

Палеонтологический журнал, 2019, № 2, стр. 3-9
Новые виды спиральных нодозариид (Foraminifera) из нижнекарнийских отложений (триас) побережья моря Лаптевых
Л. К. Левчук a, А. В. Ядренкин a, *
a Институт нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН
Новосибирск, Россия
* E-mail: YadrenkinAV@ipgg.sbras.ru
Поступила в редакцию 26.12.2017
После доработки 15.03.2018
Принята к публикации 22.03.2018
Аннотация
Выполнено монографическое изучение фораминифер из нижнекарнийских отложений верхнего триаса побережья моря Лаптевых на севере Средней Сибири. Приводится описание трех новых видов спиральных нодозариид (Dainitella insueta sp. nov., Marginulinopsis incompta sp. nov. и Astacolus dolganensis sp. nov.), которые широко распространены в осипайской свите, а также встречены в низах немцовской и чайдахской свит.
ВВЕДЕНИЕ
Спиральные нодозарииды являются характерным элементом мезозойских комплексов фораминифер бореальных регионов. Их первое появление (род Astacolus) отмечается в средней–верхней перми, однако широкое латеральное распространение и вспышка таксономического разнообразия приходится на верхний триас. Представители ряда родов на севере Средней Сибири получили широкое распространение в начале позднего триаса, что приурочено к раннекарнийской трансгрессии – периоду формирования отложений осипайской свиты. Спиральные нодозарииды родов Astacolus, Marginulinopsis и Marginulina из нижнекарнийских отложений севера Средней Сибири (Нордвикский район) впервые были установлены А.А. Герке (1961). К сожалению, им были приведены только рисованные, хотя относительно многочисленные изображения, но без монографического описания, а сами определения были выполнены в открытой номенклатуре. В целом, спиральные нодозарииды обычны в составе комплексов фораминифер из нижнего карния и являются хорошим индикатором при определении возраста, причем они характерны как для прибрежных морских обстановок (осипайская и низы немцовской и чайдахской свит, Восточный Таймыр, Нордвикский р-н, кряж Прончищева, Оленекский залив), так и для морских обстановок, удаленных от берега (станционная свита, о. Котельный).
МАТЕРИАЛЫ И МЕТОДЫ
Материалом для написания статьи послужили личные сборы А.В. Ядренкина (м. Цветкова, м. Тумул, побережье Оленекского залива, о. Таас-Ары (низовья р. Лены), кряж Прончищева. Также были переизучены коллекции З.И. Булатовой из ранних сборов (1975, 1976 гг.) из опорных разрезов триаса севера Сибири (м. Цветкова, м. Тумул), видовые списки комплексов ею были ранее опубликованы в разделе “Фораминиферы” (Дагис, Казаков,1984). Все образцы имеют точную послойную привязку в разрезах, в соответствии с их ранее опубликованными послойным описанием и годами сбора (Дагис, Казаков, 1984; Казаков и др., 2002; Константинов и др., 2007; Попов и др., 2015), а именно: м. Цветкова – обнажение Т4 (далее везде обн.) – сборы 1975 и 1992 гг., устье р. Оленек, м. Тумул (обн.18 – сборы 1976 г., обн.1001 – 2007 г., обн. ТУ2 – 2010 г.), побережье Оленекского залива: м. Улахан-Крест, обн. 16 – сборы 1976 г., УК – 2010 г.), п. Ыстаннах-Хочо (обн.15 – сборы 1976 г., ЫХ4 – 2010 г.); о. Таас-Ары (низовья р. Лены, обн.11, сборы 2004 г.); кряж Прончищева, р. Песчаная (обн. ТР31 – сборы 2012 г.) (рис. 1). Все находки имеют возрастные датировки в соответствии с последними уточненными биостратиграфическими схемами А.Г. Константинова и др. (2013).
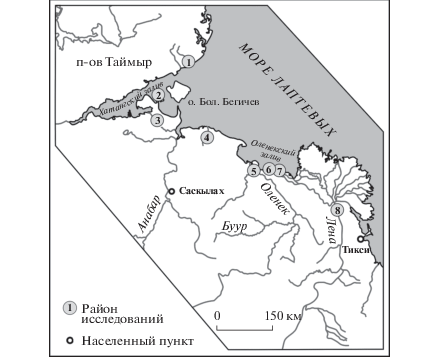
Рис. 1.
Карта-схема местоположения районов исследования нижнекарнийских отложений, из которых происходят находки описанных новых видов спиральных нодозариид: 1 – м. Цветкова (Восточное побережье п-ова Таймыр); 2 – Нордвикский участок (по: Герке, 1961), 3 – Чайдах-Гуримисский участок (по: Герке, 1961), 4 – кряж Прончищева (р. Песчаная), 5 – м. Тумул (пос. Усть-Оленек), 6 – м. Улахан-Крест (Оленекский залив), 7 – пос. Ыстаннах-Хочо (Оленекский залив), 8 – о. Таас-Ары (р. Лена).
В результате монографического изучения нами были описаны три новых вида: Dainitella insueta sp. nov., Marginulinopsis incompta sp. nov. и Astacolus dolganensis sp. nov. Представители рода Dainitella из триаса описаны впервые, ранее их первое появление отмечалось лишь в средней юре. В своей работе авторы придерживались системы фораминифер А. Леблик и Х. Таппан (Loeblich, Tappan, 1988), при описании морфологии раковин была использована методика, подробно изложенная в монографии “Проблемы …” (1975). При замерах учитывались следующие характеристики: L – длина; B – ширина; Bs – ширина спирального отдела; H – толщина раковины; Nsu – число камер в последнем обороте; N – общее число камер; Ns – число камер в спиральном отделе; Nu – число камер в прямом отделе; d – диаметр пролокулума. Все размеры, приведенные в текстовых таблицах, указаны в мм.
Коллекция хранится в лаборатории микропалеонтологии Ин-та нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН (ИНГГ СО РАН), г. Новосибирск, под №№ Т1975, Т2007, ВТ1976, ВТ1992. Работа выполнена при поддержке проекта ФНИ № 0331-2019-0005.
СИСТЕМАТИЧЕСКОЕ ОПИСАНИЕ
СЕМЕЙСТВО VAGINULINIDAE REUSS, 1860
ПОДСЕМЕЙСТВО LENTICULININAE CHAPMAN, PARR ET COLLINS, 1935
Род Dainitella Putrya, 1972
Dainitella insueta Levtchuk et Yadrenkin, sp. nov.
Табл. I , фиг. 1–8 (см. вклейку)
Название вида от insuetus лат. – необычный.
Голотип – ИНГГ СО РАН, № Т1975/1, микросферическая особь; устье р. Оленек, м. Тумул, обн. 18, обр. Я-837; верхний триас, нижний карний, аммонитовая зона omkutchanicum.
Описание. Раковина билатерально симметричная, средней удлиненности, биморфная; состоит из начальной спиральной и более поздней развернутой частей. В спиральном отделе раковина сжата с боковых сторон. Микросферические особи насчитывают 8–12 камер (от 4 до 7 камер в спиральной части, образующих от 1 до 1.2 оборота, и 3–5 в выпрямленном отделе. У мегалосферических генераций спиральная часть состоит из 2‒4 камер, очень редко из 5, которые образуют от 1/2 до 3/4 оборота спирали. Развернутая часть достаточно хорошо развита (обычно представлена 4–5 камерами), относительно объемная, округло-треугольной формы в поперечном сечении. Спиральный отдел достаточно компактно свернутый, ширина его заметно меньше общей ширины раковины. В спиральной части камеры изогнуто-треугольной формы, быстро возрастают в размере. При переходе к однорядному отделу камеры сбоку становятся скошенно-четырехугольными или прямоугольными в очертании, быстро расширяются, мало изменяясь в высоту. Швы между камерами слегка изогнутые, поверхностные, как на боковых, так и на брюшной сторонах. Боковые и брюшные стороны выпрямленного отдела у всех раковин немного выпуклые. Размер камер возрастает достаточно равномерно. Изредка последняя камера имеет недоразвитый вид, и тогда она меньше по ширине, чем предыдущая камера. В поперечном сечении раковина округленно-треугольная, иногда близка к грушевидной. Устьевая поверхность куполообразно округло-вздутая. Степень вздутости у микросферических особей выражена сильнее, чем у мегалосферических. Устье лучистое, расположено на очень низком бугорковидном возвышении у самого периферического края устьевой поверхности раковины. Стенка известковистая, с поверхности обычно матовая, непрозрачная.
Размеры, мм
| Параметры | L | B | H | dp | N | Ns | Nu |
|---|---|---|---|---|---|---|---|
| Голотип № Т1975/1 | 0.79 | 0.61 | 0.51 | 0.06 | 11 | 7 | 4 |
| Паратип № Т1975/2 | 0.62 | 0.50 | 0.37 | 0.1 | 9 | 5 | 4 |
| Паратип № Т1975/3 | 0.60 | 0.45 | 0.31 | 0.05 | 10 | 7 | 3 |
| Экз. № Т1975/5 | 0.51 | 0.37 | 0.24 | 7 | 4 | 3 | |
| Экз. № Т1975/6 | 0.46 | 033 | 0.27 | 5 | 3 | 2 | |
| Экз. № Т2007/1 | 0.46 | 0.34 | 0.24 | 0.09–0.05 | 6 | 3 | 3 |
| Экз. № Т2007/2 | 0.42 | 0.34 | 0.26 | 0.13 × 0.08 | 5 | 3 | 2 |
| Экз. № Т2007/3 | 0.42 | 0.29 | 0.22 | 0.08–0.12 | 6 | 4 | 2 |
| Пределы изменений | 1.0–0.28 | 0.7–0.25 | 0.48–0.21 | 0.05–0.16 | 4–12 | 0–7 | 2–5 |
Изменчивость. Видовая изменчивость описываемого вида связана с полиморфизмом генераций. Как известно, жизненный цикл фораминифер представляет собой чередование гаплоидного и диплоидного, т.е. полового (В) и бесполого (А) поколений – диморфизм. Иногда это чередование усложняется за счет того, что между двумя половыми поколениями (В) образуется не одно, а два бесполых поколения (А). Схема жизненного цикла тогда выглядит следующим образом: В – А1 – А2 – В. Авторам на достаточно представительном материале удалось пронаблюдать это явление у описываемого вида. Микросферические особи имеют маленькую начальную камеру (0.05–0.07 мм), более крупную и более вздутую раковину с большим общим числом камер и большим количеством камер в спиральной части раковины. Мегасферические особи поколений А1 и А2 по морфологическим признакам достаточно близки друг к другу, основным различием между ними являются размеры начальной камеры (0.08–0.10 мм у поколения А1 против 0.12–0.16 мм у поколения А2). Кроме того, раковины поколения А1 отличаются несколько большим общим числом камер, как в выпрямленной, так и спиральной частях.
Сравнение. В триасовых отложениях представители рода Dainitella не известны. По данным автора рода (Путря, 1972), они распространены в интервале юра – мел. В связи с этим, мы проводим сравнение с наиболее близкими по морфологическим признакам позднеюрскими видами дайнителл.
D. insueta sp. nov. отличается от D. explanata Putrya (Путря, 1972, с.160, табл. ХIХ, фиг. 5–6 ) более крупной раковиной (длина до 1.75 мм против максимально 1.0 мм у нового вида), особенно хорошо развитым спиральным отделом раковины, который представлен компактно свернутыми 11‒13 камерами (у нового вида 4–7). Кроме того, устьевая поверхность у раковин вида D. insueta sp. nov. куполообразно округло-вздутая по сравнению с выпуклой устьевой поверхностью у D. explanata.
Новый вид по морфологическим признакам достаточно близок к D. kunovatensis Putrya (Путря, 1972, с. 164, табл. ХХ, фиг. 9 ), отличается более широкой и более толстой раковиной, меньшим числом камер в спиральном отделе и более выпуклой формой устьевой поверхности.
Замечания. Особенные трудности при описании нового вида Dainitella insueta sp. nov. вызвало определение его родовой принадлежности. На первый взгляд, внешние морфологические признаки вроде бы свидетельствуют о принадлежности к роду Saracenella Franke, 1936, поскольку в изученном материале преобладают мегасферические особи, которые практически не имеют спиральной части или имеют начальную часть, образующую неполную быстро разворачивающуюся спираль. Кроме того, по мнению автора рода (Franke, 1936), поперечное сечение поздней части у сараценелл имеет сердцевидную форму, т. е., брюшная сторона раковин несколько вогнута в сторону спинного края. У описываемых же форм поперечное сечение округленно-треугольное, обычно с несколько выпуклыми боковыми и брюшной сторонами. Однако в изученном материале имеются микросферические особи, у которых наблюдается хорошо развитая спиральная часть, составляющая до 1.2 оборота у взрослых форм. Это в какой-то мере сближает наши формы с представителями рода Saracenaria Defrance (Defrance, 1824). Ярким отличием является поперечное сечение округленно-треугольное с выпуклыми боковыми и брюшной сторонами. Сараценарии обычно имеют поперечное сечение четкой треугольной формы, зачастую с заостренным или килеватым дорсальным периферическим краем. Межкамерные швы у них на боковых сторонах плоские, на брюшной обычно углубленные, а у описываемого вида швы поверхностные на всех сторонах. Формой раковины описанный вид достаточно близок и к представителям рода Marginulinopsis Silvestri (Silvestri, 1904), от которых отличается округленно-треугольной выпрямленной частью (у маргинулинопсисов раковина в поперечном сечении округло-овальная или яйцевидная). Таким образом, по внешним морфологическим признакам и по строению раковины формы, имеющиеся в нашем распоряжении, следует относить к роду Dainitella Putrya, 1972.
Считаем важным отметить, что относительно наиболее уплощенные экземпляры нового вида имеют некоторое сходство с раннекарнийским видом Falsopalmula pinguis Gerke (Герке, 1961; только рисованные изображения без описания), а именно, с формами, у которых нет типичных для фалзопалмул шевроновых камер (Герке, 1961, табл. LXVI , фиг. 6–7 ). На этом основании, а также по причине недостаточного на тот момент материала, в ранних работах Булатовой (Дагис, Казаков, 1984) и А.В. Ядренкина (1991а, 1994; Казаков и др., 2002; Попов и др., 2015) в приводимых ими списках фораминифер представители описываемого сейчас нового вида ошибочно относились к виду Falsopalmula pinguis Gerke.
Материал. Голотип, паратипы №№ Т1975/2, Т1975/3, экз. №№ Т1975/5, Т1975/6, Т2007/1, Т2007/2, Т2007/3 и еще 108 раковин хорошей и удовлетворительной сохранности, от 2 до 38 экз. в образце; север Средней Сибири, устье р. Оленек, мыс Тумул, обн. 18, и кряж Прончищева, р. Песчаная, обн. Тр31.
Род Marginulinopsis Silvestri, 1904
Marginulinopsis incompta Levtchuk et Yadrenkin, sp. nov.
Табл. I , фиг. 9–10 , табл. II , фиг. 1–3 (см. вклейку)
Lenticulina (Marginulinopsis) ex gr. prima (d’Orbigny): Герке, 1961, табл. XCII, фиг. 1, 2 ; табл. XCIII, фиг. 1; табл. XCIV, фиг. 6–8 ; табл. XCV, фиг. 1–9 .
Название вида от incomptus лат. – простой без прикрас.
Голотип – ИНГГ СО РАН, № ВТ1992/1, микросферическая особь; м. Цветкова, обн. 4, обр. Т4-5-п; верхний триас, нижний карний, зона по аммоноидеям tenuis-omkutchanikum.
Описание. Раковина крупная, удлиненная, биморфная, сжатая с боковых сторон в спиральном отделе; выпрямленный отдел в поперечном сечении имеет овально-яйцевидную форму. Периферический край округленно-суженный в спиральной части и округло-угловатый в выпрямленном отделе. Камеры в завитке сбоку слегка изогнуто-треугольные с уплощенной боковой поверхностью. В выпрямленном отделе – скошенно-четырехугольные, равномерно возрастающие в размере со слегка выпуклой боковой поверхностью камеры. Септальные швы почти параллельные, иногда слегка изогнутые, плоские. Общее число камер варьирует от 6 до 13. Спиральный отдел составляют 6–11, а выпрямленный – 2–5 камер. Свернутая часть раковины в большинстве случаев хорошо развита и насчитывает от 0.5 до 1.2 оборота спирали. Пупочная область плоская, заполнена прозрачным раковинным материалом, через который зачастую просвечивает начальная камера. Начальная камера у микросферических генераций имеет размеры от 0.05 до 0.08 мм, у мегасферических – 0.12–0.16 мм. Устье лучистое, расположено на устьевом возвышении у дорсального края. Устьевая поверхность слегка выпуклая, овально-яйцевидной формы. Стенка известковистая, гладкая.
Размеры, мм
| Параметры | L | B | Bs | H | dp | N | Ns | Nu |
|---|---|---|---|---|---|---|---|---|
| Голотип № ВТ1992/1 |
1.02 | 0.60 | 0.575 | 0.425 | 0.06 | 11 | 7 | 4 |
| Паратип № ВТ1992/2 |
1.2 | 0.575 | 0.625 | 0.4 | 0.14 | 11 | 7 | 4 |
| Паратип № ВТ1976/4 |
1.25 | 0.675 | 0.375 | 0.425 | 0.06 | 12 | 7 | 5 |
| Экз. № ВТ1976/3 | 0.89 | 0.64 | 0.34 | 0.05 | 10 | 9 | 1 | |
| Экз. № ВТ1976/5 | 1.39+ | 0.86 | 0.45 | 0.05 | 14+ | 10 | 4+ | |
| Пределы изменений | 1.25–0.65 | 0.68–0.50 | 0.63–0.32 | 0.43–0.30 | 0.05–0.16 | 6–13 | 5–11 | 1–5 |
Изменчивость. Описываемый вид имеет достаточно устойчивые внешние морфологические признаки. Изменчивость связана в основном с диморфизмом. Микросферические формы имеют маленькую начальную камеру (0.05–0.08 мм против 0.12–0.16 мм у мегасферических особей), несколько более крупную раковину и большее число камер в спиральном отделе (до 7–11 против 5–7 соответственно).
Сравнение. В нашем материале наиболее близким по морфологическим признакам к описанным формам является вид M. novosibirica Yadrenkin (Ядренкин, 1991б, с. 68, табл. 5 , фиг. 4–6 ) из нижнекарнийских отложений о-ва Котельный (Новосибирские острова), от которого новый вид отличается более широкой и более утолщенной раковиной, отсутствием струйчатой ребристости на поверхности раковины, отсутствием килеватости на периферическом крае. Кроме того, у M. incompta sp. nov. более развит спиральный отдел: от 0.9 до 1.2 оборота у микросферических и от 0.5 до 1.0 оборота у мегасферических особей против до 1.0 оборота у микросферических форм и до 0.33 оборота у мегасферических форм.
Новый вид достаточно близок к M. matutina (d’Orbigny) (Norvang, 1957, с. 374, фиг. 115, 117 ) из лейасовых отложений Ютландии (Дания). Отличается, в первую очередь, прямолинейностью раковины (у M. matutina спинной край изогнут в направлении брюшной стороны), плоскими межкамерными швами по всей длине раковины (у M. matutina они углубленные на брюшной стороне развернутого отдела за счет небольшой вздутости камер на ней), а также несколько большим числом оборотов в спиральном отделе (0.9–1.2 оборота против до одного оборота) и большим числом камер в нем (7–11 против 7–8).
Также внешние морфологические признаки нового вида сходны с таковыми у M. lituoides (Bornemann) из тех же отложений (Norvang, 1957, с. 375, фиг. 118, 120 ). Отличается большим числом камер и оборотов в спиральном отделе, более широкой и более утолщенной раковиной, а также плоскими швами (у M. lituoides они углубленные на брюшной стороне развернутого отдела).
Материал. Голотип, паратипы №№ ВТ1992/2, ВТ1976/4, экз. №№ ВТ1976/3, ВТ1976/5 и еще более 100 раковин хорошей и удовлетворительной сохранности, от 2 до 15 экз. в образце. Восточный Таймыр, м. Цветкова, обн. Т4; устье р. Оленек, м. Тумул, обн. 18; Оленекский залив, м. Улахан-Крест, обн. 16; п. Ыстаннах-Хочо, обн. 15; низовья р. Лены, о. Таас-Ары, обн. 11.
Род Astacolus Montfort, 1808
Astacolus dolganensis Levtchuk et Yadrenkin, sp. nov.
Табл. II , фиг. 4–7
Lenticulina (Astacolus) ex gr. varians (Bornemann): Герке, 1961, табл. C , фиг. 1, табл. CI , фиг. 1–7 .
Astacolus karnicus (Oberhausen): Oravecz-Scheffer, 1987, с. 115, табл. XXXIV , фиг. 1–3 ; Ядренкин, 1991а, табл. 4 , фиг. 1–2 .
Название вида от долгане – в честь проживающей на данной территории севера Сибири коренной народности.
Голотип – ИНГГ СО РАН, № ВТ1992/11, микросферическая особь; Восточный Таймыр, мыс Цветкова, обн.4; верхний триас, нижний карний, аммонитовая зона tenuis–omkutchanicum.
Описание. Раковина средних для рода размеров, биморфная, удлиненная с хорошо развитой спиральной частью и выпрямленным отделом, поперечное сечение которого имеет форму от суженно-овальной до яйцевидной. Умеренно удлиненная и умеренно уплощенная, со слабо выпуклыми боковыми сторонами. Спинной край раковины (в большей или меньшей степени) дугообразно изогнутый, суженный, брюшной край от почти прямого до слегка вогнутого у отдельных экземпляров. Всего насчитывает от 8 до 12 камер. Спиральная часть состоит из 5–9 камер обычно треугольной формы, которые образуют от 0.5 до 1.2 оборота. Спиральный отдел иногда выступает за пределы контура раковины. Выпрямленная часть состоит обычно из 2–4 камер, имеющих сбоку неправильно-четырехугольную форму. Причленение камер простое. Септальные швы плоские, изредка слегка углубленные между двумя последними камерами. Устье радиально-лучистое, расположено на выступающем устьевом бугорке в спинном углу последней камеры. Стенка известковистая, от стекловато-прозрачной до матовой, гладкая.
Размеры, мм
| Параметры | L | B | Bs | H | dp | N | Ns | Nu |
|---|---|---|---|---|---|---|---|---|
| Голотип № ВТ1992/11 | 0.55 | 0.30 | 0.25 | 0.20 | 0.04 | 10 | 7 | 3 |
| Паратип № ВТ1992/12 | 0.57 | 0.45 | 0.225 | 0.20 | 0.10 | 11 | 8 | 3 |
| Паратип № ВТ1992/13 | 0.75 | 0.35 | 0.325 | 0.19 | 0.06 | 11 | 8 | 3 |
| Экз. № ВТ1992/14 | 0.58 | 0.33 | 0.30 | 0.20 | 0.10 | 8 | 6 | 2 |
| Пределы изменений | 0.75–0.45 | 0.45–0.18 | 0.45–0.22 | 0.28–0.13 | 0.04–0.12 | 8–12 | 5–9 | 2–4 |
Изменчивость. Вид имеет достаточно устойчивые внешние морфологические признаки. Слегка меняется изогнутость спинного края раковины, он отклоняется в направлении брюшной стороны. У отдельных форм последняя камера бывает несколько меньше предыдущей, и тогда на брюшной стороне септальный шов становится слегка углубленным. Варьирует высота устьевого бугорка от слегка выпуклого до заметно конического. Микросферические и мегасферические формы в основном отличаются только размером начальной камеры (В-генерация имеет начальную камеру 0.04–0.06 мм, А – соответственно 0.10–0.12 мм).
Сравнение. Новый вид наиболее близок к виду A. bulatovae Yadrenkin, описанному из нижнекарнийских отложений м. Цветкова, Восточный Таймыр (Ядренкин, 1991б, с. 70, табл. 5 , фиг. 7 ). Отличается несколько менее крупной и менее удлиненной раковиной, более развитым спиральным отделом (0.5–1.2 оборота у нового вида против 0.2–0.5 у A. bulatovae), который зачастую выступает за брюшной край раковины, несколько большим числом камер и в спиральной, и в выпрямленной частях раковины, а также меньшими размерами начальной камеры у обеих генераций (микросферические формы: 0.04–0.06 мм против 0.06–0.09 мм у A. bulatovae и, соответственно, у мегасферических форм: 0.10–0.12 мм против 0.10–0.15).
По внешним морфологическим признакам описываемый вид близок к A. varians (Bornemann), описанному А. Норвангом (Norvang, 1957, с. 377, фиг. 124–134 ) из лейасовых отложений Ютландии (Дания). Лейасовый вид отличается компактной раковиной, спиральный отдел которой не превышает один оборот, а выпрямленный отдел представлен 1–3 камерами изогнуто-треугольной формы. В то же время, у A. dolganensis sp. nov. в спиральном отделе наблюдается до 1.2 оборота, а в выпрямленном отделе от 2 до 4 камер неправильно-четырехугольной формы.
Вид A. connudatus Tappan, описанный (Tappan, 1951, с.10, табл. 2 , фиг. 8–19 ) из верхнетриасовых отложений формации Shublik северного побережья Аляски, имеет определенное сходство с новым видом. Отличается наличием узкого киля на спинной стороне раковины, несколько утолщенными изогнутыми швами, большим числом камер в спиральном отделе (7–13 против 5–9 у A.dolganensis sp. nov.), более выпуклыми боковыми сторонами.
Материал. Кроме голотипа, паратипы №№ ВТ1992/12, ВТ1992/13, экз. № ВТ1992/14 и еще более 400 раковин хорошей и удовлетворительной сохранности, от 2 до 86 экз. в образце: Восточный Таймыр, м. Цветкова, обн. Т4; устье р. Оленек, м. Тумул, обн. 18; в ряде скважин Нордвикского и Чайдах-Гуримисского участков Нордвикского р-на (Герке, 1961).
Список литературы
Герке А.А. Фораминиферы пермских, триасовых и лейасовых отложений нефтеносных районов севера Центральной Сибири. Л.: Гостоптехиздат, 1961. 518 с. (Тр. НИИГА. Т. 120).
Дагис А.С., Казаков А.М. Стратиграфия, литология и цикличность триасовых отложений севера Средней Сибири. Новосибирск: Наука, 1984. 177 с. (Тр. ИГиГ СО АН СССР. Вып. 586).
Казаков А.М., Константинов А.Г., Курушин Н.И и др. Стратиграфия нефтегазоносных бассейнов Сибири. Триасовая система. Новосибирск: Изд-во СО РАН, 2002. 322 с.
Константинов А.Г., Соболев Е.С., Ядренкин А.В. Детальная биостратиграфия триасовых отложений нижнего течения реки Лена // Геол. и геофизика. 2007. Т. 48. № 9. С. 934–949.
Константинов А.Г., Соболев Е.С., Ядренкин А.В. Стратиграфия триаса восточной части побережья моря Лаптевых и Новосибирских островов // Геол. и геофизика. 2013. Т. 54. № 8. С. 1028–1046.
Попов А.Ю., Соболев Е.С., Ядренкин А.В. Строение и условия формирования осипайской и чайдахской свит (верхний триас) в районе кряжа Прончищева (Средняя Сибирь) // Изв. высш. учебн. завед. Геол. и разведка. 2015. № 3. С. 15–19.
Проблемы систематики спиральных нодозариид / Ред. А.А. Герке. Вильнюс: Минтис, 1975. 136 с.
Путря Ф.С. Лентикулиниды верхнеюрских отложений Западно-Сибирской низменности. М.: Недра, 1972. 303 с.
Ядренкин А.В. Стратиграфическое распространение фораминифер в нижнекарнийских отложениях севера Средней Сибири // Биостратиграфия и палеонтология триаса Сибири. Новосибирск: Изд-во ОИГГМ СО РАН, 1991а. С. 49–63.
Ядренкин А.В. Новые виды фораминифер из карнийских отложений севера Средней Сибири // Биостратиграфия и палеонтология триаса Сибири. Новосибирск: Изд-во ОИГГМ СО РАН, 1991б. С. 63–74.
Ядренкин А.В. Позднетриасовые фораминиферы севера Средней Сибири и их стратиграфическое значение. Автореф. … канд. г.-м. н. Новосибирск: ОИГГиМ СО РАН, 1994. 20 с.
Defrance M. In: H.M. Ducrotay de Blainville. Dictionaire des Sciences Naturelles, mollus-morf. Paris: F.G. Levrault, 1824. V. 32. P. 176.
Franke A. Die Foraminiferen des deutschen Lias // Abh. Preuss. Geol. Landes. N.F. 1936. H. 169. S. 1–138.
Loeblich A., Tappan H. Foraminiferal genera and their classification. Vol. 1–2. N.Y.: Van Nostrand Reinhold, 1988. 969 p.
Norvang A. The Foraminifera of the Lias Series in Jutland, Denmark // Medd. Dansk. Geol. For. 1957. Bd 13. H. 5. P. 279–413.
Oravecz-Scheffer A. Triassic foraminifers of the Transdanubian Central Ranges // Geol. Hung. Ser. Paleontol. 1987. Fasc. 50. 331 p.
Silvestri A. Ricerche strutturali su alcune forme dei trubi di Bonfornello (Palermo) // Mem. Accad. Nuovi Lincei. 1904. V. 22. P. 253.
Tappan H. Foraminifera from the Arctic Slope of Alaska. Pt 1. Triassic Foraminifera // U.S. Geol. Surv. Prof. Pap. 1951. № 236-A. 20 p.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал