

Палеонтологический журнал, 2019, № 1, стр. 46-52
Мшанки и брахиоподы из фаменских отложений (верхний девон) центральной части Русской платформы
З. А. Толоконникова a, b, *, А. В. Пахневич c, **
a Кубанский государственный университет
Краснодар, Россия
b Казанский (Приволжский) федеральный университет
Казань, Россия
c Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: zalatoi@yandex.ru
** E-mail: alvpb@mail.ru
Поступила в редакцию 11.11.2017
Принята к публикации 23.01.2018
Аннотация
В отложениях задонского горизонта (нижний фамен, верхний девон) Липецкой и Орловской областей обнаружены два вида трепостоматных мшанок – Leptotrypa lipovkiensis sp. nov. и Anomalotoechus promiscuus Lavrentjeva, 1974, а также брахиоподы пяти отрядов, доминирующих в бентосной ассоциации этого горизонта. Мшанки характеризуются вариабельностью формы колонии и внутренних структур. Представители обеих групп беспозвоночных, очевидно, развивались в периодически меняющихся условиях внутренней части мелководного шельфа.
ВВЕДЕНИЕ
Указания на находки позднедевонских мшанок в центральной части Русской платформы встречаются в работах П.Н. Венюкова (1886), Б.П. Марковского и Д.В. Наливкина (1934). Первые полноценные описания фаменских мшанок рассматриваемой территории сделаны В.Д. Лаврентьевой (1970, 1974) во второй половине прошлого столетия. Согласно данным этого автора, обильные по численности фрагменты колоний мшанок были отобраны из 11 местонахождений в бассейнах рек Дон и Ока из отложений задонского, елецкого и лебедянского горизонтов нижнего-среднего фамена. Однако, эти фрагменты принадлежат всего к шести видам, относящимся к родам Anomalotoechus Duncan, 1939, Leptotrypa Ulrich, 1883, Schulgina Morozova, 1957 и Leptotrypella Vinassa de Regny, 1920, из семейства Atactotoechidae Duncan, 1939 отряда Trepostomata Ulrich, 1882. Такое низкое таксономическое разнообразие объяснялось неблагоприятными условиями среды обитания, связанными, по мнению Лаврентьевой, с “…мелководностью бассейна, … периодическим опреснением и сокращением связей с другими более открытыми бассейнами” (Лаврентьева, 1974, с. 45). В подобных условиях могли существовать только такие организмы, которые обладали высокой адаптивной способностью. Вероятно, такая способность и обусловила значительную внутри- и межколониальную изменчивость морфологических признаков (формы колонии, количества и размеров автозооециев и гетерозооециев) у определенной части фаменских мшанок, изученных Лаврентьевой.
Настоящее сообщение содержит результаты обработки нового каменного материала, отобранного из ранее не опробованных на мшанки местонахождений центральной части Русской платформы. Описан новый вид Leptotrypa lipovkiensis sp. nov., проведен анализ мшанковой ассоциации, рассмотрен состав брахиопод и другой фауны из задонского горизонта Центрального девонского поля.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
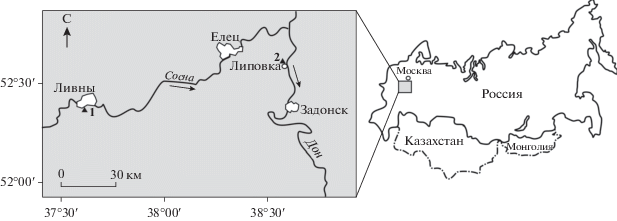
Изученные мшанки и брахиоподы происходят из двух местонахождений задонского горизонта (сборы А.В. Пахневича 1998 и 2016 гг.): первое – у с. Липовка на правом берегу р. Дон в районе родников, южнее заповедника “Галичья гора” (Липецкая область), второе – Горностаевский карьер (=карьер Известкового завода) на юго-западной окраине г. Ливны Орловской области в правом борту р. Сосна около д. Горностаевка (рис. 1). Мшанки из местонахождения у с. Липовка по внешнему облику изначально были отнесены Р.В. Горюновой к отряду Trepostomata без уточнения видовой принадлежности (Пахневич, 2005). В конце 2016 г. мшанковый материал из обоих местонахождений был большей частью расшлифован. В результате его обработки получено 40 ориентированных шлифов (восемь экземпляров), исследованных с помощью светового бинокулярного микроскопа Altami ПС0745, что позволило установить два вида мшанок, Leptotrypa lipovkiensis sp. nov. и Anomalotoechus promiscuus Lavrentjeva, 1974. Изучены также четыре целые однослойные колонии L. lipovkiensis sp. nov., инкрустирующие раковины брахиоподы Cystospirifer archiachi (Murchison, 1840), и два крупных фрагмента колонии A. рromiscuus, прямые ветви которой отходят от обрастающего основания. Описание нового таксона основывается на морфологических особенностях скелетных структур колоний и их замерах. Цифровые фотографии мшанок выполнены первым автором на микроскопе Altami ПС0745 в Кубанском государственном университете. Изученный материал хранится в Сибирском государственном индустриальном университете № 12/14.
ГЕОЛОГИЧЕСКАЯ И ФАУНИСТИЧЕКАЯ ХАРАКТЕРИСТИКА
Горностаевский карьер (=карьер Известкового завода) располагается на юго-западной окраине г. Ливны Орловской области в правом борту р. Сосна около д. Горностаевка (рис. 1). В нем обнажаются отложения верхнего девона от евлановского до задонского горизонта включительно (верхний фран-нижний фамен) (Обручева, Обручева, 1977; Молошников, 2001; Кирилишина, 2006). Отложения задонского горизонта в нижней части разреза представлены песками и песчаниками, сменяющимися в верхней части глинами и известняками. Общая мощность отложений достигает почти 10 м. Кроме мшанок, среди фоссилий обильны брахиоподы: Cystospirifer archiachi (Murchison, 1840) и C. zadonicus (Lyashenko, 1959) отряда Spiriferida, Orophomesorhynchus zadonicus (Nalivkin, 1934), O. cernosemicus (Nalivkin, 1934), O. huotinus (Verneuil, 1845), отряда Rhynchonellida, Chonopectus sp., Productella herminae Frech, 1891 отряда Productida, Athyris ex. gr. concentrica (Buch, 1834) отряда Athyridida. Интересны последние два вида, которые ранее были известны только из елецкого горизонта нижнего фамена Русской платформы (Родионова и др., 1995). Вероятно, их присутствие в отложениях задонского возраста указывает на терминальность задонских слоев на территории Горностаевского карьера и появление на этом уровне первых представителей елецкой фауны.
В литературе из задонского горизонта данного карьера описаны редкие конодонты (например, Polygnayhus acutatus Chalymbadzha, Shinkaryov, Gatovsky, 1991, Po. tichonovitchi Kuzmin et Melnikova, 1991, Icriodus iowaensis Youngquist et Peterson, 1947, Palmatolepis wolskae Ovnatanova, 1969), остатки позвоночных (например, Livnolepis zadonica (H. Obrucheva, 1983), Megapomus markovskyi Vorobyeva, 1977, Conchodus sp., Devononchus sp.) (Молошников, 2001; Кирилишина, 2006; Moloshnikov, 2008). Можно добавить, что А.В. Пахневичем обнаружены микроконхиды (ранее относимые к кольчатым червям рода Spirorbis), прямораковинные головоногие моллюски, мелкие сверления в раковинах брахиопод и ядра крупных нор. Крупные фрагменты колоний мшанки A. promiscuus были отобраны из светло-серых и темно-розовых известняков слоя № 9 согласно описанию С.В. Молошникова (2001). Координаты места сбора: 52°23′32′′ с.ш., 37°33′05′′ в.д.
Около с. Липовка (52°32′21′′ с.ш., 38°52′18′′ в.д.) на правом берегу р. Дона в районе родников, южнее заповедника “Галичья гора” (Липецкая область) (рис. 1) наблюдаются отдельные выходы коренных пород, представленных чередованием серых или желтоватых известняков и маломощных слоев серо-зеленых глин (Пахневич, 2005, 2007). По положению в разрезе и таксономическому составу брахиопод возраст отложений определен как задонский. Эти отложения содержат обильную фауну: мшанки, относящиеся к новому виду L. lipovkiensis sp. nov., табуляты, полихеты Serpula sp. и микроконхиды (Tentaculita), криноидеи, остракоды, единичные брюхоногие моллюски, двустворчатые моллюски Schizodus sp., Cimitaria sp., следы жизнедеятельности в виде нор. Однако в бентосной ассоциации доминируют брахиоподы, образующие “раковинные мостовые”. Среди них преобладают O. zadonicus (Nalivkin, 1934) (отряд Rhynchonellida), C. archiachi, C. zadonicus (отряд Spiriferida). Единично встречаются Cyrtospirifer tschernyschewi Khalfin, 1933 (отряд Spiriferida). Частыми являются Paromoeopygma koscharica (Nalivkin, 1934) (отряд Rhynchonellida), Chonopectus elcicus Nalivkin, 1934, Praewaagenoconcha oreliana (Moeller, 1871) (отряд Productida), Schuchertella sp. (отряд Strophomenida), а также неопределенные беззамковые брахиоподы с кальцитовой раковиной (Пахневич, 2007 с дополнениями и уточнениями; Savage, 2002a, b). Примечательно, что данное местонахождение располагается в стратотипическом районе задонского горизонта: стратотип последнего находится в недействующем карьере на южной окраине г. Задонск (Родионова и др., 1995).
СИСТЕМАТИЧЕСКОЕ ОПИСАНИЕ
КЛАСС STENОLAEMATA
ОТРЯД TREPOSTOMATA
ПОДОТРЯД AMPLEXOPORINA
СЕМЕЙСТВО ATACTOTOECHIDAE DUNCAN, 1939
Род Leptotrypa Ulrich, 1883
Leptotrypa lipovkiensis Tolokonnikova et Pakhnevich, sp. nov.
Табл. VI , фиг. 1, 2 (см. вклейку)
Название вида от с. Липовка.
Голотип – СибГИУ, № 12/14.2; Липецкая обл. России, местонахождение у с. Липовка; верхний девон, фаменский ярус, нижнефаменский подъярус, задонский горизонт.
Описание. Колония инкрустирующая однослойная, толщиной 0.35–1.50 мм. Эндо- и экзозоны слабо выражены. Толщина стенок в эндозоне 0.01 мм, к периферии она незначительно возрастает до 0.02 мм. В полости автозооециев могут быть развиты одна-три прямые и наклонные диафрагмы толщиной 0.01 мм. В экзозоне расстояние между ними 0.07–0.10 мм. В некоторых автозооециях диафрагмы не наблюдались. Четырех- и шестиугольные апертуры диаметром 0.08–0.11 мм, расположены беспорядочно или правильными диагональными рядами. В макулах диаметр апертур составляет 0.13–0.15 мм. На 2 мм в любом направлении приходится 8–9 апертур, в макулах – 6–7. Эксилязооеции короткие, редкие, диаметром 0.03–0.05 мм, располагаются без какой-либо закономерности. Акантостили мелкие, диаметром 0.01 мм. Они тяготеют к углам соединения стенок, располагаясь по 4–6 вокруг каждой апертуры.
Сравнение. От вида Leptotrypa donensis Lavr., 1970 (Лаврентьева, 1970, с. 48, табл. XIX , фиг. 1), описанного из франско-фаменских отложений Русской платформы, отличается большей вариацией толщины колонии (0.35–1.50 мм вместо 0.1–0.5 мм у L. donensis), мелкими апертурами (0.08–0.11 мм вместо 0.12–0.24 мм у L. donensis; в макулах 0.13–0.15 мм вместо 0.26–0.30 мм у L. donensis), меньшим количеством апертур на 2 мм (8–9 вместо 9–12 у L. donensis), мелкими эксилязооециями (0.03–0.05 мм вместо 0.04–0.15 мм у L. donensis).
Материал. Кроме голотипа, три паратипа № 12/14.3–14.5 и четыре колонии под № 12/14.1, 12/14.6–12/14.8 из того же местонахождения.
ОБСУЖДЕНИЕ
В двух местонахождениях центральной части Русской платформы (Орловская и Липецкая области) встречены мшанки отряда Trepostomata – Anomalotoechus promiscuus Lavr., 1974 и Leptotrypa lipovkiensis sp. nov. По положению в разрезе и комплексу сопутствующих окаменелостей, в первую очередь брахиопод, их возраст определен как задонский раннего фамена позднего девона. Оба вида мшанок представляют космополитные роды семейства Atactotoechidae Duncan, 1939 из отряда Trepostomata, наиболее характерного для девонских морей мира. Господство трепостомат в девоне можно объяснить фазой эволюционного расцвета мшанок данного отряда в этот период (Горюнова и др., 2004) и успешной их адаптацией к различным экологическим факторам. Согласно А. Эрнсту (Ernst, 2013), эволюционные тренды Bryozoa прослеживаются в закреплении морфологических изменений, направленных на освоение пространства, эффективность питания, защиту от хищников и размножение.
Освоение пространства изученными раннефаменскими мшанками центральной части Русской платформы реализовывалось через формирование колоний различных форм. Им свойственны однослойные тонкие инкрустирующие колонии (табл. VI , фиг. 2а), пряморастущие тонковетвистые с обрастающим разного размера основанием и массивные с точечным основанием (табл. VI , фиг. 3, 4). Колонии поселялись на раковинах брахиопод (табл. VI , фиг. 1, 3) и на водорослях.
Ветвистая прямая колония является довольно широко распространенной формой среди фанерозойских мшанок. Ее ветви, приподнятые над субстратом, имеют большую площадь для орошения и фильтрации воды, несущей пищевые частицы. Однако такая форма уязвима при сильных движениях воды, поэтому она тяготеет к относительно гидродинамически спокойным участкам дна (Cheetham, Thomsen, 1981). Инкрустирующие колонии имеют преимущество перед другими формами роста: они, обрастая любой тип субстрата (Bone, Wass, 1990), быстро осваивают большую площадь и сохраняют устойчивость к различным внешним воздействиям (McKinney, Jackson, 1989).
Большая часть колоний изученных раннефаменских мшанок, образована автозооидами с апертурами разного диаметра. Увеличенные апертуры сгруппированы иногда или часто в макулы (табл. VI, фиг. 2в). Развитие макул способствовало ускорению фильтрации воды, содержащей пищевые ресурсы: усиленный приток создавался лофофорами соседних автозооидов, собранными в группу (Ernst, 2013). Среди внутриколониальных структур у обоих изученных видов мшанок установлены редкие эксилязооеции и обильные акантостили (табл. VI , фиг. 2г, 5б). Эти типы структур характеризуются мелкими размерами. Основной функцией акантостилей, выступающих над поверхностью колонии наподобие шипов, очевидно, была защита (Астрова, 1971).
С учетом L. lipovkiensis sp. nov., ассоциация мшанок задонского горизонта центральной части Русской платформы представлена семью видами: Leptotrypa zadonskensis Lavr., L. donensis Lavr., Anomalotoechus promiscuus Lavr., Leptotrypella tenuimuralis Lavr., L. gracilis Lavr., Schulgina zushaensis Lavr. (Лаврентьева, 1970, 1974). Два вида обладают значительным интервалом распространения от задонского до лебедянского времени включительно (рис. 2). В целом, эта ассоциация характеризуется количественным обилием фрагментов колоний мшанок при их низком систематическом разнообразии. Мшанки обитали на мягком, биогенном и твердом субстратах: на раковинах брахиопод, водорослях, гальке, микроконхидах, криноидеях и конкрециях. Поверхность колоний мшанок незначительно разрушена, что может указывать на их частичный перенос и переотложение. Колонии приурочены к известнякам, чередующимся с глинистыми прослоями. Мшанки представлены преимущественно прямыми тонковетвистыми и инкрустирующими однослойными и многослойными колониями. В меньшем количестве в ассоциации присутствуют массивные и обрастающие колонии, переходящие в прямые ветвистые (Лаврентьева, 1970, 1974).
Рис. 2.
Стратиграфическое распространение мшанок в фаменских отложениях центральной части Русской платформы.

Примечательно, что у одного и того же вида встречаются разные формы колоний: у L. zadonskensis – инкрустирующие однослойные и многослойные; у A. promiscuus – инкрустирующие, массивные и инкрустирующие, переходящие в прямые ветвистые; у Sch. zushaensis – прямые ветвистые, инкрустирующие, массивные. Разнообразие колониальных форм можно рассматривать как результат адаптации к часто изменяющимся локальным условиям среды обитания (глубины, энергии воды, скорости седиментации). При всем многообразии форм колоний у одного и того же вида морфологические характеристики их внутренних структур остаются неизменными. Подобные мшанковые ассоциации возникают в условиях мелководного шельфа с глубинами 50–80 м (Amini et al., 2004) или внутреннего шельфа с глубинами 40–80 м (Hageman et al., 1997).
Согласно литолого-фациальному анализу и интерпретации данных по разным группам организмов, морской бассейн центральной части Русской платформы в течение позднего девона характеризовался неоднократным изменением глубины и площади (Тихомиров, 1995); периодической вариацией значений солености (от нормальной до близкой к ней) и глубины бассейна (Родионова и др., 1995); изменением связей с открытым океаном (Алексеев, Реймерс, 2005). Результаты анализа фаунистических комплексов раннего фамена из Горностаевского карьера и местонахождения вблизи с. Липовка хорошо укладываются в существовавшую картину.
Не только рассмотренные мшанки, но и находки криноидей, табулят (у с. Липовка), головоногих моллюсков (в Горностаевском карьере) свидетельствуют о существовании на изученной территории в начале фамена морского бассейна с нормальной соленостью. В подобных условиях жили и разнообразные многочисленные брахиоподы. В обоих местонахождениях они представлены четырьмя-пятью отрядами с доминированием Rhynchonellida. Возможно, представители этого отряда были более эвригалинными, чем остальные. В Горностаевском карьере они встречаются в песчаных и песчаниковых отложениях задонского горизонта вместе с остатками рыб и тетрапод. Накопление этих отложений, вероятно, происходило в условиях близкого расположения устья реки (О.А. Лебедев, устное сообщение, 2015 г.). Современные ринхонеллиды вида Hemithyris psittacea (Gmelin, 1790) – одни из немногих замковых брахиопод, выдерживающих некоторое опреснение морской воды (Зезина, 1999). При этом пониженная соленость влияет на темпы роста их раковин. Но в обоих местонахождениях присутствуют представители нескольких отрядов брахиопод. Это позволяет предположить, что соленость в обеих частях бассейна в задонское время раннего фамена была нормальной морской.
Значительное разнообразие эпибентосной фауны является признаком присутствия твердого субстрата. В местонахождении у с. Липовка он представлен отдельными гальками. Но основным твердым субстратом были многочисленные раковины брахиопод, что позволяло обрастателям и сверлящим организмам широко расселяться по морскому дну. В задонское время, как это было отмечено выше, раковины брахиопод формировали раковинные мостовые.
ВЫВОДЫ
1. В стратотипической местности отложений задонского горизонта нижнего фамена (верхний девон) установлен новый вид мшанки Leptotrypa lipovkiensis sp. nov. и найдены новые экземпляры эндемичного вида Anomalotoechus promiscuus Lavr. Раннефаменская ассоциация мшанок центральной части Русской платформы представлена только таксонами из отряда Trepostomata. Брахиоподы представлены пятью отрядами с доминированием Rhynchonellida.
2. Колонии задонских мшанок отличаются высокой фенотипической изменчивостью, которая проявляется в разнообразии форм колоний у одного и того же вида, в вариациях формы и размера апертур, а также параметров гетерозооециев.
3. Значительная внутри- и межколониальная изменчивость мшанок обусловлена их адаптацией к нестабильным условиям среды обитания в пределах внутренней части мелководного шельфа при достаточно устойчивой нормальной или близкой к нормальной солености, на что указывает таксономический состав мшанок, брахиопод и другой сопутствующей фауны.
* * *
Авторы благодарны С.В. Молошникову за консультации по характеристике Горностаевского карьера, Л.А. Висковой и А.В. Коромысловой за критический просмотр рукописи. З.А. Толоконникова признательна Paleontological Society International Research Program – Sepkoski Grant 2016 (проект 3000-16-62576-1) за поддержку исследования. Часть работы выполнена в рамках государственной программы повышения конкурентоспособности Казанского (Приволжского) федерального университета среди ведущих мировых научно-образовательных центров.
Список литературы
Алексеев А.С., Реймерс А.Н. Сравнительный анализ морской биоты бассейнов позднего девона и среднего-позднего карбона центральных районов Русской платформы // Палеобиология и детальная стратиграфия фанерозоя. К 100-летию со дня рождения академика В.В. Меннера. М.: РАЕН, 2005. С. 94–101.
Астрова Г.Г. К вопросу о морфологии и функциональном значении акантопор у трепостомат (Bryozoa) // Палеонтол. журн. 1971. № 3. С. 72–79.
Венюков П.Н. Фауна девонской системы Северо-Западной и Центральной России // Тр. СПб. об-ва естествоиспыт. 1886. Т. 17. С. 265–291.
Горюнова Р.В., Марков А.В., Наймарк Е.Б. Эволюция и биогеография палеозойских мшанок: результаты количественного анализа. М.: ГЕОС, 2004. 182 с.
Зезина О.Н. К размерной и возрастной характеристике современных брахиопод северных морей России // Адаптации животных и растений к условиям арктических морей (на уровне организма, популяции, экосистем). Матер. Междунар. семинара памяти акад. Е.М. Крепса (11–13 мая 1999 г., Мурманск). Апатиты, 1999. С. 68–70.
Зональная стратиграфия фанерозоя России / Науч. ред. Корень Т.Н. СПб.: ВСЕГЕИ, 2006. 255 с.
Кирилишина Е.М. Конодонты верхнефранских и нижнефаменских отложений Воронежской антеклизы. Автореф. дис. канд. геол.-мин. наук. М., 2006. 24 с.
Лаврентьева В.Д. Новые позднедевонские и раннекаменноугольные мшанки рода Leptotrypa Русской платформы // Новые виды палеозойских мшанок и кораллов. М.: Наука, 1970. C. 47–50.
Лаврентьева В.Д. Позднедевонские и раннетурнейские мшанки центральной части Русской платформы // Палеонтол. журн. 1974. № 2. С. 45–53.
Марковский Б.П., Наливкин Д.Б. Задонские и елецкие слои. М.–Л.–Новосибирск: Гос. научно-техническое горно-геолого-нефтяное изд-во, 1934. 55 с. (Тр. ГГГУ. Вып. 313).
Молошников С.В. Уникальное местонахождение позвоночных в верхнедевонских отложениях (Орловская область) // Изв. Высш. Учеб. завед. Геол. и разведка. 2001. № 3. С. 29–33.
Обручева О.П., Обручева Е.Д. Рыбы Центрального девонского поля // Очерки по филогении и систематике ископаемых рыб и бесчелюстных. М.: Наука, 1977. Вып. 2. С. 24–28.
Пахневич А.В. Палеонтологические исследования в заповеднике “Галичья гора” (предварительные результаты) //Фальцфейнивськи читання, Збірн. наук. праць. Херсон: Терра, 2005. Т. І. С. 70–73.
Пахневич А.В. Палеонтологические местонахождения на территории заповедника “Галичья гора” и сопредельных участках // Проблемы изучения и охраны объектов геологического наследия России: матер. раб. совещания Российской группы ProGEO (6–12 августа 2007 г., Миасс). М., 2007. С. 52–54.
Решение Межведомственного регионального стратиграфического совещания по среднему и верхнему палеозою Русской платформы с региональными стратиграфическими схемами. Ленинград, 1988. Девонская система. Л.: ВСЕГЕИ, 1990а. 58 с.
Решение Межведомственного регионального стратиграфического совещания по среднему и верхнему палеозою Русской платформы с региональными стратиграфическими схемами. Ленинград, 1988. Каменноугольная система. Л.: ВСЕГЕИ, 1990б. 41 с.
Родионова Г.Д., Умнова В.Т., Кононова Л.И. и др. Девон Воронежской антеклизы и Московской синеклизы. М., 1995. 265 с.
Тихомиров С.В. Этапы осадконакопления девона Русской платформы и общие вопросы развития и строения стратисферы. М.: Недра, 1995. 445 с.
Amini Z.Z., Adabi M.H., Burrett C.F., Quilty P.G. Bryozoan distribution and growth from associations as a tool in environmental interpretation, Tasmania, Australia // Sediment. Geol. 2004. V. 167. P. 1–15.
Bone Y., Wass R.E. Sub-Recent bryozoans-serpulid buildups in the Coorong Lagoon, South Australia // Aust. J. Earth Sci. 1990. V. 38. P. 207–214.
Cheetham A. X., Thomsen E. Functional morphology of arboresent animals: strength and design of cheilostome bryozoans skeletons // Paleobiology. 1981. V. 7. P. 355–383.
Ernst A. Diversity dynamics and evolutionary patterns of the Palaeozoic stenolaemate Bryozoa. Habilitationschrift (unpublished), Christian-Albrechts-Universität zu Kiel. 2013. 435 p.
Hageman, S.J., James, N.P., Bone Y. Cool water carbonate production from epizoic bryozoans on ephemeral substrates // Palaios. 1997. V. 15. P. 33–48.
Lebedev O.A., Lukševics E., Zakharenko G.V. Palaeozoogeographical connections of the Devonian vertebrate communities of the Baltica Province. Part II. Late Devonian // Palaeoworld. 2010. V. 19. P. 108–128.
McKinney, F.K., Jackson J.B.C. Bryozoan evolution. Boston: Unwin Human, 1989. 252 p.
Moloshnikov S.V. Devonian Antiarchs (Pisces, Antiarchi) from central and southern European Russia // Paleontol. J. 2008. V. 42. № 7. P. 691–773.
Savage N.M. Superfamily Rhynchotrematoidea Schuchert, 1913 // Treatise on Invertebrate Paleontology. Part H. Brachiopoda. V. 4: Rhynchonelliformea (part). Lawrence: Geol. Soc. Amer.; Univ. Kansas Press, 2002a. P. 1047–1091.
Savage N.M. Superfamily Camarotoechioidea Schuchert, 1929 // Treatise on Invertebrate Paleontology. Part H. Brachiopoda. V. 4: Rhynchonelliformea (part). Lawrence: Geol. Soc. Amer.; Univ. Kansas Press, 2002b. P. 1132–1164.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал