоссопалатинарный тип неба. Типовой род семейства – Globaura Borsuk-Białynicka, 1988; (рис. 4, а–в) – обычен для позднего мела Монголии. В первоописании (Borsuk-Białynicka, 1988) он рассматривался в таксоне Lacertoidea (sensu Estes et al., 1988), который, по нашему представлению, является сборной по составу группой (большинство его представителей попадает в состав упоминавшихся выше сомнительных сцинкоморф).
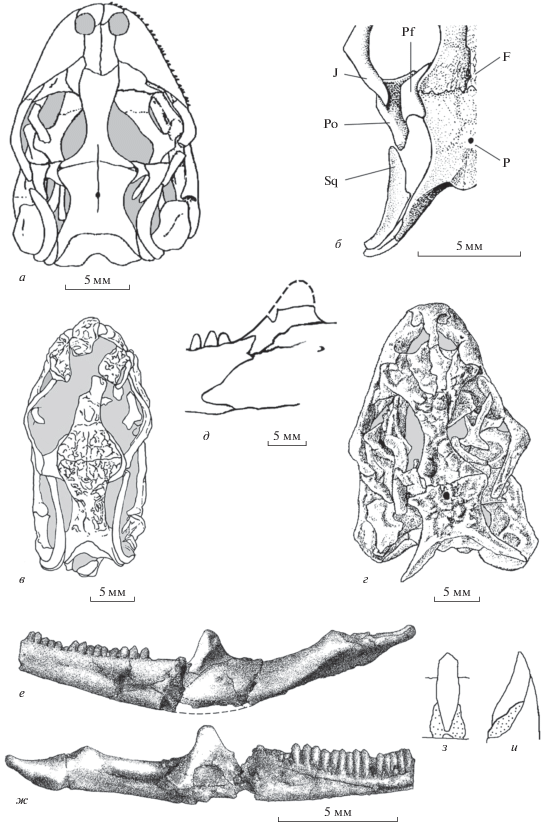
Рис. 4.
Представители семейства Globauridae: а–в – Globaura venusta Borsuk-Białynicka, 1988, реконструкция черепа дорсально (а), вентрально (б) и лабиально (в); поздний мел Монголии; г, д – Meyasaurus crusafonti (Hoffstetter, 1965), голотип IPPS, № Mont. 10, фрагмент черепа, правая ветвь нижней челюсти (г) и нижнечелюстной зуб (д) (по: Hoffstetter, 1966, фиг. 2, B); е – Meyasaurus diazromerali Evans et Barbadillo, 1997, экз. LH 13510B, передняя часть височной дуги (по: Evans, Barbadillo, 1997, фиг. 8, А); ранний мел Испании; ж – Durotrigia triconidens Hoffstetter, 1967, нижнечелюстные зубы (по: Hoffstetter, 1967, фиг. 8, С); титон–неоком Англии; з – Khereidia aptiana gen. et sp. nov., голотип ПИН, № 3334/523, правая зубная кость лингвально; Монголия, Увэрхангайский аймак, местонахождение Хобур; нижний мел, верхний апт–нижний альб, хухтыкский горизонт. Обозначения как на рис. 3.
Globauridae отличаются от Ardeosauridae и Carusiidae строением височных дуг, а именно: положением небольшого медиального отростка заглазничных костей в соответствующей ему вырезке на медиальном фланге укрупненных заднелобных костей. Такое состояние характерно для многих семейств ящериц в рамках тех или иных подгрупп (надсемейств, или микроотрядов) инфраотрядов. У глобаурид также нередко развиты двухвершинные зубы, как, например, у Meyasaurus (Ilerdaesaurus) Vidal, 1915 (рис. 4, г–е) из раннего мела Испании (Evans, Barbadillo, 1997). Род Tijubina Bonfim-Júnior еt Marques, 1997 (базальные склероглоссы: Simões, 2012), известный из нижнемеловой формации Крато в Бразилии, вероятно, также принадлежит этому же семейству, судя по наличию близ вершинки зубов, по крайней мере, нижнечелюстного ряда рострально ориентированного плеча (в первоописании этот признак не отмечается, но заметен на фотографии).
Современные обладатели двухвершинных зубов в составе, например, Lacertidae и Tupinambidae формируют представителей, несущих также несимметрично трехвершинные зубы. Наличие таких форм возможно и для глобаурид. Предположительно, к ним относится проблематичная Durotrigia Hoffstetter, 1967 (Lacertilia incertae sedis: Estes, 1983) из формации Пурбек (титон–неоком) Англии (рис. 4, ж). Не исключено, что к этому же семейству, а не к Iguanomorpha, как было предположено в первоописании, принадлежит еще одна проблематичная ящерица, Brasiliguana Nava et Martinelli, 2011, известная из позднего мела Южной Америки по левой верхнечелюстной кости. Она также обладает асимметричными по строению верхушками трехвершинных зубов. Такие зубы у каких-либо современных игуаноморф не отмечаются.
По данным кладистического анализа (Gauthier et al., 2012), Globauridae включают, кроме типового, роды Hymenosaurus Gao et Norell, 2000 (в первоописании предварительно рассматривался в составе Scincoidea; судя по небольшим глазницам и связи предлобных и заднелобных костей, может быть связан тесным родством со Slavoiidae) и Eoxanta Borsuk-Białynicka, 1988 (см. Eoxantidae). Также имеют место попытки классифицикации типового рода обсуждаемого семейства в составе Scincomorpha incertae sedis (Gao, Norell, 2000) или позднемеловых Bainguidae (Conrad, 2008). О последней группе выше говорилось, как о возможном надсемействе современного семейства Anguidae (Anguimorpha).
Cемейство Paramacellodidae. Это юрско-раннемеловая группа сцинкоморфных ящериц, характеризующаяся наличием у ее представителей прямоугольных остеодерм. Типовой род семейства – Paramacellodus Hoffstetter, 1967 – описан по челюстным костям, позвонкам и остеодермам прямоугольной формы из нижнего мела Англии. Фрагментарные находки парамацеллодид известны также из поздней юры Eвропы, Африки, Северной Америки, из раннего мела Монголии (Paramacellodidae gen. indet.: Alifanov, 1993, 2000; Алифанов, 2000, 2012) и России (Paramacellodus sp.: Averianov, Skutchas, 1999; Paramacellodidae gen. indet.: Алифанов, 2012). Прямоугольные остеодермы отмечаются у Сhangetisaurus Nessov, 1992 (рис. 5, а–в) из средней юры Кыргызстана (Федоров, Несов, 1992) и Sharovisaurus Hecht et Hecht, 1984 из поздней юры Казахстана (Хехт, Хехт, 1984). Две последние формы представлены относительно полными материалами: черепом в первом случае и скелетом (в обрамлении остеодерм) во втором. В Азии группа обнаружена также в поздней юре Китая (Mimobecklesisaurus Li, 1985). К семейству Paramacellodidae, с нашей точки зрения, следует причислить Parmeosaurus Gao et Norell, 2000 (рис. 5, г, д) из позднего мела Монголии. По строению черепа он близок к Сhangetisaurus Nessov, 1992. Остеодермы Parmeosaurus Gao et Norell, 2000 имеют разную форму их контура: прямоугольную на туловище и округлую в подбородочной области, как у Sharovisaurus Hecht et Hecht, 1984. По нашему мнению, род из позднего мела Монголии – наиболее поздний представитель Paramacellodidae.
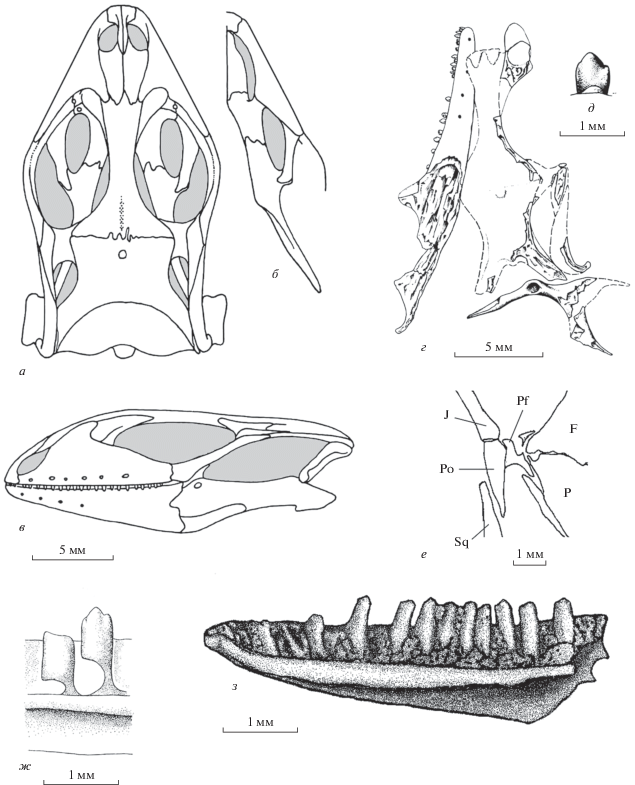
Рис. 5.
Представители семейства Paramacellodidae: а–в – Сhangetisaurus estesi Nessov, 1992, череп и нижняя челюсть лабиально (а) и дорсально (б), ростральная часть черепа, экз. ЗИН РНТ, № F89 лабиально, увеличено (в) (по: Федоров, Несов, 1992, рис. на с. 126); средняя юра Киргизии; г, д – Parmeosaurus scutatus Gao et Norell, 2000, реконструкция черепа дорсально (г) и лабиально (д) (по: Gao, Norell, 2000, фиг. 25 A, D, F); поздний мел Монголии; е, ж – Becklesisaurus scincoides Hoffstetter, 1967, экз. BMNH, № R8082, фрагмент левой ветви нижней челюсти лабиально (е) и нижнечелюстной зуб (ж) (по: Hoffstetter, 1967, фиг. 7, А, часть, 7, С); титон–неоком Англии; з – Naimanosaurus dinosauroaequalis gen. et sp. nov. (Paramacellodidae, Scincomorpha), голотип ПИН, № 3334/516, правая зубная кость лингвально; Монголия, Увэрхангайский аймак, местонахождение Хобур; нижний мел, верхний апт–нижний альб, хухтыкский горизонт.
В соответствии с мнением Р. Эстеса (Estes, 1983), Paramacellodidae родственны Cordylidae s.l. Альтернативные точки зрения, предложенные в рамках общего кладистического анализа, связывают семейство с Anguimorpha (Conrad, 2008) или с базальными Scincomorpha (Gauthier et al., 2012). Первой гипотезе противоречит специализация строения нижней челюсти, а второй – специфика остеодермального покрова (исходно ящерицам не был свойственен ни сплошной остеодермальный панцирь, ни остеодермы прямоугольной формы). Действительно, по сравнению с Cordylidae s.l., парамацеллодиды обладают внешне мало специализированым черепом, что характерно для многих ископаемых групп сцинкоморф и ящериц в целом. Однако это обстоятельство не соответствует ряду конкретных деталей: в частности, строению неба или передней части височных дуг, которые по-разному преобразованы у самых разных групп сцинкоморф.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ОТРЯД LACERTILIA
ИНФРАОТРЯД SCINCOMORPHA
МИКРООТРЯД XANTUSIOMORPHA
СЕМЕЙСТВО EOXANTIDAE ALIFANOV, 1993
Типовой род – Eoxanta Borsuk-Białynicka, 1988; поздний мел Монголии.
Диагноз. Сцинкоморфы, обладающие платинотопалатинарным типом неба и развитыми конечностями. Височные окна значительно или полностью закрыты. Кости височных дуг уплощены. Вершины зубов конические или несут ростральное плечо.
Состав. Кроме типового, род Palaeoxanta gen. nov.; ранний мел Монголии.
Род Palaeoxanta Alifanov, gen. nov.
Название рода от palaios греч. – древний и Eoxanta.
Типовой вид – Palaeoxanta conicodentata sp. nov.
Диагноз. Нижний край зубных костей прямой, утолщения не имеет. Рострально меккелев канал ориентирован вниз. На зубных костях выражено 15 площадок для прикрепления зубов. Зубы имеют слабо притупленную вершинку. Рострального плеча у вершины нет. Коронки немного расширены в средней части, а ближе к основанию заметно заужены. Соседние зубы расположены тесно, иногда с небольшим промежутком.
Видовой состав. Типовой вид.
Сравнение. От типового новый род отличается спрямленным нижним краем зубных костей, вентральной ориентацией ростральной части меккелева желобка, тесным расположением и бóльшим числом зубов, а также отсутствием рострального плеча у их вершины.
Замечания. По экз. № 3142/360 из фондов Палеонтологического ин-та им. А.А. Борисяка РАН (ПИН) нами установлено, что у Eoxanta Borsuk-Białynicka, 1988 челюстные зубы несли небольшое ростральное плечо.
Palaeoxanta conicodentata Alifanov, sp. nov.
Табл. IХ, фиг. 1 (см. вклейку)
Slavoiidae gen. nov.: Алифанов, 2000, c. 61.
Slavoiidae [Slavoia sp.]: Алифанов, 2012, с. 18.
Название вида от conicus лат. – конический и dentatus лат. – озубленный.
Голотип – ПИН, № 3334/520, левая зубная кость; Монголия, Увэрхангайский аймак, местонахождение Хобур; ранний мел, верхний апт–нижний альб, хухтыкский горизонт, свита не определена.
Описание (рис. 2, г). Зубные кости короткие и высокие, их околосимфизная часть заметно изогнута медиально. Симфизная фасетка вытянута горизонтально, имеет субтреугольную форму. Лабиально на зубной кости сформировано шесть губных отверстий, которые отстоят друг от друга на неравных расстояниях. Последнее такое отверстие соответствует по положению уровню 12‑го зуба. Короноидный отросток зубных костей удлиненный. Фланг субдентального гребня широкий, незначительно расширен рострально. Судя по следам контактов, пластинчатая кость доходила до уровня передней трети сохранившейся части зубной кости. По прикреплению зубы эуплевродонтные (т.е., имеют промежуточный между суб- и гиперплевродонтностью тип); в середине ряда они немного крупнее, чем в его начале и в конце, но самые широкие зубы находятся в предпоследней четверти ряда. За край челюстной кости зубы выступают примерно на четверть своей высоты в ростальной части и предположительно на треть высоты – в окципитальной части.
Размеры голотипа в мм. Длина сохранившейся части зубной кости – 4.8, высота заднего края зубной кости – 2.1, высота и диаметр пятого и шестого зубов – 0.8 и 0.3.
Материал. Кроме голотипа, экз. ПИН, № 3334/555, правая зубная кость с зубами; типовое местонахождение.
МИКРООТРЯД LEPTOGLOSSA
СЕМЕЙСТВО ARDEOSAURIDAE CAMP, 1923
Типовой род – Ardeosaurus Meyer, 1860; поздняя юра Германии.
Диагноз. Безостеодермные и диплоглоссопалатинарные сцинкоморфы, обладающие эуплевродонтными по типу прикрепления и одновершинными по строению зубами. Медиальный отросток заглазничных костей всегда расположен ростральнее заднелобной кости.
Состав. Кроме типового, роды: Eichstaettisaurus Kuhn, 1958, поздняя юра Германии; Yabeinosaurus Endo et Shikama, 1942, Dalinghosaurus Ji, 1998, Jehololacerta Ji et Ren, 1999, Liaoningolacerta Ji, 2005, Liushusaurus Evans et Wang, 2010, ранний мел Китая; Sakurasaurus Evans et Manabe, 1999, ранний мел Японии; Myrmecodaptria Gao et Norell, 2000, поздний мел Монголии; Norellius Conrad et Daza, 2015, Gurvelus gen. nov., ранний мел Монголии.
Замечания. Ряд перечисленных выше форм, в том числе и новая, представлены фрагментарными материалами. В этом случае к данному семейству они отнесены по отсутствию остеодерм (характерно для всех ардеозавроидных лептоглосс) и по конической форме относительно немногочисленных зубов, как у юрско-меловых ардеозаврид.
Род Gurvelus Alifanov, gen. nov.
Название рода – от гyрвэл монг. – ящерица.
Типовой вид – Gurvelus khangaicus sp. nov.
Диагноз. Мозговая капсула широкая и уплощенная. Алярные отростки переднеушных костей небольшие. Медиально края зубной кости в предсимфизной части смыкаются на короткой дистанции. Зубы сравнительно небольшие, тесно посажены. Их заостренная вершинка уплощена, имеет симметрично расположенные плечи. Основание зубов заужено и сжато ростроокципитально. Число нижнечелюстных зубов – 20–22.
Видовой состав. Типовой вид.
Сравнение. От всех представителей семейства отличается замкнутостью краев зубной кости в ростральной части и уникальной формой зубов.
Gurvelus khangaicus Alifanov, sp. nov.
Табл. IX , фиг. 2–4
Название вида – от хр. Хангай.
Голотип – ПИН, № 3334/518, фрагментарный череп; Монголия, Увэрхангайский аймак, местонахождение Хобур; нижний мел, верхний апт–нижний альб, хухтыкский горизонт, свита не определена.
Описание (рис. 3, е–и). Склеротические пластины выражены, их не менее восьми. Диаметр каждой пластины составляет примерно два миллиметра. По вентральной длине основная затылочная кость короче парасфеноида. Сфенокципитальные бугорки небольшие. Основания парокципитальных отростков широкие и уплощены дорсовентрально. Скуловые кости дугообразные, тонкие, рострально немного расширены. Верхнечелюстные кости окципитально доходят до уровня середины глазниц. Небные кости относительно узкие, их птеригоидные отростки удлиненные. Крыловидные кости вентрально несут не менее двух рядов зубов. Ростральный отросток наружных крыловидных костей лежит дорсальнее супрадентального гребня верхнечелюстных костей (это признак диплоглоссопалатинарности). Он короче крыловидного отростка этих же костей.
Зубные кости прямые. На разрушенной постдентальной части надугловой и угловой костей сохраняются следы крепления отростков зубной кости: короноидного и ангулярного. Первый лежит под углом к оси челюсти и совпадает по направлению с лабиальным отростком венечной кости, а второй идет дальше заднего края короноидного отростка венечной кости. По длине зубные кости примерно равны постдентальной части ветви нижней челюсти. Пластинчатая кость рострально продолжается до передней трети зубного ряда, т.е., до начала смыкания краев зубных костей. Постдентальная часть ветви нижней челюсти изогнута и окципитально уплощена. Аддукторная яма щелевидная. Угловые кости широкие и окципитально пересекают уровень середины надугловой кости. Переднее супраангулярное отверстие расположено на уровне заднего края венечной кости и на уровне середины надугловой кости. Венечная кость высокая, ее лабиальный отросток выражен. Терминально он несет углубление (фасетку) для связи с короноидным отростком зубной кости. Параартикулярный отросток широкий и округлый. Ретроартикулярный отросток имеет обширную дорсальную выемку и сливается с параартикулярным отростком. Последний имеет округлый контур.
Зубы сравнительно небольшие. На зубной кости наиболее высокие из них расположены в средней части ряда. Резорбционное отверстие выражено на некоторых зубах. В основании всех зубов имеют место незначительные отложения цемента.
Размеры в мм. Общая длина черепа – 17‒18, реконструкция его орбитальной ширины – 8–9, длина нижней челюсти – 20, высота нижней челюсти в области венечной кости – 4, высота нижнечелюстных зубов в средней части ряда – 1.4.
Материал. Голотип.
Замечания. Кроме голотипа G. khangaicus gen. et sp. nov., в коллекции ардеозаврид из Хобура представлены лобные кости, которые могут быть определены только как Ardeosauridae fam. indet. Судя по экз. ПИН, № 3334/552 (табл. Х, фиг. 5) и № 3334/553, указанные кости обладают относительно узкой средней частью, зачаточными субольфакторными отростками, глубокими, широко отделенными друг от друга и очень небольшими фасетками для крепления носовых костей, остеодермальной скульптурой в виде отдельных бугорков или их групп на дорсальной поверхности (остеодермальные участки могут быть ограничены друг от друга отчетливо выраженными бороздами, обозначая широкие краевые щитки). Для лобной кости, экз. ПИН, № 3334/552, устанавливается длина в 8.7 мм, ростральная ширина в 2.8 мм (реконструкция – 3.5 мм), окципитальная ширина в 6 мм и ширина в средней части – 2 мм. Указанный тип лобных костей из Хобура более всего близок по общему строению и деталям к Dalinghosaurus longidigitus Ji, 1998 из раннего мела Китая.
СЕМЕЙСТВО GLOBAURIDAE ALIFANOV, 2000
Типовой род – Globaura Borsuk-Białynicka, 1988; поздний мел Монголии.
Диагноз. Безостеодермные и диплоглоссопалатинарные сцинкоморфы, обладающие коническими, двух- и трехвершинными зубами, а также особым типом связи заглазничных и заднелобных костей: медиальный отросток первых расположен между парой латеральных отростков вторых.
Состав. Кроме типового, роды: Meyasaurus Vidal, 1915, ранний мел Испании; Durotrigia Hoffstetter, 1967, титон–неоком Англии; Tijubina Bonfim Junior et Marques, 1997, ранний мел Бразилии; Khereidia gen. nov., ранний мел Монголии.
Род Khereidia Alifanov, gen. nov.
Название рода – от древнемонгольского этнонима кераиты (керейты).
Типовой вид – Khereidia aptiana sp. nov.
Диагноз. Фланг субдентального гребня зубных костей узкий, рострально не расширен. Меккелев желобок в симфизной области ориентирован вентрально. Число зубов – не более 19–20. Зубы цилиндрические в поперечном сечении, увеличиваются окципитально, иногда немного наклонены и дугообразно изогнуты в средней части ряда. В ростральной части ряда они конические, в средней – двухвершинные, в окципитальной – трехвершинные. Боковые вершинки отделены от осевой разделительным желобком. Задняя вершинка уже, но выше передней.
Видовой состав. Типовой вид.
Сравнение. От других представителей семейства новый род отличается относительно небольшим числом нижнечелюстных зубов и слабым развитием ростральной вершинки. От Globaura Borsuk-Białynicka, 1988 дополнительно отличается серией трехвершинных зубов, а от Durotrigia Hoffstetter, 1967 – широкой окципитально зубной костью и трехвершинностью только окципитальной серии зубов.
Замечания. Филогенетически новый род, предположительно, близок к Durotrigia Hoffstetter, 1967 (рис. 4, ж) из формации Пурбек (титон–неоком) Англии по развитию несимметричной трехвершинности, поскольку в составе Globauridae только они демонстрируют данный признак.
Khereidia aptiana Alifanov, sp. nov.
Табл. IХ, фиг. 6
Название вида от апта.
Голотип – ПИН, № 3334/523, зубная кость; Монголия, Увэрхангайский аймак, местонахождение Хобур; ранний мел, верхний апт–нижний альб, хухтыкский горизонт, свита не определена.
Описание (рис. 4, з). Зубная кость постепенно и заметно расширяется окципитально. Субдентальный гребень этой кости сравнительно узкий и умеренно расширен рострально. Губные отверстия мелкие, их восемь. Зубы прямые, умеренной высоты, эуплевродонтные по прикреплению; их основания незначительно расширены. Оси зубов немного отклоняются от прямого по отношению к линии зубного края угла. В передней половине они наклонены назад, а в задней – вперед. На образце сохранилось 16 плевродонтных по прикреплению зубов.
Размеры голотипа в мм. Длина (реконструкция) зубной кости – 5.9 (7.5), окципитальная высота ее сохранившейся части – 1.8, высота средних зубов – 0.8.
Материал. Голотип.
СЕМЕЙСТВО PARAMACELLODIDAE ESTES, 1983
Типовой род – Paramacellodus Hoffstetter, 1967; поздняя юра Северной Америки, ранний мел Англии.
Диагноз. Верхние височные окна широко открыты. Теменная кость формирует длинные и относительно узкие в основании затылочные отростки. Зубы конические или притупленные, дополнительных вершинок не образуют. Остеодермы в районе лобных костей крыши черепа крупные, обычно срастаются с этими костями. Свободные остеодермы тела образуют сплошной панцирь. В подавляющем большинстве они прямоугольные по форме, иногда с килеватой внешней поверхностью. В подбородочной области остеодермы могут иметь округлую форму каудального края.
Состав. Кроме типового, роды: Changetisaurus Nessov, 1992, средняя юра Казахстана; Sharovisaurus Hecht et Hecht, 1984, поздняя юра Казахстана; Mimobecklesisaurus Li, 1985, поздняя юра Китая; Saurillodon Estes, 1983, поздняя юра Португалии; Becklesius Estes, 1983, поздняя юра Португалии и титон–неоком Англии; Becklesisaurus Hoffstetter, 1967, Pseudosaurillus Hoffstetter, 1967, титон–неоком Англии; Parmeosaurus Gao et Norell, 2000, поздний мел Монголии; Naimanosaurus gen. nov., ранний мел Монголии.
Род Naimanosaurus Alifanov, gen. nov.
Название рода от древнемонгольского этнонима найманы и sauros греч. – ящерица.
Типовой вид – Naimanosaurus dinosauroaequalis sp. nov.
Диагноз. Зубная кость низкая. Фланг субдентального гребня умеренной высоты, слабо расширяется рострально. Зубы сравнительно небольшие, прямые или немного искривлены; тип их прикрепления эу- и субплевродонтный. Вершинки зубов заостренные и несут симметрично расположенные плечи. Число нижнечелюстных зубов, судя по числу мест их прикрепления – 18.
Видовой состав. Типовой вид.
Сравнение. В составе семейства новый род представляет собой линию крупнозубых форм, к числу которых относятся Becklesisaurus Hoffstetter, 1967 (Paramacellodus oweni sensu Estes, 1983) из титон–неокома Англии (рис. 5, е, ж) и Changetisaurus Nessov, 1992 из средней юры Киргизии. От первого новая форма отличается четко выделенными плечами зубов и большей продвинутостью в сторону субплевродонтности, а от второго – небольшими, изогнутыми и более многочисленными зубами.
Замечания. Отнесение нового рода к Paramacellodidae основано на сходстве строения его зубов с зубами Changetisaurus Nessov, 1992 из средней юры Кыргызстана (Федоров, Несов 1992), который нами признается несомненным представителем семейства.
Naimanosaurus dinosauroaequalis Alifanov, sp. nov.
Табл. IХ, фиг. 7
Paramacellodidae gen. indet.: Алифанов, 2000, с. 65, рис. 28 , а; Алифанов, 2012, с. 31.
Название вида от Dinosauria и aequalis лат. – современник.
Голотип – ПИН, № 3334/516, правая зубная кость с зубами; Монголия, Увэрхангайский аймак, местонахождение Хобур; нижний мел, верхний апт–нижний альб, хухтыкский горизонт, свита не определена.
Описание (рис. 5, з). Зубные кости относительно низкие, их лабиальная сторона выпуклая и округлая. Ангулярный отросток широкий и длинный. Короноидный отросток зубных костей короткий, равен по размерам супраангулярному отростку этих же костей. Число губных отверстий – семь. Зубы постепенно увеличиваются окципитально. Их основание немного сжато ростроокципитально.
Размеры голотипа в мм. Длина зубной кости – 11, высота ее заднего края – 2.8, высота крупнейшего нижнечелюстного зуба – 1.2.
Материал. Голотип.
Список литературы
Аверьянов А.О., Федоров П.В. Раннемеловая ящерица из Ходжиабадской свиты Ферганы (Кыргызстан) // Вестн. СПбГУ. 2004. Сер. 7. Вып. 1. С. 81–82.
Алифанов В.Р. Новые ящерицы семейства Macrocephalosauridae (Sauria) из верхнего мела Монголии и критические замечания о системе Teiidae (sensu Estes, 1983) // Палеонтол. журн. 1993а. № 1. С. 57–74.
Алифанов B.Р. Ящерицы верхнего мела Монголии и проблема первого межамериканского контакта // Палеонтол. журн. 1993б. № 3. С. 79–85.
Алифанов В.Р. Макроцефалозавры и ранние этапы эволюции ящериц Монголии // Тр. Палеонтол. ин-та РАН. 2000. Т. 272. 126 с.
Алифанов В.Р. Отряд Lacertilia // Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Часть 2 / Ред. Курочкин Е.Н., Лопатин А.В. М: ГЕОС, 2012. С. 7–136.
Алифанов B.Р. Ящерицы семейства Hodzhakuliidae (Scincomorpha) из раннего мела Монголии // Палеонтол. журн. 2016. № 5. С. 73–82.
Иорданский Н.Н. Кинетизм черепа ящериц: О функциональном значении кинетизма черепа // Зоол. журн. 1966. Т. 45. № 9. С. 1398–1410.
Иорданский Н.Н. Эволюция комплексных адаптаций: Челюстной аппарат амфибий и рептилий. М.: Наука, 1990. 312 с.
Федоров П.В., Несов Л.А. Ящерица рубежа средней и поздней юры Северо-Восточной Ферганы // Вестн. СПбГУ. 1992. Сер. 7. Вып. 3. № 21. С. 9–14.
Хехт М.К., Хехт Б.М. Новая ящерица из юрских отложений Средней Азии // Палеонтол. журн. 1984. № 3. С. 135–138.
Alifanov V.R. Some peculiarities of the Cretaceous and Palaeogene lizard faunas of the Mongolian People’s Republic // Kaupia. 1993. H. 3. P. 9–13.
Averianov A.O., Skutchas P.P. Paramacellodid lizard (Squamata, Scincomorpha) from the early Cretaceous of Transbaikalia // Russ. J. Herpetol. 1999. V. 6. № 2. P. 115–117.
Borsuk-Białynicka M. Globaura venusta gen. et sp. n. and Eoxanta lacertifrons gen. et sp. n. – non-teiid lacertoids from the Late Cretaceous of Mongolia // Acta Palaeontol. Pol. 1988. V. 33. № 3. P. 211–248.
Camp C.L. Classification of the lizards // Bull. Amer. Mus. Natur. Hist. 1923. V. 48. P. 289–481.
Conrad J.L. Phylogeny and systematics of Squamata (Reptilia) based on morphology // Bull. Amer. Mus. Natur. Hist. 2008. V. 310. 182 p.
Conrad J.L., Daza J.D. Naming and rediagnosing the Cretaceous Gekkonomorph (Reptilia, Squamata) from Öösh (Övörkhangai, Mongolia) // J. Vertebr. Paleontol. 2015. V. 35. № 5. P. e980891 (5 p.).
Daza J.D., Alifanov V.R., Bauer A.M. A redescription and phylogenetic reinterpretation of lizard Hoburogekko suchanovi Alifanov 1989 (Squamata: Gekkota) // J. Vertebr. Paleontol. 2012. V. 32. № 6. P. 1303–1312.
Estes R. Sauria terrestria, Amphisbaenia. Stuttgart, N.Y.: G. Fischer Verlag, 1983. 249 s. (Handbuch der Paläeoherpetologie. T. 10A).
Estes R., de Queiroz K., Gautier J. Phylogenetic relationships within Squamata // Phylogenetic relationships within lizard families. Essays commemorating Charles L. Camp / Eds. Estes R., Pregill G. Stanford: Univ. Press, 1988. P. 119–281.
Evans S.E., Barbadillo L.J. Early Cretaceous lizards from Las Hoyas, Spain // Zool. J. Linn. Soc. 1997. V. 119. № 1. P. 23–49.
Evans S.E., Wang Y. The early Cretaceous lizard Dalinghosaurus from China // Acta Palaeontol. Pol. 2005. V. 50. № 4. P. 725–742.
Evans S.E., Wang Y. New material of the Early Cretaceous lizard Yabeinosaurus from China // Cret. Res. 2012. V. 34. P. 48–60.
Evans S.E., Wang Y., Jones M.E.H. An aggregation of lizard skeletons from the Lower Cretaceous of China // Senckenb. lethaea. 2007. Bd 87. № 1. S. 109–118.
Gao K., Norell M.A. Taxonomic revision of Carusia (Reptilia: Squamata) from the late Cretaceous of the Gobi Desert and phylogenetic relationships of anguimorphan lizards // Amer. Mus. Novit. 1998. № 3230. P. 1–51.
Gao K., Norell M.A. Taxonomic composition and systematics of late Cretaceous lizard assemblages from Ukhaa Tolgod and adjacent localities, Mongolian Gobi Desert // Bull. Amer. Mus. Natur. Hist. 2000. V. 249. P. 1–118.
Gauthier J.A., Kearney M., Maisano J.A. et al. Assembling the squamate tree of life: Perspectives from the phenotype and the fossil record // Bull. Peabody Mus. Natur. Hist. 2012. V. 53. № 1. P. 3–308.
Hoffstetter R. Les Sauria (=Lacertilia) du Jurassique supérieur du Montsech (Espagne) // Bull. Soc. Géol. France. 1966. Sér. 7. T. 7. P. 549–557.
Hoffstetter R. Coup d’oeil sur les sauriens (=lacertiliens) des couches de Purbeck (Jurassique supérieur d’Angleterre) // Problèmes Actuels de Paléontologie (Évolution des vertébrés). Coll. Intern. Centre Nat. Rech. Sci. 1967. № 163. P. 349–371.
Lucas S.G. The Psittacosaurus biochron, Early Cretaceous of Asia // Cret. Res. 2006. V. 27. № 2. P. 189–198.
Mateer N. Osteology of the Jurassic lizard Ardeosaurus brevipes (Meyer) // Palaeontology. 1982. V. 25. Pt 3. P. 461–469.
Reynoso V.-H., Callison G. A new scincomorph lizard from the early Cretaceous of Puebla, México // Zool. J. Linn. Soc. 2000. V. 130. № 2. P. 183–212.
Shuvalov V.F. The stratigraphy and palaeobiogeography of Mongolia // The age of dinosaurs in Russia and Mongolia / Eds. Benton M.J., Shishkin M.A., Unwin D.M., Kurochkin E.N. Cambridge: Univ. Press, 2000. P. 256–278.
Simões T. Redescription of Tijubina pontei, an Early Cretaceous lizard (Reptilia; Squamata) from the Crato Formation of Brazil // An. Acad. Brasil. Ciênc. 2012. V. 84. № 1. P. 79–93.
Simões T.R., Caldwell M.W., Nydam R.L., Jiménez-Huidobro P. Osteology, phylogeny, and functional morphology of two Jurassic lizard species and the early evolution of scansoriality in geckoes // Zool. J. Linn. Soc. 2017. V. 180. № 1. P. 216–241.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал