

Палеонтологический журнал, 2019, № 1, стр. 24-33
О составе моллюсков тавдинской свиты эоцена Западной Сибири
С. В. Попов a, *, Я. С. Трубин b, П. В. Смирнов b, c, В. В. Ордовский, И. А. Гончарова a, О. В. Амитров a
a Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
b Тюменский индустриальный университет
Тюмень, Россия
c Клаустальский технический университет
Клаусталь-Целлерфельд, Германия
* E-mail: serg.pop@mail.ru
Поступила в редакцию 05.04.2017
Принята к публикации 17.05.2018
Аннотация
Изучен состав двустворчатых моллюсков и гастропод из глин тавдинской свиты, найденных в Кыштырлинском карьере в 23 км к югу от г. Тюмень и в керне скважин 8 и 11 к югу от г. Омск. Малакофауна Кыштырлинского карьера представлена сидеритовыми ядрами и отпечатками, реже целыми раковинами, иногда двустворчатыми экземплярами, среди которых удалось определить 19 форм. В скважинном материале определены восемь видов восьми родов. Несомненным является сходство этих комплексов с чеганской фауной Северного Устюрта, Приаралья и Тургая, что говорит как об одновозрастности этих комплексов, так и о прямых палеогеографических связях этих бассейнов. Оба комплекса свидетельствуют о морских, вероятно, несколько опресненных условиях и относительно небольших глубинах накопления осадков ниже волноприбойной зоны. Газовый режим менялся от благоприятного для бентоса до условий дефицита кислорода.
ВВЕДЕНИЕ
Отложения тавдинской свиты широко распространены в Западной Сибири и слагают значительный объем приповерхностных отложений как в обрамлении Западно-Сибирской низменности, так и в Зауралье и в северной части Тургайского прогиба. Тавдинская свита характеризуется глинистым составом и завершает разрез морского палеогена, отражая регрессивную фазу развития бассейна. Формирование ее осадков было связано с последним этапом существования морского режима в истории Западно-Сибирского бассейна, при его устойчивой связи с Кавказско-Копетдагским морем через Тургайский прогиб.
Для тавдинской свиты до сих пор остается актуальным вопрос ее возраста и стратиграфического положения. По микропалеонтологическим данным: фораминиферам, остракодам, пыльце и спорам наземных растений он устанавливался как позднеэоценовый или даже олигоценовый (Подобина, Чернышов, 2012). Согласно последним исследованиям органикостенного микрофитопланктона, а также планктонных и бентосных фораминифер (Ахметьев и др., 2004, 2010), нижнетавдинская подсвита Западной Сибири датируется бартонским ярусом среднего эоцена, а верхнетавдинская – приабонским ярусом верхнего эоцена (Iakovleva et al., 2000; Ахметьев и др., 2004; Ахметьев, Беньямовский, 2006; Васильева и др., 2006; Васильева, 2015). Эти данные уже вошли в принятую ныне стратиграфическую схему региона (Унифицированная …, 2001).
Моллюски в Кыштырлинском карьере были собраны В.В. Ордовским, Я.С. Трубиным и П.В. Смирновым (2012–2016 гг.), коллекция хранится в Музее Тюменского Клуба юных геологов (МТКЮГ) и у Ордовского. Малакологический материал из скважин 8 и 11 в районе г. Омск был передан С.В. Попову и И.А. Гончаровой В.Н. Бень-ямовским и хранится в лаб. моллюсков Палеонтологического ин-та им. А.А. Борисяка РАН (ПИН).
Макрофауна тавдинской свиты изучена еще очень слабо. Это связано как с плохой обнаженностью отложений свиты, так и с редкостью находок. Поэтому определение и описание состава собранных моллюсков представляется важным не столько для целей стратиграфии, сколько для восстановления экологических условий в тавдинском бассейне, его палеонтологической характеристики, гидродинамического, солевого и газового режима, палеогеографических связей.
МОЛЛЮСКИ ТАВДИНСКОЙ СВИТЫ
Моллюски из Кыштырлинского карьера, расположенного в 4 км к юго-востоку от пос. Винзили, в 2 км к северо-востоку от д. Кыштырла, в 5 км к северо-западу от железнодорожной станции Богандинская и в 23 км к югу от г. Тюмени (рис. 1). Породы представлены толщей бейделлитовых зеленовато- и голубовато-серых глин, видимой мощностью до 20–25 м. В основании разреза глины однородные, неслоистые, с редкими мелкими включениями кристаллов пирита. По минералогическому составу они представлены гидрослюдистыми глинами с незначительным содержанием минералов хлоритовой группы. Выше глины становятся слоистыми; слоистость обусловлена присыпками и мелкими линзочками алеврита, встречаются небольшие стяжения марказита и караваеобразные включения сидерита, размером до 0.3 м. Глины тавдинской свиты с размывом перекрываются четвертичными аллювиальными отложениями первой и второй надпойменных террас. Последние представлены зеленовато-серыми, серыми кварц-полевошпатовыми песками с линзами и прослоями (до 3–4 м) буровато-серых плотных глин. На контакте глин тавдинской свиты с вышележащими породами встречаются друзы и кристаллы гипса.
Рис. 1.
Cхема расположения Кыштырлинского карьера, его положение на геологической карте и фотография карьера. Обозначения: 1 – скважины 8 и 11 к югу от г. Омск; 2, 3 – Кыштырлинский карьер.
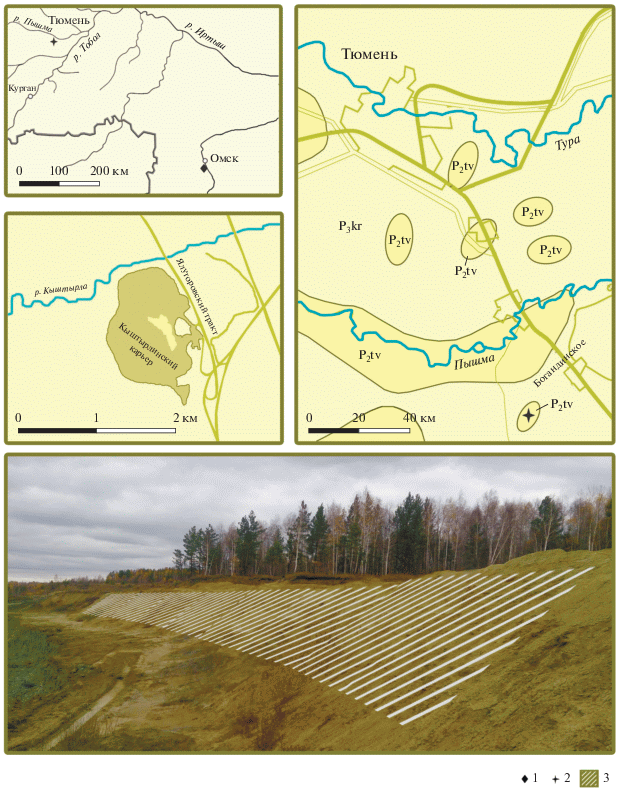
Ископаемые остатки моллюсков, которые послужили объектом исследования, сохранились как в виде отпечатков и ядер (иногда сложных ядер, несущих наружную скульптуру), так и в виде раковин. Иногда встречаются двустворчатые экземпляры, захороненные в прижизненном положении и более или менее равномерно распределенные по всей толще глин, вскрытой карьером. Ключевую роль в полноте данных по этому местонахождению сыграли многолетние сборы Ордовского и сделанные им фотографии, которыми в основном и проиллюстрирована эта работа.
Встречено несколько крупных линзовидных конкреций и плит, набитых раковинами, перенесенными сюда течениями, с резким доминированием в них остатков двустворчатых моллюсков, вложенных друг в друга (рис. 2), предположительно рода Arctica, и брюхоногих моллюсков рода Turritella, с единичными остатками гетеродонтных двустворчатых моллюсков (рис. 3). Раковины Turritella, по данным Ордовского, приурочены только к верхней части отложений, вскрытых в карьере. Они имеют размер от 2.0 до 3.5 см, реже до 4.0 см, ориентированы хаотично, но чаще в плоскости напластования. Кроме двустворчатых и брюхоногих моллюсков, на некоторых раковинах видны мшанки, селившиеся на них, как на субстрате; сохранились и следы прикрепления балянусов.
Рис. 2.
Скопление раковин преимущественно двустворчатых моллюсков, транспортированных течениями, обр. МТКЮГ № 1/100.

Рис. 3.
Скопление отпечатков раковин туррителлидных гастропод Torculoidea cf. oxyceras, обр. МТКЮГ № 1/101.

Так как весь материал, доступный для изучения, представлен лишь немногими экземплярами, часто ядрами и отпечатками, и имеется очень мало литературных данных по этой фауне, определить многие формы удалось лишь в открытой номенклатуре, часто со знаком cf. В этой фауне несомненно присутствие новых видов, описать которые пока не представляется возможным, так как все они имеют либо недостаточно хорошую сохранность, либо представлены единичными экземплярами. Список определений насчитывает 19 форм, определенных с разной степенью достоверности.
Двустворчатые моллюски: Nucula cf. kasakstanica Ruchin, Nuculana sp. nov. 1 (табл. III , фиг. 1; см. вклейку), Nuculana sp. 2, Yoldia sp. nov. (табл. III , фиг. 2), Anomia psamatheis Bayan (табл. III , фиг. 8, 9), Cultellus orientalis Alexeiev (рис. 4), Arctica alexeevi (Ovetschkin) (табл. III , фиг. 6), A. cf. rotundata (A. Braun), Pelecyora (Cordiopsis) polytropa Anderson (табл. III , фиг. 7), Pitar sp., ?Mya sp. (табл. III , фиг. 10), Sphenia kirgizensis (Romanovsky) (рис. 5), Panopea intermedia (Sowerby) (рис. 6).
Рис. 4.
Cultellus orientalis Alexeiev, обр. МТКЮГ № 1/111, двустворчатые экземпляры, захороненные на месте жизни.

Рис. 5.
Sphenia kirgizensis (Romanovsky), обр. МТКЮГ № 1/112, раковины на породе, ×1.5: а – изнутри, виден глубокий синус и хондрофор левой створки (вверху) и отвечающая ему выемка под макушкой правой створки (внизу); б – раковины снаружи.

Рис. 6.
Panopea intermedia (Sowerby), экз. МТКЮГ № 1/113, сложное ядро правой створки с хорошо сохранившейся наружной скульптурой, ×1.

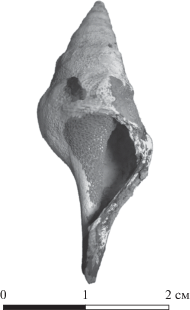
Гастроподы: Aporrhais sp. (табл. III , фиг. 5), Euspira achatensis Recluz (рис. 7), Sigaretotrema clathrata (Gmelin) (табл. III , фиг. 3), Scalaspira grossiventrosa (Amitrov) (рис. 8), Bathytoma sp. (рис. 9), Ringicula sp. nov. (табл. III , фиг. 4).
Рис. 7.
Euspira achatensis Recluz, экз. МТКЮГ № 1/114: а – со стороны макушки; б – сбоку; в – со стороны пупка. Размер линейки здесь и далее соответствует 1 см.

Рис. 8.
Scalaspira grossiventrosa (Amitrov), экз. МТКЮГ № 1/116, со слегка обломанным устьем: а – сбоку, б – со стороны устья.

Рис. 9.
Bathytoma sp., экз. МТКЮГ № 1/115, окатанный экземпляр с обломанным устьем, обросшим мшанкой.
Многие одиночные раковины представлены парными створками (Arctica alexeievi, Nucula, Nuculana, Yoldia, Cultellus orientalis, Pitar, Sphenia kirgizensis, ?Mya) и являются автохтонными, захороненными на месте жизни.
Моллюски из скважин 8 и 11 к югу от г. Омск. Кроме моллюсков, собранных в карьере, был исследован состав этой группы из двух скважин к югу от г. Омск.
Скважина 8 расположена в 8 км к югу от г. Омск, моллюски происходят с глубин 132–134 м из глин тавдинской свиты.
С глубины 132.5 м поднят двустворчатый экземпляр Arctica alexeievi (рис. 10, а) достаточно хорошей сохранности и многочисленные фрагменты крупных раковин, включая характерные обломки замка арктик (рис. 10, б). Двустворчатый экземпляр этого вида характеризуется выпуклой раковиной с заметно выступающей макушкой и четко выраженным килевым перегибом, что отличает его от другого известного здесь и в чегане вида – A. rotundata.
Рис. 10.
Моллюски тавдинской свиты из скв. 8, к югу от г. Омска, ×1: a, б – Arctica alexeievi (а – экз. ПИН, № 1470/43301, двустворчатый экземпляр со стороны левой створки, б – экз. ПИН, № 1470/43302, замок правой створки); в – Cultellus orientalis, экз. ПИН, № 1470/43303, левая створка изнутри.

С глубины 134.4 м определен вид Cultellus orientalis (рис. 10, в), представленный также двустворчатым экземпляром. В результате препаровки удалось вскрыть замок и большую часть левой створки изнутри. Этот вид ранее был описан из чегана (Алексеев, 1960; Мерклин, 1974), но в коллекции ПИН экземпляр такой сохранности, с замком левой створки, отсутствовал. Кроме этих видов, были найдены два отпечатка с фрагментами раковины, определенные как Nucula sp. Захоронение автохтонное, о чем свидетельствуют парные створки. Обе формы, определенные до вида, ранее были описаны из чеганской свиты Приаралья и Тургая.
В скважине 11 к юго-востоку от Омска моллюски найдены на глубинах 290–333 м в зеленовато-серых неясно слоистых алевритистых глинах с блестками слюды тавдинской свиты.
На глубине 290.8 м найдены замок раковины Pitar sp., неполная раковина Callista sp. и примакушечный фрагмент левой створки Lutraria sp. juv., напоминающий вид L. aralensis Merklin (Мерклин, 1974, с. 130, табл. 43 , фиг. 7–10), так же, как и этот вид, отличающийся от типичных представителей рода более выступающей макушкой. От других представителей рода Lutraria и от вида L. aralensis из олигоцена Северного Устюрта этот экземпляр отличается довольно хорошо развитыми боковыми зубами, отодвинутыми от макушки.
На глубине 330.5 м найдены Pitar cf. circularis (Koenen) – двустворчатый экземпляр и фрагменты раковин с замком; обломок раковины Callista sp., отличающийся от известных из чегана C. latilamella (Lukovic) более высокой раковиной; и несколько неполных экземпляров Pelecyora (Cordiopsis) cf. polytropa с обломками замков.
На глубине 333.1 м найдена неполная раковина и противоотпечаток Musculus cf. samensis Merkl., описанный Р.Л. Мерклиным (1974) из чеганской свиты Северного Устюрта.
С глубины 333.5 м была поднята раздробленная раковина Aporrhais cf. cornuta I. Korobkov.
ЗАМЕЧАНИЯ К СИСТЕМАТИЧЕСКОМУ СОСТАВУ МОЛЛЮСКОВ
Nuculana sp. nov. 1 (табл. III , фиг. 1), найденная в Кыштырлинском карьере, наиболее близка к виду N. aff. galeottiana (Nyst), известному из хатта Бельгии и Германии, но отличается более крупной толстостенной раковиной со значительно большим числом ребер (более 30, против 15–18 у N. galeottiana, см. Marquet et al., 2012) и более широкой макушкой, чем у этого западноевропейского вида (Albrechl, Valk, 1943, табл. 9 , рис. 326–331 ).
Yoldia sp. nov. (табл. III , фиг. 2) значительно отличается от единственного вида этого рода – Y. khanabadensis (Ivanova), известного в чеганской и белоглинской свитах эоцена и пшехской свите олигоцена, более высокой раковиной субквадратных очертаний с более центральной макушкой и характером выпуклости, распределенной более равномерно.
Arctica alexeievi (Ovetschkin) (табл. III , фиг. 6) – эндемичный вид, ранее известный только в чегане Устюрта, Приаралья и Тургая (Овечкин, 1954, с. 124; Мерклин, 1974, с. 86, 87). От встречающегося совместно с ним A. rotundata (Braun) отличается более выпуклой и более удлиненной раковиной с двумя килевыми перегибами, идущими к задне-нижнему углу раковины и отделяющими щиток.
?Mya sp. nov. (табл. III , фиг. 10). По форме раковины изображенный экземпляр более всего похож на современных и неогеновых представителей рода Mya. Однако для уверенного определения родовой принадлежности необходимо изучить замок и связочный аппарат, которые остались не вскрытыми.
Sigaretotrema clathrata (Gmelin) (табл. III , фиг. 3). Этот вид ранее определяли, как “Sigaretus clathratus” (Буртман в: Липман и др., 1960, с. 121, табл. 18 , фиг. 8а–г; Алексеев, 1963, с. 58, 59, табл. 12 , фиг. 23–26). Их сходство подтвердил и французский исследователь Ж. Пако (J.-M. Pacaud), изучающий эту группу, по посланной ему фотографии. Он прислал нам фотографии топотипов S. clathrata из лютета Парижского бассейна (рис. 11). Эта форма отличается от вида “Sinum rotundatum (Koenen)”, известного в чегане (Амитров, 2005, с. 43), более широким открытым завитком, глубоким пупком и четким подразделением ребер последнего оборота на основные и вставочные, что также сближает ее с типичными S. clathrata из Парижского бассейна.
Рис. 11.
Sigaretotrema clathrata (Gmelin, 1791), Национальный музей естественной истории, Париж; лютет Парижского бассейна.

Scalaspira grossiventrosa (Amitrov) (рис. 8) описана из нижнего олигоцена, верхней части ащеайрыкской свиты Северного Устюрта (Амитров, 1966), и неизвестна из чеганской свиты. От типовых олигоценовых тавдинская форма отличается хуже выраженной концентрической скульптурой, отчетливой лишь на нижней части последнего оборота. Тем не менее, не располагая более представительным материалом, мы пока не склонны придавать этим отличиям систематического значения.
Ringicula sp. nov. (табл. III , фиг. 4). В чеганской свите ренгикулы неизвестны, а несколько близких по форме раковины латдорфских видов отличаются от тавдинской формы строением устья.
Bathytoma sp. (рис. 9). От чеганского вида этого рода – B. alexeevi Amitrov et Mironova (=“Pleurotoma tuberculata” Alexeev) отличается расположением бугорчатой скульптуры на верхнем склоне оборота, а не на центральном ребре.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Сведения о находках моллюсков в тавдинской свите немногочисленны. Двустворчатые моллюски и гастроподы тавдинской свиты были описаны Е.С. Буртман (Липман и др., 1960), но представленные на таблицах моллюски не отличаются достаточно хорошим качеством, поэтому многие определения остаются сомнительными. Несомненным по этим данным представляется присутствие родов и нескольких видов двустворок: Nucula, Nuculana (Leda у Буртман), Chlamys (Chlamys) meissonieri (Mayer) [=Pecten (Pecten) picta], Siliqua, Arctica (=Cyprina), Astarte, Pitar [=Meretrix (Pitaria)], Callista, Pelecyora (Cordiopsis) [Meretrix (Cardiopsis) у Буртман], Varicorbula gibba (Olivi) [у Буртман, табл. 17 , фиг. 11а – Marcia (Textivenus) texta Lmk], Sanguinolaria binotata (=Libitina и Panopea у Буртман) (подчеркнуты формы, дополняющие описанный выше материал), а также 10 семейств гастропод (Амитров, 1993, с. 62). Все семейства и роды известны также и в чегане (Миронова и др., 1962; Алексеев, 1963; Мерклин, 1974; Амитров, 1993). Вид Chlamys (Chlamys) meissonieri, в синонимику которого следует включить и Chlamys kassini Ovetschkin, 1954 (Овечкин, 1954, c. 126, 127, табл. 4 , фиг. 2), Н.К. Овечкиным был впервые отмечен в верхах эоцена (зона Pinna lebedevi) центральной части Тургайской впадины. Наиболее тепловодными в этой фауне являются гастроподы: туррителлиды и синиды (Амитров, 1993, с. 105).
Двустворчатые моллюски, гастроподы, брахиоподы в рассматриваемых отложениях указывались при проведенном в Западной Сибири геологическом картировании (Государственная …, 2011). Список моллюсков из верхней подсвиты приведен В.А. Мартыновым и др. (1975, с. 322) в томе “Палеогеновая система”. Находки единичных моллюсков упоминаются в работе Л.Б. Бакиевой (2005, с. 14). Однако эти отрывочные данные уже не дополняют существенно приведенные списки.
Экологические условия в Тавдинском бассейне. Все перечисленные более или менее представительные находки малакофауны происходят, вероятно, из одной батиметрической зоны Тавдинского бассейна. В основном это – неглубоко зарывающаяся инфауна, обитавшая на мягких илисто-алевритовых грунтах на небольших глубинах, но вне волноприбойной зоны. Поэтому мы можем судить лишь об этой краевой зоне в юго-восточной части Тавдинского моря, привлекая данные по ближайшим смежным бассейнам.
Несомненно, в приабоне здесь продолжали существовать морские условия, но было ли Тавдинское море полносоленым? Во всяком случае, в составе двустворчатых моллюсков преобладали относительно эвригалинные группы, способные жить при довольно значительном опреснении. Представители большинства встреченных здесь родов выдерживают понижение солености до 20‰, и лишь присутствие Anomia, Nuculana, Aporrhais может свидетельствовать о несколько большей солености. То же можно сказать о массовых в верхах разреза туррителлидах, выдерживающих понижение солености до 20‰ (Татишвили и др., 1968; Fretter, Gracham, 1981). О неполной солености и специфических условиях может свидетельствовать и относительно бедный состав этой фауны, иногда с резким доминированием немногих видов туррителл (рис. 3), корбулид (Липман и др., 1960, табл. 17 , фиг. 11а); даже при столь неполных данных обращает на себя внимание сходство систематического состава из разных местонахождений, представленного небольшим числом таксонов.
Опресненность верхнего слоя воды, приводившая к резко выраженной галинной стратификации, для этого бассейна указывалась ранее по диноцистовым данным (Akhmet’ev, 2011). Стратификация вод должна была иметь следствием затрудненный газовый режим в основной акватории бассейна. Последним, вероятно, и объясняется редкость находок макрофауны, обедненность комплексов и наблюдаемая нами (см. выше) и описанная в литературе (Мартынов и др., 1975), а также частая пиритизация глин.
Если привлечь фаунистические данные по прилегающей тургайской части Чеганского бассейна [по разрезам Атамбас-чинк, Челкар-Нура; сборы О.В. Амитрова и Л.В. Мироновой (ВСЕГЕИ), переданные в ПИН], то кажется несомненной резкая галинная стратификация вод и в Тургайском проливе, которая могла быть обусловлена опресненностью вод, выходивших из Тавдинского моря, и большей соленостью чеганских вод, поступавших в Тавдинский бассейн. Эту ситуацию можно сравнить с современной в Мраморном и Азовском морях, где у поверхности идут опресненные воды речного стока, а глубже располагается водная масса морского генезиса. Такая стратификация приводит к дефициту кислорода на дне, частым заморам при отсутствии ветрового перемешивания, и повторным заселениям дна одной возрастной генерацией одного или немногих видов. Резко доминировавшие виды в таких условиях могут сменять друг друга. Так, в акватории Азовского моря C.B. Попову удавалось наблюдать большие площади, заселенные после замора годовалыми моллюсками Cerastoderma glaucum в районе Таганрога. В Тургайском проливе в чегане такими доминантами чаще всего становились Cultellus orientalis, Callista latilamella, Musculus aff. bernai, Lentidium donaciforme из двустворчатых моллюсков и Turritella oxiceras, T. paucicarinata, Sigmesalia fasciata termenbesica среди гастропод.
В составе моллюсков практически отсутствуют группы, характерные для тепловодных субтропических морей. Среди гастропод наиболее тепловодными являются представители семейств Sinidae и Turritellidae. Современные представители последнего семейства живут в теплых и умеренных водах до берегов Англии (Fretter, Gracham, 1981), а в палеогене встречались в относительно тепловодных бассейнах северного типа (Амитров, 1993, с. 105). Представители родов Arctica, Astarte, Yoldia, Mya, широко распространившиеся в северных и умеренно-тепловодных морях только с олигоценового времени, можно считать холодноводными для эоцена группами. Вероятно, тавдинские моллюсковые комплексы следует оценивать, как умеренно тепловодные, и наиболее холодноводными из известных эоценовых ассоциаций атлантического происхождения. Если в дальнейшем подтвердится правильность определения родовой принадлежности ?Mya, это будет одна из самых ранних находок этого рода в атлантическом секторе.
Присутствие крупных форм, обрастание раковин балянусами, мшанками, массовое развитие туррителл свидетельствуют о временами наступавшем благоприятном газовом режиме на дне и небольших глубинах, не превышавших первые десятки метров.
* * *
Авторы благодарны А.А. Березовскому за плодотворное обсуждение систематического состава моллюсков и помощь в их определении, Ж. Пако (J.-M. Pacaud) за помощь в определении гастропод, А.В. Гужову за помощь в поисках литературы и определении гастропод, В.Н. Беньямовскому за переданный им материал из скважин в районе г. Омска. Проведенные исследования были поддержаны грантом РФФИ № 17-05-00047.
Список литературы
Алексеев А.К. Палеогеновая фауна моллюсков Северного Приаралья. Ереван: Изд-во АН АрмССР, 1963. 230 с.
Амитров О.В. О распространении некоторых видов Aquilofusus в палеогене Закаспия // Палеонтол. журн. 1966. № 1. С. 28–37.
Амитров О.В. История гастропод палеогеновых морей запада Евразии. М: Наука, 1993. 208 с. (Тр. ПИН РАН. Т. 254).
Амитров О.В. Комплексы морских гастропод эоцена и олигоцена Западного Казахстана // Бюлл. МОИП. Отд. геол. 2005. Т. 80. Bып. 1. С. 37–55.
Ахметьев М.А., Александрова Г.Н., Беньямовский В.Н. и др. Новые данные по морскому палеогену Западно-Сибирской плиты. Ст. 1 // Стратигр. Геол. корелляция. 2004. Т. 12. № 2. С. 58–96.
Ахметьев М.А., Беньямовский В.Н. Палеоцен и эоцен российской части Западной Евразии // Стратигр. Геол. корелляция. 2006. Т. 14. № 1. С. 69–93.
Ахметьев М.А., Запорожец Н.И., Орешкина Т.В. и др. Сравнительный анализ разрезов и биоты морского палеогена Западной Сибири и Арктики // Стратигр. Геол. корелляция. 2010. Т. 18. № 6. С. 78–103.
Бакиева Л.Б. Палинология и стратиграфия палеогена зауральского и центрального литологофациальных районов Западной Сибири. Тюмень: ТюмГНГУ, 2005. 190 с.
Беньямовский В.Н. Палеогеновые меридиональные проливы Северной Евразии // Барабошкин Е.Ю., Найдин Д.П., Беньямовский В.Н. и др. Проливы Северного полушария в мелу и палеогене. М.: Изд-во Геол. ф-та МГУ, 2007. С. 80–118.
Васильева О.Н., Малышкина Т.П., Амон Э.О. Корреляция биозональных шкал по диноцистам, фораминиферам и ихтиофауне для палеогена Зауралья и Северного Тургая // Топорковские чтения. Вып. 7. Т. 1. Рудный: Рудненский индустриальный ин-т, 2006. С. 380–389.
Васильева О.Н. О возрасте тавдинской свиты в Тургайском прогибе // Ежегодник-2014. Тр. ИГГ УрО РАН. 2015. Вып. 162. С. 8–20.
Государственная геологическая карта Российской Федерации (третье поколение). Серия Уральская. Лист О-41 (Екатеринбург). – 1 : 1 000 000. М.: ФГУП “ВСЕГЕИ”, 2011.
Липман Р.Х., Буртман Б.С., Хохлова И.А. Стратиграфия и фауна палеогеновых отложений Западно-Сибирской низменности. Л.: Изд-во ВСЕГЕИ, 1960. 232 с. (Тр. ВСЕГЕИ. Нов. сер. Т. 28).
Мартынов В.А., Сигов А.П., Чирва С.А. Западно-Сибирская плита // Стратиграфия СССР. Палеогеновая система. М.: Недра, 1975. С. 315–329.
Мерклин Р.Л. Определитель двустворчатых моллюсков олигоцена юга СССР. М.: Наука, 1974. 172 с.
Миронова Л.В., Овечкин Н.К., Яркин В.И. Биостратиграфия палеогеновых отложений Тургайского прогиба и Северного Приаралья. Пластинчатожаберные моллюски // Матер. к биостратиграфии палеогеновых отложений Зап. Казахстана. Л.: Изд-во ВСЕГЕИ, 1962. 353 с. (Тр. ВСЕГЕИ. Нов. сер. Т. 82).
Овечкин Н.К. Отложения среднего палеогена Тургайской впадины и Северного Приаралья. М., 1954. 170 с.
Подобина В.М., Чернышов А.И. Стратиграфия и микрофаунистическая характеристика тавдинской свиты Западной Сибири. Томск: Изд-во Томск. гос. ун-та, 2012. 5 с.
Татишвили К.Г., Багдасарян К.Г., Казахашвили Ж.Р. Справочник по экологии морских брюхоногих. М.: Наука, 1968. 169 с.
Унифицированные региональные стратиграфические схемы неогеновых и палеогеновых отложений Западно-Сибирской равнины. Объяснительная записка. Новосибирск: СНИИГГиМС, 2001. 84 с.
Akhmet’ev M.A. Problems of Paleogene stratigraphy and paleogeography in the middle latitudes of Eurasia // Russ. Geol. and Geophysics. 2011. V. 52. P. 1075–1091.
Albrecht L., Valk W. Oligozäne Invertebraten von Sud-Limburg // Med. Geol. Stichting. Maastricht. Ser. C. 1943. Bd 4. № 13. 163 s.
Fretter V., Gracham A. The prosobranch molluscs of Britain and Denmark. Pt 6 // J. Moll. Stud. 1981. Suppl. 9. P. 285–363.
Iakovleva A.I., Rosseau D.-D., Cavagnetto C. Paleocene – Eocene dinoflagellate cysts and continental palynomorphs from borehole N 4 (Vasugan Basin, Central Western Siberia) // Palinology. 2000. V. 24. P. 187–200.
Marquet R., Lenaerts J., Laporte J. A systematic study of the Bivalvia (Mollusca) from the Glimmertingen Sand Member and from the Klimmen Member (Early Oligocene) in Belgium and the Netherlands // Palaeontos. 2012. V. 22. 151 p.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал