

Палеонтологический журнал, 2019, № 2, стр. 94-105
Уточнение интерпретации пыльцевых зерен Wodehouseia spinata Stanley из позднего маастрихта Китая
М. В. Теклева a, *, С. В. Полевова b, Е. В. Бугдаева c, В. С. Маркевич c, Сунь Ге d
a Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
b Московский государственный университет им. М.В. Ломоносова
Москва, Россия
c Федеральный научный центр биоразнообразия наземной биоты Восточной Азии ДВО РАН
Владивосток, Россия
d Колледж палеонтологии, Шэньянский педагогический университет
Шэньян, Китай
* E-mail: tekleva@mail.ru
Поступила в редакцию 25.05.2018
После доработки 30.09.2018
Принята к публикации 15.10.2018
Аннотация
Исследованы дисперсные пыльцевые зерна Wodehouseia spinata Stanley неизвестного ботанического родства из маастрихтских отложений Дальнего Востока с помощью светового, сканирующего и трансмиссионного электронных микроскопов. Растения, продуцировавшие данный тип пыльцевых зерен, возможно, были водными или околоводными, приспособленными к резким сменам водного режима в течение сезона вегетации. Характер скульптуры и ультраструктуры спородермы позволяет предполагать участие насекомых в процессе опыления. Гребень и эндэкзина неравномерной толщины могли служить для выполнения гармомегатной функции. Наиболее вероятным для Wodehouseia представляется квадратный тип тетрады. Характер инфратектума предполагает принадлежность Wodehouseia к высокоорганизованным двудольным.
ВВЕДЕНИЕ
Дисперсные пыльцевые зерна из верхнемеловых отложений США и России, характеризующиеся шиповатой поверхностью, четырьмя попарно расположенными порами и гребневидной структурой, опоясывающей пыльцевое зерно по периметру, были описаны практически одновременно в 1961 г. Э.А. Стэнли (Stanley, 1961) как Wodehouseia Stanley, С.Р. Самойлович (1961) как Kryshtofoviana Samoilovitch и Regina Samoilovitch, и А.Ф. Хлоновой (1961) как Deplexipollis Chlonova. Приоритетным является родовое название Wodehouseia с типовым видом Wodehouseia spinata Stanley.
Самойлович (1961), изучив пыльцу своеобразного строения из верхнего мела–палеоцена Сибири, выделила надгруппу Kryshtofoviacites с группами Kryshtofoviana, Singularia Samoilovitch, Regina и Azonia Samoilovitch. Пыльцу из групп Kryshtofoviana и Regina отличало наличие латеральной оторочки, отсутствующей у представителей Singularia и Azonia. Хлонова (1961) для подобной пыльцы предложила использовать название морфологического типа oculata.
Разные виды вудхаузии выделены на основании отличия в размерах, форме апертур (поровидных или бороздовидных), степени развитости гребня, деталей скульптуры. Виды Azonia, другого представителя морфотипа oculata, образно говоря, представляющие собой “тело” вудхаузии без гребня, также относятся рядом палинологов к вудхаузии, утратившей гребень (см. обзор в: Wiggins, 1976). Для окончательного разрешения этого вопроса необходимо детальное изучение морфологии и ультраструктуры пыльцевых зерен Azonia. В.Ф. Тарасевич (Ботанический ин-т им. В.Л. Комарова РАН, С.-Петербург) было проведено исследование ультраструктуры пыльцевых зерен Azonia с помощью ТЭМ, но результаты еще не опубликованы (устн. сообщ.). В настоящий момент опубликованных результатов таких исследований не существует, поэтому данная проблема остается пока нерешенной.
Узкий временной диапазон существования и широкое географическое распространение такой легко распознаваемой пыльцы, как Wodehouseia, обуславливают важность этого рода для стратиграфических целей. Например, для российского Дальнего Востока В.С. Маркевич (1983, 1995) ввела этот таксон в практику палиностратиграфических работ, выделив палинозоны Wodehouseia spinata – Aquilapollenites subtilis (маастрихт), Orbiculapollis lucidus – Wodehouseia avita (поздний маастрихт), Wodehouseia fimbriata – Ulmoideipites krempii (ранний даний). К сожалению, до сих пор пыльцевые зерна такого типа не были описаны in situ и не установлены филогенетические связи растений, продуцировавших подобную пыльцу. Это значительно ограничивает потенциал находок этого палинотипа, оставляя пока в основном прикладное использование известных данных для целей стратиграфии. Очевидно, что необходимо детализировать интерпретацию находок пыльцы Wodehouseia и попытаться выявить ее возможные родственные связи и экологические предпочтения.
Хлонова (1961) детально рассмотрела возможные варианты ориентировки осей пыльцевых зерен Wodehouseia и также предположила их сходство с пыльцевыми зернами современных Impatiens L. и Jollydora Pierre ex Gilg. Самойлович (1961) также обсуждала ориентировку осей пыльцевых зерен Wodehouseia, но высказала иную точку зрения, сблизив этот род с современными Acanthaceae (см. детальный обзор в: Leffingwell et al., 1970). Обе упомянутые работы базируются на изучении пыльцевых зерен в световом микроскопе. Для более точной интерпретации необходимо всестороннее изучение морфологии и ультраструктуры пыльцевых зерен Wodehouseia. Подобное исследование с помощью светового (СМ), сканирующего (СЭМ) и трансмиссионного (ТЭМ) электронных микроскопов было выполнено для Wodehouseia spinata из маастрихта США (Leffingwell et al., 1970). В результате был сделан вывод, что данные СЭМ и ТЭМ опровергают сближение Wodehouseia как с Balsaminaceae, так и с Acanthaceae. Но вопрос относительно возможного родства Wodehouseia так и остался открытым.
В настоящей работе мы изучили пыльцевые зерна самого распространенного вида Wodehouseia spinata из маастрихта Зейско-Буреинского бассейна Приамурья, сравнили наши данные с таковыми, полученными Х. Лефингвел с соавт. (Leffingwell et al., 1970), обсудили новые сравнения с семенными растениями и идеи относительно возможной экологической ниши материнских растений.
Авторы благодарны рецензентам и научному редактору за ценные замечания по улучшению текста статьи. Исследование поддержано грантом РФФИ № 17-04-01094 (МТ, СП) и грантом РФФИ № 17-04-01582 (ВМ, ЕБ).
МАТЕРИАЛ И МЕТОДЫ
Материал отобран из формации Фурао, скважины № XHY2008, Зейско-Буреинского бассейна (Markevich et al., 2011). Проба взята из керна скважины № XHY2008 с глубины 23.25 м из верхнемаастрихтских отложений (Markevich et al., 2011). Исследованные пыльцевые зерна принадлежат к спорово-пыльцевому комплексу Aquilapollenites stelckii–Pseudointegricorpus clarireticulatum маастрихтского возраста. Содержание пыльцевых зерен Wodehouseia в комплексе очень низкое, чаще всего доли процента, и только в изученном образце оно составляет 2.4%. Номер коллекции XHY2008, она хранится в Исследовательском центре палеонтологии и стратиграфии Цзилиньского ун-та, г. Чанчунь, КНР (Research Center of Paleontology & Stratigraphy Jilin University, Changchun, China).
Дисперсные пыльцевые зерна Wodehouseia spinata идентифицировались под световым микроскопом Olympus CX41, переносились поодиночке на новое предметное стекло в каплю глицерина. Полученный таким образом монопрепарат фотографировали при увеличении ×100 с помощью светового микроскопа Zeiss Axioplan-2 с цифровой камерой Leica DFC-420. Далее покровное стекло снимали, пыльцевое зерно промывали в капле спирта и переносили на кусочек фотопленки, прикрепленной на лак для ногтей эмульсионной стороной вверх на столик для СЭМ. Столик для СЭМ с перенесенными на него пыльцевыми зернами напылялся золотом и изучался с помощью СЭМ TESCAN VEGA-II XMU (ускоряющее напряжение 30 кВ). Для изучения в ТЭМ часть пыльцевых зерен была снята со столика СЭМ и перенесена в смесь эпоксидных смол согласно методике, описанной Н.Е. Завьяловой с соавт. (2018). С полученных блоков сделаны ультратонкие срезы с помощью ультратома LKB Leica UC6. Часть срезов дополнительно контрастировались цитратом свинца и уранил ацетатом и изучались с помощью ТЭМ Jeol 100 B и Jeol 1011 (ускоряющее напряжение 80 кВ). Другая часть срезов была изучена без дополнительного контрастирования. Пыльцевые зерна также исследовались с помощью конфокального микроскопа LSM 780 (результаты и методика описаны в статье О.А. Гавриловой с соавт. (Gavrilova et al., 2018). Измерения пыльцевых зерен сделаны в СМ и СЭМ. Всего изучено 20 пыльцевых зерен. При описании пыльцевых зерен были использованы термины, предложенные в руководстве по морфологии и ультраструктуре палиноморф М. Хессе с соавт. (Hesse et al., 2009).
В работе применялось оборудование центра коллективного пользования ПИН РАН (СМ и СЭМ) и Биологического фак-та МГУ им. М.В. Ломоносова (ТЭМ).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
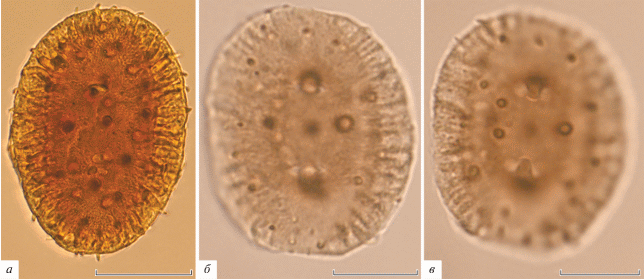
СМ. Пыльцевые зерна эллипсоидальные, средних размеров, 31.9 (27.6–38.9) × 23.9 (19.7–27.0) µm, уплощенные, с гребнем по периметру пыльцевого зерна. Апертурный тип поровый или короткобороздный, апертуры простые, в числе четырех, видны как щелевидные структуры, 4.5 (3.7–5.5) × 1.3 (0.8–2.1) µm, вытянутые по короткой оси пыльцевого зерна, сближенные попарно на противоположных сторонах пыльцевого зерна (рис. 1, а). Скульптура поверхности шиповатая, шипы трех размерных классов (рис. 1, б, в). Крупные шипы располагаются по одному по краям каждой поры и в центральной части уплощенного пыльцевого зерна, мелкие многочисленные шипы развиты на периферии уплощенного по длинной оси пыльцевого зерна, шипы среднего размера располагаются между крупными и мелкими более или менее равномерно. Оболочка неравномерная по толщине, с участками максимальной толщины в области гребня и с некоторым утончением этих участков на закругленных концах эллипсоидального пыльцевого зерна, двуслойная, внутренний слой состоит из хорошо заметных столбиковидных элементов (рис. 1, а).
Рис. 1.
Wodehouseia spinata Stanley, СМ: a – совмещенные фото, сделанные на разном фокусе; б, в – одно и то же пыльцевое зерно с разным фокусом, видны поры, разноразмерные шипы и столбики инфратектума. Длина масштабной линейки 10 мкм.
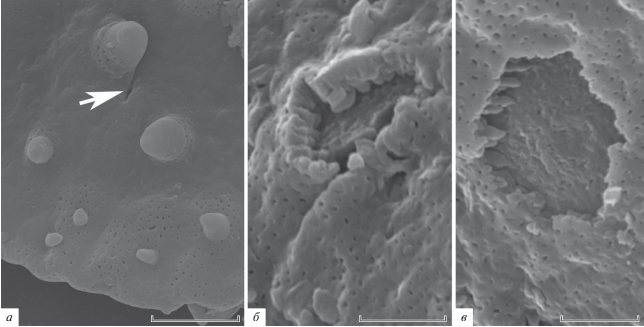
СЭМ. Поверхность шиповатая, перфорированная (рис. 2, а–в; 3, а). Шипы крупные (2.7–3.4 µm, основание шипа 2.1–3.6 µm, в количестве 8–12), средние (1.3–2.0 µm, основание шипа 1.1–1.7 µm, около 20) и мелкие (0.6–2.0 µm, основание шипа 0.3–0.8 µm, около 40–50). Перфорации располагаются между шипами, часто в центральной части сплющенного пыльцевого зерна перфораций меньше.
Рис. 2.
Wodehouseia spinata Stanley, СЭМ: а, б – общий вид пыльцевого зерна, видна пора и шипы разных размеров; в – участок поверхности при большем увеличении, видны перфорации, в том числе в основании шипов среднего размерного класса. Длина масштабной линейки: а, б – 10 мкм; в – 2 мкм.

Шипы конусовидные, крупные и средние по размерам шипы с относительно широким основанием и более узкой верхней половиной, мелкие шипы приблизительно одинакового диаметра по всей высоте, с притупленной верхушкой, основания крупных и зачастую средних шипов перфорированы. На сколах оболочки виден покров и инфратектум из ветвящихся столбиковидных элементов (рис. 3, б, в). Апертуры 4.6 (3.5–5.6) µm длиной.
Рис. 3.
Wodehouseia spinata Stanley, участки поверхности при большом увеличении, СЭМ: а – видна пора (cтрелка), перфорированность экзины и шипы трех размерных классов с перфорациями в основании крупных и средних шипов; б, в – сколы оболочки, видна внутренняя структура экзины. Длина масштабной линейки: а – 5 мкм; б, в – 2 мкм.
ТЭМ. Экзина неравномерная по толщине в пределах пыльцевого зерна, от 0.6 до 1.5 µm, представлена тремя слоями: менее электронно-плотными покровом и инфратектумом и более электронно-плотным внутренним слоем, вероятно, соответствующим эндэкзине (рис. 4, а–г). Покров тонкий, 0.18 (0.12–0.23) µm, относительно равномерный по толщине по всему периметру пыльцевого зерна. Инфратектум представлен высокими и иногда разветвленными столбиками, 0.49 (0.31–0.7) µm высотой и 0.25 (0.17–0.39) µm шириной, в латеральных областях располагаются крупные полости между инфратектумом и внутренним слоем (рис. 4, а–д; 5, а–в). Внутренний слой неравномерной толщины, 0.21 (0.12–0.27) µm в латеральных областях и 0.67 (0.5–0.82) µm в остальных областях пыльцевого зерна, выглядит гомогенным, с просветами (рис. 6); возможно, истинная природа этого слоя – ламеллятная. Утончения внутреннего слоя приурочены к латеральным областям. Шипы сплошные, в основном гомогенные (рис. 4, а–г). Поры представляют собой внезапный перерыв во всех слоях экзины; поры располагаются попарно, напротив друг друга на противоположных сторонах пыльцевого зерна (рис. 4, а; 5, а).
Рис. 4.
Wodehouseia spinata Stanley, срезы через пыльцевые зерна, общий вид, ТЭМ: а – срез прошел через обе поры (звездочки), видна эндэкзина разной толщины, столбиковый инфратектум и средние шипы (головки стрелок); б – срез прошел через одну из пор (звездочка), видна эндэкзина разной толщины, столбиковый инфратектум, полости в экзине в области гребня (стрелка) и крупный шип (головки стрелок); в – срез прошел в области между порами (в центральной части пыльцевого зерна), хорошо видна эндэкзина разной толщины, столбиковый инфратектум, полости в экзине в области гребня (стрелка) и крупный, два средних и один мелкий шип (головки стрелок); г – скошенный срез, прошедший через обе поры и крупные шипы, приуроченные к ним (головки стрелок); д – парадермальный срез, срезаны столбики инфратектума (стрелка) и средние и мелкие шипы (головки стрелок). Длина масштабной линейки: а – 1 мкм; б–г – 2 мкм; д – 5 мкм.
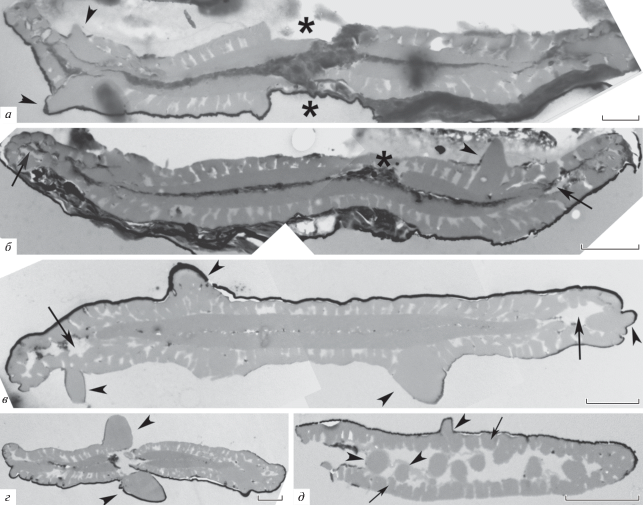
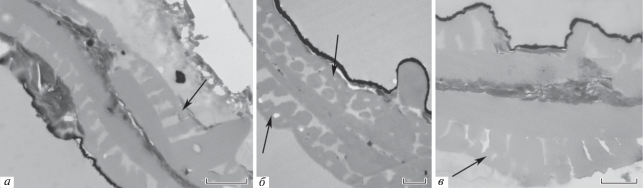
Рис. 5.
Wodehouseia spinata Stanley, участки оболочки при большом увеличении, ТЭМ: а – срез через пору, в которой слои экзины отсутствуют; также заметна эндэкзина разной толщины, столбики инфратектума, кое-где заметны разветвленные столбики (стрелка) и крупный шип; б – скошенный срез, инфратектум с разветвленными (стрелки) столбиками и средний шип; в – срез через край поры, эндэкзина присутствует в области поры, заметны разветвленные столбики (стрелка). Длина масштабной линейки: а – 1 мкм; б – 0,5 мкм; в – 0.67 мкм.

ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ ИССЛЕДОВАНИЯ
Необычная и сложноустроенная оболочка, как по морфологии, так и по ультраструктуре экзины, подразумевает высокую степень специализации материнского растения и приспособления его к условиям произрастания. На настоящий момент в составе рода описано 26 видов (http://paleobotany.ru/palynodata). До сих пор пыльцевые зерна Wodehouseia были найдены только в дисперсном состоянии и, таким образом, о возможной систематической принадлежности и экологии продуцировавшего их растения можно судить только на основании данных по морфологии и ультраструктуре спородермы, характеру вмещающей породы и сопутствующим растениям.
Находки пыльцевых зерен Wodehouseia на Дальнем Востоке
Считается, что Wodehouseia впервые появилась в туроне Хатангской впадины на севере Сибири (Синопсис …, 1980), однако, в Зейско-Буреинском бассейне Приамурья мы зафиксировали первые находки этой пыльцы в кундурской свите сантон-кампанского возраста. Была найдена единичная W. aspera (Samoilovitch) Wiggins в местонахождении 1808, в котором обнажаются сантонские озерные отложения. В раннекампанском местонахождении 15 междуречья Мутная-Удурчукан обнаружены единичные W. aspera, W. spinata и W. gracile Samoilovitch. Здесь же собраны, наряду с фитофоссилиями, остатки лимнофауны – остракоды, двустворки, рыбы, существовавшие в условиях обширного озера (Бугдаева и др., 2001; Маркевич и др., 2005). Наиболее разнообразной в этом бассейне пыльца Wodehouseia становится в маастрихте; ее видовой состав включает W. aspera, W. spinata, W. gracile, W. stanleyi Srivastava и W. wigginsii Frederiksen. На границе мела и палеогена эти таксоны исчезают, в дании найден только один вид – W. fimbriata Stanley.
В Приморье редкие пыльцевые зерна Wodehouseia sp. появляются позже. Первые ее находки сделаны в раннекампанской осадочной толще бассейна р. Алчан, которая представлена песчаниками, алевролитами, аргиллитами с прослоями углей; она содержит многочисленные растительные остатки (Маркевич, 1995). Фациально находки пыльцы приурочены к нормально-осадочным отложениям (угли, алевролиты, аргиллиты, песчаники), по всей видимости, озерно-речного происхождения, зажатым между вулканическими толщами.
На Сахалине редкие пыльцевые зерна Wodehouseia появляются в среднебыковской подсвите раннекампанского возраста морского генезиса, содержащей аммониты и двустворки. Выше по разрезу эта пыльца становится разнообразнее; она выявлена в палиноспектрах, выделенных из морских отложений красноярковской свиты позднекампанского-маастрихтского возраста.
На о-ве Юрий Малой Курильской гряды три вида Wodehouseia [W. spinata, W. aspera и редко встречающийся (1–2%) вид W. avita Wiggins] были обнаружены в палинокомплексе алевролитов юрьевской толщи пограничного маастрихт-датского возраста (Красилов и др., 1988). Здесь же встречены остатки растений, фораминифер, мелких двустворок, гастропод и морских ежей, что свидетельствует о прибрежно-морском генезисе отложений.
На протяжении сантона-дания на юге российского Дальнего Востока в палинофлорах Wodehouseia встречается единично, что может говорить или о редкой распространенности продуцирующих ее растений, или об удаленных от захоронений местах их обитания. Последнее кажется более предпочтительным, ввиду встречаемости этой пыльцы на столь обширной территории в фациях различного генезиса (морские, озерные, речные, вулканические).
Мы полагаем, что материнские растения Wodehouseia произрастали на склонах. Если исключить для них анемофильный характер опыления и, следовательно, транспортацию пыльцы воздушными потоками в места захоронения, можно предположить ее доставку в низины водным путем. Возможно, некоторые продуценты Wodehouseia произрастали в водной или околоводной среде.
Что дает морфология и ультраструктура Wodehouseia для понимания “образа жизни” материнского растения?
Существует два основных способа опыления – ветром и насекомыми – и водные и околоводные покрытосеменные растения в этом отношении не исключение (Manabale, 1968). Среди них значительное количество стали водными вторично, и часть видов сохранила свои “сухопутные” пыльцевые зерна, а адаптация к водным условиям отразилась в строении вегетативных органов (Manabale, 1968). Поэтому далеко не у всех наблюдается строение спородермы, характерное для водных растений, например, с редуцированной экзиной и упрощенным строением оболочки.
В нашем случае строение спородермы как раз нетипично для “настоящих” водных или околоводных растений – шиповатая скульптура, хорошо развитая эктэкзина, апертуры, занимающие небольшую часть общего периметра пыльцевого зерна. Очевидно, что, если материнское растение действительно было приурочено к какому-либо водоему, адаптация происходила, главным образом, за счет вегетативной части.
Развитая скульптура, представленная как надпокровными образованиями – шипами, так и сеткой, у современных растений всегда связана с энтомофилией (напр., Tanaka et al., 2004), тогда как характер скульптуры (надпокровные элементы – шипы, палочки, булавы или полупокровные элементы – сетка, перфорации) связан скорее с филогенетическими (таксономическими) группами. Нам представляется логичным утверждение, что разный размер шипов также должен быть связан с приспособлением к определенным условиям произрастания материнского растения – например, как способ увеличения вероятности нужной ориентации пыльцевого зерна при попадании на рыльце и дальнейшего прорастания (крупные шипы и высокий гребень создают общую архитектуру пыльцевого зерна, которому в результате удобнее “расположиться” своими двумя порами к рыльцу). Также крупные шипы около поры могут при частичной дегидратации прикрывать еe для уменьшения испарения в результате того, что более тонкие участки оболочки вокруг апертуры прогибаются. Последнее предположение, однако, ставит под сомнение околоводный образ жизни материнского растения, либо подразумевает периодически подсыхающий водоем. Крупные скульптурные элементы (в данном случае – шипы) на поверхности спородермы создают некоторую прослойку воздуха вокруг апертуры, не мешающую пыльцевому зерну прорастать, но, возможно, препятствуя тому, чтобы пыльцевое зерно утонуло в воде. Мы не знаем, была ли у пыльцевых зерен Wodehouseia необходимость распространения по поверхности воды, лежало ли рыльце материнского растения на воде или располагалось над водой, однако, судя по морфологии пыльцевых зерен Wodehouseia, рыльце не было погружено в воду. У пыльцевых зерен некоторых современных растений (напр., у представителей Plumbaginaceae, Valerianaceae) шипы и палочки привязаны к апертуре только в случае, если они расположены на оперкулюме. При этом оперкулюм может быть шиповатым, а мезокольпиум струйчатым или струйчато-перфорированным, как, например, у Aesculus hippocastanum L. Представители Plumbaginaceae и Valerianaceae не считаются классическими околоводными видами, а скорее являются обитателями засоленных местообитаний семи- и аридного климата.
Наличие гребня и соответствующих полостей в эктэкзине и уплощение пыльцевого зерна можно также рассматривать как возможное приспособление к опылению ветром или водой, аналогично мешкам у хвойных ветроопыляемых растений. Хотя не исключена возможность, что уплощенность пыльцевого зерна также способствует определенной ориентировке при попадании его на рыльце. Полости в спородерме обычно несут гармомегатную функцию, а у пыльцевых зерен покрытосеменных растений часто служат для вмещения веществ, способствующих энтомофилии и/или прикреплению к рыльцу.
Апертурный тип вудхаузии также не является типичным для водных или околоводных растений. Пыльцевые зерна вудхаузии описываются как поровые, но в принципе, эти апертуры могут быть названы и бороздками (короткими бороздами). Похожий пример наблюдается, например, у Myriophyllum L., чей апертурный тип описывается то как поровый (Praglowski, 1970; www.paldat.org), то как бороздный (Alwadie, 2008). Все это семейство (Haloragaceae) представлено водными растениями, однако пыльца его представителей характеризуется мелкошиповатой скульптурой и порами (=бороздками) с ободком и утолщенной нэкзиной. Отметим, что, как и у вудхаузии, бороздки этих пыльцевых зерен ориентированы меридионально (то есть, по меньшему диаметру), пыльцевые зерна сплющенные.
Ультраструктура оболочки, с одной стороны, дает основания полагать, что материнское растение было приспособлено к энтомофилии – большая площадь свободного пространства в эктэкзине обеспечивалa так называемую витализацию экзины. Лефингвел с соавт. (Leffingwell et al., 1970) также отмечали наличие включений в пространствах между столбиками. Однако надо иметь в виду, что, например, у хвойных ячеистость тоже была сильно развита и использовалась для создания воздушных мешков и адаптации к анемофилии, выполнению гармомегатной функции и ориентации на опылительной капле. Была также высказана гипотеза, что мешки способствуют более быстрому забиванию желудка насекомого и, таким образом, меньшему потреблению пыльцевых зерен. Подобная гипотеза, однако, нуждается в подтверждении фактическими данными.
Образование полостей в инфратектуме или аналогичном слое происходило во многих группах независимо, даже у спор плауновидных (Tryon, Lugardon, 1990). Возможно, это связано с водным (болотным) местообитанием, как у Trapa L., или не связано с подобным местообитанием, как у Centaurea L.
Эндэкзина пыльцевых зерен Wodehouseia неравномерной толщины, с утонченными участками в области гребня. Ранее (Leffingwell et al., 1970) она была описана как гомогенная, однако, мы наблюдали просветы в толще этого слоя (рис. 6). Возможно, структура эндэкзины не позволяла выдерживать сильные изменения объема пыльцевого зерна, в связи с чем и наблюдается неравномерная утолщенность этого слоя. Отсутствие подстилающего слоя, возможно, обуславливало большую “гибкость” слоев экзины относительно друг друга. Неравномерности в высоте слоев экт- и особенно эндэкзины, вероятно, способствовали “стабилизации” оболочки, имеющей многочисленные перепады в толщине и структурах (гребень, полости, шипы разного размера, поры).
Сравнение Wodehouseia с пыльцевыми зернами голосеменных
Шиповатая или шипиковатая скульптура экзины крайне редко встречается у пыльцевых зерен голосеменных растений [напр., Gnetum L. (Tekleva, 2016), Cyclusphaera Elsik (Del Fueyo et al., 2012)]. Пыльцевые зерна Wodehouseia сравнивали с пыльцевыми зернами голосеменных из-за наличия мешковидных структур (= гребня). Действительно, строение оболочки в латеральных утолщенных областях в значительной степени напоминает таковое у мешковых пыльцевых зерен голосеменных – та же полость между инфратектумом и нижележащими слоями экзины. Морфологически это выражено не воздушными мешками, характерными для многих хвойных, а уплощенной гребневидной структурой. Сходная по степени развития и выраженности в морфологии структура наблюдается у ископаемых дисперсных пыльцевых зерен Cryptosacciferites pabularis Krassilov et Tekleva (Krassilov et al., 2003) и Zolerella tricolpata Scheuring (Scheuring, 1978), но у этих видов данная структура располагается не по всему периметру пыльцевого зерна, а лишь в латеральных областях, подобно мешкам у двухмешковых пыльцевых зерен. У Cryptosacciferites pabularis и Zolerella tricolpata инфрактектум ячеистый, ячеи в основном удлиненные и располагаются в один ряд, в целом напоминая столбиковидную структуру на срезах, однако, в отличие от изученного нами вида, стенки ячей гораздо тоньше, чем столбики инфратектума Wodehouseia. Также, Cryptosacciferites pabularis и Zolerella tricolpata характеризуются гладкой или шероховатой поверхностью экзины и тремя бороздами, в отличие от шиповатых поровых пыльцевых зерен Wodehouseia.
Поровый апертурный тип, в целом, характерен только для покрытосеменных растений, хотя у некоторых таксонов ископаемых голосеменных (напр., Cyclusphaera, Classopollis Pflug, Cupressoideae, Taxodiaceae) известны поровидные структуры (Zavialova et al., 2010; Del Fueyo et al., 2012; обзор в: Завьялова, Соколова, 2017). Апертурные области у голосеменных с поровидными структурами представлены сильно утонченной эктэкзиной, часто редуцированной до подстилающего слоя, и эндэкзиной разной степени развитости. У пыльцевых зерен с дифференцированной апертурной областью (наличием оперкулюма, папиллы или поры) наблюдается вариабельность строения эндэкзины: утолщение (область оперкулюма у Cyclusphaera: Zavada, 1992; Del Fueyo et al., 2012) или исчезновение (область папиллы или поры у таксодиевых) (напр., Kurmann, 1990, 1991).
Сравнение Wodehouseia с пыльцевыми зернами покрытосеменных
Апертурный тип и скульптура поверхности. Необычный апертурный тип, характерный для Wodehouseia, довольно редко встречается среди современных покрытосеменных, и среди них наиболее сходное строение имеют пыльцевые зерна рода Impatiens, в составе которого есть и околоводные виды. Однако скульптура и ультраструктура спородермы пыльцевых зерен представителей Impatiens довольно сильно отличаются от таковых у Wodehouseia.
Шиповатая скульптура поверхности спородермы встречается по филетическому древу цветковых неравномерно. Среди палеотрав шиповатые формы есть у Nymphaeales (Taylor et al., 2015), Lauraceae (Van Der Merwe et al., 1990) и Piperaceae (Ligong, Hanxing, 1998); среди однодольных у Alismatales (Furness, 2013), редко у Liliaceae (Kosenko, 1999) и Arecaceae (Sannier et al., 2009); среди примитивных двудольных только у Ranunculales (Blackmore et al., 1995). У продвинутых двудольных шипы и шипики встречаются редко, в основном, у представителей Saxifragales, Myrtales, Rosaceae, Cucurbitaceae и Malvaceae (Christensen, 1986). Гораздо чаще шипики и шипы встречаются у Caryophyllales (есть околоводные среди Droseraceae), Dipsacales и Asterales (Полевова, 2004; Теклева и др., 2011).
Среди палеотрав, например, у Nymphaeaceae, шипики бывают при сулькатном апертурном типе, в то время как у представителей Laurales и Piperales обычны безапертурные шиповатые пыльцевые зерна. Многие представители Piperaceae предпочитают переувлажненные местообитания (Кирпичников, Комар, 1980; Tebbs, 1993).
Следует отметить, что шиповатыми формами с порами или короткими (“поровидными”) бороздками обладают пыльцевые зерна Ranunculales, Cucurbitales, Malvales, Caryophyllales и Dipsacales (Erdtman, 1966). Из них последние самые интересные, т.к. у них шипы могут быть разных размерных классов, часто оперкулюм с палочками и ободок у поры тоже покрыты шипами. Однако околоводные формы есть только у Caprifoliaceae (Godfrey, Wooten, 2011). Неодинаковая группировка шипов наблюдается, например, у Lemna L., но этот род характеризуется однопоровыми пыльцевыми зернами (Alwadie, 2008).
У пыльцевых зерен покрытосеменных четыре апертуры встречаются нечасто. Если не рассматривать таксоны с четырехбороздными и четырехборозднооровыми пыльцевыми зернами, то для анализа остаются представители семейств Bromeliaceae, Betulaceae, Campanulaceae, Haloragaceae, Apocinaceae, Malpigiaceae (www.paldat.org). Среди них шипы или шипики характерны только для пыльцевых зерен представителей Betulaceae, Campanulaceae (Золала и др., 2011) и Haloragaceae (Praglovsky, 1970). Некоторые виды Lobelia (Campanulaceae) являются водными или околоводными растениями, но для них характерны трехбороздные пыльцевые зерна и отсутствие шиповатой скульптуры спородермы (Тахтаджан, 1981; Lammers, 2007; Золала и др., 2009).
Ультраструктура спородермы. Пыльцевые зерна с усложненным покровом или ветвистыми столбиками и неравномерной по толщине экзиной с экт- и эндэкзиной встречаются только у высокоорганизованных Asterids: Apiaceae, Asteraceae, Campanulaceae, Caprifoliaceae. Если у Asteraceae с Caprifoliaceae сильнее утолщена оболочка в мезокольпиумах (Polevova, 2006), то у многих других, в том числе у Apiaceae (Shu, She, 2001), утолщена экзина вокруг апертур, например, у Hacquetia epipactis DC. (www.paldat.org), Trapa (Zetter, Ferguson, 2001) и Morina L. (Blackmore, Cannon, 1983).
Ультраструктура пор Wodehouseia ближе к таковой у покрытосеменных с просто устроенной порой (Fumariaceae; Romero et al., 2003), что выражается в отсутствии заметного изменения характера слоев оболочки вблизи апертурной области, где все слои экзины резко прерываются. С другой стороны, нельзя исключить возможность присутствия сложноустроенной поры у вудхаузии, выраженной в строении интины, которая в ископаемом состоянии не сохранилась.
Возможное расположение пыльцевых зерен в тетраде. Если считать пару пор у Wodehouseia за концы одной апертуры, то две сложные апертуры располагаются каждая на своем полюсе (самый невероятный вариант), либо две борозды опоясывают пыльцевое зерно по экватору, а на полюсах располагаются гребни, либо две борозды идут по меридиану, как и гребень, который на полюсах становится немного меньше (рис. 7, а).
Рис. 7.
Модели расположения пыльцевых зерен Wodehouseia spinata Stanley в тетрадах: а – полярная ось вдоль длинной оси, апертурных областей две, в каждой по две поры, тетраэдрическая (левее) и квадратная (правее) тетрады; б – полярная ось вдоль короткой оси, четыре бороздки в экваториальной области, тетраэдрическая (левее) и квадратная (правее) тетрады; в – полярная ось вдоль короткой оси, апертурных областей две, на каждом полюсе по две поры, тетраэдрические тетрады.

При экваториальном расположении пор кайма идет от полюса к полюсу между парами апертур, а в чередующимся промежутке между порами экзина тонкая (рис. 7, б). При полярном расположении пор у Wodehouseia ее пыльцевое зерно становится билатерально-симметричным, по экватору экзина сильно утолщена в виде каймы, а на полюсах располагаются по две поры с шипами по краю (рис. 7, в). Также возможны другие варианты расположения пыльцевых зерен в тетрадах квадратного типа (рис. 8). Нам представляется самым логичным квадратный тип тетрад; при этом не возникает “уродств” в архитектуре пыльцевых зерен вудхаузии. Это самый удобный тип для формирования двух или четырех апертур.
ВЫВОДЫ
Систематическое положение Wodehouseia остается загадочным. Полифациальный характер отложений Дальнего Востока, в которых встречаются характерные для этого рода пыльцевые зерна, позволяет предположить, что материнские растения могли слагать склоновые растительные сообщества, произрастая вдоль ручьев и рек. Усложненная скульптура поверхности и внутреннее строение спородермы указывают на несомненную принадлежность Wodehouseia покрытосеменным. Комплекс признаков спородермы затрудняет прямое сопоставление с каким-либо современным таксоном, хотя характер инфратектума указывает на принадлежность Wodehouseia к высокоорганизованным двудольным. Тип и строение апертуры, а также характер скульптуры поверхности (в том числе, разноразмерные шипы), скорее, обусловлены генетически, в то время как гребень и эндэкзина неравномерной толщины могли служить приспособлением к условиям обитания. Появление полостей в инфратектуме или аналогичных слоях наблюдается в нескольких неродственных группах семенных и споровых растений.
Необычный апертурный тип, характерный для пыльцевых зерен Wodehouseia, довольно редко встречается у современных покрытосеменных. Скульптура и ультраструктура спородермы резко отличают этот род от других сходных по апертурному типу родов покрытосеменных. Подобная ситуация отмечается и для остальных признаков Wodehouseia, аналоги которых по отдельности можно наблюдать у разных таксонов, но их уникальный для Wodehouseia комплекс более нигде не отмечен. Приходится согласиться с мнением Лефингвел и др. (Leffingwell et al., 1970) о том, что Wodehouseia, очевидно, представляет собой тупиковую ветвь эволюции. Существование Wodehouseia в течение более 20 миллионов лет на обширной территории Азии и Северной Америки может свидетельствовать о том, что материнские растения имели неплохой адаптационный потенциал, реализовавшийся в приспособлении к определенной экологической нише. Не исключено, что именно узкая специализация способствовала полному вымиранию этой линии.
Список литературы
Бугдаева Е.В., Маркевич В.С., Сорокин А.П., Болотский Ю.Л. Стратиграфия // Флора и динозавры на границе мела и палеогена Зейско-Буреинского бассейна. Владивосток: Дальнаука, 2001. С. 25–43.
Завьялова Н.Е., Соколова А.Б. Применение признаков морфологии и ультраструктуры пыльцевых зерен в систематике семейства Cupressaceae // Систематика и эволюционная морфология растений: Матер. конф., посвящ. 85-летию со дня рожд. В.Н. Тихомирова (31 января–3 февраля 2017 г., Москва). М.: МАКС, 2017. С. 168–171.
Завьялова Н.Е., Теклева М.В., Полевова С.В., Богданов А.Г. Исследование палинологических объектов методами электронной микроскопии. М.: РИПОЛ классик, 2018. 360 с. / Zavialova N., Tekleva M., Polevova S., Bogdanov A. Electron microscopy for morphology of pollen and spores. Moscow: RIPOL Classic Press, 2018. 360 p.
Золала Х.А., Полевова С.В., Северова Е.Э. Палиноморфология представителей семейства Колокольчиковые (Cаmpanulaceae Juss.) // Бюлл. МОИП. Отд. биол. 2009. Т. 114. № 1. С. 39–54.
Золала Х.А., Полевова С.В., Северова Е.Э. Особенности ультраструктуры спородермы поровых пыльцевых зерен представителей подсемейства Сampanuloideae (Сampanulaceae) // Ботан. журн. 2011. Т. 96. № 8. С. 1076–1084.
Кирпичников М.Э., Комар Г.А. Порядок Перцевые (Piperales) // Жизнь растений / Ред. А.Л. Тахтаджан. М.: Просвещение, 1980.Т. 5. № 1. С. 169–172.
Красилов В.А., Блохина Н.И., Маркевич В.С., Серова М.Я. Мел-палеоген Малой Курильской гряды (новые данные по палеонтологии и геологической истории). Владивосток: ДВО АН СССР, 1988. 140 с.
Маркевич В.С. Пыльца рода Wodehouseia Stanley 1961 // Палеоботаника и фитостратиграфия востока СССР / Ред. В.А. Красилов. Владивосток: ДВНЦ АН СССР, 1983. С. 80–84.
Маркевич В.С. Меловая палинофлора севера Восточной Азии. Владивосток: Дальнаука, 1995. 200 с.
Маркевич В.С., Головнева Л.Б., Бугдаева Е.В. Флористическая характеристика сантон-кампанских отложений Зейско-Буреинского бассейна (Приамурье) // Современные проблемы палеофлористики, палеофитогеографии и фитостратиграфии. Тр. Междунар. палеобот. конф. Москва, 17–18 мая 2005 г. Вып. 1. М.: ГЕОС, 2005. С. 198–206.
Полевова С.В. Особенности ультраструктуры шипиковатых пыльцевых зерен астровых (Asteraceae) и некоторых других семейств // Бюлл. МОИП. Отд. биол. 2004. Т. 109. № 1. С. 22–30.
Самойлович С.А. Kryshtofoviacites // Пыльца и споры Западной Сибири. Юра – палеоцен / Ред. С.Р. Самойлович, Н.Д. Мчедлишвили. Л., 1961. С. 230–243 (Тр. ВНИГРИ. Вып. 177).
Синопсис: ключевые таксоны пыльцы покрытосеменных (поздний мел-ранний палеоген). Основные местонахождения / Ред. Е.Д. Заклинская. М.: Наука, 1980. 95 с.
Тахтаджан А.Л. Порядок Колокольчиковые (Campanulales) // Жизнь растений / Ред. А.Л. Тахтаджан. М.: Просвещение, 1981. Т. 5. № 2. С. 447–461.
Теклева М.В., Северова Е.Э., Полевова С.В., Леунова В.М. Сопоставление пыльцевых зерен некоторых насекомоядных растений // Матер. XIII Росс. палинол. конф. ИГ Коми НЦ УрО РАН. 2011. Т. 1. С. 64–67.
Хлонова А.Ф. Споры и пыльца верхней половины верхнего мела восточной части Западно-Сибирской низменности // Тр. Ин-та геол. и геофиз. СО АН СССР. 1961. Вып. 7. С. 3–137.
Alwadie H.M. Pollen morphology of six aquatic angiosperms from Saudi Arabia // Asian J. Biol. Sci. 2008. V. 1. № 1. P. 45–50.
Blackmore S., Cannon M.J. Palynology and systematics of Morinaceae // Rev. Palaeobot. Palynol. 1983. V. 40. P. 207–266.
Blackmore S., Stafford P., Persson V. Palynology and systematics of Ranunculiflorae // Systematics and Evolution of the Ranunculiflorae / Eds. Jensen U., Kadereit J.W. Vienna: Springer, 1995. P. 71–82 (Plant Syst. Evol. Suppl. 9).
Christensen P.B. Pollen morphological studies in the Malvaceae // Grana. 1986. V. 25. № 2. P. 95–117.
Del Fueyo G.M., Archangelsky S., Archangelsky A. An ultrastructural study of the araucarian pollen grain Cyclusphaera radiata Archangelsky from the Albian of Patagonia // Rev. Palaeobot. Palynol. 2012. V. 173. P. 57–67.
Erdtman G. Pollen morphology and plant taxonomy. Angiosperms. N. Y.: Hafner, 1966. 553 p. (An introduction to palynology. I.)
Furness C.A. Diversification of pollen and tapetum in early angiosperms // Early Events in Monocot Evolution / Eds. Wilkin P., Mayo S. Cambridge: Univ. Press, 2013. P. 82–98 (Syst. Assoc. Spec. V. 83).
Gavrilova O., Zavialova N., Tekleva M., Karasev E. Potential of CLSM in studying some modern and fossil palynological objects // J. Microsc. 2018. V. 269. № 3. P. 291–309.
Godfrey R.K., Wooten J.W. Aquatic and Wetland Plants of Southeastern United States: Dicotyledons. Athens: Univ. Georgia Press, 2011. 944 p.
Hesse M., Halbritter H., Weber M. et al. Pollen terminology – an illustrated handbook. N. Y.: Springer, 2009. 266 p.
Kosenko V.N. Contribution to the pollen morphology and taxonomy of Liliaceae // Grana. 1999. V. 38. № 1. P. 20–30.
Krassilov V.A., Tekleva M.V., Meyer-Melikian N.R., Rasnitsyn A.P. New pollen morphotype from gut compression of a Cretaceous insect, and its bearing on palynomorphological evolution and palaeoecology // Cret. Res. 2003. V. 24. № 2. P. 149–156.
Kurmann M.H. Exine formation in Cunninghamia lanceolata (Taxodiaceae) // Rev. Palynol. Paleobot. 1990. V. 64. P. 175–179.
Kurmann M.H. Pollen ultrastructure in Elatides willamsonii (Taxodiaceae) from the Jurassic of North Yorkshire // Rev. Palynol. Paleobot. 1991. V. 69. P. 291–298.
Lammers T.G. Campanulaceae // The Families and Genera of Vascular Plants. V. 8: Flowering Plants: Eudicots – Asterales / Eds. Kadereit J.W., Jeffrey C. Heidelberg: Springer Sci. Business Media, 2007. P. 25–56.
Leffingwell H.A., Larson D.A., Valencia M.J. A study of the fossil pollen Wodehouseia spinata. I. Ultrastructure and comparisons to selected modern taxa. II. Optical microscopic recognition of foot layers in differentially stained fossil pollen and their significance // Bull. Canad. Petrol. Geol. 1970. V. 18. № 2. P. 238–262.
Ligong L., Hanxing L. Pollen morphology and its taxonomic significance of Piperaceae // Acta Bot. Yunnanica. 1998. V. 20. № 4. P. 429–433.
Manabale T.S. Spores and pollen grains of water plants and their dispersal // Rev. Palynol. Paleobot. 1968. V. 7. P. 285–296.
Markevich V.S., Bugdaeva E.V., Ashraf A.R., Sun Ge. Boundary of Cretaceous and Paleogene continental deposits in Zeya-Bureya Basin, Amur (Heilongjiang) river region // Global Geol. 2011. V. 14. № 3. P. 144–159.
PalDat – a palynological database (2000 onwards, www.paldat.org).
http://paleobotany.ru/palynodata.
Polevova S.V. Review of the sporoderm ultrastructure of members of the Asterales // Paleontol. J. 2006. V. 40. P. 656–663.
Praglowski J. The pollen morphology of the Haloragaceae with reference to taxonomy // Grana. 1970. V. 10. № 3. P. 159–239.
Romero A.T., Salinas M.J., Fernandez M.C. Pollen wall development in Hypecoum imberbe Sm. (Fumariaceae) // Grana. 2003. V. 42. P. 91–101.
Sannier J., Baker W.J., Anstett M.-C., Nadot S. A comparative analysis of pollinator type and pollen ornamentation in the Araceae and the Arecaceae, two unrelated families of the monocots // BMC Res. Notes. 2009. V. 2. P. 145.
Scheuring B.W. Mikrofloren aus den Meridekalken des Mts. San Giorgio (Kanton Tessin) // Schweiz. Paläontol. Abh. 1978. V. 88. P. 1–119.
Shu P., She M.L. Pollen photographs and flora of Umbelliferae in China. Shanghai: Shanghai Sci. Techn. Publ., 2001. 331p. (in Chinese)
Stanley E.A. A new sporomorph genus from South Dakota // Pollen et Spores. 1961. V. 3. № 1. P. 155–162.
Tanaka N., Uehara K., Murata J. Correlation between pollen morphology and pollination mechanisms in the Hydrocharitaceae // J. Plant Res. 2004. V. 117. P. 265–276.
Taylor M.L., Cooper R.L., Schneider E.L., Osborn J.M. Pollen structure and development in Nymphaeales: Insights into character evolution in an ancient angiosperm lineage // Amer. J. Bot. 2015. V. 102. № 10. P. 1685–1702.
Tebbs M.C. Piperaceae // Flowering Plants. Dicotyledons: Magnoliid, Hamamelid and Caryophyllid Families. V. 2 / Eds. Kubitzki K., Rohwer J.G., Bittrich V. Heidelberg: Springer Sci. Business Media, 1993. P. 516–521.
Tekleva M.V. Pollen morphology and ultrastructure of several Gnetum species // Plant Syst. Evol. 2016. V. 302. № 3. P. 291–303.
Tryon A.F., Lugardon B. Spores of the Pteridophyta. Surface, wall structure and evolution based on electron microscope studies. N. Y.: Springer Verlag, 1990. 648 p.
Van der Merwe J.J.M., Van Wyk A.E., Kok P.D.F. Pollen types in the Lauraceae // Grana. 1990. V. 29. P. 185–196.
Wiggins V.D. Fossil oculata pollen from Alaska // Geosci. and Man. 1976. V. 15. P. 51–76.
Zavada M.S. The wall structure of fossil discoid pollen // Bull. Torrey Bot. Club. 1992. V. 119. P. 44–49.
Zavialova N.E., Tekleva M.V., Smirnova S.B., Mroueh M. Exine ultrastructure in pollen grains of Classopollis Pflug from the Cretaceous of Lebanon // Paleontol. J. 2010. V. 44. № 10. P. 1353–1367.
Zetter R., Ferguson D.K. Trapaceae pollen in the Cenozoic // Acta Palaeobot. 2001. V. 41. P. 321–339.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал