

Палеонтологический журнал, 2019, № 1, стр. 61-72
Палеогеновые черепахи Крыма
Е. А. Звонок a, *, И. Г. Данилов b, **
a Луганский национальный университет им. Тараса Шевченко
Луганск, Украина
b Зоологический институт РАН
Санкт-Петербург, Россия
* E-mail: evgenij-zvonok@yandex.ua
** E-mail: igordanilov72@gmail.com
Поступила в редакцию 29.11.2017
Принята к публикации 30.01.2018
Аннотация
Описаны ископаемые черепахи из пяти палеогеновых местонахождений Крыма: двух – датского возраста (Ак-Кая 2 и Сахарная Головка) и трех – бартонского возраста (Ак-Кая 1, Бахчисарай и Пролом). Наиболее изученной является фауна черепах местонахождения Ак-Кая 1, которая включает три таксона: морских черепах Argillochelys sp. (Cheloniidae) и Cosmochelys sp. (Dermochelyidae), а также неопределенную трехкоготную черепаху (Trionychidae indet.). Argillochelys sp. обнаруживает сходство с A. antiqua (König, 1825) из эоцена Европы. Новые данные расширяют представления о внутривидовой изменчивости и распространении черепах родов Argillochelys и Cosmochelys. Из других палеогеновых местонахождений Крыма описаны неопределенные хелонииды (Бахчисарай, Пролом и Сахарная Головка), дермохелииды (Бахчисарай и Пролом) и трионихиды (Ак-Кая 2 и Пролом).
Ископаемые черепахи Крыма долгое время были известны только из неогеновых и четвертичных отложений (Прендель, 1875; Рябинин, 1945; Хозацкий, 1946, 1947, 1948, 1953; Таращук, 1971), хотя Л.И. Хозацкий (1949, 1951) писал о находке в Крыму остатков “эоценовой черепахи (надо полагать, морской)”, которая, по-видимому, была утрачена. Позднее в литературе появились упоминания остатков палеогеновых черепах, собранных в Крыму Н.И. Удовиченко (Несов, 1987, 1992; Чхиквадзе, 1990; Averianov, 2002). Публикация этих, а также дополнительных материалов по палеогеновым черепахам Крыма (часть коллекции из местонахождения Ак-Кая 1 и экземпляры из местонахождения Сахарная Головка) началась сравнительно недавно (Звонок, 2011а, 2014; Звонок и др., 2013). В настоящей статье подробно описываются эти и другие материалы, хранящиеся в палеогерпетологической коллекции Зоологического ин-та РАН (ZIN PH) и происходящие из пяти местонахождений Крыма: Ак-Кая 1 и 2, Бахчисарай, Пролом и Сахарная Головка (рис. 1).
Рис. 1.
Местонахождения палеогеновых черепах на геологической карте Южного Крыма (на основе Геологической карты…, 2009).
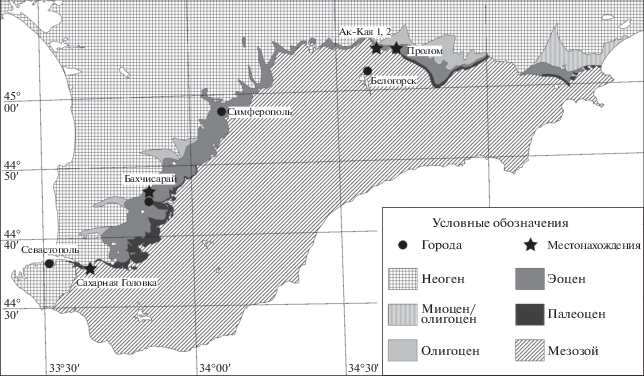
Мергели с фосфоритами местонахождения Ак-Кая 1 (=Белая Скала), Белогорский р-н (45°06′ N, 34°38′ E), относятся к бартонскому ярусу эоценового отдела, зона CP14b (верхняя часть NP17; В.А. Мусатов, личн. сообщ., данные по результатам изучения комплекса известкового нанопланктона). Материал по черепахам был собран Удовиченко в 2003 г. и Е.А. Звонком в 2010–2012 гг. Ранее из этого местонахождения упоминались Argillochelys sp., Cheloniidae indet. и Trionychidae indet., а также были описаны эпитекальные пластинки панциря Dermochelyidae indet. (Звонок, 2011а; Звонок и др., 2013). Из других позвоночных этого местонахождения в литературе описан богатый комплекс рыб и змеи-палеофииды трех видов (Братишко, Удовиченко, 2007; Братишко, 2011; Звонок, 2011а).
Примерно в 500 м западнее местонахождения Ак-Кая 1 находится карьер по добыче нуммулитового известняка – местонахождение Ак-Кая 2, где в известняках белокаменского региояруса (датский ярус палеоценового отдела) Удовиченко был найден фрагмент скульптированной пластинки панциря черепахи.
Мергели местонахождения Бахчисарай, Бахчисарайский р-н (44°46′ N, 33°50′ E), относятся к бартонскому ярусу эоценового отдела, зона CP14b (верхняя часть NP17) (В.А. Мусатов, личн. сообщ., данные по результатам изучения комплекса известкового нанопланктона). Материал по черепахам был собран Удовиченко в 2002 г. и А.Ф. Банниковым в 2009 г. Ранее из этого местонахождения был описан позвонок Trionychidae indet. (Звонок, 2011а; ошибочное определение, измененное в настоящей статье, см. Обсуждение). В числе других позвоночных данного местонахождения известны разнообразные рыбы и змеи-палеофииды (Снетков, Банников, 2010; Братишко, 2011; Малышкина и др., 2013). Из палеоценовых отложений окрестностей г. Бахчисарай также упоминались остатки морских черепах, однако место их хранения неизвестно (Averianov, 2002).
Мергели с фосфоритами местонахождения Пролом (=Белогорск), Белогорский р-н (45°05′ N, 34°39′ E), относятся к бартонскому ярусу (Братишко, Удовиченко, 2007). Материал по черепахам был собран Удовиченко в 1980–1990-х г.г. и Н.К. Гором в 1994 г. Ранее из местонахождения упоминались остатки морских черепах (см. Averianov, 2002). В числе других позвоночных из комплекса известны разнообразные рыбы и морские змеи-палеофииды (Averianov, 1997; Братишко, 2011).
Известняки белокаменского региояруса местонахождения Сахарная Головка, Севастопольский р-н (44°35′ N, 33°37′ E), относятся к датскому ярусу палеоценового отдела (Звонок, 2014). Фрагменты пластинок панциря черепахи были собраны Ю.О. Жуковым в 2013 г. и описаны Е.А. Звонком (2014).
В работе использованы следующие сокращения наименований учреждений: IRSNB, Королевский музей естественных наук Бельгии, Брюссель, Бельгия; NHM, Музей естественной истории, Лондон, Великобритания.
Анатомическая терминология дается по Ю. Гэффни (Gaffney, 1979) для черепных костей и по В.Б. Суханову (1964) – для костей посткрания.
Сравнение Argillochelys sp. из местонахождения Ак-Кая 1 осуществлялось со следующими представителями этого рода: A. antiqua (König, 1825): экз. IRSNB, № 1653, неполный череп, местонахождение Эрквелиннес, Бельгия, танет (Zangerl, 1971); экз. NHM, № 38952, неполный посткраний, местонахождение Шеппи, Великобритания, лондонские глины, ипр (голотип Chelone subcristata Owen, 1849; Owen, Bell, 1849; Lydekker, 1889); экз. ZIN PH, №№ 2-4/36, 8/36, неполный скелет; местонахождение Вышгород, Украина, лютет или бартон (Zvonok, Danilov, 2017); A. cuneiceps (Owen, 1849): экз. NHM, № 41636 (голотип), неполный череп, местонахождение Шеппи, Великобритания, лондонские глины, ипр (Owen, Bell, 1849; Lydekker, 1889). Cравнение описанных в статье материалов по Argillochelys sp. также проводилось с другими эоценовыми таксонами хелониид: “Argillochelys” africana Tong et Hirayama, 2008; Eochelone brabantica Dollo, 1903; Osonachelus decorata de Lapparent de Broin et al., 2014 и Puppigerus camperi (Gray, 1831) (Owen, Bell, 1849; Lydekker, 1889; Casier, 1968; Moody, 1974; Tong, Hirayama, 2008; de Lapparent de Broin et al., 2014).
ОПИСАНИЕ МАТЕРИАЛА ПО CHELONIIDAE
Argillochelys sp. (местонахождение Ак-Кая 1) представлен следующим материалом: семь предлобных костей, экз. ZIN PH, № 29/153 и шесть экз. без номеров (далее – б/н) в коллекции ZIN PH, № 153; четыре лобные кости, экз. ZIN PH, № 30/153 и б/н; шесть теменных костей, экз. ZIN PH, №№ 31-32/153 и б/н; две скуловые кости, экз. ZIN PH, № 33/153 и б/н; чешуйчатая кость, экз. ZIN PH, № 34/153; три заглазничные кости, экз. ZIN PH, № 35/153 и б/н; три предчелюстные кости, экз. ZIN PH, №№ 36-37/153 и б/н; пять верхнечелюстных костей, экз. ZIN PH, № 38/153 и б/н; крыловидная кость, экз. ZIN PH, № 39/153; квадратная кость, экз. ZIN PH, № 40/153; 23 зубные кости, их симфизы и фрагменты, экз. ZIN PH, №№ 41-42/153 и б/н; надугловая кость, экз. ZIN PH, № 43/153; 38 невральных пластинок, экз. ZIN PH, №№ 44-47/153 и б/н; супрапигальная пластинка, экз. ZIN PH, № 48/153; пигальная пластинка, экз. ZIN PH, № 49/153; восемь костальных пластинок, экз. ZIN PH, № 50/153 и б/н; 96 периферальных пластинок, экз. ZIN PH, №№ 51-54/153 и б/н; семь фрагментов гио- или гипопластронов, экз. ZIN PH, № 55/153 и б/н; ксифипластрон, экз. ZIN PH, № 56/153; VI шейный позвонок, экз. ZIN PH, № 57/153; две лопатки, экз. ZIN PH, № 58/153 и б/н; 14 плечевых костей и их фрагментов, экз. ZIN PH, №№ 59-60/153; проксимальный фрагмент бедренной кости, экз. ZIN PH, № 61/153.
Предлобные кости (рис. 2, а). Почти полностью сохранились две предлобных кости, включая экз. ZIN PH, № 29/153. Остальные экземпляры неполные, два из них более широкие и массивные. Кости вытянуты спереди назад. Постеромедиальный край длинный и ориентирован диагонально по отношению к сагиттальной плоскости, что свидетельствует о том, что лобные кости вместе глубоко вклинивались между предлобными (см. ниже). Предлобная кость формировала антеродорсальный край глазницы и дорсальную часть края наружного носового отверстия. Предлобных щитков одна пара.
Рис. 2.
Кости черепа Argillohelys sp. из местонахождения Ак-Кая 1 (Крым; бартон, средний эоцен): а – экз. ZIN PH, № 29/153, левая предлобная кость сверху; б – экз. ZIN PH, № 30/153, правая лобная кость сверху; в – экз. ZIN PH, № 31/153, правая теменная кость сверху; г – экз. ZIN PH, № 32/153, правая теменная кость сверху; д, е – экз. ZIN PH, № 33/153, правая скуловая кость изнутри (д) и сбоку (е); ж – экз. ZIN PH, № 34/153, левая чешуйчатая кость сбоку; з – экз. ZIN PH, № 35/153, правая заглазничная кость сбоку; и – экз. ZIN PH, № 36/153, правая предчелюстная кость спереди; к, л – экз. ZIN PH 37/153, задняя часть правой предчелюстной кости снизу (к) и медиально (л); м, н – экз. ZIN PH, № 38/153, правая верхнечелюстная кость сверху (м) и медиально (н); о – экз. ZIN PH, № 39/153, левая крыловидная кость сверху; п – экз. ZIN PH, № 40/153, правая квадратная кость медиально; р, с – экз. ZIN PH, № 42/153, задняя часть левой зубной кости медиально (р) и латерально (с); т–ц – экз. ZIN PH, № 41/153, симфиз зубных костей сверху (т), снизу (у), сзади (ф), сбоку (х) и спереди (ц); ч, ш – экз. ZIN PH, № 43/153, левая надугловая кость сверху (ч) и сбоку (ш). Обозначения: aam – area articularis mandibularis; cc – canalis cavernosus, cci – canalis caroticus internus; ddm – углубление m. depressor mandibulae; df – смещенный фрагмент зубной кости; F – лобный щиток; fai – foramen alveolare inferius; fas – foramen alveolare superius; fdm – foramen dentofaciale majus; fna – foramen nervi auriculotemporalis; fsm – foramen supramaxillare; FP – лобно-теменной щиток; IP – межтеменной щиток; ica – incisura columellae auris; lar – лабиальный гребень; lir – лингвальный гребень; MA – верхнечелюстной щиток; P – теменной щиток; PF – предлобный щиток; PO – заглазничный щиток; pp – поверхность слома предлобного отростка; rms – гребень на медиальной поверхности скуловой кости; scm – sulcus cartilaginis meckelii; SO – надглазничный щиток; sw – вздутие на альвеолярной поверхности предчелюстной и верхнечелюстной костей; syr – симфизный гребень; TE – височный щиток. Длина масштабной линейки – 1 см.

Лобные кости (рис. 2, б). Почти полностью сохранились две лобных кости, включая экз. ZIN PH, № 30/153; у еще двух отломаны латеральные части. Лобные кости четырехугольной формы, глубоко вклинивались между предлобными. Они формировали небольшую часть дорсального края глазницы, как у всех известных эоценовых хелониид, за исключением “Argillochelys” africana. Лобный щиток заостренный спереди.
Теменные кости (рис. 2, в, г). Все теменные кости повреждены. Борозда между лобным и теменным щитками сохранилась у двух экз. б/н (не изображена) и проходит примерно по шву между лобной и теменной пластинками. Задний край лобно-теменного щитка виден у экз. ZIN PH, №№ 31-32/153 и двух экз. б/н. У экз. ZIN PH, № 31/153 и двух экз. б/н лобно-теменной щиток почти равномерно сужается в заднем направлении, как у Argillochelys antiqua (экз. ZIN PH, № 8/36) и Eochelone brabantica Dollo, 1903, и в отличие от A. antiqua (экз. IRSNB, № 1653), голотипа A. cuneiceps (экз. NHM, № 41636), а также других эоценовых хелониид (рис. 2, в). У экз. ZIN PH, № 32/153 в задней части лобно-теменного щитка имеется вырезка для вклинивающегося вперед межтеменного щитка (рис. 2, г). Теменные кости формировали дорсальный край задней височной вырезки.
Скуловые кости (рис. 2, д, е). Сохранились задние части экз. ZIN PH, № 33/153 и одного экз. б/н. На латеральной поверхности видна борозда между заглазничным и верхнечелюстным щитками. На медиальной поверхности экз. ZIN PH, № 33/153 гребень, проходящий вдоль края глазницы, слабо выражен и не достигает дорсального края кости; у второго экземпляра он высокий, острый и достигает дорсального края кости (не изображен). Кость формировала постеровентральный край глазницы и передний край нижней височной вырезки.
Чешуйчатые кости (рис. 2, ж). Сохранилась вентральная часть чешуйчатой кости экз. ZIN PH, № 34/153. Углубление для крепления m. depressor mandibulae расположено близко к заднему краю кости, узкое и глубокое, как у “Argillochelys” africana, голотипа Argillochelys cuneiceps (экз. NHM, № 41636) и A. antiqua (экз. ZIN PH, № 8/36) и отличается от широкого углубления у Eochelone brabantica и Puppigerus camperi. У Argillochelys antiqua (экз. IRSNB, № 1653) это углубление широкое, как у Puppigerus camperi, но более глубокое. Чешуйчатая кость формировала боковой край задней височной вырезки и задний край cavum tympani.
Заглазничные кости (рис. 2, з). У заглазничных костей экз. ZIN PH, № 35/153 и двух экз. б/н повреждены задние и нижние части. Кости вытянуты в длину, широкие. У экз. ZIN PH, № 35/153 надглазничный щиток разделен на два, которые сзади граничат с крупным височным щитком. Заглазничная кость формировала постеродорсальный край глазницы.
Предчелюстные кости (рис. 2, и–л) представлены лабиальным гребнем (экз. ZIN PH, № 36/153) и небной частью (экз. ZIN PH, № 37/153), возможно, одной особи, а также еще одним лабиальным гребнем (экз. б/н). На всех фрагментах видны питательные отверстия. Лабиальный гребень высокий, острый, не образовывал вырезку или выраженный крюк. Небная часть кости экз. ZIN PH, № 37/153 субтреугольной формы при виде снизу и сильно утолщается в заднем направлении при виде с медиальной стороны, образуя мощное вздутие в месте контакта с верхнечелюстной костью (сходно с A. antiqua, экз. ZIN PH, № 2/36; Zvonok, Danilov, 2017).
Верхнечелюстные кости (рис. 2, м, н) представлены экз. ZIN PH, № 38/153 и одним экз. б/н с отломанными предлобными отростками; остальные экземпляры представляют собой задние части костей. Лабиальный гребень высокий, острый, увеличивается в высоту и толщину в заднем направлении. Небный отросток сужается в заднем направлении. В антеромедиальной части небного отростка имеется мощное вздутие, продолжающееся на предчелюстной кости (см. выше). На дорсальной поверхности, у постеромедиального края предлобного отростка видно крупное foramen alveolare superius. Foramen supramaxillare находится у заднего края кости.
Крыловидные кости (рис. 2, о) представлены средней частью кости экз. ZIN PH, № 39/153. Canalis caroticus internus и canalis cavernosus не сливаются в единый канал. Область контакта между крыловидными костями на вентральной поверхности повреждена, поэтому наличие сагиттального гребня установить невозможно.
Квадратные кости (рис. 2, п) представлены экз. ZIN PH, № 40/153, у которого повреждена или обломана часть, прилегающая к condylus mandibularis, а также дорсальная и медиальная части кости. Incisura columellae auris открыта сзади.
Зубные кости (рис. 2, р–ц). Альвеолярная поверхность в области симфиза сохранилась у экз. ZIN PH, № 41/153 и еще у семи экземпляров б/н. Она субтреугольной формы при виде сверху, как у A. antiqua (экз. ZIN PH, №№ 3-4/36). Присутствуют одновременно лабиальные, симфизный и лингвальные гребни, которые варьируют по высоте у разных экземпляров. Sulcus cartilaginis meckelii очень глубокий в районе симфиза, как у A. antiqua (экз. ZIN PH, №№ 3-4/36). Foramen dentofaciale majus и foramen alveolare inferius видны на изолированной ветви зубной кости (экз. ZIN PH, № 42/153) и расположены далеко позади симфиза (рис. 2, р, с).
Надугловая кость (рис. 2, ч, ш) представлена дорсальной частью (экз. ZIN PH, № 43/153), которая формировала латеральный край fossa meckelii и латеральную часть area articularis mandibularis. На латеральной поверхности кости виден foramen nervi auriculotemporalis.
Невральные пластинки (рис. 3, а–г). Среди невральных пластинок имеются шестиугольные с короткими передне-боковыми сторонами, слабо суженные сзади (напр., экз. ZIN PH, № 44/153), занимавшие более переднее положение в невральном ряду (из них некоторые пересечены интерцентральной бороздой); такие же по форме, но сильнее суженные сзади (напр., экз. ZIN PH, № 45/153), занимавшие среднее положение в ряду; а также пятиугольные (напр., экз. ZIN PH, № 46/153), располагавшиеся в конце ряда. Три пластинки, включая экз. ZIN PH, № 47/153, имеют киль в передней части, подобно A. antiqua (экз. NHM, № 38952), еще восемь пластинок выпуклые сверху, а остальные плоские.
Рис. 3.
Кости посткрания Argillochelys sp. из местонахождения Ак-Кая 1 (Крым; бартон, средний эоцен): а–г – невральные пластинки сверху (а–в) и спереди (г): а – экз. ZIN PH, № 44/153; б – экз. ZIN PH, № 45/153; в – экз. ZIN PH, № 46/153; г – ZIN PH 47/153; д – экз. ZIN PH, № 48/153, II супрапигальная пластинка сверху; е – экз. ZIN PH, № 49/153, пигальная пластинка сверху; ж – экз. ZIN PH, № 50/153, костальная пластинка сверху; з, и – экз. ZIN PH, № 51/153, II правая периферальная пластинка сверху (з) и спереди (и); к, л – экз. ZIN PH, № 52/153, III правая периферальная пластинка снизу (к) и сзади (л); м, н – экз. ZIN PH, № 53/153, средняя периферальная пластинка сверху (м) и спереди (н); о, п – экз. ZIN PH, № 54/153, задняя периферальная пластинка сверху (о) и спереди (п); р, с – экз. ZIN PH, № 55/153, фрагмент гио- или гипопластрона снизу (р) и спереди или сзади (с); т – экз. ZIN PH, № 56/153, правый ксифипластрон снизу; у–ц – экз. ZIN PH, № 57/153, VI шейный позвонок снизу (у), сбоку (ф), сзади (х) и спереди (ц); ч – экз. ZIN PH, № 58/153, фрагмент левой лопатки спереди; ш – экз. ZIN PH, № 59/153, левая плечевая кость снизу; щ–я – экз. ZIN PH, № 60/153, медиальная часть правой плечевой кости снизу (щ), сверху (ы), сзади (э), проксимально (ю) и спереди (я); аа, бб – экз. ZIN PH, № 61/153, медиальная часть левой бедренной кости снизу (аа) и сзади (бб). Обозначения: aas – передняя сочленовная поверхность тела позвонка; AS – анальный щиток; cf – головка бедренной кости; ch – головка плечевой кости; CS – центральный щиток; df – углубление у основания малого трохантера; dlt – углубление для mm. latissimus dorsi et teres major; FS – бедренный щиток; gn – шейка гленоида; if – межтрохантерная яма; k – киль; MS – маргинальный щиток; nh – вырезка для гипопластрона; np – вырезка на периферальной пластинке; pas – задняя сочленовная поверхность тела позвонка; pm – медиальный отросток; pl – латеральный отросток; pp – презигапофиз; PS – плевральный щиток; tma – большой трохантер; tmi – малый трохантер; tp – поперечный отросток; vk – вентральный киль. Длина масштабной линейки – 1 см.

II супрапигальная пластинка (рис. 3, д; экз. ZIN PH, № 48/153) вытянута спереди назад, сужается кзади (к контакту с пигальной пластинкой), ее латеральные края свободные (ограничивают косто-периферальные фонтанели). В задней части пластинки видна борозда между последним центральным и одним из маргинальных щитков.
Пигальная пластинка (рис. 3, е; экз. ZIN PH, № 49/153) имеет небольшое углубление спереди для вхождения заднего края II супрапигальной пластинки. Задняя часть пластинки повреждена, но латеральные части заднего края значительно толще, чем его медиальная часть, что говорит о наличии вырезки сзади.
Костальные пластинки (рис. 3, ж) в различной степени повреждены. Две из них, включая экз. ZIN PH, № 50/153, с сохранившейся латеральной частью, имели свободные края, что свидетельствует о наличии косто-периферальных фонтанелей.
Периферальные пластинки (рис. 3, з–п). II периферальная пластинка (экз. ZIN PH, № 51/153) без ямки для свободного ребра на медиальной поверхности, каплевидное сечение при виде спереди. Крупная III периферальная пластинка (напр., экз. ZIN PH, № 52/153) сильно увеличивается в ширину в заднем направлении и имеет ямку для свободного ребра примерно посредине медиальной поверхности. Она имеет вырезку латерального (свободного) края, служащую, по-видимому, для удержания самки самцом когтем (у Carolinochelys wilsoni Hay, 1923 эта вырезка была на II периферальной пластинке; Weems, Sanders, 2014). Средние периферальные пластинки (напр., экз. ZIN PH, № 53/153) имеют близкий к прямому угол между дорсальной и вентральной сторонами и ямки для свободных ребер. Задние периферальные пластинки (напр., экз. ZIN PH, № 54/153) имеют острый угол между дорсальной и вентральной сторонами, могут иметь (более передние) или не иметь (более задние) ямки для свободных ребер. Одна из задних периферальных пластинок (?XI; экз. ZIN PH, № 54/153) в средней части латерального края имеет вырезку, подобную той, которая есть на крупной III периферальной пластинке (экз. ZIN PH, № 52/153).
Гио- или гипопластроны (рис. 3, р, с) представлены только фрагментами, которые не позволяют точно определить, какой именно элемент представлен. Два из них, включая экз. ZIN PH, № 55/153, имеют мощные парасагиттальные гребни.
Ксифипластрон (рис. 3, т; экз. ZIN PH, № 56/153) обломан спереди и сзади. Его передняя часть уже задней. Постеромедиальная часть пластинки имеет не менее одного заостренного отростка, направленного медиально. В передней части видна борозда между феморальным и анальным щитками.
VI шейный позвонок (рис. 3, у–ц; экз. ZIN PH, № 57/153) с обломанными верхними частями невральных дуг, постзигапофизами и частично поперечными отростками, вентральный киль стерт. Передняя сочленовная поверхность тела позвонка вогнутая, дорсовентрально приплюснутая, овальной формы. Задняя сочленовная поверхность тела позвонка выпуклая, такой же формы, как передняя, но менее широкая. Презигапофизы ориентированы почти параллельно оси тела позвонка.
Лопатка (рис. 3, ч; экз. ZIN PH, № 58/153) с обломанными скапулярным и акромиальными отростками, что не позволяет измерить угол между ними. Шейка гленоида короткая.
Плечевые кости (рис. 3, ш–я). Проксимальная часть плечевой кости сохранилась только у двух экземпляров (экз. ZIN PH, №№ 59-60/153). Processus lateralis расположен близко к головке плечевой кости, округленный. Processus medialis короткий, что, по-видимому, связано с молодым возрастом черепах (Zug et al., 1986). Головка сферическая. Углубление для mm. latissimus dorsi et teres major крупное, расположено ближе к заднему краю. Дистальные части костей сильно повреждены и детали их строения не видны.
Бедренная кость (рис. 3, аа, бб; экз. ZIN PH, № 61/153) представлена сильно поврежденной проксимальной частью. Гребня между трохантерами нет. У основания малого трохантера на дорсальной стороне имеется ямка.
Cheloniidae indet. (местонахождение Бахчисарай) представлены следующим материалом: VIII шейный позвонок, экз. ZIN PH, № 1/151; ?IV периферальная пластинка, экз. ZIN PH, № 2/151; фрагменты периферальных пластинок, экз. ZIN PH, №№ 4-5/151; эпипластрон, экз. ZIN PH, № 3/151.
VIII шейный позвонок (рис. 4, а–в) представлен невральной дугой с постзигапофизами. Задняя часть невральной дуги значительно выше передней. На ее антеродорсальной поверхности сохранилась сочленовная поверхность для прикрепления к загривковой пластинке. Постзигапофизы маленькие, расположены близко друг к другу.
Рис. 4.
Остатки черепах из разных палеогеновых местонахождений Крыма: а–е – Cheloniidae indet. (местонахождение Бахчисарай; бартон, средний эоцен): а–в – экз. ZIN PH, № 1/151, VIII шейный позвонок сбоку (а), сзади (б) и сверху (в); г – экз. ZIN PH, № 3/151, левый эпипластрон снизу; д, е – экз. ZIN PH, № 2/151, ?IV периферальная пластинка сверху или снизу (д), спереди или сзади (е); ж–и – Cheloniidae indet. (местонахождение Пролом; бартон, средний эоцен): ж ‒ экз. ZIN PH, № 1/18, костальная пластинка сверху; з, и – экз. ZIN PH, № 3/18, периферальная пластинка сверху или снизу (з), спереди или сзади (и); к–у – Cosmochelys sp. (местонахождение Ак-Кая 1; бартон, средний эоцен): к–н – экз. ZIN PH, № 62/153, VIII шейный позвонок сбоку (к), спереди (л), сзади (м) и сверху (н); о – экз. ZIN PH, № 62/153, поперечный отросток сверху или снизу; п – экз. ZIN PH, № 10/153, две эпитекальных пластинки карапакса в сочленении сверху; р, с – экз. ZIN PH, № 64/153, краевая эпитекальная пластинка сверху или снизу (р) и спереди или сзади (с); т – экз. ZIN PH, № 65/153, маргинальная эпитекальная пластинка спереди или сзади; у – экз. ZIN PH, № 63/153, эпитекальная пластинка карапакса с гребнем спереди или сзади; ф – экз. ZIN PH, № 4/151, эпитекальная пластинка Dermochelyidae indet. (местонахождение Бахчисарай; бартон, средний эоцен) сверху; х – экз. ZIN PH, № 5/18, эпитекальная пластинка Dermochelyidae indet. (местонахождение Пролом; бартон, средний эоцен) сверху; ц–ш – Trionychidae indet. (местонахождение Ак-Кая 1; бартон, средний эоцен): ц – экз. ZIN PH, № 67/153, невральная пластинка сверху; ч, ш – экз. ZIN PH, № 68/153, фрагмент костальной пластинки сверху (ч), спереди или сзади (ш); щ – экз. ZIN PH, № 7/18, фрагмент пластинки панциря Trionychidae indet. (местонахождение Пролом; бартон, средний эоцен) сверху или снизу; ы – экз. ZIN PH 1/241, фрагмент пластинки панциря Trionychidae indet. (Ак-Кая 2; даний, нижний палеоцен) сверху или снизу; э, ю – экз. ZIN PH, № 7/151, фрагмент левой зубной кости Testudines indet. (местонахождение Бахчисарай; бартон, средний эоцен) сбоку (э) и медиально (ю); я – экз. ZIN PH, № 1/240, фрагмент пластинки панциря Cheloniidae indet. (местонахождение Сахарная Головка; даний, нижний палеоцен) сверху или снизу. Обозначения: aas – передняя сочленовная поверхность тела позвонка; asn – сочленовная поверхность для нухальной пластинки; CS – центральный щиток; lar – лабиальный гребень; lir – лингвальный гребень; poz – постзигапофиз; PS – плевральный щиток; prz – презигапофиз; rls – гребнеподобная структура; sa – поверхность для сочленения с угловой костью; scm – sulcus cartilaginis meckelii; sr – борозда заднего края рамфотеки; tp – поперечный отросток; tu – бугорки. Длина масштабной линейки – 1 см.

IV (?) периферальная пластинка ювенильной особи (рис. 4, д, е) целая, узкая медиолатерально и высокая дорсовентрально, передний и задний края примерно одинаковой высоты.
Эпипластрон (рис. 4, г) обломан сзади, вытянут спереди назад. Значительная часть постеромедиального края, судя по его шероховатой поверхности и небольшому перегибу позади этого участка, контактировала с энтопластроном.
Cheloniidae indet. (местонахождение Пролом) представлены следующим материалом: две средние костальные пластинки, экз. ZIN PH, № 1-2/18; две задние периферальные пластинки, экз. ZIN PH, № 3-4/18.
Костальные пластинки (рис. 4, ж) без выраженной скульптуры и с обломанными дистальными частями. Периферальные пластинки (рис. 4, з–и) с острым углом между дорсальной и вентральной поверхностями.
Cheloniidae indet. (местонахождение Сахарная Головка) представлены следующим материалом: экз. ZIN PH, № 1-5/240, пять фрагментов пластинок, по-видимому, одной особи (Звонок, 2014).
Край пластинки панциря (рис. 4, я; экз. ZIN PH, № 1/240) имеет форму ломаной линии из трех отрезков с тупыми углами между собой; два из них свободные, один имеет контактную поверхность.
ОПИСАНИЕ МАТЕРИАЛА ПО DERMOCHELYIDAE
Cosmochelys sp. (местонахождение Ак-Кая 1) представлен следующим материалом: два фрагмента одного VIII шейного позвонка, экз. ZIN PH, № 62/153; 172 эпитекальные пластинки, экз. ZIN PH, №№ 1-13, 63-65/153 и б/н.
VIII шейный позвонок (рис. 4, к–о). Сохранилась большая часть позвонка, у которого повреждены нижняя и задняя части тела и отломаны поперечные отростки, но большая часть одного из них сохранилась отдельно. Передняя сочленовная поверхность тела позвонка вогнутая и почти круглая. Задняя часть невральной дуги значительно выше передней, область для сочленения с загривковой пластинкой стерта. Презигапофизы не выходят латерально за уровень тела позвонка. Постзигапофизы маленькие, расположены близко друг к другу.
Эпитекальные пластинки (рис. 4, п–у). Серия эпитекальных пластинок из Ак-Каи 1 ранее описана как Dermochelyidae indet. (Звонок и др., 2013). Большинство пластинок имеют форму многогранников округлых очертаний. Пластинки крупных особей более толстые, не имеют рельефа наружной поверхности или имеют рельеф в виде слабовыраженных бугорков. Пластинки ювенильных особей более тонкие, имеют рельеф в виде таких же, но сильнее выраженных бугорков. Краевой элемент крупной особи с углом между дорсальной и вентральной частями около 80° (Звонок и др., 2013).
Одна из ранее не описанных эпитекальных пластинок ювенильной особи (экз. ZIN PH, № 63/153) имеет высокий заостренный в сечении гребень, как у Cosmochelys dolloi (Andrews, 1919), Psephophorus polygonus von Meyer, 1847 и Dermochelys coriacea (Wood et al., 1996; Delfino et al., 2013). Края пластинки обломаны. Она менее толстая, чем у других палеогеновых дермохелиид, но ее небольшая толщина соответствует другим эпитекальным пластинкам ювенильных особей из Ак-Каи 1.
Пять ранее не описанных краевых элементов, включая экз. ZIN PH, №№ 64-65/153, принадлежавших ювенильным особям, имеют сильно варьирующий угол между карапаксальной и пластральной поверхностями, от 35° до 55°, что может объясняться различным положением пластинок на краю эпитекальной мозаики.
Dermochelyidae indet. (местонахождение Бахчисарай) представлены тремя эпитекальными пластинками, экз. ZIN PH, №№ 4-6/151. Они имеют округлые очертания (рис. 4, ф) и рельеф в виде слабо выраженных бугорков, не отличаясь от некоторых Dermochelyidae из Ак-Каи 1.
Dermochelyidae indet. (местонахождение Пролом) представлены двумя эпитекальными пластинками, экз. ZIN PH, №№ 5-6/18. Эти пластинки (рис. 4, х) имеют округлые очертания и рельеф в виде слабо выраженных бугорков, не отличаясь от некоторых Dermochelyidae из Ак-Каи 1 и Бахчисарая.
ОПИСАНИЕ МАТЕРИАЛА ПО TRIONYCHIDAE
Trionychidae indet. (местонахождение Ак-Кая 1) представлены следующим материалом: невральная пластинка, экз. ZIN PH, № 67/153; фрагмент костальной пластинки, экз. ZIN PH, № 68/153.
Невральная пластинка (рис. 4, ц) шестиугольная с короткими передне- или заднебоковыми сторонами, вытянута спереди назад. На наружной поверхности имеется скульптура в виде мелких ямок, характерная для Trionychidae. Удлиненность пластинки свидетельствует о ее происхождении из передней или средней части неврального ряда.
Фрагмент костальной пластинки (рис. 4, ч, ш) с такой же скульптурой наружной поверхности не имеет особенностей, позволяющих определить ее положение.
Trionychidae indet. (местонахождение Пролом) представлены двумя фрагментами неопределенных пластинок, экз. ZIN PH, №№ 7-8/18, которые имеют скульптуру (рис. 4, щ) в виде ямок и извивающихся борозд и гребней, характерную для Trionychidae.
Trionychidae indet. (местонахождение Ак-Кая 2) представлены фрагментом пластинки панциря, экз. ZIN PH 1/241 (рис. 4, ы) со скульптурой в виде заостренных бугорков и гребней, характерной для Trionychidae.
ОПИСАНИЕ МАТЕРИАЛА ПО TESTUDINES INDET.
Testudines indet. (местонахождение Бахчисарай) представлены средней частью левой ветви зубной кости, экз. ZIN PH, № 7/151 (рис. 4, э, ю). На латеральной поверхности кости в передней части фрагмента сохранились многочисленные крупные питательные отверстия, позади которых располагается широкая диагональная борозда, соответствующая заднему краю рамфотеки. Sulcus cartilaginis meckelii расположен высоко. Вентральнее него кость толстая и закругленная снизу. Постеромедиальная часть фрагмента образует шовную поверхность с угловой костью. Также видны основания обломанных лабиального и лингвального гребней. В задней части лабиальный гребень переходит в небольшой вырост – вероятно, переднюю часть processus coronoideus или представлявший собой дорсальный отросток. Лингвальный гребень вплотную примыкал к верхнему краю sulcus cartilaginis meckelii. В задней части эти гребни не смыкаются между собой, между ними видно основание еще одной гребнеподобной структуры, возможно, лежавшей в основании венечной кости.
ОБСУЖДЕНИЕ
Бартонские черепахи Крыма известны по многочисленным материалам, большинство из которых происходит из местонахождения Ак-Кая 1. Остатки хелониид из Ак-Каи 1 отнесены к роду Argillochelys на основании того, что все зубные кости хелониид с сохранившейся альвеолярной поверхностью (восемь экземпляров) имеют одновременно лабиальные, симфизный и лингвальные гребни, и часть невральных пластинок (три экземпляра) имеют киль – признаки рода Argillochelys (Zvonok, Danilov, 2017). Остальные кости хелониид из Ак-Каи 1, скорее всего, также принадлежат к роду Argillochelys, так как отличаются от других эоценовых хелониид. Так, лобные кости отличаются от “Argillochelys” africana, конфигурация щитков на теменных костях – от “Argillochelys” africana, Osonachelus decorata и Puppigerus camperi, чешуйчатая кость – от Eochelone brabantica и Puppigerus camperi, предчелюстная и верхнечелюстные кости – от Puppigerus camperi (Casier, 1968; Moody, 1974; Tong, Hirayama, 2008; de Lapparent de Broin et al., 2014).
Субтреугольные вытянутые симфизы зубных костей и вытянутый в заднем направлении лобно-теменной щиток являются признаками A. antiqua, однако разобщенность анатомических элементов и отсутствие в материалах передней части крыловидных костей и, соответственно, отсутствие данных о форме наружного крыловидного отростка, не позволяют уверенно определить видовую принадлежность аргиллохелисов из местонахождения Ак-Кая 1, которые определяются как Argillochelys sp.
Хелонииды Бахчисарая и Пролома известны по более фрагментарным материалам, поэтому их родовую принадлежность уточнить невозможно. VIII шейный позвонок хелониида из Бахчисарая (экз. ZIN PH, № 1/151) ранее был ошибочно отнесен к трионихидам (Звонок, 2011б). В этой работе он определен как хелониид, так как имеет сочленение нухальной пластинки с VIII шейным позвонком – синапоморфию Сhelonioidea, отсутствующую у трионихид, а также не имеет существенных отличий от хелониид Caretta caretta (L., 1758), Chelonia mydas L., 1758 и Procolpochelys grandaeva (Leidy, 1851) (Zangerl, Turnbull, 1955; Joyce, 2007).
Вторым бартонским таксоном черепах Крыма является дермохелиид, лучше всего изученный по материалам из Ак-Каи 1. Большая часть данных о нем была опубликована ранее (Звонок и др., 2013). Новый материал свидетельствует о том, что эти черепахи имели эпитекальные пластинки с гребнями. Наличие данных гребней исключает принадлежность к Natemys peruvianus Wood et al., 1996 и “Psephophorus” rupeliensis Van Beneden, 1883, сходство с которыми отмечалось ранее (Wood et al., 1996; Звонок и др., 2013). Само наличие эпитекальных пластинок исключает принадлежность к Eosphargis spp., а отсутствие вздутий пластинок и наличие губчатого слоя кости – к Arabemys crassiscutata Tong et al., 1999 (Wood et al., 1996: ch. 8; Звонок и др., 2013). Дермохелиид из Ак-Каи 1 не относится к Egyptemys, так как гребень на эпитекальной пластинке ZIN PH, № 64/153 в сечении заостренный, а не полукруглый (Wood et al., 1996). Заостренная форма гребня наблюдается у Cosmochelys dolloi – единственного вида дермохелиид, описанного из среднего эоцена (Wood et al., 1996). Наличие этого признака, а также среднеэоценовый возраст, позволяют отнести дермохелиида из Ак-Каи 1 к роду Cosmochelys. Вместе с тем, скульптура наружной поверхности эпитекальных пластинок зрелых особей дермохелиида из Ак-Каи 1 в целом менее выражена, чем у Cosmochelys dolloi, и напоминает более поздних Egyptemys eocaenus и Psephophorus calvertensis Palmer, 1909 (Tong et al., 1999), поэтому мы определяем дермохелиида из Ак-Каи 1 как Cosmochelys sp.
VIII шейный позвонок из Ак-Каи 1 (экз. ZIN PH, № 62/153) можно отнести к дермохелиидам, так как примерно одинаковые высота и ширина сочленовных поверхностей тел задних шейных позвонков описаны у Dermochelyoidae; у всех хелониид сочленовная поверхность задних шейных позвонков уплощенная (Hirayama, Chitoku, 1996: ch. 62; Hirayama, 1998: ch. 47).
Конфигурация гребней зубной кости неопределенной черепахи из Бахчисарая (экз. ZIN PH, № 7/151) не характерна для Argillochelys, у которого лабиальный и лингвальный гребни сходятся в задней части зубной кости, и лингвальный гребень расположен значительно дорсальнее sulcus cartilaginis meckelii (см. выше). От современного Dermochelys coriacea отличается наличием выроста в постеродорсальной части кости (Gaffney, 1979: фиг. 204). Если этот вырост является передней частью processus coronoideus, то кость может принадлежать Trionychidae, для которых характерен высокий pr. coronoideus (Gaffney, 1979).
Обилие в местонахождении Ак-Кая 1 остатков только одного вида (Argillochelys sp.), преобладание костей ювенильных особей и принадлежность двух периферальных пластинок (экз. ZIN PH, №№ 52/15 и 53/153) взрослым особям (вероятно, зрелым самкам), возможно, указывают на то, что в районе Ак-Каи 1 в бартонском веке был пляж, на котором эти черепахи откладывали яйца. Подобное местонахождение с массовыми материалами только по одному виду хелониид – Tasbacka aldabergeni Nessov, 1987 – известно из палеоцена Казахстана (танетский ярус, Джилга 1; Несов, 1987).
Трионихиды могут вести литоральный образ жизни, но при этом всегда связаны с устьями рек. Они могут перемещаться вдоль морского побережья на значительные расстояния и переплывать небольшие проливы, подобно тому, как Trionyx triunguis заселяют побережье островов Эгейского моря (Taskavak, Can Akcinar, 2009). В связи с этим, присутствие трионихид указывает на наличие поблизости местонахождений Ак-Кая 1 и Пролом (восточная часть северных предгорий Крыма) устья реки.
Датские черепахи Крыма пока представлены только неопределенными хелониидами и трионихидами. Присутствие трионихид в Ак-Кае 2 также, вероятнее всего, указывает на близость суши.
* * *
Авторы благодарят Н.И. Удовиченко, А.Ф. Банникова, Н.К. Гора и Ю.О. Жукова за передачу собранных ими материалов по палеогеновым черепахам Крыма в Зоологический ин-т РАН, В.А. Мусатова за определение геологического возраста местонахождений, Р. Хираяму за предоставление фотографий Argillochelys spp. из коллекций NHM и IRSNB и анонимного рецензента за замечания, которые позволили улучшить статью.
Работа выполнена в рамках темы Зоологического ин-та РАН № AAAA-A17-117030310017-8.
Список литературы
Братишко А.В. Отолиты и зубы костистых рыб палеогена Украины. Автореф. дис. канд. геол. наук. К.: ИГН НАН Украины, 2011. 25 с.
Братишко А.В., Удовиченко Н.И. Ихтиофауна верхней части эоценовых отложений в районе Белогорска (Крым) // Палеонтологічні дослідження України: історія, сучасний стан та перспективи. Київ, 2007. С. 238–244.
Геологическая карта Горного и Предгорного Крыма. Сост. В.В. Юдин. Симферополь: Союзкарта, 2009.
Звонок Е.А. Лютетские рептилии местонахождения Ак-Кая (Крым, Украина): предварительные данные // Пробл. стратиграфии и корреляции фанерозойских отложений Украины: Матер. XXXIII сессии Палеонтол. об-ва НАН Украины. Киев, 2011а. С. 72–73.
Звонок Є. Нові дані про місцезнаходження й таксономічне розмаїття еоценових крокодилів і черепах України // Палеонтол. збірн. Львів, 2011б. № 43. С. 107–120.
Звонок Е.А. О пластинке панциря морской черепахи из отложений датского яруса окрестностей Севастополя (Крым, Украина) // Еволюція органічного світу та етапи геологічного розвитку Землі: Матер. XXXV сесії Палеонтол. тов-ва НАН України. К., 2014. С. 83–84.
Звонок Е.А., Данилов И.Г., Сыромятникова Е.В. Первая достоверная находка кожистых черепах (Dermochelyidae) на территории Северной Евразии (средний эоцен Украины) // Палеонтол. журн. 2013. № 2. С. 70–73.
Малышкина Т.П., Гонсалес-Барба Х., Банников А.Ф. Находки зубов эласмобранхий в бартоне Северного Кавказа (Россия) и Крыма (Украина) // Палеонтол. журн. 2013. № 1. С. 76–80.
Несов Л.А. О морских черепахах палеогена Южного Казахстана и филогенетических соотношениях Toxochelyidae и Cheloniidae // Палеонтол. журн. 1987. № 4. С. 76–87.
Несов Л.А. Обзор местонахождений остатков птиц мезозоя и палеогена СССР и описание новых находок // Русс. орнитол. журн. 1992. Т. 1. № 1. С. 7–50.
Прендель Р. Сарматские образования Севастополя и его окрестностей // Зап. Новоросс. об-ва естествоиспыт. 1875. Т. 3. Вып. 1. С. 1–30.
Рябинин А.Н. Черепаха из мэотиса Крыма // Ежегодн. Всеросс. палеонтол. об-ва. 1945. Т. 12. С. 126–132.
Снетков П.Б., Банников А.Ф. Позвонки морских змей из эоцена Крыма // Палеонтол. журн. 2010. № 6. С. 97–100.
Суханов В.Б. Подкласс Testudinata. Тестудинаты // Основы палеонтологии. Земноводные, пресмыкающиеся и птицы. М.: Наука, 1964. С. 354−438.
Таращук В.И. Черепахи неогеновых и антропогеновых отложений Украины. Сообщ. I. Семейство большеголовых черепах (Platysternidae) // Вестн. зоол. 1971. № 2. С. 56–62.
Хозацкий Л.И. Остатки пресноводной черепахи Emys orbicularis L. из четвертичных отложений Крыма // Бюлл. Комисс. по изуч. четверт. пер. 1946. № 8. С. 78–81.
Хозацкий Л.И. Наземная черепаха из мэотических отложений Крыма // Докл. АН СССР. 1947. Т. 58. № 9. С. 2059–2062.
Хозацкий Л.И. Об остатках черепах из сармата Крыма // Бюлл. МОИП. Отд. геол. 1948. Т. 23. Вып. 3. С. 17–22.
Хозацкий Л.И. История фауны черепах СССР в свете палеогеографии // Тр. 2-го Всесоюз. географ. съезда. М.: Изд-во геол. лит-ры, 1949. Т. С. С. 221–230.
Хозацкий Л.И. Палеонтологическое и стратиграфическое значение ископаемых черепах // Вопросы палеонтол. Т. 1. Л., 1951. С. 20–31.
Хозацкий Л.И. О мэотических черепахах Керченского полуострова // Ежегодн. Всесоюз. палеонтол. об-ва. 1953. Т. 14. С. 237–256.
Чхиквадзе В.М. Палеогеновые черепахи СССР. Тбилиси: Мецниереба, 1990. 96 с.
Averianov A.O. Paleogene sea snakes from the eastern part of Tethys // Russ. J. Herpetol. 1997. № 4. P. 128–142.
Averianov A.O. Review of Mesozoic and Cenozoic sea turtles from the former USSR // Russ. J. Herpetol. 2002. V. 9. № 2. P. 137–154.
Casier E. Le squelette céphalique de Eochelone brabantica L. Dollo, du Bruxellien (Lutétien inférieur) de Belgique, et sa comparaison avec celui de Chelone mydas Linné // Bull. Inst. Roy. Sci. Natur. Belg. 1968. T. 44. P. 1–22.
Delfino M., Scheyer T.M., Chesi F. et al. Gross morphology and microstructure of type locality ossicles of Psephophorus polygonus Meyer, 1847 (Testudines, Dermochelyidae) // Geol. Mag. 2013. V. 150. № 5. P. 767–782.
Gaffney E.S. Comparative cranial morphology of recent and fossil turtles // Bull. Amer. Mus. Natur. Hist. 1979. V. 164. P. 65–376.
Hirayama R. Oldest known sea turtle // Nature. 1998. V. 392. P. 705–708.
Hirayama R., Chitoku T. Family Dermochelyidae (Superfamily Chelonioidea) from the Upper Cretaceous of North Japan // Trans. Proc. Palaeontol. Soc. Japan. N. Ser. 1996. № 184. P. 597–622.
Joyce W. Phylogenetic relationships of Mesozoic turtles // Bull. Peabody Mus. Natur. Hist. 2007. V. 48. № 1. P. 3–102.
Lapparent de Broin de F., Murelaga X., Farrés F., Altimiras J. An exceptional cheloniid turtle, Osonachelus decorata nov. gen., nov. sp., from the Eocene (Bartonian) of Catalonia (Spain) // Geobios. 2014. V. 47. № 3. P. 111–132.
Lydekker R. Catalogue of the Fossil Reptilia and Amphibia in the British Museum (Natural History). Part III. The Order Chelonia. L.: Brit. Mus. (Natur. Hist.), 1889. 239 p.
Moody R.T.J. The taxonomy and morphology of Puppigerus camperi (Gray), an Eocene sea turtle from northern Europe // Bull. Brit. Mus. (Natur. Hist.) Geol. 1974. V. 25. P. 153–186.
Owen R., Bell A. Monograph of the Fossil Reptilia of the London Clay. Part I. Chelonia. L.: Palaeontogr. Soc., 1849. 76 p.
Taskavak E., Can Akcinar S. Marine records of the Nile soft–shelled turtle, Trionyx triunguis // Marine Biodiversity Records. 2009. № 2. doi 10.1017/S1755267208000092
Tong H., Buffetaut E., Thomas H. et al. A new dermochelyid turtle from the Late Paleocene – Early Eocene of Saudi Arabia // C.R. Acad. Sci. Earth Planet. Sci. 1999. V. 329. P. 913–919.
Tong H., Hirayama R. A new species of Argillochelys (Testudines: Cryptodira: Cheloniidae) from the Ouled Abdoun phosphate basin, Morocco // Bull. Soc. Géol. Fr. 2008. V. 179. № 6. P. 623–630.
Weems R.E., Sanders A.E. Oligocene pancheloniid sea turtles from the vicinity of Charleston, South Carolina, U.S.A. // J. Vertebr. Paleontol. 2014. V. 34. P. 80–99.
Wood R.C., Johnson-Gove J., Gaffney E.S., Maley K.F. Evolution and phylogeny of leatherback turtles (Dermochelyidae), with descriptions of new fossil taxa // Chelonian Conserv. Biol. 1996. № 2. P. 266–286.
Zangerl R. Two toxochelyid sea turtles from the Landenian sands of Erquelinnes (Hainaut) of Belgium // Mém. Inst. Roy. Sci. Natur. Belg. 1971. T. 169. P. 1–32.
Zangerl R., Turnbull W.D. Procolpochelys grandaeva (Leidy), an early carettine sea turtle // Fieldiana Zool. 1955. № 37. P. 345–382.
Zug G.R., Wynn A.H., Ruckdeschel C. Age determination of loggerhead sea turtles, Caretta caretta, by incremental growth marks in the skeleton // Smithson. Contrib. Zool. 1986. № 127. P. 1–34.
Zvonok E.A., Danilov I.G. A revision of fossil turtles from the Kiev clays (Ukraine, middle Eocene) with comments on the history of the collection of fossil vertebrates of A.S. Rogovich // Proc. Zool. Inst. RAS. 2017. V. 321. № 4. P. 485–516.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал