

Палеонтологический журнал, 2019, № 2, стр. 26-32
О внутреннем строении раковины Semiplanella carinthica Sarytcheva et Legrand-Blain (Brachiopoda, Productida)
А. В. Пахневич *
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: alvpb@mail.ru
Поступила в редакцию 05.07.2016
После доработки 28.11.2017
Принята к публикации 14.12.2017
Аннотация
Изучено внутреннее строение спинной створки раковины брахиоподы Semiplanella carinthica Sarytcheva et Legrand-Blain, 1977. Предложена реконструкция расположения оснований брахиальных петель и мускулов-аддукторов. Необычное положение аддукторов и брахиальных петель являются результатом их поворота. Причинами необычного положения стали сильная скрученность раковины и необходимость поднятия переднего края раковины над поверхностью дна. Сходное строение брахиальных петель, а также, отчасти, замочного отростка, наблюдается у рода Beleutella Litvinovich, 1967 и, возможно, у Talasoproductus Litvinovich et Vorontsova, 1983. Предложено объединить эти три рода в новую трибу Semiplanellini tribe nov. в подсемействе Gigantoproductinae.
ВВЕДЕНИЕ
Среди каменноугольных брахиопод подсемейства Gigantoproductinae, семейства Monticuliferidae (по: Brunton et al., 2000), распространенных на территории Европы, наименее изученным является род Semiplanella Sarytcheva et Legrand-Blain, 1977, описанный в работе Т.Г. Сарычевой и М. Легран-Блан (1977). По мнению авторов рода (Сарычева, Легран-Блан, 1977), возможно, к нему принадлежал и вид Gigantoproductus rectestrius (Gröber, 1909) из нижнекаменноугольных отложений Казахстана. Также, с долей сомнения, этот род указывался из Северной Африки (Brunton et al., 2000).
Внутреннее строение брюшной створки этих продуктид неизвестно. О внутреннем строении спинной створки можно судить только по паратипу [ПИН, № 3704/1] типового вида Semiplanella carinthica Sarytcheva et Legrand-Blain, 1977. Но даже при беглом просмотре внутренней поверхности спинной створки видно необычное взаимное положение отпечатков аддукторов и брахиальных петель.
Авторы рода и вида подчеркивают (Сарычева, Легран-Блан, 1977), что раковина Semiplanella сильно спирально свернутая. Кардинальный отросток с массивной срединной лопастью. Следы прикрепления аддукторов удлиненно-овальные; они ветвистые в задней части и продольно-штриховатые в передней. Брахиальные петли под углом отходят от середины аддукторов. Эти признаки сильно отличают данный род от других родов подсемейства Gigantoproductinae.
Тем не менее, анализа такого расположения отпечатков аддукторов и брахиальных петель, а также морфофункциональной интерпретации этих данных так и не последовало. В связи с этим возникла необходимость подробно изучить внутреннее строение S. carinthica.
МАТЕРИАЛ И МЕТОДИКА
Изучение проводилось с помощью бинокулярных микроскопов МБС-10 и Leica M165C.
Исследована серия типовых экземпляров вида Semiplanella carinthica и дополнительный материал, происходящий из Казахстана. Голотип и 5 паратипов, колл. № 3704, и паратип И-51/1а Semiplanella carinthica Sarytcheva et Legrand-Blain, 1977 хранятся в Палеонтологическом ин-те им. А.А. Борисяка РАН (ПИН), Москва; нижний карбон, верхний визейский подъярус; Карнийские Альпы, Каринтия. 2 экз., колл. № 1019, хранятся на Кафедре региональной и исторической геологии геологического фак-та Московского государственного ун-та им. М.В. Ломоносова (МГУ); нижний карбон, серпуховский ярус; хр. Каратау, Казахстан.
В качестве сравнительного материала по морфологии продуктид подсемейства Gigantoproductinae изучены экземпляры из коллекций в Музее землеведения Московского государственного ун-та им. М.В. Ломоносова (МЗ МГУ), Государственном геологическом музее им. В.И. Вернадского (ГГМ), Москва, Музее естественной истории (Muséum d’Histoire Naturellе, [MNHN]), Париж, Франция:
Latiproductus latissimus (Sowerby, 1822), МЗ МГУ, № 31/139, спинная створка; нижний карбон, серпуховский ярус, белеутинский горизонт; р. Белеуты, обн. 4, сл. 24, Казахстан;
Latiproductus (?) edelburgensis subsahariensis Legrand-Blain, 1980, MNHN, паратип JRV 244 a 2, спинная створка; нижний карбон, низы серпуховского яруса; местонахождение Таудени (Taoudenni), Южная Сахара, Мали; MNHN A 3219 a 1, спинная створка; нижний карбон, низы серпуховского яруса; Южная Сахара, Мали;
Semiplanus semiplanus (Schwetzov, 1922), типовой экземпляр ГГМ, VI-30/35, раковина; нижний карбон, визейский ярус, низы алексинского горизонта; Калужская область, д. Стопкино; экз. ГГМ, VI-30/37, спинная створка; нижний карбон, низы алексинского горизонта; Калужская обл., р. Ока, д. Вороново.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ОБСУЖДЕНИЕ
На раковине паратипа вида Semiplanella carinthica наблюдаются следующие особенности внутреннего строения спинной створки. Следы прикрепления парных аддукторов по обе стороны септы располагаются почти друг над другом, но граница между ними не перпендикулярна септе (оси симметрии), а скошена (наклонена под углом к септе) (паратип ПИН, № 3704/1, табл. VII , фиг. 2а, см. вклейку). Поэтому для них более всего подходят названия: задние и передние. Места прикрепления аддукторов утолщены. На отпечатках задних аддукторов наблюдается ветвистая исчерченность, характерная для отпечатков аддукторов гигантопродуктид и ряда других продуктид. На передних – продольная исчерченность. Сохранность не позволяет реконструировать начальные и конечные отрезки брахиальных петель. Лучше сохранились большая часть нисходящей и восходящей дуг брахиальных петель, а также сам их изгиб (табл. VIII, фиг. 1б, 1в, см. вклейку). Можно констатировать, что восходящие ветви проходят вдоль передних аддукторов, что не свойственно другим гигантопродуктидам, у которых брахиальные петли расположены ниже отпечатков аддукторов. Непонятно, как расположена нисходящая часть петли у основания ее выхода между мускулами. Возможно, она опускается вперед, что изображено на рис. 1, а. Изгибы петель упираются в небольшие возвышения, которые являются брахиальными конусами (табл. VIII, фиг. 1г). Брахиальные конусы должны огибаться петлями, восходящие ветви которых отделяют их от отпечатков аддукторов (см. Brunton et al., 2000, с. 550, фиг. 385 c ). Но в изгибе петель у S. carinthica брахиальных конусов обнаружено не было. У гигантопродуктид они выполняют функцию поддержания мягкого лофофора. Брахиальные конусы у семипланелл значительно редуцированы и расположены у вершины изгиба брахиальных петель, т.е., гораздо ближе к переднему краю, чем обычно это бывает у гигантопродуктид. Вероятно, функцию поддержания лофофора они уже не выполняли.
Рис. 1.
Схематичное изображение внутреннего строения спинных створок Semiplanella carinthica Sarytcheva et Legrand-Blain, 1977 и Beleutella rara Litvinovich, 1969: а – S. carinthica, паратип ПИН, № 3704/1, ×1; Карнийские Альпы, Каринтия; нижний карбон, верхнeвизейский подъярус; б – B. rara, экз. МЗ МГУ, № 31/349, ×1; Центральный Казахстан, р. Белеуты, обн. 4, сл. 24; нижний карбон, серпуховский ярус, белеутинский горизонт. Обозначения: с.с – срединная септа; о.п.а – отпечаток переднего аддуктора; б.п – брахиальная петля; б.к – брахиальный конус; л.з.о – лопасть замочного отростка; м – миофор; о.з.а – отпечатки задних аддукторов; н.б.п – начало брахиальной петли; у – утолщение; я.у – якоревидное утолщение.
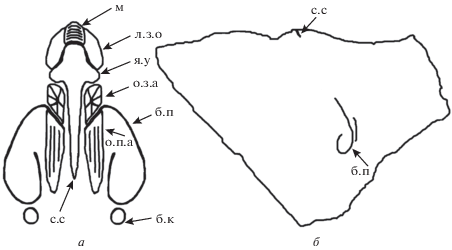
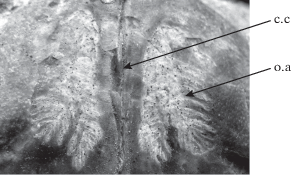
Рис. 2.
Отпечатки парных аддукторов на поверхности спинной створки Moderatoproductus moderatus Litvinovich et Vorontsova, 1983, экз. ГГМ, № II-108/21 из колл. М.А. Болховитиновой, ×1; Тульская обл., карьер у с. Граворонки; нижний карбон. Обозначения: с. с. – срединная септа, о. а. – отпечатки аддукторов.
Не совсем понятно, насколько близко подходят к изгибам брахиальных петель и брахиальным конусам отпечатки передних аддукторов. Наиболее четко видна часть следа прикрепления переднего аддуктора, примыкающая непосредственно к границе между парными аддукторами. Если радиальная исчерченность, действительно, соответствует отпечаткам передних аддукторов, то они должны очень близко подходить к брахиальным конусам и изгибу брахиальных петель. Не вызывает сомнения, что отпечаток переднего аддуктора, хотя бы ненамного, длиннее заднего (табл. VII , фиг. 2а; табл. VIII, фиг. 1б).
Строение следов прикрепления мускулов у S. carinthica отличается от таковых у представителей трибы Gigantoproductini. У брахиопод Gigantoproductus Prentice, 1950 и Moderatoproductus Litvinovich et Vorontsova, 1983 отпечатки парных мускулов аддукторов расположены на спинной створке почти параллельно друг другу, под небольшим углом (рис. 2). У Semiplanella они располагаются друг за другом. У Gigantoproductus и Moderatoproductus между отпечатками аддукторов с каждой стороны септы начинается брахиальная петля. Далее она почти перпендикулярно септе отходит в бок, к латеральному краю раковины, изгибается и резко опускается к переднему краю, частично огибая брахиальный конус. Принцип расположения аддукторов и строения брахиальной петли у этих брахиопод одинаков (рис. 3) (см. Brunton et al., 2000, с. 550, фиг. 385 c ).
Рис. 3.
Внутренняя поверхность спинной створки Moderatoproductus moderatus Litvinovich et Vorontsova, 1983, лектотип ГГМ, № VI-30/9, ×1; Рязанская обл., Еринские каменоломни; нижний карбон, визейский ярус, верхи алексинского горизонта. Обозначения см. на рис. 1.

Интересно пронаблюдать, насколько распространен такой тип положения аддукторов у древнейших продуктид или хонетид. У девонских линопродуктоидей аддукторы в спинной створке располагались ближе к макушке (замочному краю), их пары были по бокам от септы и каждый из парных мускулов не находился за другим, они были наклонены так же, как это наблюдается у родов Gigantoproductus и Moderatoproductus (см. Brunton et al., 2000, фиг. 383 -1f). Между ними начиналась брахиальная петля. Совершенно определенно такой план взаимного расположения в паре аддукторов сохранился и у раннекаменноугольных гигантопродуктид, но не у Semiplanella.
У хонетид, предков продуктид, аддукторы располагаются таким же образом (см. Афанасьева, 1988; Racheboeuf, 2000), как у раннекаменноугольных Gigantoproductus и Moderatoproductus, то есть этот тип расположения аддукторов оказывается весьма консервативным признаком, четко выраженным еще у предков продуктид. Поэтому расположение мускулов у Semiplanella является вновь приобретенным признаком, так как начало петли направлено от септы не к латеральному краю раковины, а к замочному краю. Тогда и изгиб брахиальной петли, после выхода между аддукторами, должен находиться выше границы их соприкосновения. А в случае, если основание петли направлено к латеральному краю, то петля не делает изгиба над границей заднего и переднего аддукторов.
Semiplanella отнесена к трибе Semiplanini (Brunton et al., 2000). Можно ожидать, что схожее строение будет и у других представителей группы. Особенно внешне близка Semiplanella к роду Semiplanus Sarytcheva in Sarytcheva et Sokolskaya, 1952. Она схожа с ним по характеру ребристости и положению игл на обеих створках (табл. VII, фиг. 1, 2г).
По строению замочного отростка Semiplanella сходна с родом Latiproductus Sarytcheva et Legrand-Blain, 1977: для этих двух родов характерен трехлопастный замочный отросток. Разница заключается в том, что у Semiplanella центральная лопасть приподнята над поверхностью спинной створки, а боковые лежат на ней. Подобная особенность строения замочного отростка не характерна для рода Latiproductus. Тем не менее, строение брахиальных петель и расположение аддукторов у Latiproductus и Semiplanus такое же, как и у Gigantoproductus и Moderatoproductus. Для рода Semiplanus это можно наблюдать на примере экземпляра ГГМ, VI-30/37 (см. также Сарычева, Сокольская, 1952, рис. 121 ), а также в статье Г. Жаковой (Żakowa, 1986, табл. I , фиг. 3). Подобное строение было отмечено мной и на ряде экземпляров, принадлежащим к различным видам рода Latiproductus. К роду Latiproductus относятся виды, для которых характерны следующие признаки: четкие, иногда утолщенные ребра, редкие иглы, которые постепенно укрупняются к переднему краю и в зависимости от этого располагаются на ребрах, не нарушая ребристость (маленькие иглы на висцеральном диске), или нарушают ее (крупные иглы на шлейфе). На шлейфе ребра огибают иглы, или их количество уменьшается после иглы, или увеличивается. Такие признаки присущи, в частности, экземплярам, для которых известно внутреннее строение спинной створки. Это экземпляры: L. edelburgensis subsahariensis (MNHN A 3219 a 1), найденный в Южной Сахаре (Мали) (MNHN JRV 244 a 2) из местонахождение Таудени из низов серпуховского яруса Южной Сахары (Мали), L. daguini (MNHN ML 409 с 4; см. Legrand-Blain, 1980, фиг. 22 ) из серпуховского яруса Алжирской Сахары, L. aff. volgensis (MNHN ML 574 a 4; см. Legrand-Blain, 1980, фиг. 10 ) из серпуховского яруса Алжирской Сахары, Latiproductus sp. 1 (MNHN A 1998 a 5; см. Legrand-Blain, 1980, фиг. 8 ) из серпуховского яруса Алжирской Сахары. Сходное строение брахиальных петель имеет и экземпляр L. latissimus (МЗ МГУ, № 31/137; см. Литвинович и др., 1969, табл. XIX , фиг. 1) из серпуховских отложений белеутинского горизонта Казахстана. У перечисленных экземпляров брахиальные петли расположены таким же образом, как у представителей родов Semiplanus, Moderatoproductus и Gigantoproductus, то есть, они огибают брахиальные конусы, отсекая их от мускулов-аддукторов.
Таким образом, Semiplanella является уникальным представителем гигантопродуктид со своеобразным положением аддукторов и брахиальных петель на спинной створке. Основным фактором, приведшим к столь значительным изменениям, стала очень сильная скрученность раковины (табл. VII , фиг. 2в), не настолько выраженная у Semiplanus и Latiproductus. Еще одна особенность, связанная с сильной скрученностью раковины – приподнятая над поверхностью створки центральная лопасть трехлопастного замочного отростка и лежащие на внутренней поверхности спинной створки боковые лопасти (табл. VII , фиг. 2а). Это было необходимо, чтобы мышцы-дидукторы не повторяли полный изгиб скрученной раковины, огибая макушечную область. Помимо этого, у заднего края спинной створки (в правой части) S. carinthica из Карнийских Альп обнаружено утолщение, которое располагается почти параллельно замочному краю (табл. VII , фиг. 2а). Возможно, оно есть и с противоположной стороны створки, но в связи с сохранностью экземпляра это видно не столь ясно. После этого утолщения скрученность задней части раковины усиливается. На голотипе ПИН, № 3704/2 в этом районе раковина сдавлена, и перегиб, следующий после утолщения, не наблюдается. Зато на раковинах серпуховских S. carinthica из Казахстана этот перегиб виден даже на не отпрепарированных раковинах. Этот признак, а также частые сближенные ребра с многочисленными мелкими иглами, короткие не ясно обособленные ушки, замочный отросток с сильно развитой центральной лопастью и боковыми лопастями, лежащими на поверхности спинной створки, – все это позволяет достоверно отнести упомянутые экземпляры к виду S. carinthica, и расширить его географическое и геологическое распространение до серпуховского яруса Казахстана.
Вероятно, своеобразное взаимное положение парных мускулов-аддукторов явилось результатом их поворота, что повлекло за собой и перемещение брахиальных петель. Так, восходящая дуга брахиальной петли стала проходить вдоль переднего парного аддуктора. При этом передний аддуктор удлинился по сравнению с задним (рис. 4). Произошло удлинение шлейфа раковины и сокращение висцеральной полости. Повлияло это и на строение лофофора. Если у таких родов, как Gigantoproductus и Moderatoproductus, брахиальные конусы служили для поддержки бесскелетной мягкой части лофофора и были хорошо развиты, то у семипланеллы эта их функция утеряна. Тогда каким же должен быть лофофор родов Gigantoproductus и Moderatoproductus? Лофофор ли это, как у Falafer epidelus Grant, 1972, или же это спиролоф, который закручивался вокруг брахиальных конусов? Этот вопрос требует отдельного исследования.
Рис. 4.
Схема расположения основания брахиальной петли (короткая черная черта) между отпечатками парных аддукторов и брахиальных конусов (круг). Стрелкой показано направление поворота основания брахиальной петли.
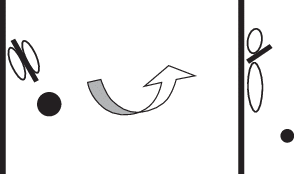
Но принадлежит ли к роду Semiplanella вид Gigantoproductus rectestrius из отложений нижнего карбона Казахстана? Автор вида П. Грёбер (Gröber, 1909) не изобразил внутреннее строение створок, без которого нельзя сделать заключение о принадлежности данного вида к роду Semiplanella. Также отсутствует и описание внутреннего строения. Не изучено внутреннее строение G. rectestrius и коллективом авторов, изучавшим раннекарбоновых брахиопод западной части Центрального Казахстана (Литвинович и др., 1969).
Для уточнения родовой принадлежности G. rectestrius был изучен экземпляр этого вида МЗ МГУ, № 31/146 к работе Н.В. Литвиновича и др. (1969). Он представлен брюшной створкой. Особенностей внутреннего строения отмечено не было. Скульптура G. rectestrius на брюшной створке сходна с таковой у S. carinthica, хотя ребра у G. rectestrius толще. Иглы встречаются крайне редко. Есть ли они на спинной створке, неизвестно. В более поздней работе (Литвинович, Воронцова, 1991) в качестве экземпляра Latiproductus rectestrius выдается еще и экз. МЗ МГУ, № 31/140, который ранее (Литвинович и др., 1969) был отнесен к виду Gigantoproductus edelburgensis (Phillips, 1836). В связи с вышесказанным, принадлежность вида G. rectestrius к роду Semiplanella остается недоказанной.
Однако близким по некоторым особенностям внутреннего строения является вид Beleutella rara Litvinovich, 1967 из отложений белеутинского горизонта Центрального Казахстана. Внутреннее строение брюшной створки B. rara известно. Для нее характерны широкие отпечатки дидукторов. Отпечатки аддукторов и дидукторов располагаются почти на одном уровне, отпечатки аддукторов чуть ближе к макушке створки. Возможно, есть небольшие брахиальные конусы (Литвинович и др., 1969, табл. XV , фиг. 1). Но внутреннее строение спинной створки известно не полностью. Хорошо заметны: большой цельный замочный отросток на массивном основании, который изначально имел, помимо одной крупной лопасти, еще две небольшие боковые лопасти; овальные отпечатки аддукторов; срединная септа (Литвинович и др., 1969, табл. XIV , фиг. 1б–1г, рис. 46 ). Описание, в основном, составлено по спинной створке МЗ МГУ, № 31/342. Но брахиальные петли не описаны, поскольку эта часть поверхности створки разрушена. Однако одна из брахиальных петель слабо проступает на другом экземпляре МЗ МГУ, № 31/349. Это брюшная створка с сильно стертой поверхностью. Местами наблюдается поверхность спинной створки. С одной стороны от предполагаемой срединной септы наблюдается еле заметная брахиальная петля (рис. 1, б). Она узкая, расположена вдоль септы по направлению к переднему краю, не огибает никаких структур. Подобное строение брахиальной петли сходно с таковой у S. carinthica. Поэтому род Beleutella Litvinovich, 1967 является наиболее близким к роду Semiplanella.
На фотографиях поверхности спинной створки Talasoproductus turlanensis Litvinovich et Vorontsova, 1983 (см. Brunton et al., 2000) хорошо видны сильно удлиненные отпечатки аддукторов. А это является одной из характерных особенностей внутреннего строения рода Semiplanella. К сожалению, ничего не известно о строении брахиальных петель Talasoproductus Litvinovich et Vorontsova, 1983. Вдоль замочного края идет невысокое утолщение, как у Semiplanella. В связи с тем, что найти типовой материал типового вида не удалось, род плохо изучен, приходится полагаться только на это изображение. Вероятно, род Talasoproductus также близок к Semiplanella.
В связи со значительными отличиями во внутреннем строении необходимо выделить роды Semiplanella и Beleutella, а возможно и Talasoproductus, в отдельную трибу в составе подсемейства Gigantoproductinae.
ОТРЯД PRODUCTIDA
НАДСЕМЕЙСТВО LINOPRODUCTOIDEA STEHLI, 1954
CЕМЕЙСТВО MONTICULIFERIDAE MUIR-WOOD ET COOPER, 1960
ПОДСЕМЕЙСТВО GIGANTOPRODUCTINAE MUIR-WOOD ET COOPER, 1960
Триба Semiplanellini Pakhnevich, tribe nov.
Диагноз. Раковины средних и крупных размеров. Радиальная скульптура в виде многочисленных сближенных ребер. Тонкие иглы располагаются на обеих створках. Замочный отросток имеет несколько лопастей или, в результате редукции лопастей, становится цельным. Срединная септа в спинной створке хорошо развита. Парные аддукторы располагаются друг за другом. Передние аддукторы удлинены. Брахиальные петли проходят вдоль передних аддукторов. Брахиальные конусы слабо развиты.
Родовой состав. Semiplanella, Beleutella и, возможно, Talasoproductus. Нижний карбон (середина визейского яруса – серпуховский ярус).
ЗАКЛЮЧЕНИЕ
Таким образом, брахиоподы, относящиеся к родам Semiplanella, Beleutella и, возможно, Talasoproductus являются небольшой, самостоятельной ветвью гигантопродуктид трибы Semiplanellini tribe nov. У рода Semiplanella, в связи с большой скрученностью раковины, произошел поворот парных мускулов-аддукторов, в результате чего они заняли почти параллельное горизонтальное положение относительно друг друга. За этим процессом произошел и поворот брахиальных петель. Их восходящие ветви проходили вдоль удлинившегося переднего аддуктора и располагались около аддукторов. Причиной таких кардинальных изменений стала сильная скрученность раковины. На ее фоне передний край (шлейф) высоко поднимается над поверхностью дна. За счет образовавшегося утолщения около замочного края спинной створки сформировался перегиб, что способствовало ориентации переднего края раковины высоко над поверхностью дна. Удлинение мускульного поля аддукторов, укорочение длины мускулов также способствовало высокому поднятию переднего края над субстратом. Почти вертикальное положение раковины было у рода Beleutella. В данном случае это было достигнуто за счет увеличение центральной лопасти замочного отростка и его сильного удлинения, а также выпрямления утолщенной макушки брюшной створки, которая стала опорой для тяжелой раковины.
Список литературы
Афанасьева Г.А. Брахиоподы отряда Chonetida (история развития, функциональная морфология, филогенез и система). М.: Наука, 1988. 123 с. (Тр. ПИН АН СССР. Т. 228).
Литвинович Н.В., Аксенова Г.Г., Разина Т.П. Стратиграфия и литология отложений нижнего карбона западной части Центрального Казахстана. М.: Недра, 1969. 448 с.
Литвинович Н.В., Воронцова Т.Н. Гигантоидные брахиоподы СССР, их распространение и стратиграфическое значение. М.: Наука, 1991. 61 с.
Сарычева Т.Г., Легран-Блан М. Семейство Semiplanidae (Brachiopoda), его состав и развитие // Палеонтол. журн. 1977. № 2. С. 70–82.
Сарычева Т.Г., Сокольская А.Н. Определитель палеозойских брахиопод Подмосковной котловины. М.: Изд-во АН СССР, 1952. 272 с. (Тр. ПИН АН СССР. Т. 38).
Brunton C.H.C., Lazarev S.S., Grant R.E., Jin Yu-gan. Productidina // Treatise on Invertebrate Paleontology. Part H. Brachiopoda. V. 3. Boulder, Colorado; Lawrence, Kansas: Geol. Soc. America and Univ. Kansas Press, 2000. P. 424–643.
Gröber P. Carbon und Carbonfossilien des nördlichen und zentralen Tian-Schan // Aus den wissenschaftlichen Ergebnissen der Merzbacherschen Tian-Schan-Expedition. München: Verl. K.B. Akad. Wiss., 1909. S. 341–384.
Legrand-Blain M. Les Gigantoproductides (Brachiopodes) du Sahara Algerien III – Semiplanidae Viseens et Namuriens // Bull. Soc. Hist. Natur. Afrique du Nord. 1980. T. 69. Fasc. 1, 2. P. 3–86.
Racheboeuf P.R. Suborder Chonetidina // Treatise on Invertebrate Paleontology. Part H. Brachiopoda. V. 2. Boulder, Colorado; Lawrence, Kansas: Geol. Soc. America and Univ. Kansas Press, 2000. P. 362–423.
Żakowa H. Brachiopods of the family Semiplanidae Sarytcheva, 1960 from the Upper Visean of Poland // Biul. Inst. Geol. 1986. V. 355. Geology of Poland. V. VII. P. 49–70.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал