

Палеонтологический журнал, 2019, № 2, стр. 72-83
Ящерицы cемейств Dorsetisauridae и Xenosauridae (Anguimorpha) из апта-альба Монголии
В. Р. Алифанов *
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: valifan@paleo.ru
Поступила в редакцию 28.11.2017
После доработки 15.03.2018
Принята к публикации 12.04.2018
Аннотация
В работе описаны два новых вида ящериц в составе инфраотряда Anguimorpha из апт–альбских отложений Монголии. Один из них – Paradorsetisaurus postumus gen. et sp. nov. – самый поздний в семействе Dorsetisauridae, а другой – Xenostius futilus gen. et sp. nov. – самый ранний в семействе Xenosauridae s. str. Статья включает также комментарии, поясняющие значение находок из Монголии для реконструкции становления разнообразия ящериц в Центральной Азии до начала позднего мела.
ВВЕДЕНИЕ
Местонахождение ископаемых наземных позвоночных Хобур (Ховур, Гучин-Ус) расположено в Центральной Монголии (Гучинская депрессия, Увэрхангайский аймак). Оно сложено апт–альбскими по возрасту песчано-глинистыми отложениями озерного и озерно-речного генезиса. Коллекция остатков тетрапод (млекопитающие, динозавры, птерозавры, крокодилы, черепахи и ящерицы) из Хобура в фондах Палеонтологического ин-та им. А.А. Борисяка РАН (ПИН РАН) составлена, главным образом, по итогам камерального разбора промытой в поле костеносной породы. Работы проводили разные отряды Совместной Советско-Монгольской палеонтологической экспедиции, в основном, в период с 1969 по 1972 гг. Наибольшее число костных остатков в сборах принадлежит ящерицам.
Ящерицы Хобура представлены многочисленными (сотни образцов) изолированными элементами скелета, черепа и нижней челюсти. Такие материалы трудно идентифицировать. Однако опыт первых и во многом предварительных определений (Alifanov, 1993, 2000; Алифанов, 2000) позволил установить большое количество семейств (рис. 1). Данный факт ставит ассоциацию ящериц Хобура в положение самой богатой в мире для интервала от середины юры до конца раннего мела. Она также имеет ключевое значение для выяснения деталей становления разнообразия обсуждаемой группы в ранний период ее истории.
Рис. 1.
Семейственный состав (по данным разных работ) и фаунистическая структура ассоциации ящериц из местонахождения Хобур (Монголия, Увэрхангайский аймак; нижний мел, верхний апт–нижний альб, хухтыкский горизонт).
Часть ящериц Хобура изучена подробнее. В составе Gekkonidae s.l. (Gekkota) выявлена древнейшая форма – Hoburogekko suchanovi (Алифанов, 1989; Daza et al., 2012). Семейство Hodzhakuliidae Alifanov, 1993 (Scincomorpha) представлено описанным ранее из позднего альба Узбекистана Hodzhakulia magna Nessov, 1985 и новыми видами: Carnoscincus eublepharus, Bagaluus primigenius и Platynotoides altidentatus (Алифанов, 2016). Охарактеризованы также данные о семействе Temujiniidae (Temujiniidae gen. indet.: Алифанов, 2018), составляющем часть микроотряда Iguanomorpha (Iguania).
Настоящая работа продолжает изучение разнообразия ящериц Хобура. На этот раз в центр внимания поставлены находки, относящиеся к инфраотряду Anguimorpha, а именно – к семействам Dorsetisauridae и Xenosauridae. Систематической части предшествует краткий очерк по развитию представлений на их классификацию, филогенез и географическое распространение. В заключительном разделе статьи обсуждаются некоторые подробности формирования разнообразия ящериц на территории Центральной Азии в течение юры и раннего мела.
Работа поддержана грантом РФФИ, № 16-05-00408.
ПРОБЛЕМЫ ИЗУЧЕНИЯ СЕМЕЙСТВ DORSETISAURIDAE И XENOSAURIDAE
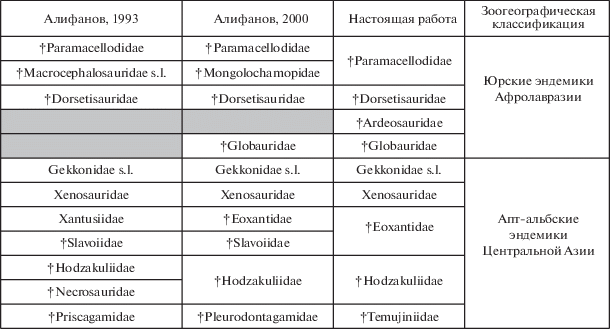
Семейство Dorsetisauridae выделил Р. Хоффштеттер (Hoffstetter, 1967) для установленного им же по фрагментам черепа и нижней челюсти Dorsetisaurus purbeckensis из формации Пурбек (титон–неоком) в Англии. У данного вида отмечаются остеодермальная скульптура на костях крыши черепа в виде щитков среднего размера, положение теменного отверстия в поле теменной кости, а также немногочисленные зубы с высокой и ланцетовидной по форме апикальной частью (рис. 2, а–в). Позднее последнее семейство оказалось зафиксировано в поздней юре США (Prothero, Estes, 1980) и Португалии (Seiffert, 1973; Broschinski, 2000), а также в раннем мелу Монголии (Alifanov, 1993, 2000; Алифанов, 2000).
Рис. 2.
Ящерицы семейства Dorsetisauridae (Anguimorpha): а–в – Dorsetisaurus purbeckensis Hoffstetter, 1967 (Hoffstetter, 1967, фиг. 10 A, 11 С): а, б – правая верхнечелюстная кость в сочленении с предлобной и слезной лабиально (а) и вентрально (б); экз. BMNH, № R8248, в – реконструкция левой ветви нижней челюсти лабиально; Англия, титон–нижний неоком; г, д – Kuwajimalla kagaensis Evans et Manabe, 2008 (по: Evans, Manabe, 2008, текст-фиг. 4, B, 5, А), правая зубная кость (г) лабиально (экз. SBEI, № 1538) и нижнечелюстные зубы (д) лингвально (экз. SBEI 557); Япония, баррем–апт; е – Chometokadmon fitzingeri Costa, 1864; голотип MPN, № 539 (по: Evans et al., 2006, фиг. 2, B), реконструкция черепа дорсально; Италия, нижний мел; ж – Paradorsetisaurus postumus gen. et sp. nov.; голотип ПИН, № 3334/517, правая верхнечелюстная кость лабиально; Монголия, Увэрхангайский аймак, местонахождение Хобур; нижний мел, верхний апт–нижний альб, хухтыкский горизонт. Длина масштабной линейки – 5 мм для а–в, е и ж, 1 мм для г и д.
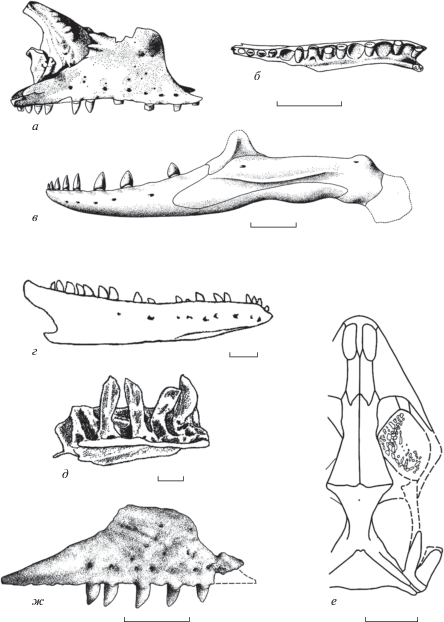
Первоначально Dorsetisauridae рассматривались в составе Anguoidea (Hoffstetter, 1967) или в промежуточном положении между современными Xenosauridae s.l. и Anguidae (Estes, 1983). Типовой род ископаемой группы относился и к базальным ангвиморфам (Evans et al., 2006), и к базальным платинотам (Conrad, 2008). Однако его cвязь с Anguimorpha пока строго не доказана из-за фрагментарности данных. В этом случае все гипотезы родства носят очень предварительный характер. Именно в подобном ключе нами (Алифанов, 2000, 2012) допускалась близкая филогенетическая связь дорсетизаврид и Xenosauridae s.l.
Новый свет на Dorsetisauridae проливает такая ящерица, как Kuwajimalla kagaensis Evans et Manabe, 2008 из раннего мела Японии (Исикава). Этот вид, основанный на находке фрагментов черепа и нижней челюсти с зубами, вершина которых имеет ланцетовидную форму, классифицирован в первоописании (Evans, Manabe, 2008) в кладистическом таксоне Borioteiioidea Nydam et al., 2007. Последний представляет собой сумму выделенных нами ранее (Алифанов, 2000, 2012) позднемеловых микроотрядов Polyglyphanodontia (Agamognatha) и Macrocephalosauria (Chamaeleomorpha). Отметим, что их несомненных раннемеловых представителей в Центральной Азии нет. Кроме этого, у K. kagaensis не отмечается свойственного Polyglyphanodontia и Macrocephalosauria субплевродонтного подтипа прикрепления нижнечелюстных зубов, что ставит под сомнение первую реконструкцию филогенетических связей данного вида. Сомнительна также приведенная в первоописании вида интерпретация пильчато-зубчатого строения кромок верхушечной части зубов в качестве многовершинности. С другой стороны, ланцетовидная форма зубов, эуплевродонтный (промежуточный между суб- и гиперплевродонтией) тип прикрепления их на нижней челюсти и наличие выемок под зубные основания (рис. 2, г, д) допускают родство K. kagaensis с Dorsetisauridae.
При обсуждении проблем, связанных с Dorsetisauridae, интересны данные по Chometokadmon fitzingeri Costa, 1864 (рис. 2, е) из альба Центральной Италии. Переизучение позволило связать эту форму с Anguimorpha (Evans et al., 2006). У Ch. fitzingeri лобные и теменная кости совпадают по строению с Dorsetisaurus purbeckensis Hoffstetter, 1967. Кроме этого, отмечается наличие свободных остеодерм в области головы, как у Xenosauridae s.l. (Bever et al., 2005). Тем не менее, С. Эванс и др. (Evans et al., 2006) отметили генерализованность Ch. fitzingeri, сблизив его, кроме D. purbeckensis, с Parviraptor Evans, 1994 из поздней юры и неокома США и Англии. Интересно, что в отношении последней формы, известной по изолированным костям скелета, существуют и другие представления, например, о ее возможной связи со змеями (Caldwell et al., 2015). С нашей точки зрения, парвираптор может быть классифицирован в таксоне Mosasauria, представители которого, как и змеи, нередко обладают заостренными зубами, а также зигосфен-зигантральным сочленением позвонков.
Судя по деталям строения черепа, нижней челюсти и зубной системы, эмбрионы в яйцах с необычной для ящериц твердой скорлуповой оболочкой из раннего мела Таиланда также принадлежат представителю Dorsetisauridae, а не Platynota (для раннего мела варанообразные ящерицы достоверно не известны), как было предположено в первоописании (Fernandez et al., 2015). Наше определение учитывает представленную в последней работе реконструкцию черепа, на которой отмечено вхождение верхнечелюстных костей в состав суборбитальных окон, чего у вараноидных ящериц не бывает, а также охарактеризовано строение лобных костей (они широкие, парные и скульптурированы), выявлено наличие удлиненного рострального отростка венечных костей, глубокой вырезки заднего края в целом коротких зубных костей и уплощенной верхушки зубов с боковыми кромками.
Напротив, отнесение Changetisaurus estesi Nessov, 1992 из средней юры Кыргызстана к дорсетизавридам (Федоров, Несов, 1992) спорно из-за развития крупного ангулярного отростка зубных костей при наличии прямоугольных остеодерм. Эти признаки одновременно свойственны ископаемым Paramacellodidae (Cordyloidea, Scincomorpha), например, Sharovisaurus karatauensis Hecht et Hecht, 1984 из поздней юры Казахстана и Parmeosaurus scutatus Gao et Hou, 2000 из позднего мела Монголии.
В отличие от Dorsetisauridae, Xenosauridae s.l., представленные современными родами Xenosaurus (Мексика) и Shinisaurus (Китай), – несомненные Anguimorpha. Исторически их сближали с Anguidae в составе микроотряда Diploglossa, сестринского по отношению Platynota (Camp, 1923; McDowell, Bogert, 1954). Кладистический анализ определяет ксенозаврид к базальным Anguimorpha (Gauthier et al., 2012) или Platynota (Сonrad, 2008). Платинотная гипотеза относится к Shinisaurus и близким к этому роду ископаемых формам. С нашей точки зрения, она противоречит ключевым признакам строения сравниваемых таксонов.
Заметим, что для Shinisaurus, как и для всех Diploglossa (кроме Anniella: Алифанов, 2000, 2012), характерна диплоглоссопалатинарность (отсутствие связи латеральных отростков наружных крыловидных и небных костей и их положение вентральнее субдентальных отростков верхнечелюстных костей, которые составляют обрамление в край суборбитальных окон). Этот тип может быть противопоставлен другому распространенному у ящериц платинотопалатинарному типу (верхнечелюстные кости изолированы от края суборбитальных окон медиальным контактом небных и наружных крыловидных костей). По нашим данным, оба состояния несводимы друг к другу, поскольку составляют части разных ветвей дивергентного морфо-типологического ряда. Вывести одно такое состояние из другого непосредственно без учета промежуточных стадий невозможно (Алифанов, 2012, 2016).
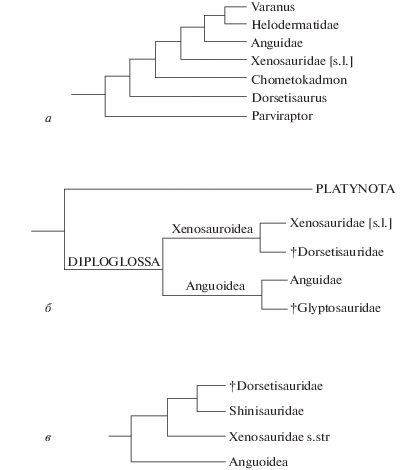
В свете указанных проблем и данных следует обратить внимание на строение зубов у Shinisaurus и Xenosaurus. У первой формы они несут неявно выраженные симметрично расположенные плечи центральной вершинки (Conrad, 2004, фиг. 17, A, C ), а у второй – ростральное плечо, придающее верхушке зуба асимметричное строение. Если Xenosauridae s.l. родственны Dorsetisauridae, то возникает обстоятельство, которое в ряде вышеназванных работ не учитывалось, в том числе и нами (рис. 3, а, б). Оно состоит в разной степени родства двух современных форм по отношению к ископаемому семейству. В частности, в обозначенной группе сравнения по строению зубов обособленно выглядит Xenosaurus. Данное обстоятельство позволяет рассматривать две современные ксенозавроидные формы в отдельных семействах (рис. 3, в), что не раз допускалось ранее (Bever et al., 2005; Conrad, 2008).
Рис. 3.
Реконструкция филогенетических связей семейств Dorsetisauridae и Xenosauridae (Anguimorpha) по разным авторам: а – C. Эванс и др. (Evans et al., 2006), б – В.Р. Алифанову (2012), в – настоящая работа.
С Xenosauridae s.l. иногда синонимизируют позднемеловых Carusiidae Borsuk-Białynicka, 1987 или их типовой род (Gao, Norell, 1998, 2000; Conrad, 2008). Такому определению, однако, противоречит строение зубных костей. У представителей карусиид обнаруживается сцинкоморфный тип их строения (супраангулярный отросток зубных костей зачаточный или отсутствует, ангулярный отросток широкий в основании и часто удлиненный, а короноидный отросток налегает на латеральную поверхность лабиального и/или дорсального отростков венечных костей). Сцинкоморфность отмечалась и в первоописании обсуждаемой группы (Borsuk-Białynicka, 1985, 1987). В современных работах допускается ее базальное положение в составе Scincoidea (Gauthier et al., 2012) или в качестве одного из семейств надсемейства Ardeosauroidea (Алифанов, 2012, 2016). Представителей типового семейства последнего таксона в кладистических работах обычно рассматривают в родстве с Gekkota (напр., Conrad, 2008; Gauthier et al., 2012).
В Сев. Америке Xenosauridae s.str. представлены вымершими родами Exostinus Соре, 1873 (поздний мел–ранний олигоцен; включает два, возможно, немонофилетичных вида: Bhullar, 2011) и Restes Gauthier, 1982 (поздний палеоцен). Древнейшие находки группы имеют здесь сеноманский возраст (Nydam, 2013). В Азии семейство известно по Oxia karakalpakensis Nessov, 1985 (рис. 4, а) из позднего альба Узбекистана (Gao, Nessov, 1998). Вид описан по несущей притупленные зубы зубной кости, обладающей характерным для ангвиморф строением ее заднего края: короноидный отросток развит, супраангулярный и ангулярный отростки не разрастаются окципитальнее уровня центра венечных костей.
Рис. 4.
Ящерицы семейства Xenosauridae (Anguimorpha): а – Oxia karakalpakensis Nessov, 1985; голотип CMGE, № 28/12000 (по: Gao, Nessov, 1998, фиг. 3, часть), левая зубная кость лингвально; Узбекистан, верхний апт; б – “Xenosauridae” gen. indet. (?Ardeosauridae fam. indet.); экз. ПИН, № 3334/515 (Алифанов, 2000, рис. 28 , г), задний край левой зубная кости лабиально; в – Xenostius futilus gen. et sp. nov. (Xenosauridae, Anguimorpha), голотип ПИН, № 3334/522, левая зубная кость лингвально; Монголия, Увэрхангайский аймак, местонахождение Хобур; нижний мел, верхний апт–нижний альб, хухтыкский горизонт. Длина масштабной линейки – 3 мм для а, 1 мм для б и в.
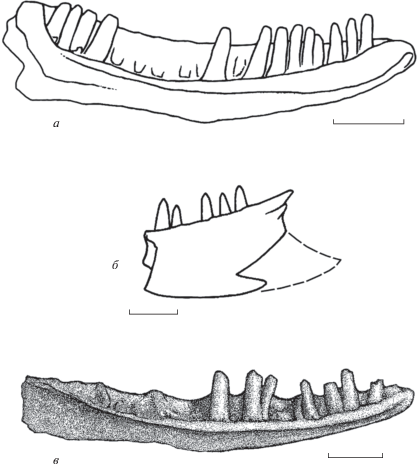
Открытие Oxia karakalpakensis Nessov, 1985 указывает на раннее (допозднемеловое) появление ксенозаврид. Гипотеза их происхождения в Центральной Азии должна быть подкреплена более древними данными. В качестве последних мы рассматривали экз. ПИН, № 3334/515 (рис. 4, б) из местонахождения Хобур (Алифанов, 2000). Однако наше переизучение этого образца, представленного фрагментом зубной кости с зубами, показало: его задний край, строение которого, как отмечалось, имеет диагностическое значение, обломан, при этом имитирует строение заднего края зубной кости Anguimorpha. Скорее всего, образец принадлежит представителю сцинкоморф. Тем не менее, ксенозавриды в ассоциации ящериц Хобура присутствуют, о чем позволяют судить представленные в данной работе образцы.
Загадочно происхождение Shinisauridae, обитающих ныне на небольшой территории, охватывающей Юж. Китай и Сев. Вьетнам. О том, что в прошлом это семейство было расселено шире, указывают находки его представителей в эоцене Северной Америки (Conrad, 2006) и миоцене Европы (Klembara, 2008). Судя по всему, группа возникла на одной из трех перечисленных территорий. Нам кажется вероятным ее происхождение в меловой Европе, с последующим распространением в Юго-Восточную Азию в постмезозойское время либо через Северную Америку и Центральную Азию, либо через Переднюю Азию и Индостан (не исключено, что последняя территория была первично связана зоогеографически не с южными континентами, а с Европой: Каландадзе, Раутиан, 1992; Kalandadze, Rautian, 1997).
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ОТРЯД LACERTILIA
ИНФРАОТРЯД ANGUIMORPHA
МИКРООТРЯД DIPLOGLOSSA
НАДСЕМЕЙСТВО XENOSAUROIDEA COPE, 1868
Диагноз. Диплоглоссы, отличающиеся наличием свободных остеодерм щиткового типа в области головы и отсутствием их в области туловища.
Состав. Семейства: Xenosauridae Cope, 1868; ранний мел Узбекистана и Монголии, поздний мел – современность Северной Америки; Dorsetisauridae Hoffstetter, 1967; поздняя юра Северной Америки, титон – ранний неоком Европы, ранний мел Центральной Азии; Shinisauridae Ahl, 1930; эоцен Северной Америки, ранний миоцен Европы, современность Юж. Китая и Сев. Вьетнама.
Замечания. Как отмечалось выше, для ангвиморф микроотряда Diploglossa, принимаемых нами в составе надсемейств Xenosauroidea и Anguoidea, характерен диплоголоссопалатинарный тип неба. Надсемейства отличаются по степени развития остеодерм, которые у первых концентрируются только в области головы, а у вторых развиваются в покрывающий все тело панцирь. Как и Xenosauridae s. str., надсемейство Anguoidea требует ревизии состава и внутригрупповых филогенетических связей. Ревизия должна касаться ископаемых Glyptosauridae, а также современных Anguidae (эту группу, возможно, необходимо разделить на два-три семейства). Судя по типу неба, семейство Anniellidae является частью таксона Platynota, а не Diploglossa, как обычно считается. В составе платинот они проявляют сходство по ряду краниологических признаков и, видимо, родство с Lanthanotidae (Алифанов, 2012).
СЕМЕЙСТВО DORSETISAURIDAE HOFFSTETTER, 1967
Типовой род – Dorsetisaurus Hoffstetter, 1967; поздняя юра Северной Америки и Европы, поздняя юра – ранний мел Европы.
Диагноз. Носовые отверстия удлиненные. Лобные кости парные. Латеральные отростки заднелобных костей раздвоены для сочленения с медиальным отростком заглазничных костей. Остеодермальная скульптура покровных костей черепа среднещиткового типа. Зубы по прикреплению эу- и субплевродонтные. Их верхушки ланцетовидные по форме.
Состав. Помимо типового, роды: Chometokadmon Costa, 1864; ранний мел Италии; Kuwajimalla Evans et Manabe, 2008; ранний мел Японии; Paradorsetisaurus gen. nov.; ранний мел Монголии.
Замечания. Диагноз семейства составлен с учетом его нового состава. Филогенетические связи родов Dorsetisauridae показаны на рис. 5. Схема отражает несомненное близкое родство Dorsetisaurus Hoffstetter, 1967 и Paradorsetisaurus gen. nov., основанное на значительном сходстве строения зубов, а также базальность Chometokadmon Costa, 1864, судя по слабому расширению верхушечной части зубов и их увеличенному числу.
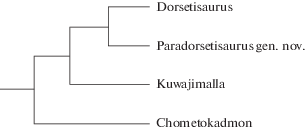
Род Paradorsetisaurus Alifanov, gen. nov.
Название рода от para греч. – подле и Dorsetisaurus.
Типовой вид – Paradorsetisaurus postumus sp. nov.
Диагноз. Премаксиллярный и скуловой отростки верхнечелюстных костей относительно длинные, последний заострен на конце. Дорсально на медиальной ветви премаксиллярного отростка выражен бугорок. Крупнейшие зубы расположены в середине ряда. Их вершины уплощены, изогнуты немного вовнутрь, повернуты под небольшим углом к оси челюсти и выходят за край челюстной кости примерно на половину их собственной высоты. Кромки вершинок протяженные и гладкие. Число верхнечелюстных зубов – 15.
Видовой состав. Типовой вид.
Сравнение. От других родов семейства новый отличается развитием дорсального бугорка на медиальной ветви премаксиллярного отростка и меньшим числом зубов. В сравнении с типовым родом он специфичен по относительно длинным премаксиллярному и скуловому отросткам верхнечелюстных костей и повороту вершинок зубов под углом к оси челюсти. Дополнительно новый род отличается от Kuwajimalla Evans et Manabe, 2008 гладкими режущими кромками, а от Chometokadmon Costa, 1864 – более высокими зубами и расширенными верхушками.
Paradorsetisaurus postumus Alifanov, sp. nov.
Табл. XIV , фиг. 1–4
Dorsetisaurus [sp.]: Alifanov, 1993, с. 9; 2000, с. 382.
Название вида postumus лат. – родившийся последним.
Объяснение к таблице XIV
Фиг. 1–4. Paradorsetisaurus postumus gen. et sp. nov. (Dorsetisauridae), голотип ПИН, № 3334/517, правая верхнечелюстная кость: 1 – лабиально, 2 – лингвально, 3 – вентрально; 4 – зубы лабиально, увеличено. Обозначение: Tpmx – дорсоростральный бугорок.
Фиг. 5 – Lacertilia (?Dorsetisauridae) fam. indet., экз. ПИН, № 3334/519, фрагмент лобной кости дорсально.
Фиг. 6 . Xenostius futilus gen. et sp. nov. (Xenosauridae s.str.), голотип ПИН, № 3334/522, левая зубная кость лингвально.
Монголия, Увэрхангайский аймак, местонахождение Хобур; нижний мел, верхний апт–нижний альб, хухтыкский горизонт.

Голотип – ПИН, №3334/517, правая верхнечелюстная кость; Монголия, Увэрхангайский аймак, местонахождение Хобур; нижний мел, верхний апт–нижний альб, хухтыкский горизонт.
Описание (рис. 2, ж). Резкого перехода от дорсального к окципитальному отростку верхнечелюстных костей нет. Окципитальный отросток этих костей составляет половину от длины всей кости. Предчелюстной отросток широкий в основании. Его длина превышает четверть от общей длины кости. Реконструкция обломанной латеральной части рострального отростка показывает, что носовые отверстия были укрупненными. Длина основания дорсального отростка составляет треть от общей длины. Лабиально на внешней поверхности верхнечелюстной кости расположены многочисленные отверстия, в нижнем ряду их не менее семи. Супрадентальный гребень широкий поперечно, особенно в третьей четверти кости, то есть, на уровне 8–11 зубов. Альвеолярное отверстие расположено на дорсальной поверхности этого гребня на уровне десятого зуба. Зубной край верхнечелюстной кости фестончатый. Зубы сидят в неглубоких лунках. По прикреплению они субплевродонтные. В основании зубов обычно сформировано небольшое резорбционное отверстие. Незначительные отложения цемента также развиты. За край зубного гребня зубы выходят больше чем на половину собственной высоты.
Размеры голотипа в мм. Длина верхнечелюстной кости (реконструкция) – 14.5 (15.5), ее высота – 4.3, высота наиболее крупного верхнечелюстного зуба – 2.5.
Замечания. Из коллекции остатков ящериц местонахождения Хобур интересен экз. ПИН, № 3334/519 (табл. XIV , фиг. 5). Это непарная лобная кость, которая принадлежит сравнительно крупной форме. Кость широкая, со скульптурированной дорсальной поверхностью и серией отверстий для питающих сосудов, расположенных в два ряда от условной срединной линии. Субольфакторные отростки низкие и широкие в окципитальной части костей, а также тонкие и высокие в их ростральной части. Фасетки для крепления предлобных костей глубокие. Их задний край расположен на уровне середины орбиты. В целом форма, представленная экз. ПИН, № 3334/519, внешне сходна по строению лобной кости c Dorsetisaurus, но отличается непарностью и отсутствием поперечных желобков на дорсальной поверхности. Не исключено, что последний образец принадлежит к описанному выше виду, однако до появления новых и более полных данных он формально не может быть определен точнее, чем Lacertilia fam. indet.
Материал. Голотип.
СЕМЕЙСТВО XENOSAURIDAE COPE, 1868
Типовой род – Xenosaurus Соре, 1886, современная Мексика.
Диагноз. На костях крыши черепа обычно развита остеодермальная скульптура гранулярного или мелкощиткового типа. Лобные кости всегда непарные и узкие. Чешуйчатые кости уплощены окципитально. Зубы притупленные, несут ростральное плечо.
Состав. Помимо типового, роды Exostinus Соре, 1873, поздний мел–ранний олигоцен Северной Америки; Restes Gauthier, 1982, поздний палеоцен Северной Америки; Oxia Nessov, 1985, ранний мел Узбекистана; Xenostius gen. nov., ранний мел Центральной Монголии.
Род Xenostius Alifanov, gen. nov.
Название рода – анаграмма от Exostinus.
Типовой вид – Xenostius futilus sp. nov.
Диагноз. Зубные кости длинные, тонкие, выпрямленные и слабо расширены окципитально. Задний край этих костей не формирует глубокой вырезки. Фланг субдентального гребня узкий на всем протяжении. Зубы относительно крупные, прямые и слабо расширены книзу. Их выступающая за дорсальную кромку зубной кости часть составляет от трети до половины общей высоты. Крупнейшие зубы формируются в средней части ряда. На верхушках развито небольшое ростральное плечо. Общее число зубов – 20.
Видовой состав. Типовой вид.
Сравнение. Новый род уникален в составе семейства, судя по небольшим размерам и тонкости зубной кости. От Oxia Nessov, 1985 он дополнительно отличается слабой ростральной расширенностью фланга субдентального гребня, наличием четко выраженного рострального плеча близ вершины зубов, а также более высокой частью зубов, выступающей за кромку зубного гребня.
Xenostius futilus Alifanov, sp. nov.
Табл. XIV , фиг. 6
Название вида futilus лат. – хрупкий.
Голотип – ПИН, № 3334/522, левая зубная кость; Монголия, Увэрхангайский аймак, местонахождение Хобур; нижний мел, верхний апт – нижний альб, хухтыкский горизонт.
Описание (рис. 4, в). Зубные кости низкие и почти прямые. Их лабиальная сторона заметно округлая. Ростральная часть меккелева канала ориентирована вентромедиально, что обычно характерно для ангвиморф. Расширенное основание крупнейших зубов выделенной переходной зоны от верхушечной части не имеет. Все зубы в целом ориентированы вертикально, иногда с небольшим отклонением назад или вперед. Фасетку, разделяющую центральную вершинку зуба от ее рострального плеча, у формы из Хобура установить не удается.
Размеры голотипа в мм. Длина зубной кости – 7.4, высота ее заднего края – 1.2 (реконструкция – 1.5), высота самого высокого нижнечелюстного зуба – 1.1.
Материал. Кроме голотипа, экз. ПИН, № 3334/521; левая зубная кость с зубами; типовое местонахождение.
О РАЗНООБРАЗИИ ЯЩЕРИЦ ЦЕНТРАЛЬНОЙ АЗИИ В ЮРЕ И РАННЕМ МЕЛУ
Ассоциация ящериц Хобура включает две группы семейств (рис. 1). Часть одной из них имела широкое распространение не только в раннем мелу Центральной Азии, но и за ее пределами: она представлена в юре Европы, Северной Америки (Paramacellodidae и Dorsetisauridae) и Африки (Paramacellodidae). Другая часть отмечается в юре (Ardeosauridae) и раннем мелу Европы (Globauridae). Все представители этой группы, видимо, имели распространение на территории юрской Лавразии, или на территории Афролавразии, т.е. Лавразии, соединенной с Африкой.
Для второй группы характерна эндемичность для территории раннемеловой Центральной Азии. Составляющие ее семейства (Eoxantidae, Gekkonidae s.l., Hodzhakuliidae, Temujiniidae и Xenosauridae) впервые появились в раннем мелу Центральной Азии и приурочены к территориальной изоляции этого палеоконтинента, длившейся на протяжении заключительной части юры и большей части раннего мела. Изоляция Центральной Азии, как часть процесса распада Афролавразии, была предложена Н.Н. Каландадзе и А.С. Раутианом (1992; Kalandadze, Rautian, 1997).
За пределами Монголии ассоциации ящериц Хобура по возрасту, несомненно, соответствуют две ящерицы семейства Hodzhakuliidae: Pachygenys thlastesa Gao et Cheng, 1999 из формации душань (апт–альбский возраст данной формации установлен С. Лукасом: Lucas, 2006) в Китае (Шаньдун); и другой вид того же рода – P. adashii Ikeda et al., 2015 из нижней части формации сасаяма (рубеж апта и альба) в Японии (Хего).
Любопытен состав единственной постхобурской (позднеальбской) ассоциации ящериц из нижней части ходжакульской свиты в Узбекистане, в которой установлено четыре группы (Gao, Nessov, 1998). По данным наших определений, помимо обсуждавшихся выше Xenosauridae, ходжакульская ассоциация включает также семейства Hodzhakuliidae, Ardeosauridae gen. indet. и Temujiniidae gen. indet. Такой состав, даже с поправкой на неполноту данных, указывает на снижение разнообразия ящериц в постхобурское время. Судя по данным из позднего мела Монголии, из числа известных в Хобуре семейств в последующее время вымерли только Hodzhakuliidae и Dorsetisauridae. Семейство Xenosauridae s. str. также исчезает в Центральной Азии, но появляется в позднем мелу другого континента – Северной Америки.
Разнообразие ящериц вблизи границы апта и альба контрастирует с низким разнообразием ящериц более раннего времени. Например, в юрских отложениях Центральной Азии представлено только одно семейство – Paramacellodidae (Changetisaurus estesi Nessov, 1992, Sharovisaurus karatauensis Hecht et Hecht, 1984, средняя и поздняя юра Казахстана соответственно; Mimobecklesisaurus gansuensis Li, 1991, поздняя юра Китая). Единственная для Центральной Азии ящерица неокомского (видимо, ранненеокомского) возраста происходит из ходжиабатской свиты Кыргызстана (Lacertilia gen. indet.: Аверьянов, Федоров, 2004; Ardeosauridae: Алифанов, 2012).
Среди дохобурских ящериц в Монголии отмечаются находки Norellius nyctisaurops Conrad et Daza, 2015 (Ardeosauridae, опр. автора) из ундурухинской свиты (формация оош), которая выходит на поверхность в Центральной Монголии и Bavarisauridae gen. indet. (Алифанов, 2000, 2012) из цаганцабской свиты в Западной Монголии. Отмеченные стратоны входят в цаганцабский горизонт, для которого известны данные абсолютного датирования. Последние укладываются в интервал от 141 до 119 млн лет (Shuvalov, 2000), соответствующий, скорее, готерив–баррему, но не всему неокому (Shuvalov, 2000) и не баррем–апту (Lucas, 2006).
За пределами Монголии ящерицы предположительно баррем–аптского возраста известны в Японии из серии Тетори: Kaganaias hakusanensis Evans et al., 2006 (Dolichosauridae), Kuwajimalla kagaensis Evans et Manabe, 2008 (Dorsetisauridae; опр. автора) и Sakurasaurus shokawensis Evans et Manabe, 1999 (Ardeosauridae; опр. автора) из формаций куваджима (Исикава) и окуродани (Гифу) соответственно. Есть данные о находках сходного возраста из илекской (Paramacellodidae indet., Xenosauridae indet.: Аверьянов, Файнгерц, 2001) и муртойской (Paramacellodidae indet.: Averianov, Skutchas, 1999) свит в России (Кемеровская обл. и Забайкалье, соответственно). Многочисленны находки ящериц (Jehololacerta formosa Ji et Ren, 1999; Dalinghosaurus longidigitus Ji, 1998; Liaoningolacerta brevirostra Ji, 2005; Liushusaurus acanthocaudata Evans et Wang, 2010; Yabeinosaurus tenuis Endo et Shikama, 1942) из отложений серии Джехол (баррем–апт) в Северо-Восточном Китае. Все они, скорее всего, принадлежат сцинкоморфам семейства Ardeosauridae, которое, по нашим представлениям, являлось доминантной группой на территории Центральной Азии в течение всего раннего мела.
На фоне в целом невысокого разнообразия раннемеловых ящериц Центральной Азии примечателен его апт-альбский максимум, установленный с помощью данных из местонахождения Хобур. Подъем связан, видимо, не только с удачными для аккумуляции остатков тафономическими условиями, но и сопровождался формированием благоприятных для мелких пресмыкающихся условий (потепление, рост разнообразия и биомассы насекомых). Это первый из двух меловых подъемов разнообразия ящериц в Центральной Азии с момента их появления в палеонтологической летописи. Второй, и гораздо более высокий пик разнообразия фиксируется в кампане (Алифанов, 2000).
Важно отметить, что юрско-раннемеловой временной отрезок представляет собой отдельный этап исторического развития ящериц. Отметим связанное с ним преобладание сцинкогеккономорф, особенно Scincomorpha (Paramacellodidae в юре, Ardeosauridae в раннем мелу в целом, Hodzhakuliidae в позднем апте и альбе). Такой состав доминант резко контрастирует с другими подобными этапами: позднемеловым, в течение которого преобладали ящерицы инфраотряда Chamaeleomorpha (Macrocephalosauria и Priscagamia), и палеогеновым, в течение которого наибольшего разнообразия достигли ящерицы микроотряда Pachyglossa (Changjiangosauridae, Uromastycidae и Agamidae).
С учетом сказанного выше, в рамках юрско-раннемелового этапа исторического развития ящериц Центральной Азии удается реконструировать несколько фаз. Они приходятся на вторую половину юры (доминирование Paramacellodidae), неоком – ранний апт (доминирование Ardeosauridae и поддержание архаического разнообразия), поздний апт – ранний альб (формирование максимума разнообразия в связи с появлением эндемичных групп) и поздний альб (сокращение разнообразия предыдущей фазы).
Список литературы
Аверьянов А.О., Файнгерц А.В. Ящерицы раннего мела Западной Сибири // Вопросы герпетологии. Матер. I съезда Герпетол. oб-ва им. А.М. Никольского. Пущино–Москва, 2001. С. 6–8.
Аверьянов А.О., Федоров П.В. Раннемеловая ящерица из ходжиабадской свиты Ферганы (Кыргызстан) // Вестн. СПбГУ. Сер. 7. 2004. Вып. 1. С. 81–82.
Алифанов В.Р. Древнейший геккон (Lacertilia, Gekkonidae) из нижнего мела Монголии // Палеонтол. журн. 1989. № 1. С. 124–126.
Алифанов В.Р. Макроцефалозавры и ранние этапы эволюции ящериц Монголии. М: ГЕОС, 2000. 126 с. (Тр. Палеонтол. ин-та РАН. Т. 272).
Алифанов В.Р. Отряд Lacertilia // Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Часть 2 / Ред. Курочкин Е.Н., Лопатин А.В. М: ГЕОС, 2012. С. 7–136.
Алифанов B.Р. Ящерицы семейства Hodzhakuliidae (Scincomorpha) из раннего мела Монголии // Палеонтол. журн. 2016. № 5. С. 73–82.
Алифанов B.Р. Ящерицы семейства Temujiniidae (Iguanomorpha): находки из апт-альба Монголии, классификация и географическое происхождение // Палеонтол. журн. 2018. № 6. С. 63–73.
Каландадзе Н.Н., Раутиан А.С. Система млекопитающих и историческая зоогеография // Филогенетика млекопитающих / Ред. Россолимо О.Л. М.: Изд-во МГУ, 1992. С. 44–152 (Сб. тр. Зоол. музея МГУ. Т. 29).
Федоров П.В., Несов Л.А. Ящерица рубежа средней и поздней юры Северо-Восточной Ферганы // Вестн. СПбГУ. Сер. 7. 1992. Вып. 3. № 21. С. 9–14.
Alifanov V.R. Some peculiarities of the Cretaceous and Palaeogene lizard faunas of the Mongolian People’s Republic // Kaupia. 1993. H. 3. P. 9–13.
Alifanov V.R. The fossil record of Cretaceous lizards of Mongolia // The Age of Dinosaurs in Russia and Mongolia / Eds. Benton M., Shishkin M.A., Unwin D.M., Kurochkin E.N. Cambridge: Univ. Press, 2000. P. 368–389.
Averianov A.O., Skutchas P.P. Paramacellodid lizard (Squamata, Scincomorpha) from the early Cretaceous of Transbaikalia // Russ. J. Herpetol. 1999. V. 6. № 2. P. 115–117.
Bever G.S., Bell C.J., Maisano J.A. The ossified braincase and cephalic osteoderms of Shinisaurus crocodilurus (Squamata, Shinisauridae) // Palaeontol. Electron. 2005. V. 8. № 1. P. 1–36.
Bhullar B.-A. S. The power and utility of morphological characters in systematics: a fully resolved phylogeny of xenosaurus and its fossil relatives (Squamata: Anguimorpha) // Bull. Mus. Comp. Zool. 2011. V. 160. № 3. P. 65–181.
Borsuk-Białynicka M. Carolinidae, a new family of Xenosaurid-like lizards from the Upper Cretaceous of Mongolia // Acta Palaeontol. Pol. 1985. V. 30. № 3–4. P. 151–176.
Borsuk-Białynicka M. Carussia, a new name for the late Cretaceous lizards from the Upper Cretaceous of Mongolia // Acta Palaeontol. Pol. 1987. V. 32. № 1–2. P. 153.
Broschinski A. The lizards from the Guimarota mine // Guimarota, a Jurassic ecosystem / Eds. Martin T., Krebs B. München: Dr. F. Pfeil Verlag, 2000. P. 59–68.
Caldwell M.W., Randall L., Nydam R.L. et al. The oldest known snakes from the Middle Jurassic-Lower Cretaceous provide insights on snake evolution // Nature Communicat. 2015. № 5976. P. 1–11.
Camp C.L. Classification of the lizards // Bull. Amer. Mus. Natur. Hist. 1923. V. 48. P. 289–481.
Conrad J.L. Skull, mandible, and hyoid of Shinisaurus crocodilurus Ahl (Squamata, Anguimorpha) // Zool. J. Linn. Soc. 2004. V. 141. № 3. P. 399–434.
Conrad J.L. An Eocene shinisaurid (Reptilia, Squamata) from Wyoming, USA // J. Vertebr. Paleontol. 2006. V. 26. № 1. P. 113–126.
Conrad J.L. Phylogeny and systematics of Squamata (Reptilia) based on morphology // Bull. Amer. Mus. Natur. Hist. 2008. V. 310. 182 p.
Daza J.D., Alifanov V.R., Bauer A.M. A redescription and phylogenetic reinterpretation of lizard Hoburogekko suchanovi Alifanov 1989 (Squamata: Gekkota) // J. Vertebr. Paleontol. 2012. V. 32. № 6. P. 1303–1312.
Estes R. Sauria terrestria, Amphisbaenia. Stuttgart, N.Y.: G. Fischer Verlag, 1983. 249 s. (Handbuch der Paläoherpetologie. T. 10A).
Evans S.E., Manabe M. An early herbivorous lizard from the Lower Cretaceous of Japan // Palaeontology. 2008. V. 51. Pt 2. P. 487–498.
Evans S.E., Raia P., Barbera C. The Lower Cretaceous lizard genus Chometokadmon from Italy // Cret. Res. 2006. V. 27. № 5. P. 673–683.
Fernandez V., Buffetaut E., Suteethorn V. et al. Evidence of egg diversity in Squamate evolution from Cretaceous anguimorph embryos // PLoS One. 2015. V. 10. № 7. P. 1–20.
Gao K., Nessov L.A. Early Cretaceous squamates from the Kyzylkum Desert, Uzbekistan // N. Jb. Geol. Paläontol. Abh. 1998. Bd 207. S. 289–309.
Gao K., Norell M.A. Taxonomic revision of Carusia (Reptilia: Squamata) from the late Cretaceous of the Gobi Desert and phylogenetic relationships of anguimorphan lizards // Amer. Mus. Novit. 1998. № 3230. 51 p.
Gao K., Norell M.A. Taxonomic composition and systematics of late Cretaceous lizard assemblages from Ukhaa Tolgod and adjacent localities, Mongolian Gobi Desert // Bull. Amer. Mus. Natur. Hist. 2000. V. 249. P. 1–118.
Gauthier J.A., Kearney M., Maisano J.A. et al. Assembling the squamate tree of life: Perspectives from the phenotype and the fossil record // Bull. Peabody Mus. Natur. Hist. 2012. V. 53. № 1. P. 3–308.
Hoffstetter R. Coup d’oeil sur les sauriens (=lacertiliens) des couches de Purbeck (Jurassique supérieur d’Angleterre) // Problèmes Actuels de Paléontologie (Evolution des Vertebres). Coll. Intern. CNRS. 1967. T. 163. P. 349–371.
Kalandadze N.N., Rautian A.S. Historical zoogeography of terrestrial tetrapods and new method of global palaeogeographical reconstructions // Evolution of the biosphere / Eds. Rozanov A.Yu., Vickers-Rich P. Launceston: C. Tassel, 1997. P. 95–98 (Rec. Queen Victoria Mus., Art Gallery. №104).
Klembara J. A new anguimorph lizard from the lower Miocene of north-west Bohemia, Czech Republic // Palaeontology. 2008. V. 51. Pt 1. P. 81–94.
Lucas S.G. The Psittacosaurus biochron, Early Cretaceous of Asia // Cret. Res. 2006. V. 27. № 2. P. 189–198.
McDowell S.B.J., Bogert CM. The systematic position of Lanthanotus and the affinities of the anguinomorphan lizards // Bull. Amer. Mus. Natur. Hist. 1954. V. 105. № 1. P. 1–105.
Nydam R.L. Squamates from the Jurassic and Cretaceous of North America // Palaeobio. Palaeoenv. 2013. V. 93. № 4. P. 535–565.
Prothero D., Estes R. Late Jurassic lizards from Como Bluff, Wyoming, and their palaeobiogeographic significance // Nature. 1980. V. 286. № 5772. P. 484–486.
Seiffert J. Upper Jurassic lizards from Central Portugal // Mem. Serv. Geol. Port. 1973. V. 22. P. 1–85.
Shuvalov V.F. The stratigraphy and palaeobiogeography of Mongolia // The Age of Dinosaurs in Russia and Mongolia / Eds. Benton M. J., Shishkin M.A., Unwin D.M., Kurochkin E.N. Cambridge: Univ. Press, 2000. P. 256–278.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал